
CRISPR/Cas9 Mediated Fluorescent Tagging of Caenorhabditis elegans SPE-38 Reveals a Complete Localization Pattern in Live Spermatozoa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Worm Strains, Brood Sizing, Statistical Analysis, Sperm Preparations, and C. elegans Culture
2.2. CRISPR/Cas9 Mediated Fluorescent Tagging of Endogenous SPE-38
2.3. Microscopic Imaging of SPE-38::wrmScarlet-I Localization in Spermatids and Spermatozoa
3. Results
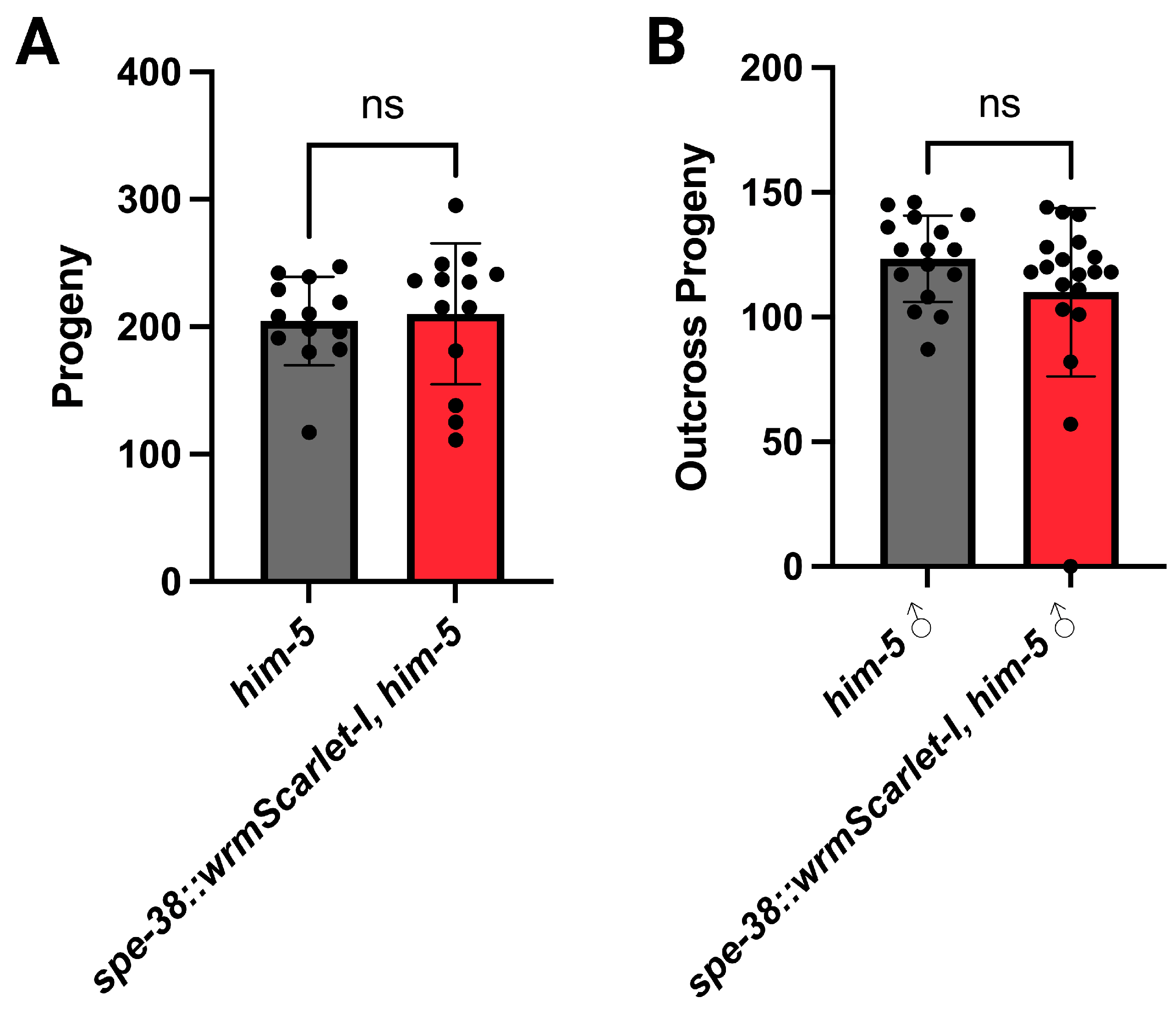
3.1. Genome Edited Animals Are Fertile
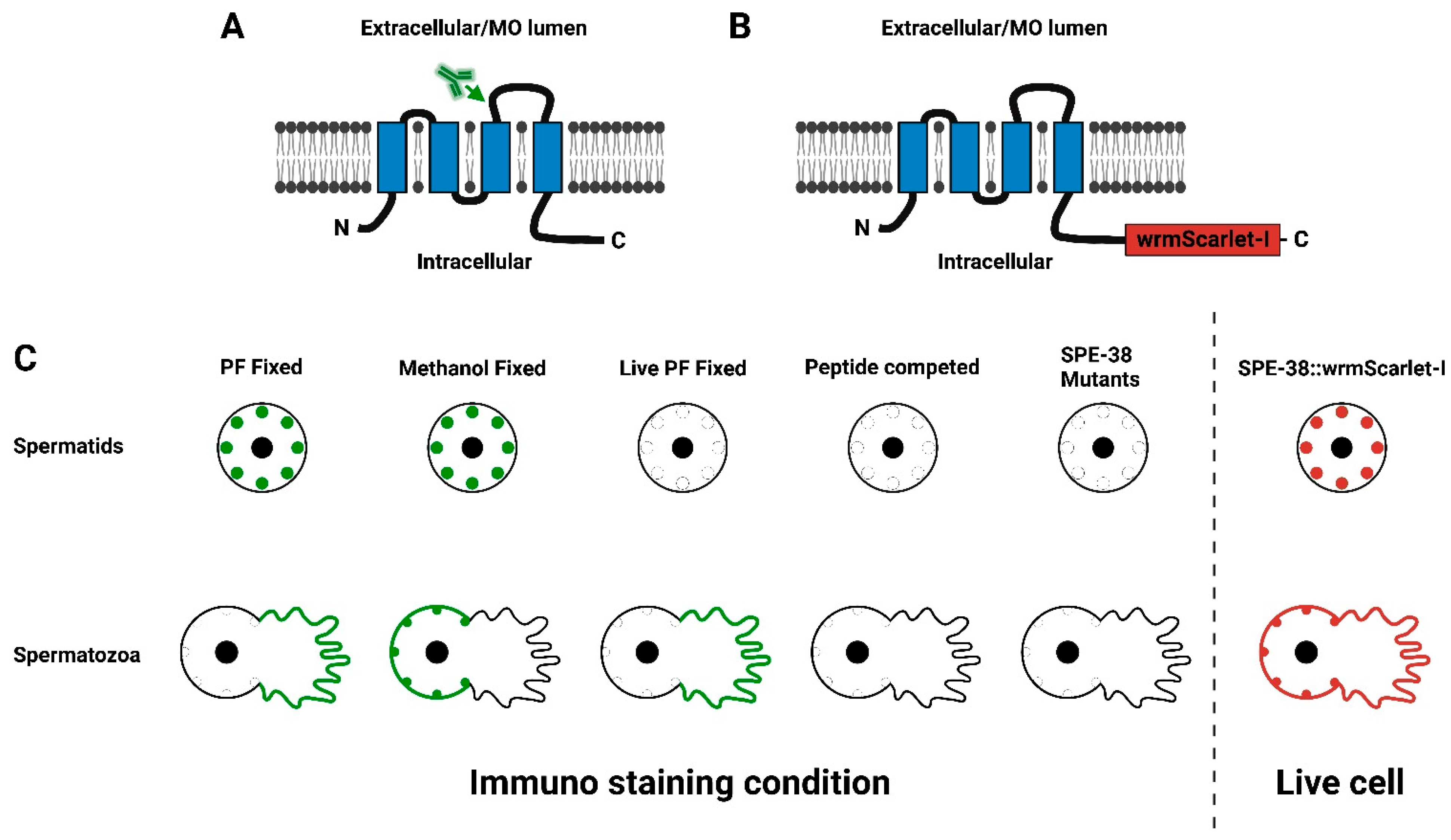
3.2. SPE-38::wrmScarlet-I Is Localized to the Membranous Organelles in Spermatids
3.3. SPE-38::wrmScarlet-I Redistributes to the Plasma Membrane of the Cell Body and the Pseudopod in Mature Sperm
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krauchunas, A.R.; Marcello, M.R.; Singson, A. The molecular complexity of fertilization: Introducing the concept of a fertilization synapse. Mol. Reprod. Dev. 2016, 83, 376–386. [Google Scholar] [CrossRef]
- Mei, X.; Singson, A.W. The molecular underpinnings of fertility: Genetic approaches in Caenorhabditis elegans. Adv. Genet. 2020, 2, e10034. [Google Scholar] [CrossRef]
- Lu, Y.; Ikawa, M. Eukaryotic fertilization and gamete fusion at a glance. J. Cell Sci. 2022, 135, 260296. [Google Scholar] [CrossRef]
- Deneke, V.E.; Pauli, A. The Fertilization Enigma: How Sperm and Egg Fuse. Annu. Rev. Cell Dev. Biol. 2021, 37, 391–414. [Google Scholar] [CrossRef]
- Chatterjee, I.; Richmond, A.; Putiri, E.; Shakes, D.C.; Singson, A. The Caenorhabditis elegans spe-38 gene encodes a novel four-pass integral membrane protein required for sperm function at fertilization. Development 2005, 132, 2795–2808. [Google Scholar] [CrossRef]
- Singson, A. Every Sperm Is Sacred: Fertilization in Caenorhabditis elegans. Dev. Biol. 2001, 230, 101–109. [Google Scholar] [CrossRef]
- Singaravelu, G.; Chatterjee, I.; Rahimi, S.; Druzhinina, M.K.; Kang, L.; Xu, X.S.; Singson, A. The sperm surface localization of the TRP-3/SPE-41 Ca2+-permeable channel depends on SPE-38 function in Caenorhabditis elegans. Dev. Biol. 2012, 365, 376–383. [Google Scholar] [CrossRef]
- Umeda, R.; Satouh, Y.; Takemoto, M.; Nakada-Nakura, Y.; Liu, K.; Yokoyama, T.; Shirouzu, M.; Iwata, S.; Nomura, N.; Sato, K.; et al. Structural insights into tetraspanin CD9 function. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Marcello, M.R.; Druzhinina, M.; Singson, A. Caenorhabditis elegans sperm membrane protein interactome. Biol. Reprod. 2018, 98, 776–783. [Google Scholar] [CrossRef]
- Samuel, A.D.; Murthy, V.N.; Hengartner, M.O. Calcium dynamics during fertilization in C. elegans. BMC Dev. Biol. 2001, 1, 8. [Google Scholar] [CrossRef]
- Xu, X.-Z.; Sternberg, P.A. C. elegans Sperm TRP Protein Required for Sperm-Egg Interactions during Fertilization. Cell 2003, 114, 285–297. [Google Scholar] [CrossRef]
- Takayama, J.; Onami, S. The Sperm TRP-3 Channel Mediates the Onset of a Ca2+ Wave in the Fertilized C. elegans Oocyte. Cell Rep. 2016, 15, 625–637. [Google Scholar] [CrossRef]
- Nelson, G.A.; Ward, S. Vesicle fusion, pseudopod extension and amoeboid motility are induced in nematode spermatids by the ionophore monensin. Cell 1980, 19, 457–464. [Google Scholar] [CrossRef]
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis. Curr. Biol. 2017, 27, R988–R994. [Google Scholar] [CrossRef]
- Mello, C.; Kramer, J.; Stinchcomb, D.; Ambros, V. Efficient gene transfer in C.elegans: Extrachromosomal maintenance and integration of transforming sequences. EMBO J. 1991, 10, 3959–3970. [Google Scholar] [CrossRef]
- Kadandale, P.; Chatterjee, I.; Singson, A. Germline Transformation of Caenorhabditis elegans by Injection. Methods Mol. Biol. 2009, 518, 123–133. [Google Scholar] [CrossRef]
- Praitis, V.; Casey, E.; Collar, D.; Austin, J. Creation of Low-Copy Integrated Transgenic Lines in Caenorhabditis elegans. Genetics 2001, 157, 1217–1226. [Google Scholar] [CrossRef]
- Praitis, V.; Maduro, M.F. Transgenesis in C. elegans. Methods Cell Biol. 2011, 106, 159–185. [Google Scholar] [CrossRef]
- Geldziler, B.D.; Marcello, M.R.; Shakes, D.C.; Singson, A. The Genetics and Cell Biology of Fertilization. Methods Cell Biol. 2011, 106, 343–375. [Google Scholar] [CrossRef]
- Krauchunas, A.R.; Werner, M.; Britt, N.; Chen, D.S.; Maddox, A.S.; Singson, A. C. elegans CYLC-2 localizes to sperm. Micropubl. Biol. 2020, 2020, 314. [Google Scholar]
- Kelly, W.G.; Xu, S.; Montgomery, M.K.; Fire, A. Distinct requirements for somatic and germline expression of a generally expressed Caenorhabditis elegans gene. Genetics 1997, 146, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Frøkjær-Jensen, C.; Davis, M.; Ailion, M.; Jorgensen, E.M. Improved Mos1-mediated transgenesis in C. elegans. Nat. Methods 2012, 9, 117–118. [Google Scholar] [CrossRef] [PubMed]
- Nance, J.; Frøkjær-Jensen, C. The Caenorhabditis elegans Transgenic Toolbox. Genetics 2019, 212, 959–990. [Google Scholar] [CrossRef]
- Dickinson, D.J.; Goldstein, B. CRISPR-Based Methods for Caenorhabditis elegans Genome Engineering. Genetics 2016, 202, 885–901. [Google Scholar] [CrossRef]
- Heppert, J.K.; Dickinson, D.J.; Pani, A.M.; Higgins, C.D.; Steward, A.; Ahringer, J.; Kuhn, J.R.; Goldstein, B. Comparative assessment of fluorescent proteins for in vivo imaging in an animal model system. Mol. Biol. Cell 2016, 27, 3385–3394. [Google Scholar] [CrossRef]
- Hutter, H. Fluorescent Protein Methods: Strategies and Applications. Methods Cell Biol. 2012, 107, 67–92. [Google Scholar] [CrossRef]
- Bindels, D.S.; Haarbosch, L.; van Weeren, L.; Postma, M.; Wiese, K.E.; Mastop, M.; Aumonier, S.; Gotthard, G.; Royant, A.; Hink, M.A.; et al. mScarlet: A bright monomeric red fluorescent protein for cellular imaging. Nat. Methods 2017, 14, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Maddox, A.S.; Maddox, P.S. High-Resolution Imaging of Cellular Processes in Caenorhabditis elegans. Methods Cell Biol. 2012, 107, 1–34. [Google Scholar] [CrossRef]
- Goudeau, J.; Sharp, C.S.; Paw, J.; Savy, L.; Leonetti, M.D.; York, A.G.; Updike, D.L.; Kenyon, C.; Ingaramo, M. Split-wrmScarlet and split-sfGFP: Tools for faster, easier fluorescent labeling of endogenous proteins in Caenorhabditis elegans. Genetics 2021, 217, 14. [Google Scholar] [CrossRef]
- El Mouridi, S.; Lecroisey, C.; Tardy, P.; Mercier, M.; Leclercq-Blondel, A.; Zariohi, N.; Boulin, T. Reliable CRISPR/Cas9 Genome Engineering in Caenorhabditis elegans Using a Single Efficient sgRNA and an Easily Recognizable Phenotype. G3 Genes Genomes Genet. 2017, 7, 1429–1437. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Shakes, D.C.; Ward, S. Initiation of spermiogenesis in C. elegans: A pharmacological and genetic analysis. Dev. Biol. 1989, 134, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Singaravelu, G.; Chatterjee, I.; Marcello, M.R.; Singson, A. Isolation and in vitro activation of Caenorhabditis elegans sperm. J. Vis. Exp. JoVE 2011, 47, e2336. [Google Scholar] [CrossRef]
- Mei, X.; Druzhinina, M.; Dharia, S.; Krauchunas, A.R.; Ni, J.; Singaravelu, G.; Gu, S.G.; Shakes, D.; Grant, B.D.; Singson, A.W. SPE-51, a sperm secreted protein with an immunoglobulin-like domain, is required for sperm-egg fusion in C. elegans. bioRxiv 2021. [Google Scholar] [CrossRef]
- Miné-Hattab, J. When fixation creates fiction. Elife 2023, 12, 85671. [Google Scholar] [CrossRef] [PubMed]
- Krauchunas, A.R.; Marcello, M.R.; Looper, A.M.; Mei, X.; Putiri, E.; Singaravelu, G.; Ahmed, I.I.; Singson, A. The EGF-motif containing protein SPE-36 is a secreted protein required for sperm function at fertilization in C. elegans. bioRxiv 2021. [Google Scholar] [CrossRef]
- Geldziler, B.; Chatterjee, I.; Kadandale, P.; Putiri, E.; Patel, R.; Singson, A. A comparative study of sperm morphology, cytology and activation in Caenorhabditis elegans, Caenorhabditis remanei and Caenorhabditis briggsae. Dev. Genes Evol. 2006, 216, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Shakes, D.C.; Miller, D.M., 3rd; Nonet, M.L. Immunofluorescence Microscopy. Methods Cell Biol. 2012, 107, 35–66. [Google Scholar] [CrossRef]
- Lee, C.W.; Vitriol, E.A.; Shim, S.; Wise, A.L.; Velayutham, R.P.; Zheng, J.Q. Dynamic Localization of G-Actin during Membrane Protrusion in Neuronal Motility. Curr. Biol. 2013, 23, 1046–1056. [Google Scholar] [CrossRef]
- Caroti, F.; Thiels, W.; Vanslambrouck, M.; Jelier, R. Wnt Signaling Induces Asymmetric Dynamics in the Actomyosin Cortex of the C. elegans Endomesodermal Precursor Cell. Front. Cell Dev. Biol. 2021, 9, 702741. [Google Scholar] [CrossRef]
- Parry, J.M.; Velarde, N.V.; Lefkovith, A.J.; Zegarek, M.H.; Hang, J.S.; Ohm, J.; Klancer, R.; Maruyama, R.; Druzhinina, M.K.; Grant, B.D.; et al. EGG-4 and EGG-5 Link Events of the Oocyte-to-Embryo Transition with Meiotic Progression in C. elegans. Curr. Biol. 2009, 19, 1752–1757. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.A.; Roberts, T.M.; Ward, S. Caenorhabditis elegans spermatozoan locomotion: Amoeboid movement with almost no actin. J. Cell Biol. 1982, 92, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Foor, W.E. Zygote Formation in Ascaris Lumbricoides (Nematoda). J. Cell Biol. 1968, 39, 119–134. [Google Scholar] [CrossRef]
- Yanagimachi, R. Mammalian fertilization. In The Physiology of Reproduction; Knobil, E., Neill, J.D., Eds.; Raven Press: New York, NY, USA, 1994; pp. 189–317. [Google Scholar]
- Gleason, E.J.; Lindsey, W.C.; Kroft, T.L.; Singson, A.W.; L’Hernault, S.W. spe-10 Encodes a DHHC–CRD Zinc-Finger Membrane Protein Required for Endoplasmic Reticulum/Golgi Membrane Morphogenesis during Caenorhabditis elegans Spermatogenesis. Genetics 2006, 172, 145–158. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, Y.; Mei, X.; Singson, A. CRISPR/Cas9 Mediated Fluorescent Tagging of Caenorhabditis elegans SPE-38 Reveals a Complete Localization Pattern in Live Spermatozoa. Biomolecules 2023, 13, 623. https://doi.org/10.3390/biom13040623
Zuo Y, Mei X, Singson A. CRISPR/Cas9 Mediated Fluorescent Tagging of Caenorhabditis elegans SPE-38 Reveals a Complete Localization Pattern in Live Spermatozoa. Biomolecules. 2023; 13(4):623. https://doi.org/10.3390/biom13040623
Chicago/Turabian StyleZuo, Yamei, Xue Mei, and Andrew Singson. 2023. "CRISPR/Cas9 Mediated Fluorescent Tagging of Caenorhabditis elegans SPE-38 Reveals a Complete Localization Pattern in Live Spermatozoa" Biomolecules 13, no. 4: 623. https://doi.org/10.3390/biom13040623
APA StyleZuo, Y., Mei, X., & Singson, A. (2023). CRISPR/Cas9 Mediated Fluorescent Tagging of Caenorhabditis elegans SPE-38 Reveals a Complete Localization Pattern in Live Spermatozoa. Biomolecules, 13(4), 623. https://doi.org/10.3390/biom13040623