
MiRNA and Exosomal miRNA as New Biomarkers Useful to Phenotyping Severe Asthma
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
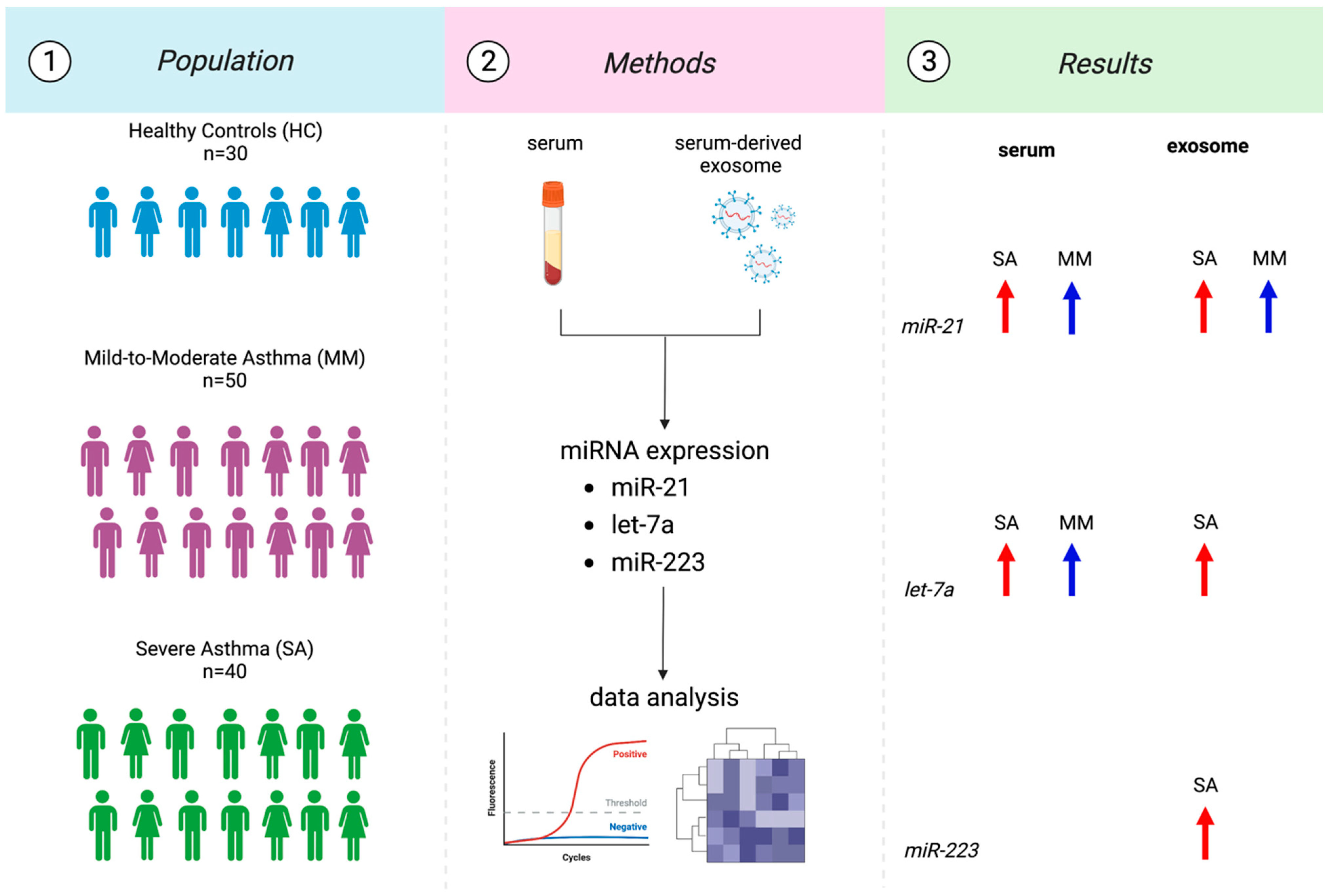
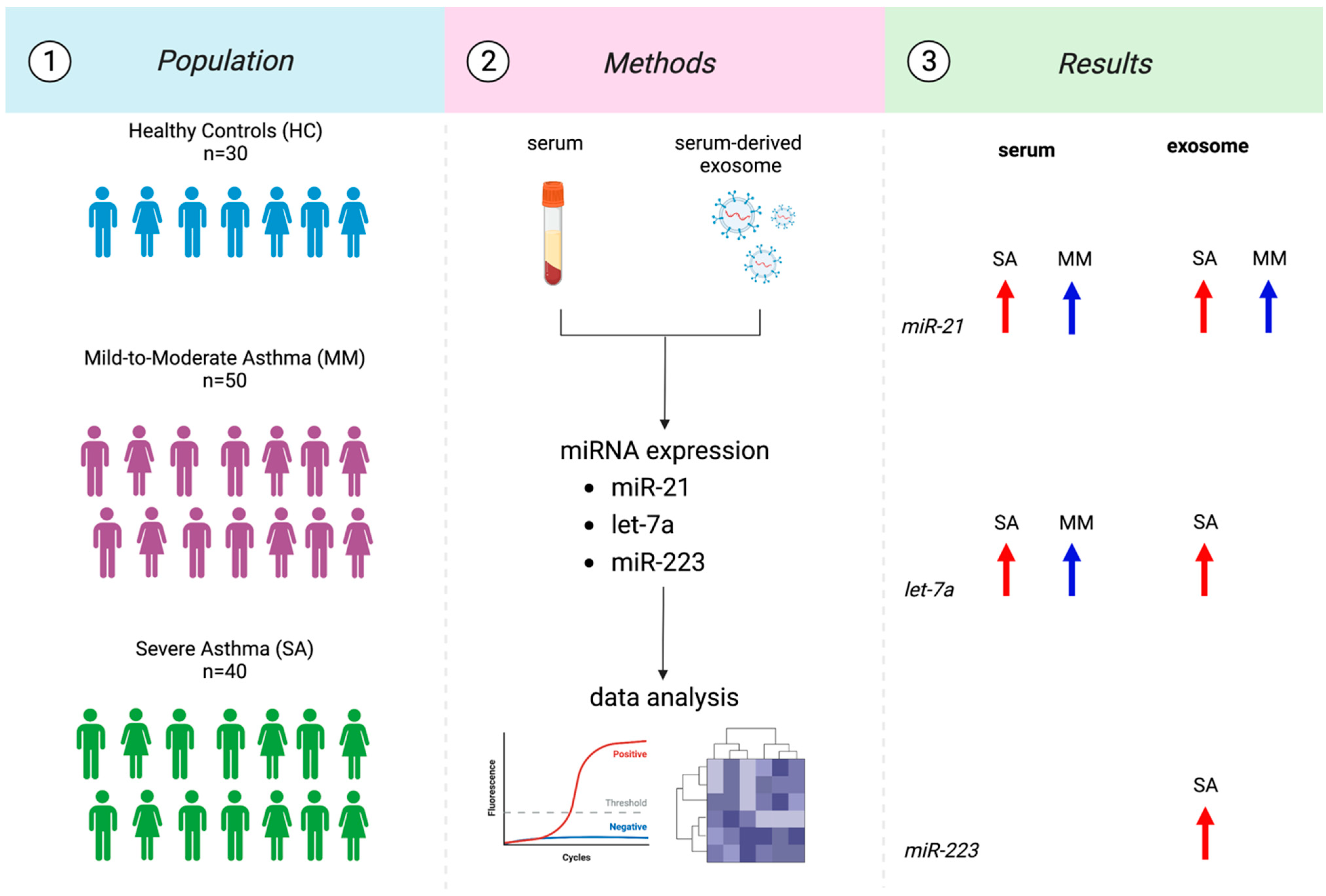
2.1. Study Population and Sampling
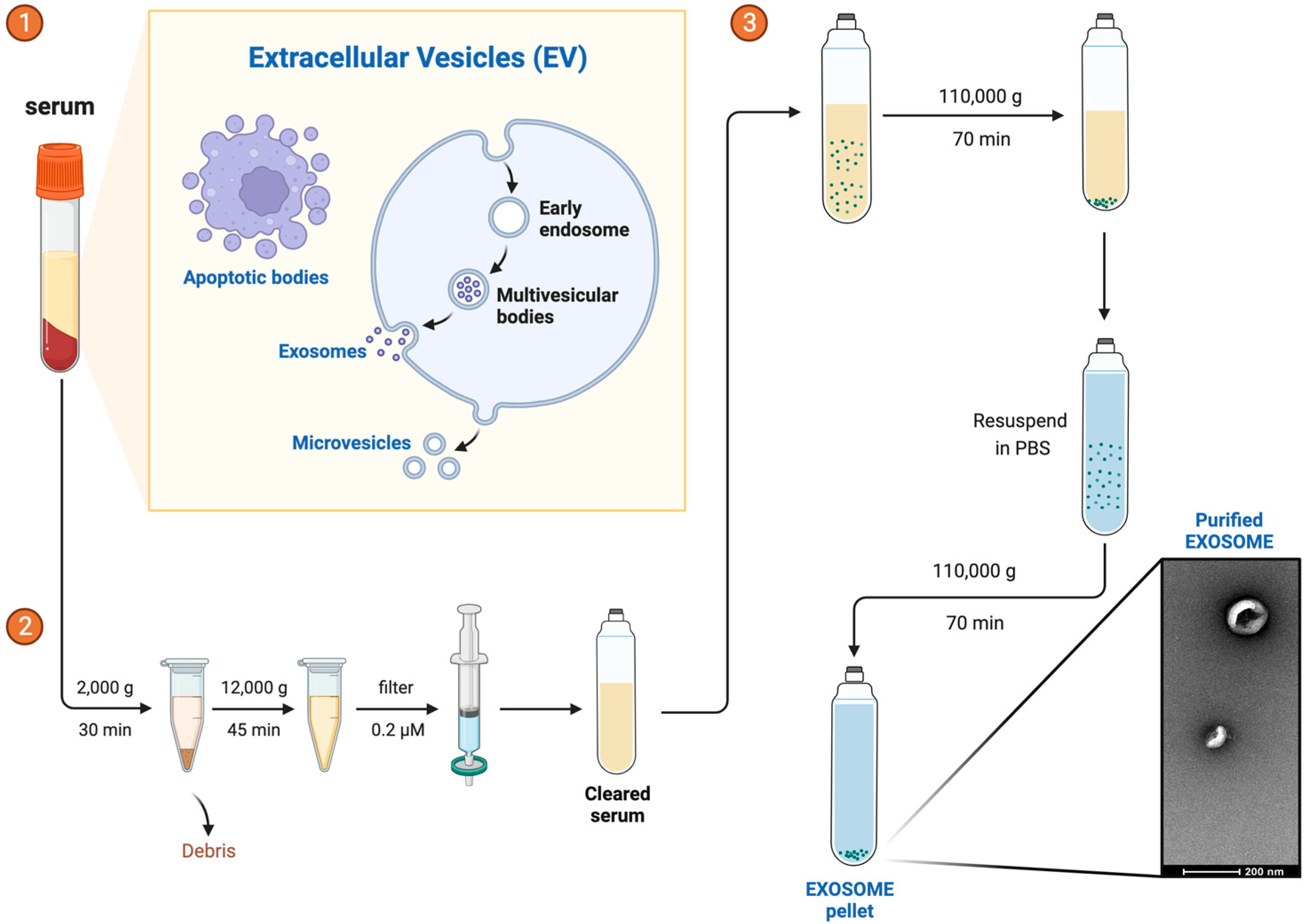
2.2. Exosome Isolation
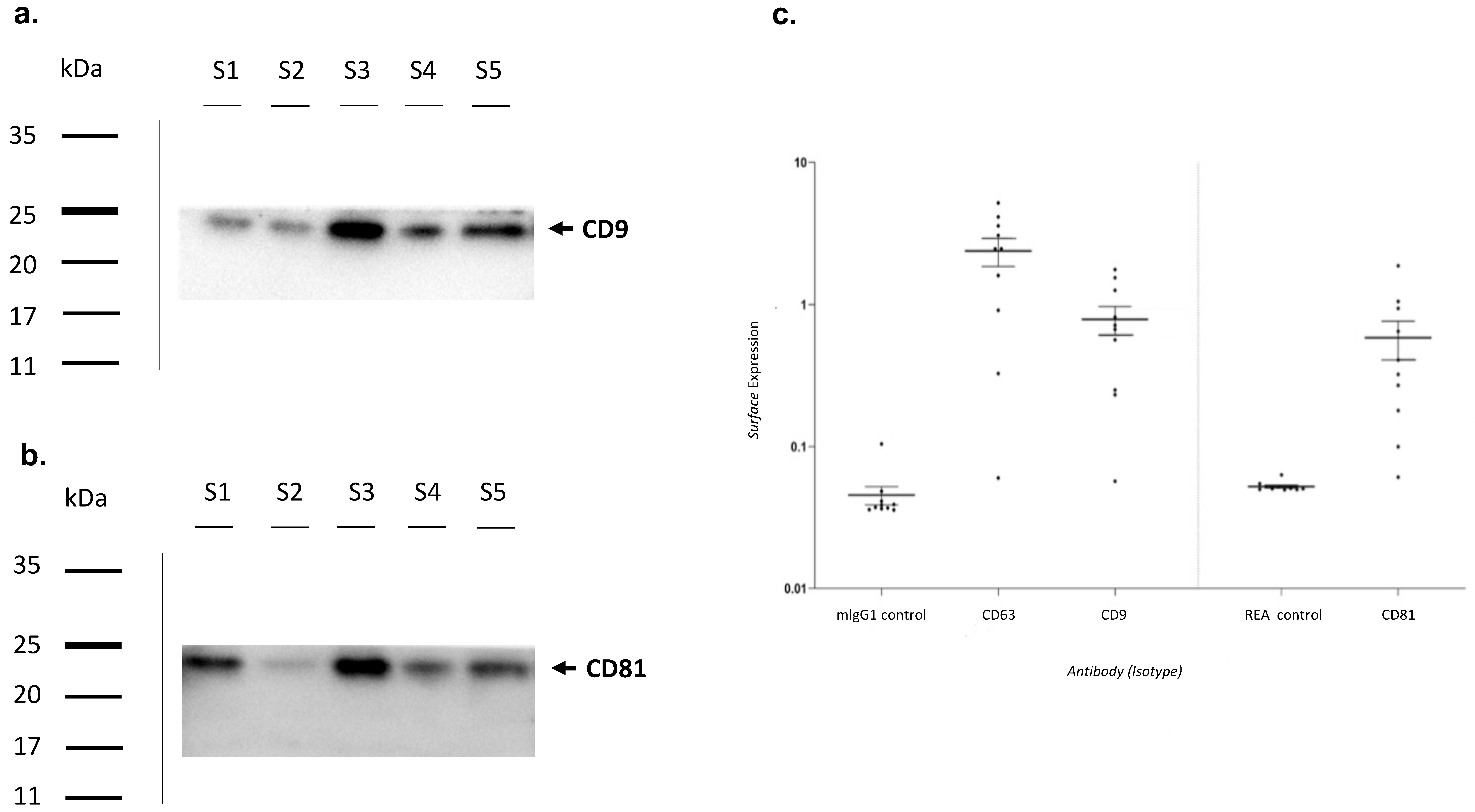
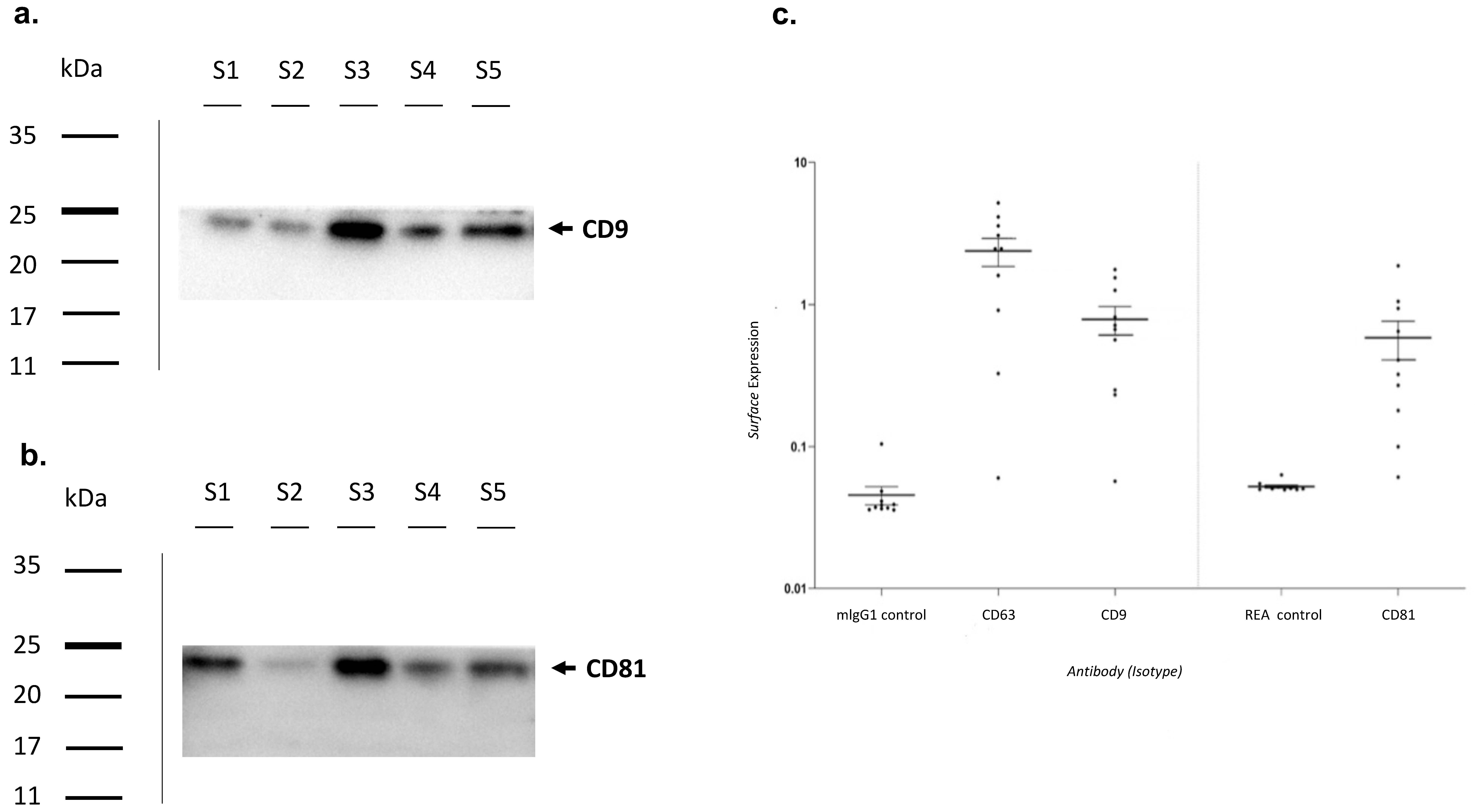
2.3. Exosome Characterization by Western Blot
2.4. Exosome Characterization by Multiplex Surface Marker Analysis
2.5. Total RNA Extraction
2.6. RNA Reverse Transcription and miRNA’s Expression through qRT-PCR
2.7. Statistical Analysis
3. Results
3.1. Demographic and Clinical Characteristics of the Study Population
3.2. Exosomes Characterization
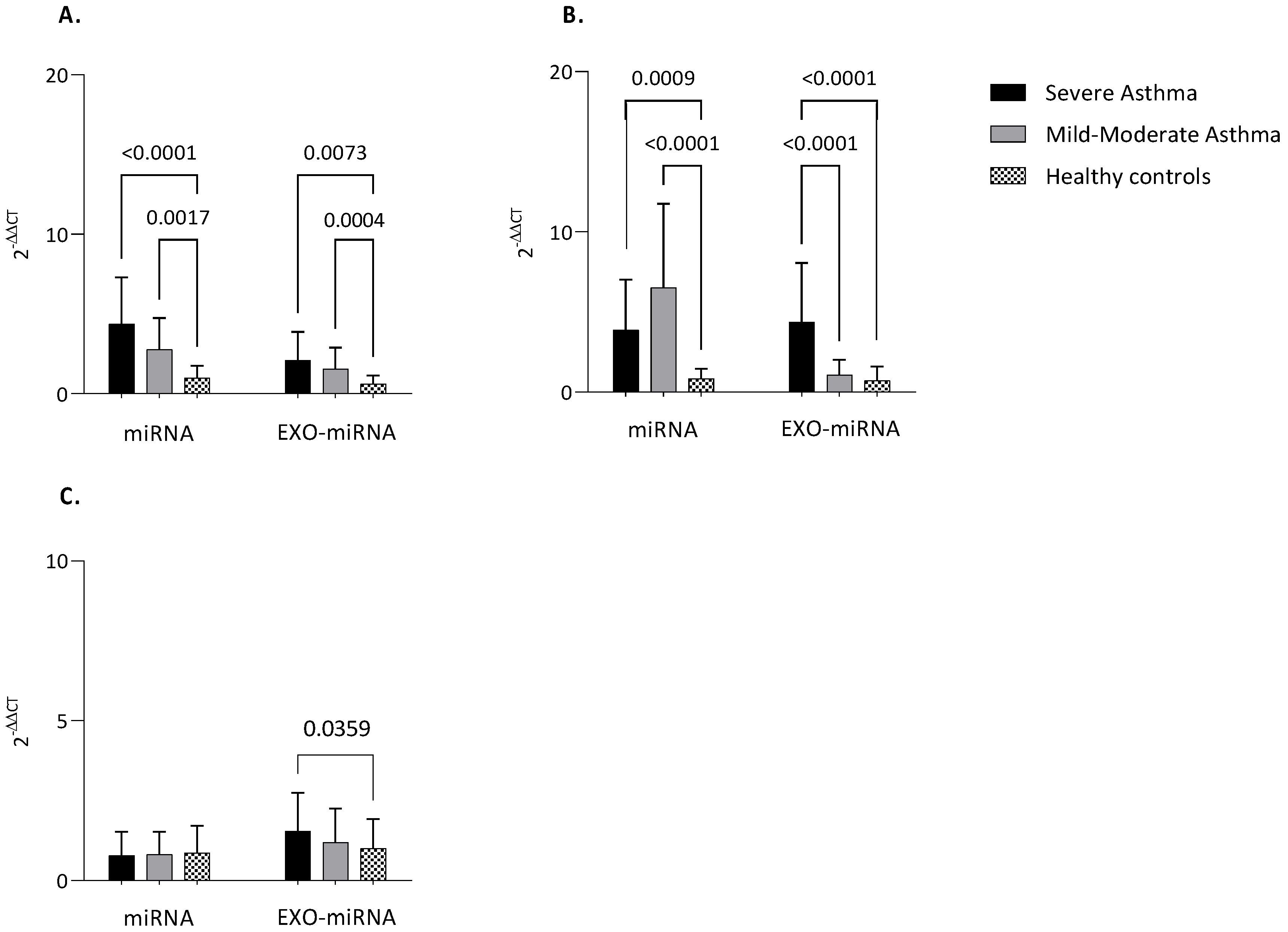
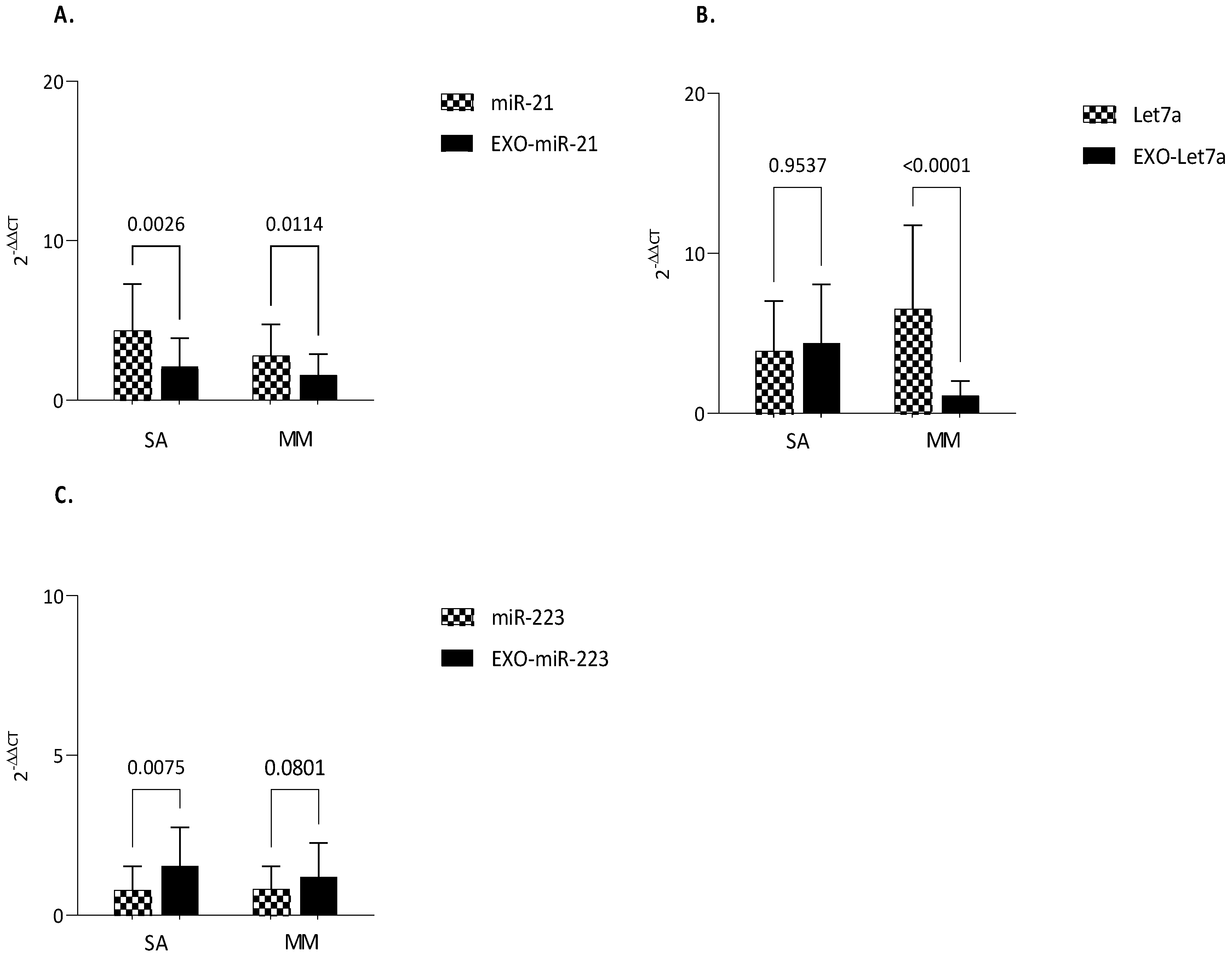
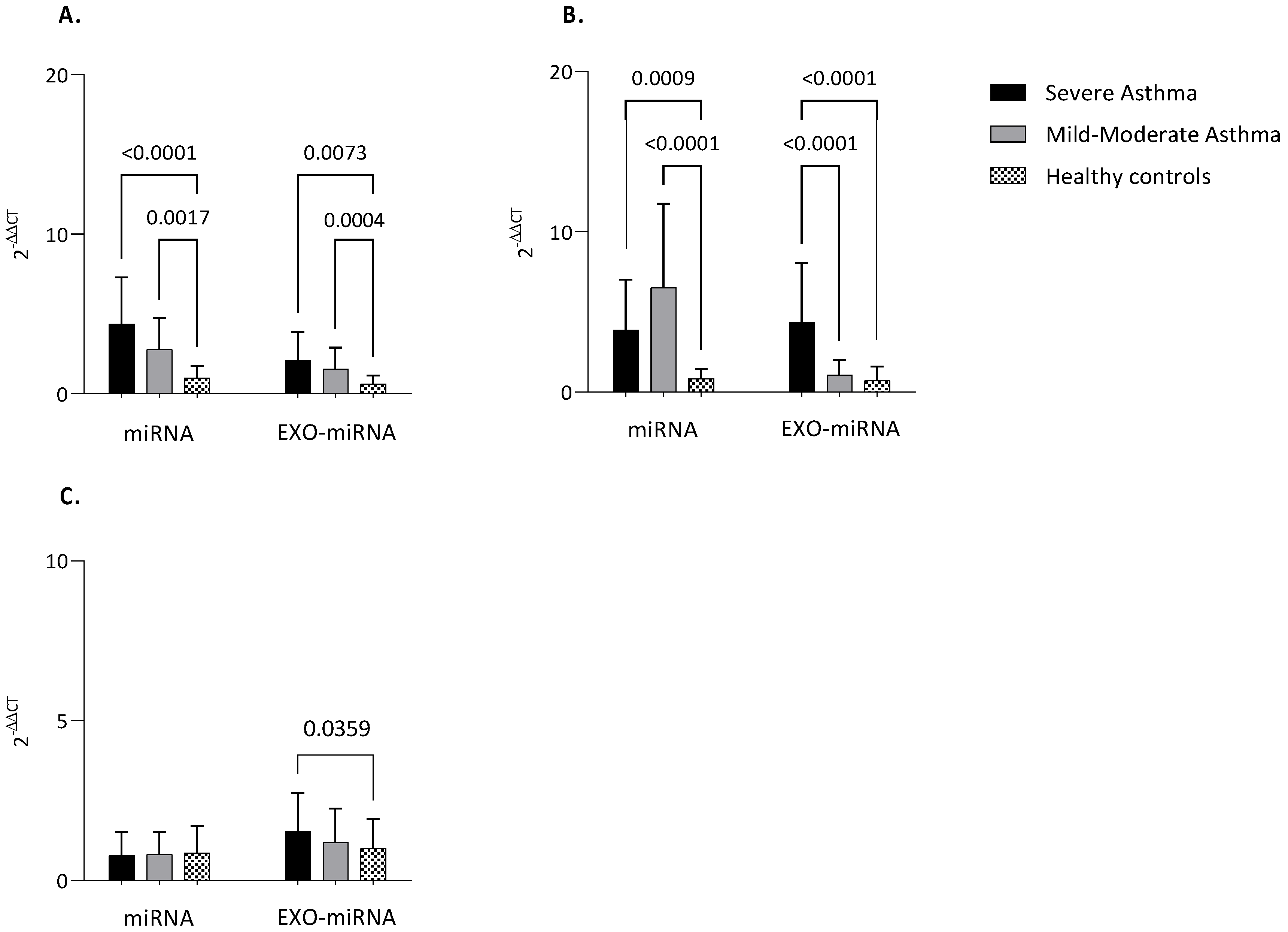
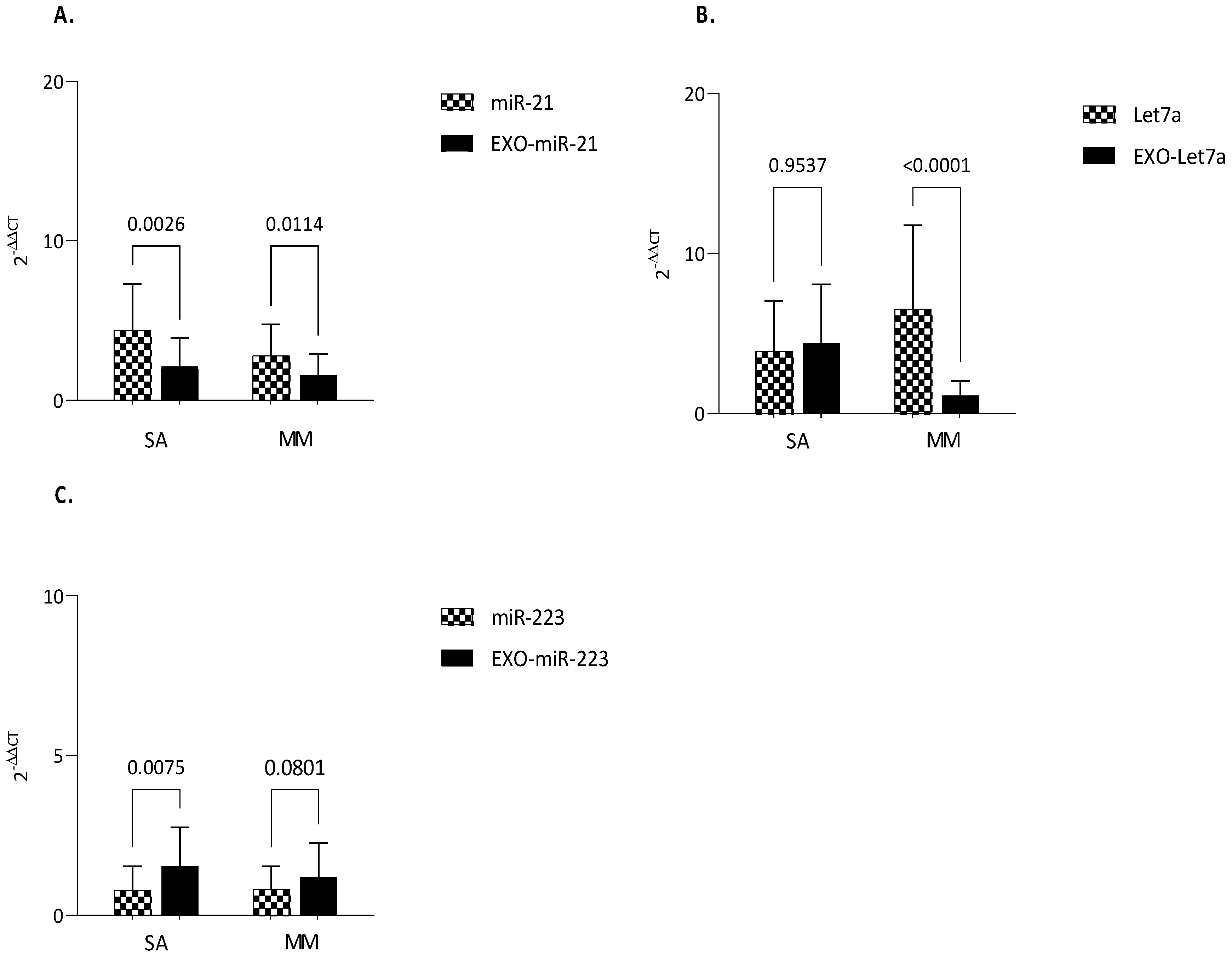
3.3. MiRNAs Expression
3.4. Clinical Correlations
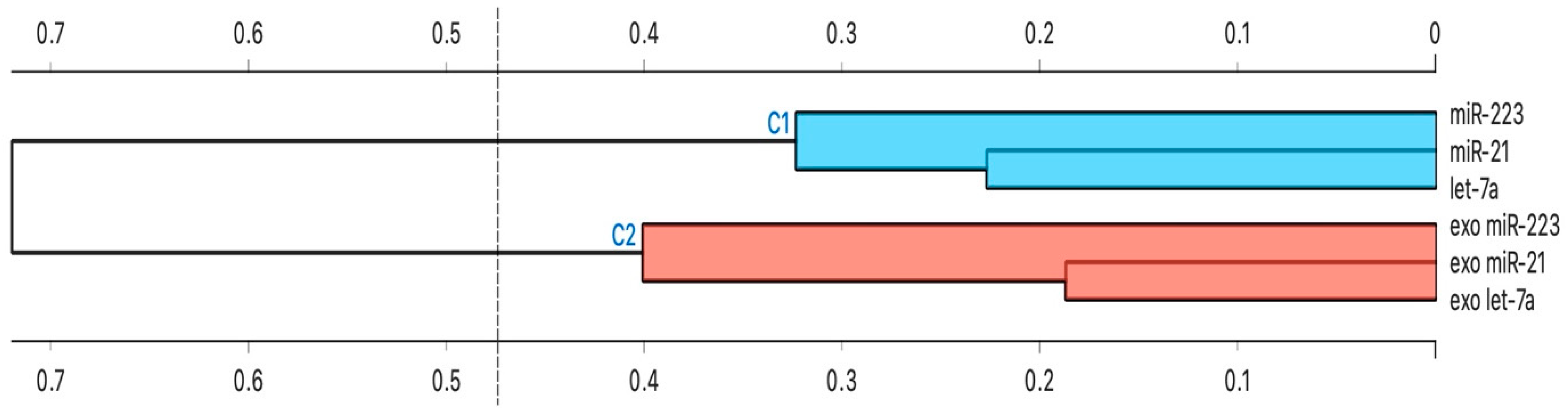
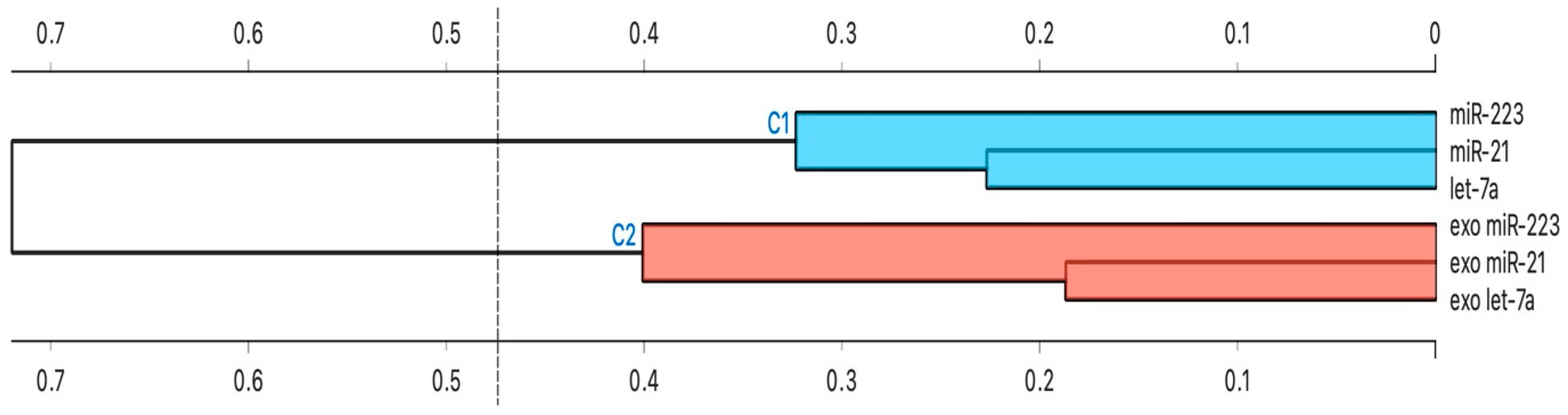
3.5. Cluster Analysis
4. Discussion
Limitations of the Study
5. Conclusions
Take Home Message
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Global Initiative for Asthma. Global Strategy for Asthma Management and Prevention. 2022. Available online: www.ginasthma.org (accessed on 2 September 2022).
- Bagnasco, D.; Paggiaro, P.; Latorre, M.; Folli, C.; Testino, E.; Bassi, A.; Milanese, M.; Heffler, E.; Manfredi, A.; Riccio, A.M.; et al. Severe asthma: One disease and multiple definitions. World Allergy Organ. J. 2021, 14, 100606. [Google Scholar] [CrossRef] [PubMed]
- Gans, M.D.; Gavrilova, T. Understanding the immunology of asthma: Pathophysiology, biomarkers, and treatments for asthma endotypes. Paediatr. Respir. Rev. 2020, 36, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef] [PubMed]
- Tsitsiou, E.; Williams, A.E.; Moschos, S.A.; Patel, K.; Rossios, C.; Jiang, X.; Adams, O.-D.; Macedo, P.; Booton, R.; Gibeon, D.; et al. Transcriptome analysis shows activation of circulating CD8+ T cells in patients with severe asthma. J. Allergy Clin. Immunol. 2012, 129, 95–103. [Google Scholar] [CrossRef] [PubMed]
- van der Ploeg, E.K.; Krabbendam, L.; Vroman, H.; van Nimwegen, M.; de Bruijn, M.J.W.; de Boer, G.M.; Bergen, I.M.; Kool, M.; Tramper-Standers, G.A.; Braunstahl, G.J.; et al. Type-2 CD8+ T-cell formation relies on interleukin-33 and is linked to asthma exacerbations. Nat. Commun. 2023, 14, 5137. [Google Scholar] [CrossRef]
- Levy, M.L.; Bacharier, L.B.; Bateman, E.; Boulet, L.-P.; Brightling, C.; Buhl, R.; Brusselle, G.; Cruz, A.A.; Drazen, J.M.; Duijts, L.; et al. Key recommendations for primary care from the 2022 Global Initiative for Asthma (GINA) update. NPJ Prim. Care Respir. Med. 2023, 33, 7. [Google Scholar] [CrossRef]
- van den Berge, M.; Tasena, H. Role of microRNAs and exosomes in asthma. Curr. Opin. Pulm. Med. 2019, 25, 87–93. [Google Scholar] [CrossRef]
- Mohr, A.M.; Mott, J.L. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar] [CrossRef]
- Weidner, J.; Bartel, S.; Kılıç, A.; Zissler, U.M.; Renz, H.; Schwarze, J.; Schmidt-Weber, C.B.; Maes, T.; Rebane, A.; Krauss-Etschmann, S.; et al. Spotlight on microRNAs in allergy and asthma. Allergy 2021, 76, 1661–1678. [Google Scholar] [CrossRef]
- Habib, N.; Pasha, M.A.; Tang, D.D. Current Understanding of Asthma Pathogenesis and Biomarkers. Cells 2022, 11, 2764. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Fabbiano, F.; Corsi, J.; Gurrieri, E.; Trevisan, C.; Notarangelo, M.; D’Agostino, V.G. RNA packaging into extracellular vesicles: An orchestra of RNA-binding proteins? J. Extracell. Vesicles 2020, 10, e12043. [Google Scholar] [CrossRef] [PubMed]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Purghè, B.; Manfredi, M.; Ragnoli, B.; Baldanzi, G.; Malerba, M. Exosomes in chronic respiratory diseases. Biomed. Pharmacother. 2021, 144, 112270. [Google Scholar] [CrossRef]
- Grueso-Navarro, E.; Navarro, P.; Laserna-Mendieta, E.J.; Lucendo, A.J.; Arias-González, L. Blood-Based Biomarkers for Eosinophilic Esophagitis and Concomitant Atopic Diseases: A Look into the Potential of Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 3669. [Google Scholar] [CrossRef]
- Li, Y.; Yin, Z.; Fan, J.; Zhang, S.; Yang, W. The roles of exosomal miRNAs and lncRNAs in lung diseases. Signal. Transduct. Target. Ther. 2019, 4, 47. [Google Scholar] [CrossRef]
- Alhamwe, B.A.; Miethe, S.; Von Strandmann, E.P.; Potaczek, D.P.; Garn, H. Epigenetic Regulation of Airway Epithelium Immune Functions in Asthma. Front. Immunol. 2020, 11, 1747. [Google Scholar] [CrossRef]
- Boateng, E.; Kovacevic, D.; Oldenburg, V.; Rådinger, M.; Krauss-Etschmann, S. Role of airway epithelial cell miRNAs in asthma. Front. Allergy 2022, 3, 962693. [Google Scholar] [CrossRef]
- Gil-Martínez, M.; Lorente-Sorolla, C.; Naharro, S.; Rodrigo-Muñoz, J.M.; Del Pozo, V. Advances and Highlights of miRNAs in Asthma: Biomarkers for Diagnosis and Treatment. Int. J. Mol. Sci. 2023, 24, 1628. [Google Scholar] [CrossRef] [PubMed]
- Feketea, G.; Bocsan, C.I.; Popescu, C.; Gaman, M.; Stanciu, L.A.; Zdrenghea, M.T. A Review of Macrophage MicroRNAs’ Role in Human Asthma. Cells 2019, 8, 420. [Google Scholar] [CrossRef]
- Alhamwe, B.A.; Potaczek, D.P.; Miethe, S.; Alhamdan, F.; Hintz, L.; Magomedov, A.; Garn, H. Extracellular Vesicles and Asthma-More Than Just a Co-Existence. Int. J. Mol. Sci. 2021, 22, 4984. [Google Scholar] [CrossRef] [PubMed]
- Alhamdan, F.; Greulich, T.; Daviaud, C.; Marsh, L.M.; Pedersen, F.; Thölken, C.; Pfefferle, P.I.; Bahmer, T.; Potaczek, D.P.; Tost, J.; et al. Identification of extracellular vesicle microRNA signatures specifically linked to inflammatory and metabolic mechanisms in obesity-associated low type-2 asthma. Allergy 2023. Early View. [Google Scholar] [CrossRef]
- Vázquez-Mera, S.; Martelo-Vidal, L.; Miguéns-Suárez, P.; Saavedra-Nieves, P.; Arias, P.; González-Fernández, C.; Mosteiro-Añón, M.; Corbacho-Abelaira, M.D.; Blanco-Aparicio, M.; Méndez-Brea, P.; et al. Serum exosome inflamma-miRs are surrogate biomarkers for asthma phenotype and severity. Allergy 2023, 78, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Schindler, V.E.M.; Alhamdan, F.; Preußer, C.; Hintz, L.; Alhamwe, B.A.; Nist, A.; Stiewe, T.; von Strandmann, E.P.; Potaczek, D.P.; Thölken, C.; et al. Side-Directed Release of Differential Extracellular Vesicle-associated microRNA Profiles from Bronchial Epithelial Cells of Healthy and Asthmatic Subjects. Biomedicines 2022, 10, 622. [Google Scholar] [CrossRef]
- Causin, R.L.; Souza, K.C.B.; Evangelista, A.F.; Reis, R.M.V. Identification and performance evaluation of housekeeping genes for microRNA expression normalization by reverse transcription-quantitative PCR using liquid-based cervical cytology samples. Oncol. Lett. 2019, 18, 4753–4761. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Essandoh, K.; Li, Y.; Huo, J.; Fan, G.C. MiRNA-Mediated Macrophage Polarization and its Potential Role in the Regulation of Inflammatory Response. Shock 2016, 46, 122–131. [Google Scholar] [CrossRef]
- Zhou, X.; Fragala, M.S.; McElhaney, J.E.; Kuchel, G.A. Conceptual and methodological issues relevant to cytokine and inflammatory marker measurements in clinical research. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 541–547. [Google Scholar] [CrossRef]
- Fatemi, F.; Sadroddiny, E.; Gheibi, A.; Farsani, T.M.; Kardar, G.A. Biomolecular markers in assessment and treatment of asthma. Respirology 2014, 19, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Wang, X.-P.; Luoreng, Z.-M.; Yang, J.; Jia, L.; Ma, Y.; Wei, D.-W. miR-223: An Effective Regulator of Immune Cell Differentiation and Inflammation. Int. J. Biol. Sci. 2021, 17, 2308–2322. [Google Scholar] [CrossRef]
- Roffel, M.P.; Bracke, K.R.; Heijink, I.H.; Maes, T. miR-223: A Key Regulator in the Innate Immune Response in Asthma and COPD. Front. Med. 2020, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Roffel, M.P.; Boudewijn, I.M.; van Nijnatten, J.L.; Faiz, A.; Vermeulen, C.J.; van Oosterhout, A.J.; Affleck, K.; Timens, W.; Bracke, K.R.; Maes, T.; et al. Identification of asthma-associated microRNAs in bronchial biopsies. Eur. Respir. J. 2022, 59, 2101294. [Google Scholar] [CrossRef]
- Williams, A.E.; Larner-Svensson, H.; Perry, M.M.; Campbell, G.A.; Herrick, S.E.; Adcock, I.M.; Erjefalt, J.S.; Chung, K.F.; Lindsay, M.A. MicroRNA expression profiling in mild asthmatic human airways and effect of corticosteroid therapy. PLoS ONE 2009, 4, e5889. [Google Scholar] [CrossRef]
- Rijavec, M.; Korošec, P.; Žavbi, M.; Kern, I.; Malovrh, M.M. Let-7a is differentially expressed in bronchial biopsies of patients with severe asthma. Sci. Rep. 2014, 4, 6103. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wang, Y.; Ma, Y.; Yang, J. MiR-223 plays a protecting role in neutrophilic asthmatic mice through the inhibition of NLRP3 inflammasome. Respir. Res. 2020, 21, 116. [Google Scholar] [CrossRef]
- Specjalski, K.; Niedoszytko, M. MicroRNAs: Future biomarkers and targets of therapy in asthma? Curr. Opin. Pulm. Med. 2020, 26, 285–292. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. Diagnostic, functional, and therapeutic roles of microRNA in allergic diseases. J. Allergy Clin. Immunol. 2013, 132, 3–13, quiz 14. [Google Scholar] [CrossRef]
- Lu, T.X.; Munitz, A.; Rothenberg, M.E. MicroRNA-21 is up-regulated in allergic airway inflammation and regulates IL-12p35 expression. J. Immunol. 2009, 182, 4994–5002. [Google Scholar] [CrossRef]
- Wu, S.Q.; Wang, G.L.; Li, L.Y.; Ji, J. Effects of microRNA-21 on the interleukin 12/signal transducer and activator of transcription 4 signaling pathway in asthmatic mice. Cent. Eur. J. Immunol. 2014, 39, 40–45. [Google Scholar] [CrossRef]
- Wu, X.-B.; Wang, M.-Y.; Zhu, H.-Y.; Tang, S.-Q.; You, Y.-D.; Xie, Y.-Q. Overexpression of microRNA-21 and microRNA-126 in the patients of bronchial asthma. Int. J. Clin. Exp. Med. 2014, 7, 1307–1312. [Google Scholar] [PubMed]
- Zou, Y.; Zhou, Q.; Zhang, Y. MicroRNA-21 released from mast cells-derived extracellular vesicles drives asthma in mice by potentiating airway inflammation and oxidative stress. Am. J. Transl. Res. 2021, 13, 7475–7491. [Google Scholar] [PubMed]
- Russo, F.; Di Bella, S.; Nigita, G.; Macca, V.; Laganà, A.; Giugno, R.; Pulvirenti, A.; Ferro, A. miRandola: Extracellular circulating microRNAs database. PLoS ONE 2012, 7, e47786. [Google Scholar] [CrossRef]
- Ghosh, B. Involvement of microRNA in Asthma: New perspective in respiratory biology. Indian J. Allergy Asthma Immunol. 2013, 27, 3–8. [Google Scholar] [CrossRef]
- Zhang, H.H.; Li, C.X.; Tang, L.F. The Differential Expression Profiles of miRNA-let 7a, 7b, and 7c in Bronchoalveolar Lavage Fluid from Infants with Asthma and Airway Foreign Bodies. J. Evid. Based Integr. Med. 2019, 24, 2515690X18821906. [Google Scholar] [CrossRef] [PubMed]
- Polikepahad, S.; Knight, J.M.; Naghavi, A.O.; Oplt, T.; Creighton, C.J.; Shaw, C.; Benham, A.L.; Kim, J.; Soibam, B.; Harris, R.A.; et al. Proinflammatory role for let-7 microRNAS in experimental asthma. J. Biol. Chem. 2010, 285, 30139–30149. [Google Scholar] [CrossRef]
- Yang, G.; Wang, T.; Qu, X.; Chen, S.; Han, Z.; Chen, S.; Chen, M.; Lin, J.; Yu, S.; Gao, L.; et al. Exosomal miR-21/Let-7a ratio distinguishes non-small cell lung cancer from benign pulmonary diseases. Asia Pac. J. Clin. Oncol. 2020, 16, 280–286. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay Name: | Assay ID | Species | miRNA Sequence |
|---|---|---|---|
| hsa-miR-16-5p | 000391 | human | UAGCAGCACGUAAAUAUUGGCG |
| hsa-miR-21-5p | 000397 | human | UAGCUUAUCAGACUGAUGUUGA |
| hsa-let-7a-5p | 000377 | human | UGAGGUAGUAGGUUGUAUAGUU |
| hsa-miR-223-5p | 002098 | human | CGUGUAUUUGACAAGCUGAGUU |
| ALL n = 120 | HC n = 30 | MM n = 50 | SA n = 40 | p-Value | TUKEYP OST-HOC | |
|---|---|---|---|---|---|---|
| Demographic data | ||||||
| sex, M/F | 43/77 | 16/14 | 17/33 | 9/31 | 0.0248 * | SA > HC = MM |
| age, years | 53 ± 13.0 | 50 ± 9.7 | 56 ± 15.7 | 56 ± 10.6 | 0.1976 | - |
| BMI, kg/m2 | 29 ± 6.7 | 27 ± 5.4 | 30 ± 6.5 | 29 ± 7.2 | 0.4689 | - |
| Lung function parameters | ||||||
| FEV1, % | 85 ± 20.0 | 96 ± 20.3 | 86 ± 16.5 | 78 ± 20.8 | 0.0046 * | HC > MM > SA |
| FEV1/FVC, % | 76 ± 12.9 | 81 ± 6.0 | 80 ± 11.9 | 69 ± 13.6 | <0.0001 ** | HC = MM > SA |
| FVC, % | 99 ± 17.5 | 93 ± 17.4 | 100 ± 15.3 | 101 ± 15.0 | 0.2387 | - |
| Asthma biomarkers | ||||||
| FeNO50, ppb | 27 ± 35.1 | 9.9 ± 3.5 | 25 ± 20.8 | 28 ± 41.1 | 0.5546 | - |
| Blood eosinophils, % total cells | 14.7 ± 42.8 | 1.9 ± 0.7 | 4 ± 3.2 | 11 ± 27.5 | 0.6637 | - |
| Heading | Heading | FEV1, % | FVC, % | FEV1/FVC, % | FeNO50, % |
|---|---|---|---|---|---|
| miR-21 | serum | −0.1433 | 0.0714 | 0.0359 | 0.0285 |
| exosome | −0.1139 * | 0.1478 | −0.1551 * | 0.0893 | |
| let-7a | serum | −0.0344 | −0.1773 | −0.2152 | 0.0623 |
| exosome | −0.2418 ** | −0.1165 | −0.3680 ** | 0.1039 | |
| miR-223 | serum | 0.0536 | 0.1006 | 0.1590 | −0.0133 |
| exosome | −0.0004 * | −0.0131 | −0.0780 | −0.0827 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soccio, P.; Moriondo, G.; Lacedonia, D.; Tondo, P.; Pescatore, D.; Quarato, C.M.I.; Carone, M.; Foschino Barbaro, M.P.; Scioscia, G. MiRNA and Exosomal miRNA as New Biomarkers Useful to Phenotyping Severe Asthma. Biomolecules 2023, 13, 1542. https://doi.org/10.3390/biom13101542
Soccio P, Moriondo G, Lacedonia D, Tondo P, Pescatore D, Quarato CMI, Carone M, Foschino Barbaro MP, Scioscia G. MiRNA and Exosomal miRNA as New Biomarkers Useful to Phenotyping Severe Asthma. Biomolecules. 2023; 13(10):1542. https://doi.org/10.3390/biom13101542
Chicago/Turabian StyleSoccio, Piera, Giorgia Moriondo, Donato Lacedonia, Pasquale Tondo, Dalila Pescatore, Carla Maria Irene Quarato, Mauro Carone, Maria Pia Foschino Barbaro, and Giulia Scioscia. 2023. "MiRNA and Exosomal miRNA as New Biomarkers Useful to Phenotyping Severe Asthma" Biomolecules 13, no. 10: 1542. https://doi.org/10.3390/biom13101542
APA StyleSoccio, P., Moriondo, G., Lacedonia, D., Tondo, P., Pescatore, D., Quarato, C. M. I., Carone, M., Foschino Barbaro, M. P., & Scioscia, G. (2023). MiRNA and Exosomal miRNA as New Biomarkers Useful to Phenotyping Severe Asthma. Biomolecules, 13(10), 1542. https://doi.org/10.3390/biom13101542