
Aggravated Gut Microbiota and Metabolomic Imbalances Are Associated with Hypertension Patients Comorbid with Atrial Fibrillation


Abstract
1. Introduction
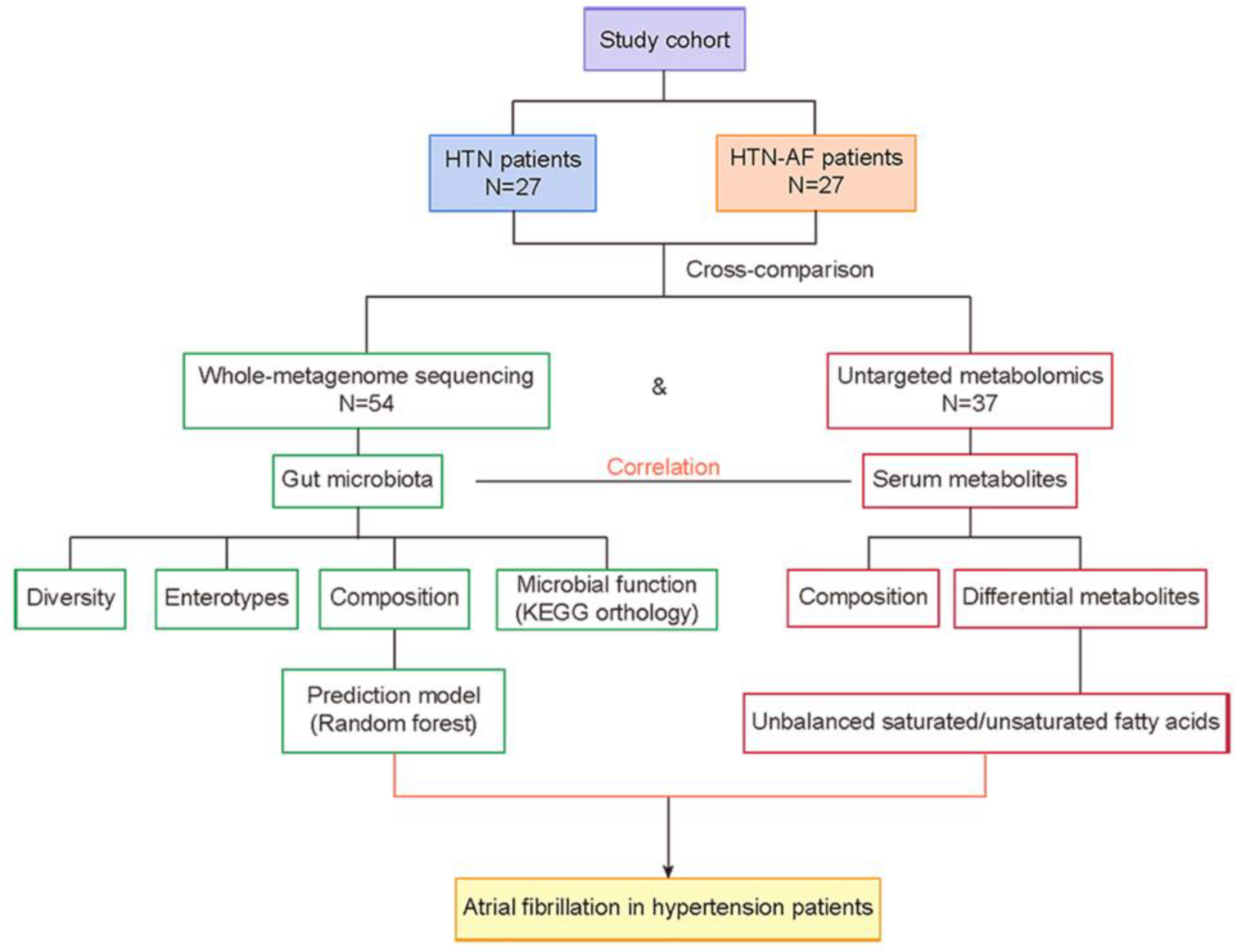
2. Materials and Methods
2.1. Study Cohort
2.2. GM Analysis by Metagenomics and Metabolomics
2.3. Statistical Analysis
3. Results
3.1. Baseline Characteristics of the Study Cohort
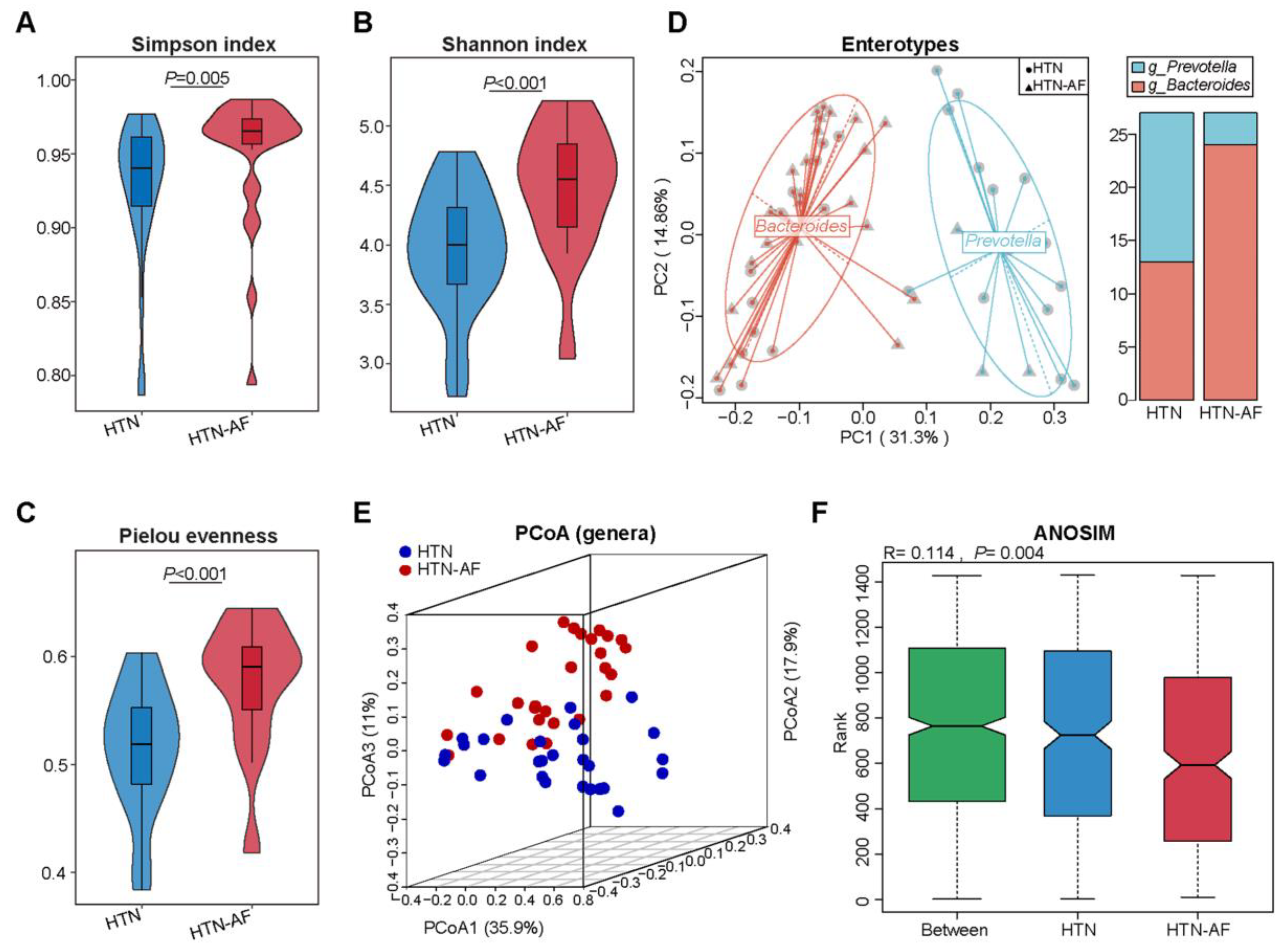
3.2. Altered Gut Microbial Diversity and Enterotype Distribution in HTN Patients with AF
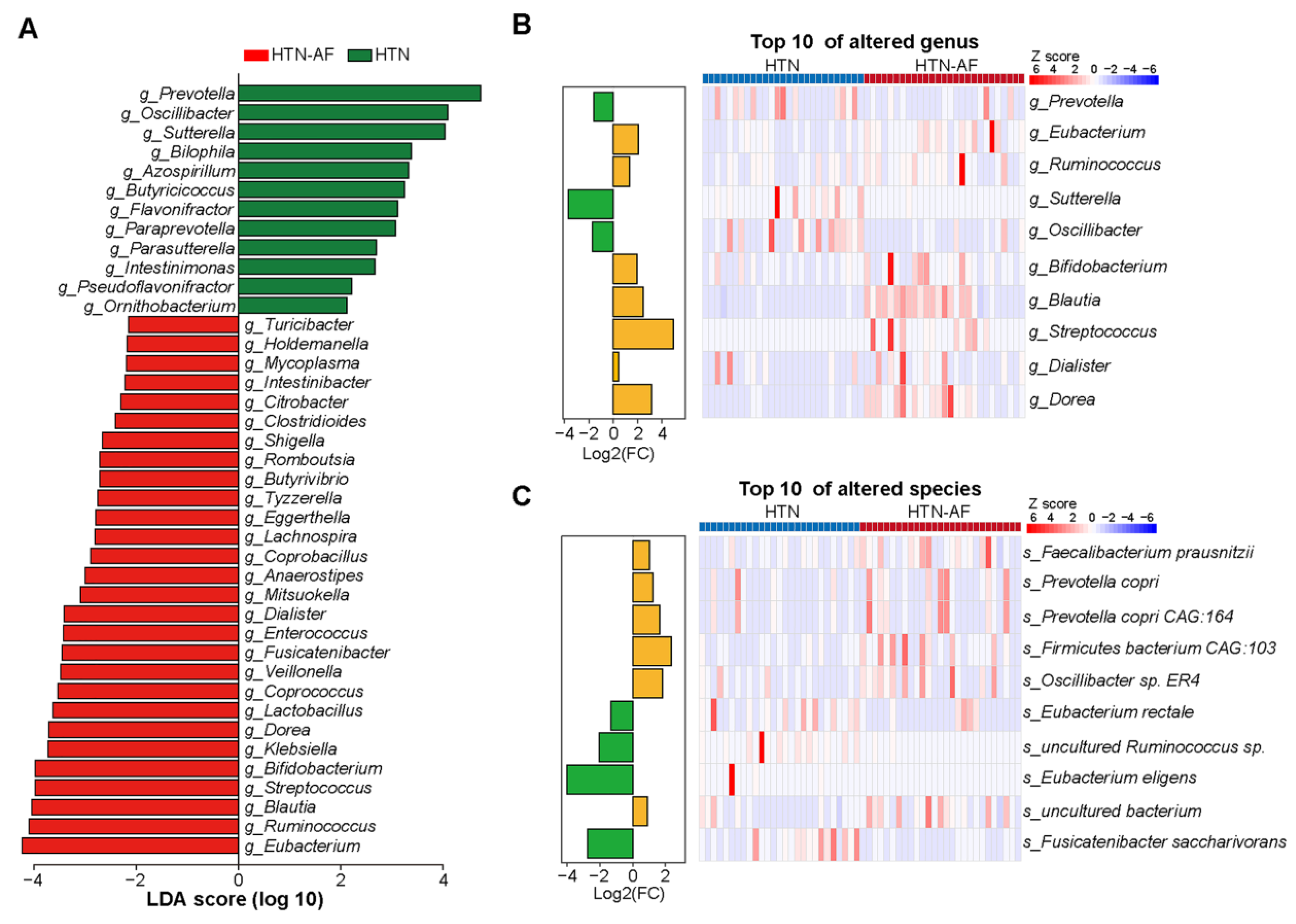
3.3. Distinct Gut Microbial Compositions between HTN and HTN-AF Groups
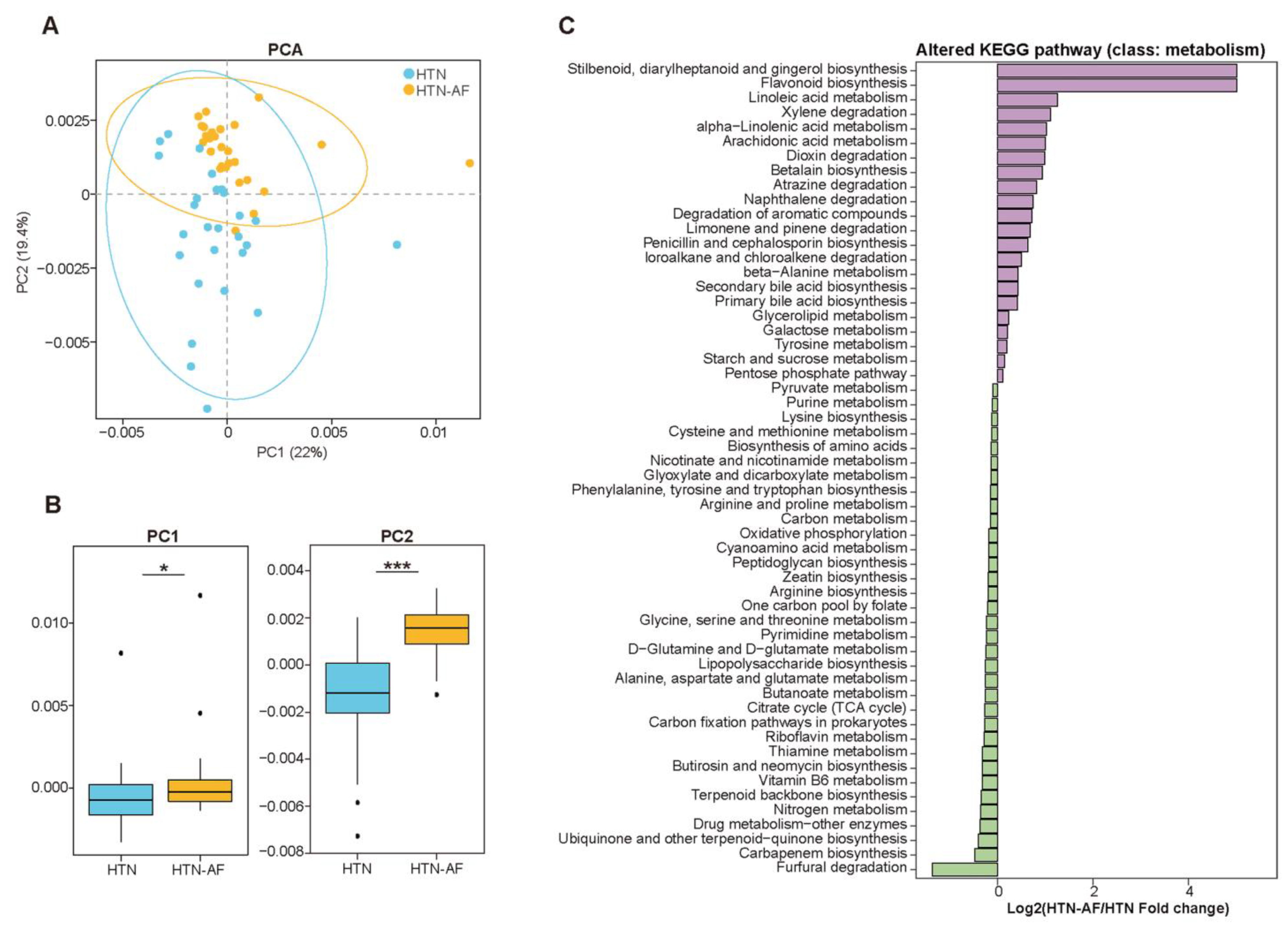
3.4. Disturbance of GM Functions in HTN-AF Patients
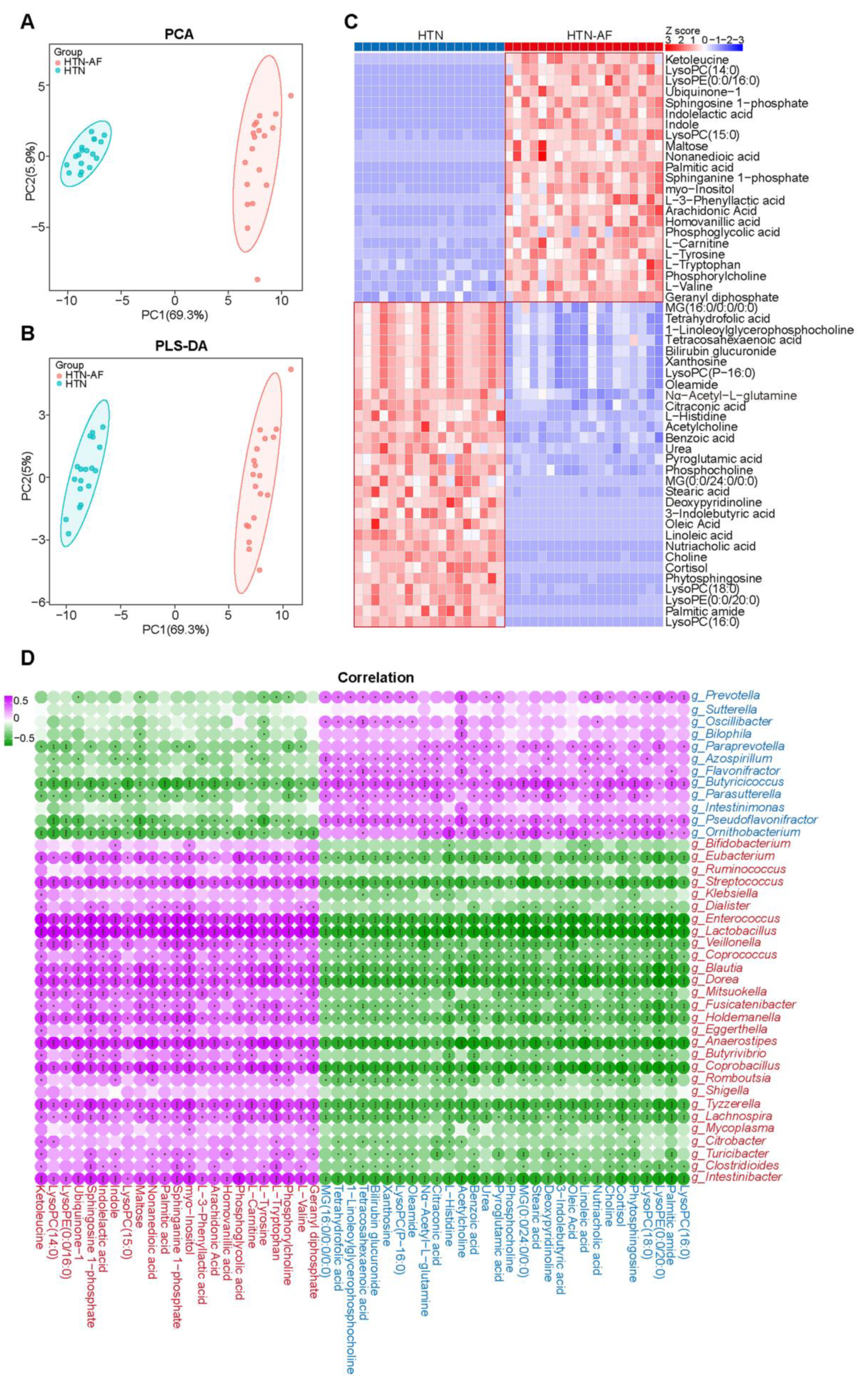
3.5. AF Occurrence in HTN Patients was Related to Changed Serum Metabolomics
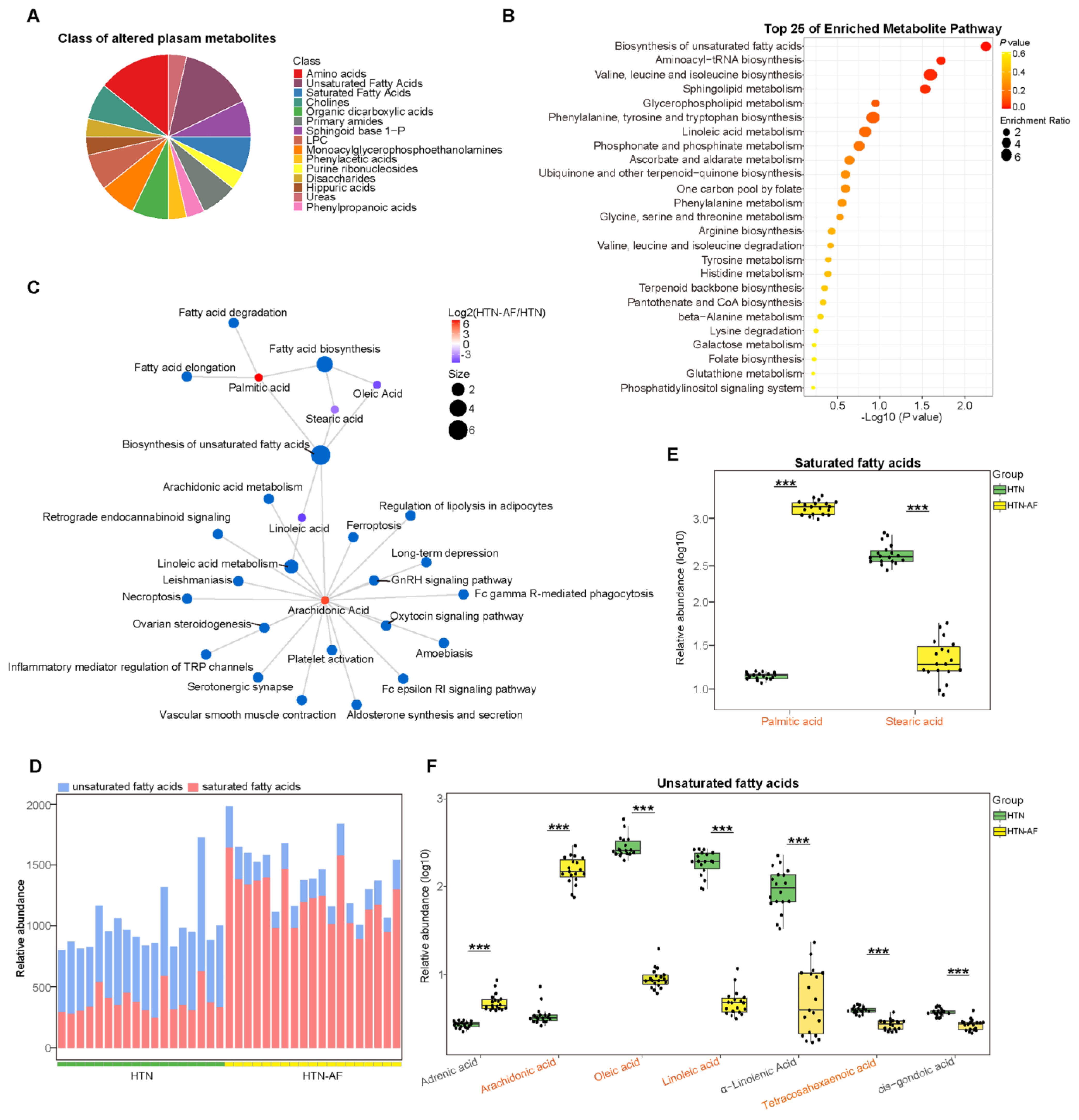
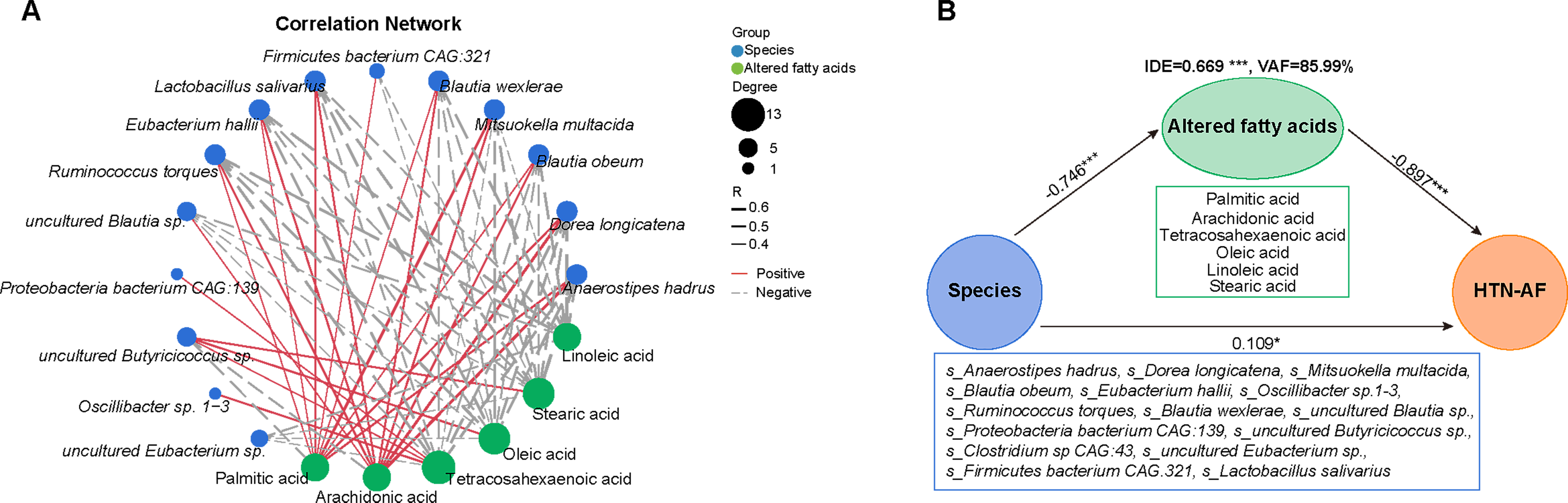
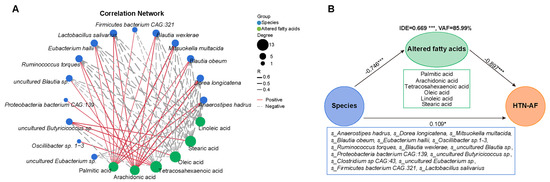
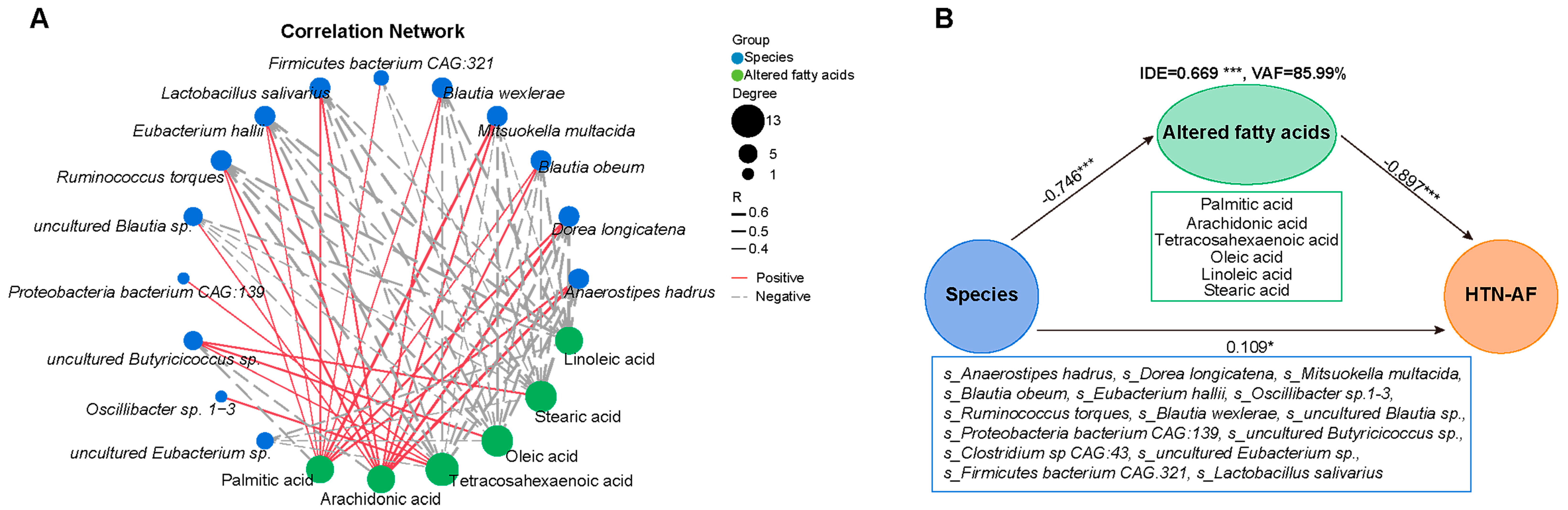
3.6. Unbalanced Saturated and Unsaturated Fatty Acids in HTN Patients with AF
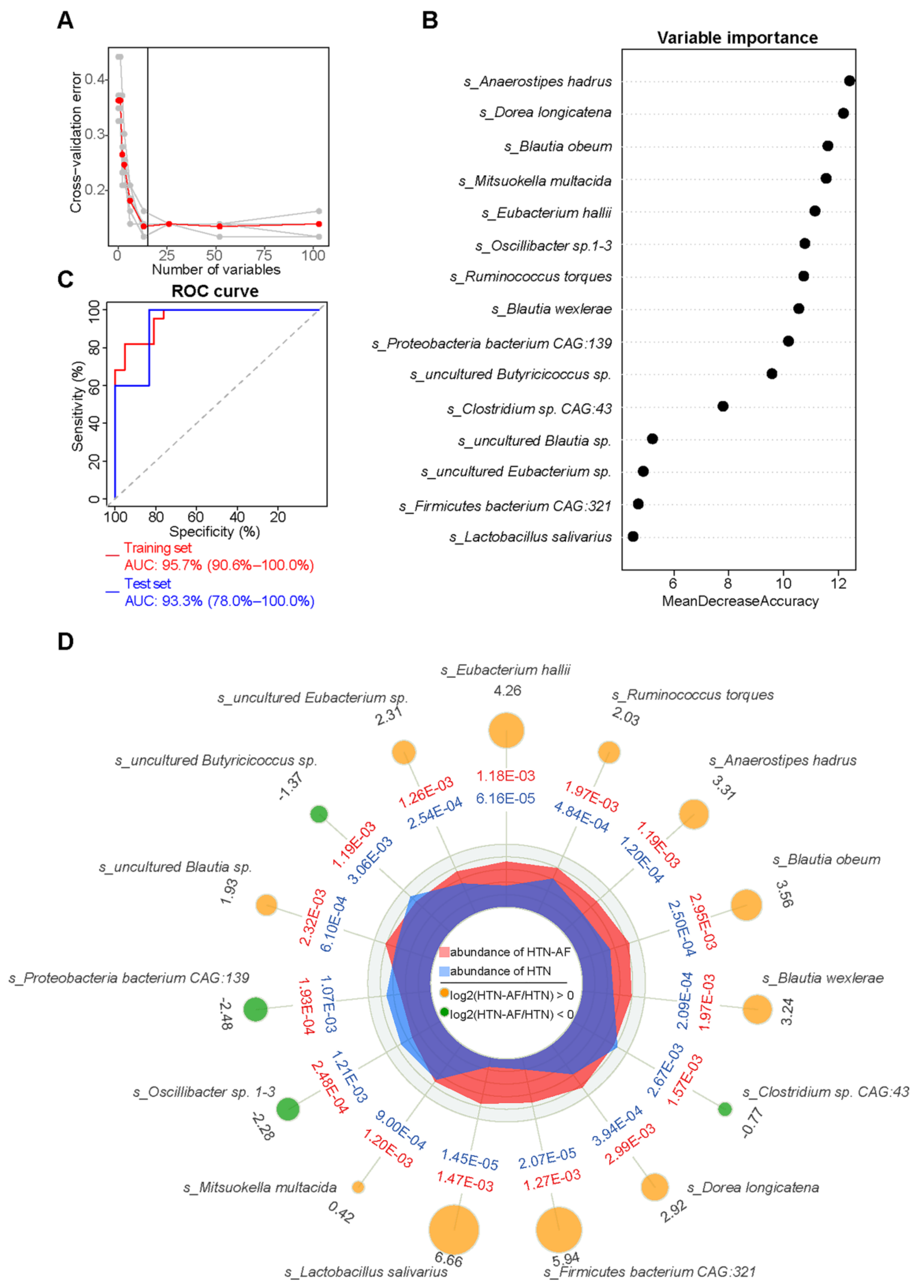
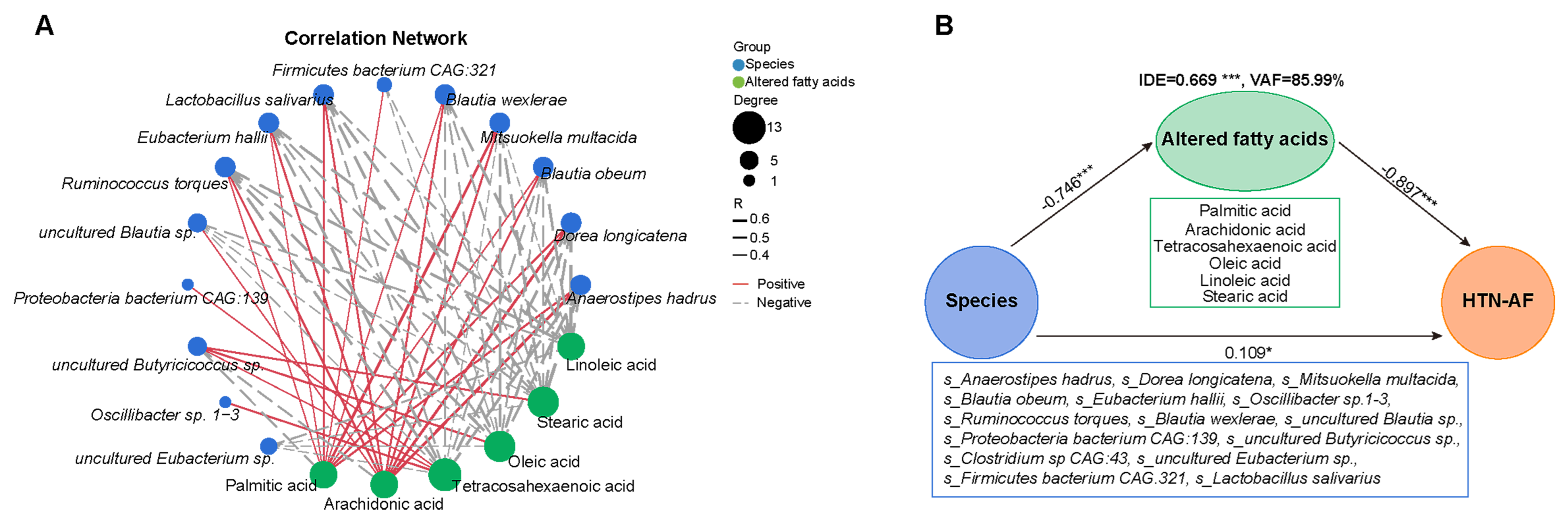
3.7. A Risk Prediction Model based on Gut Microbial Signatures for AF in HTN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Go, A.S.; Hylek, E.M.; Phillips, K.A.; Chang, Y.; Henault, L.E.; Selby, J.V.; Singer, D.E. Prevalence of diagnosed atrial fibrillation in adults: National implications for rhythm management and stroke prevention: The AnTicoagulation and Risk Factors in Atrial Fibrillation (ATRIA) Study. JAMA 2001, 285, 2370–2375. [Google Scholar] [CrossRef] [PubMed]
- Huxley, R.R.; Lopez, F.L.; Folsom, A.R.; Agarwal, S.K.; Loehr, L.R.; Soliman, E.Z.; Maclehose, R.; Konety, S.; Alonso, A. Ab-solute and attributable risks of atrial fibrillation in relation to optimal and borderline risk factors: The Atherosclerosis Risk in Communities (ARIC) study. Circulation 2011, 123, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Levy, D.; Vaziri, S.M.; D’Agostino, R.B.; Belanger, A.J.; Wolf, P.A. Independent risk factors for atrial fibrillation in a population-based cohort. The Framingham Heart Study. JAMA 1994, 271, 840–844. [Google Scholar] [CrossRef]
- Parcha, V.; Patel, N.; Kalra, R.; Kim, J.; Gutiérrez, O.M.; Arora, G.; Arora, P. Incidence and Implications of Atrial Fibrilla-tion/Flutter in Hypertension: Insights From the SPRINT Trial. Hypertension 2020, 75, 1483–1490. [Google Scholar] [CrossRef]
- Witkowski, M.; Weeks, T.L.; Hazen, S.L. Gut Microbiota and Cardiovascular Disease. Circ. Res. 2020, 127, 553–570. [Google Scholar] [CrossRef] [PubMed]
- Gawałko, M.; Agbaedeng, T.A.; Saljic, A.; Müller, D.N.; Wilck, N.; Schnabel, R.; Penders, J.; Rienstra, M.; van Gelder, I.; Jespersen, T.; et al. Gut microbiota, dysbiosis and atrial fibrillation. Arrhythmogenic mechanisms and potential clinical impli-cations. Cardiovasc. Res. 2022, 118, 2415–2427. [Google Scholar] [CrossRef] [PubMed]
- Verhaar, B.J.H.; Prodan, A.; Nieuwdorp, M.; Muller, M. Gut Microbiota in Hypertension and Atherosclerosis: A Review. Nutrients 2020, 12, 2982. [Google Scholar] [CrossRef] [PubMed]
- Muralitharan, R.R.; Jama, H.A.; Xie, L.; Peh, A.; Snelson, M.; Marques, F.Z. Microbial Peer Pressure: The Role of the Gut Mi-crobiota in Hypertension and Its Complications. Hypertension 2020, 76, 1674–1687. [Google Scholar] [CrossRef]
- Zuo, K.; Yin, X.; Li, K.; Zhang, J.; Wang, P.; Jiao, J.; Liu, Z.; Liu, X.; Liu, J.; Li, J.; et al. Different Types of Atrial Fibrillation Share Patterns of Gut Microbiota Dysbiosis. mSphere 2020, 5, e00071-20. [Google Scholar] [CrossRef]
- Li, J.; Zuo, K.; Zhang, J.; Hu, C.; Wang, P.; Jiao, J.; Liu, Z.; Yin, X.; Liu, X.; Li, K.; et al. Shifts in gut microbiome and metabolome are associated with risk of recurrent atrial fibrillation. J. Cell. Mol. Med. 2020, 24, 13356–13369. [Google Scholar] [CrossRef]
- Zuo, K.; Li, J.; Wang, P.; Liu, Y.; Liu, Z.; Yin, X.; Liu, X.; Yang, X. Duration of Persistent Atrial Fibrillation Is Associated with Alterations in Human Gut Microbiota and Metabolic Phenotypes. mSystems 2019, 4, e00422-19. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zuo, K.; Zhang, W.; Zhong, J.; Li, J.; Xu, L.; Yang, X. Association between Gut Microbiota Dysbiosis and the CHA2DS2-VASc Score in Atrial Fibrillation Patients. Int. J. Clin. Pract. 2022, 2022, 7942605. [Google Scholar] [CrossRef] [PubMed]
- Zuo, K.; Li, J.; Li, K.; Hu, C.; Gao, Y.; Chen, M.; Hu, R.; Liu, Y.; Chi, H.; Wang, H.; et al. Disordered gut microbiota and al-terations in metabolic patterns are associated with atrial fibrillation. Gigascience 2019, 8, giz058. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.W.; Pham, H.P.; Bridonneau, C.; Aubry, C.; Lamas, B.; Martin-Gallausiaux, C.; Moroldo, M.; Rainteau, D.; Lapaque, N.; Six, A.; et al. Microorganisms linked to inflammatory bowel disease-associated dysbiosis differentially impact host physiology in gnotobiotic mice. ISME J. 2016, 10, 460–477. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.Y.; Liu, Q.Q.; Su, H.Z.; Zhang, H.P.; Fan, J.M.; Yang, J.H.; Hu, A.K.; Liu, Y.Q.; Chou, D.; Zeng, Y.M. Gut microbiota in obstructive sleep apnea-hypopnea syndrome: Disease-related dysbiosis and metabolic comorbidities. Clin. Sci. 2019, 133, 905–917. [Google Scholar] [CrossRef]
- Zhuang, Z.Q.; Shen, L.L.; Li, W.W.; Fu, X.; Zeng, F.; Gui, L.; Lü, Y.; Cai, M.; Zhu, C.; Tan, Y.L.; et al. Gut Microbiota is Altered in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in ath-erosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Ter Horst, R.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the Human Gut Microbiome to Inflammatory Cytokine Production Capacity. Cell 2016, 167, 1125–1136. [Google Scholar] [CrossRef]
- Klemm, S.; Ruland, J. Inflammatory signal transduction from the Fc epsilon RI to NF-kappa B. Immunobiology 2006, 211, 815–820. [Google Scholar] [CrossRef]
- Neefs, J.; van den Berg, N.W.; Limpens, J.; Berger, W.R.; Boekholdt, S.M.; Sanders, P.; de Groot, J.R. Aldosterone Pathway Blockade to Prevent Atrial Fibrillation: A Systematic Review and Meta-Analysis. Int. J. Cardiol. 2017, 231, 155–161. [Google Scholar] [CrossRef]
- Reil, J.C.; Hohl, M.; Selejan, S.; Lipp, P.; Drautz, F.; Kazakow, A.; Münz, B.M.; Müller, P.; Steendijk, P.; Reil, G.H.; et al. Al-dosterone promotes atrial fibrillation. Eur. Heart J. 2012, 33, 2098–2108. [Google Scholar] [CrossRef]
- Milovanovic, M.; Fransson, E.; Hallert, C.; Järemo, P. Atrial fibrillation and platelet reactivity. Int. J. Cardiol. 2010, 145, 357–358. [Google Scholar] [CrossRef] [PubMed]
- Bakogiannis, C.; Sachse, M.; Stamatelopoulos, K.; Stellos, K. Platelet-derived chemokines in inflammation and atherosclerosis. Cytokine 2019, 122, 154157. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Kong, B.; Shuai, W.; Xiao, Z.; Dai, C.; Qin, T.; Gong, Y.; Zhu, J.; Liu, Q.; Huang, H. Ferroportin-mediated ferroptosis involved in new-onset atrial fibrillation with LPS-induced endotoxemia. Eur. J. Pharmacol. 2021, 913, 174622. [Google Scholar] [CrossRef]
- Yu, Y.; Yan, Y.; Niu, F.; Wang, Y.; Chen, X.; Su, G.; Liu, Y.; Zhao, X.; Qian, L.; Liu, P.; et al. Ferroptosis: A cell death connecting oxidative stress, inflammation and cardiovascular diseases. Cell. Death Discov. 2021, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Jiang, T.; Sun, H.; Li, T.; Gao, F.; Fan, B.; Li, X.; Qin, X.; Zheng, Q. Necroptosis is required for atrial fibrillation and involved in aerobic exercise-conferred cardioprotection. J. Cell. Mol. Med. 2021, 25, 8363–8375. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Hu, Y.F.; Chen, Y.J.; Lin, Y.J.; Chen, S.A. Inflammation and the pathogenesis of atrial fibrillation. Nat. Rev. Cardiol. 2015, 12, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, H.; Zhai, L.; Ye, B.; Cheng, Y.; Zhai, C. ALA/LA ameliorates glucose toxicity on HK-2 cells by attenuating oxidative stress and apoptosis through the ROS/p38/TGF-β1 pathway. Lipids Health Dis. 2017, 16, 216. [Google Scholar] [CrossRef] [PubMed]
- Al-Shudiefat, A.A.; Sharma, A.K.; Bagchi, A.K.; Dhingra, S.; Singal, P.K. Oleic acid mitigates TNF-α-induced oxidative stress in rat cardiomyocytes. Mol. Cell. Biochem. 2013, 372, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Haworth, R.A.; Potter, K.T.; Russell, D.C. Role of arachidonic acid, lipoxygenase, and mitochondrial depolarization in reper-fusion arrhythmias. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H165–H174. [Google Scholar] [CrossRef]
- Mangali, S.; Bhat, A.; Udumula, M.P.; Dhar, I.; Sriram, D.; Dhar, A. Inhibition of protein kinase R protects against palmitic acid-induced inflammation, oxidative stress, and apoptosis through the JNK/NF-kB/NLRP3 pathway in cultured H9C2 car-diomyocytes. J. Cell. Biochem. 2019, 120, 3651–3663. [Google Scholar] [CrossRef] [PubMed]
- Jalife, J.; Kaur, K. Atrial remodeling, fibrosis, and atrial fibrillation. Trends Cardiovasc. Med. 2015, 25, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Fretts, A.M.; Mozaffarian, D.; Siscovick, D.S.; Djousse, L.; Heckbert, S.R.; King, I.B.; McKnight, B.; Sitlani, C.; Sacks, F.M.; Song, X.; et al. Serum phospholipid saturated fatty acids and incident atrial fibrillation: The Cardiovascular Health Study. J. Am. Heart. Assoc. 2014, 3, e000889. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HTN | HTN-AF | p Value | |
|---|---|---|---|
| Number | 27 | 27 | |
| Male (%) | 26 (96.30) | 18 (66.67) | 0.011 |
| DM (%) | 0 (0.00) | 7 (25.93) | 0.010 |
| BMI, kg/m2 | 25.80 ± 3.06 | 27.10 ± 5.56 | 0.298 |
| Age, years | 53.59 ± 4.89 | 68.74 ± 8.40 | <0.001 |
| TC, mmol/L | 4.59 ± 1.04 | 3.89 ± 1.11 | 0.020 |
| TG, mmol/L | 1.43 ± 0.87 | 1.67 ± 1.53 | 0.478 |
| LDL, mmol/L | 3.06 ± 3.27 | 2.16 ± 0.97 | 0.176 |
| ALT, U/L | 21.56 ± 9.65 | 21.67 ± 11.10 | 0.971 |
| sCr, μmol/L | 70.00 (59.75, 92.54) | 69.30 (59.00, 86.30) | 0.663 |
| FBG, mmol/L | 5.29 (4.55, 5.71) | 4.79 (4.50, 6.01) | 0.961 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, C.; Zuo, K.; Fu, Y.; Zhu, X.; Li, J.; Zhong, J.; Xu, L.; Yang, X. Aggravated Gut Microbiota and Metabolomic Imbalances Are Associated with Hypertension Patients Comorbid with Atrial Fibrillation. Biomolecules 2022, 12, 1445. https://doi.org/10.3390/biom12101445
Fang C, Zuo K, Fu Y, Zhu X, Li J, Zhong J, Xu L, Yang X. Aggravated Gut Microbiota and Metabolomic Imbalances Are Associated with Hypertension Patients Comorbid with Atrial Fibrillation. Biomolecules. 2022; 12(10):1445. https://doi.org/10.3390/biom12101445
Chicago/Turabian StyleFang, Chen, Kun Zuo, Yuan Fu, Xiaoming Zhu, Jing Li, Jiuchang Zhong, Li Xu, and Xinchun Yang. 2022. "Aggravated Gut Microbiota and Metabolomic Imbalances Are Associated with Hypertension Patients Comorbid with Atrial Fibrillation" Biomolecules 12, no. 10: 1445. https://doi.org/10.3390/biom12101445
APA StyleFang, C., Zuo, K., Fu, Y., Zhu, X., Li, J., Zhong, J., Xu, L., & Yang, X. (2022). Aggravated Gut Microbiota and Metabolomic Imbalances Are Associated with Hypertension Patients Comorbid with Atrial Fibrillation. Biomolecules, 12(10), 1445. https://doi.org/10.3390/biom12101445