
Decoupling of Plant Growth and Accumulation of Biologically Active Compounds in Leaves, Roots, and Root Exudates of Hypericum perforatum L. by the Combination of Jasmonate and Far-Red Lighting



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Performance
2.2. Growth Conditions
2.3. Experimental Setup
2.4. Lightning Conditions and FR Supply
2.5. Application of MeJA
2.6. Root Exudate Collection
2.7. Final Sampling
2.8. Analysis of Roots and Root Hairs
2.9. Extraction of Bioactive Compounds from Leaves and Roots
2.10. Collection of Root Exudates
2.11. Assay of Total Phenolic Content
2.12. Quantitative Determination of Phenolic Compounds by UHPLC
2.13. Quantitative Determination of Pigments, Sugars, and Ions
2.14. Statistics
3. Results
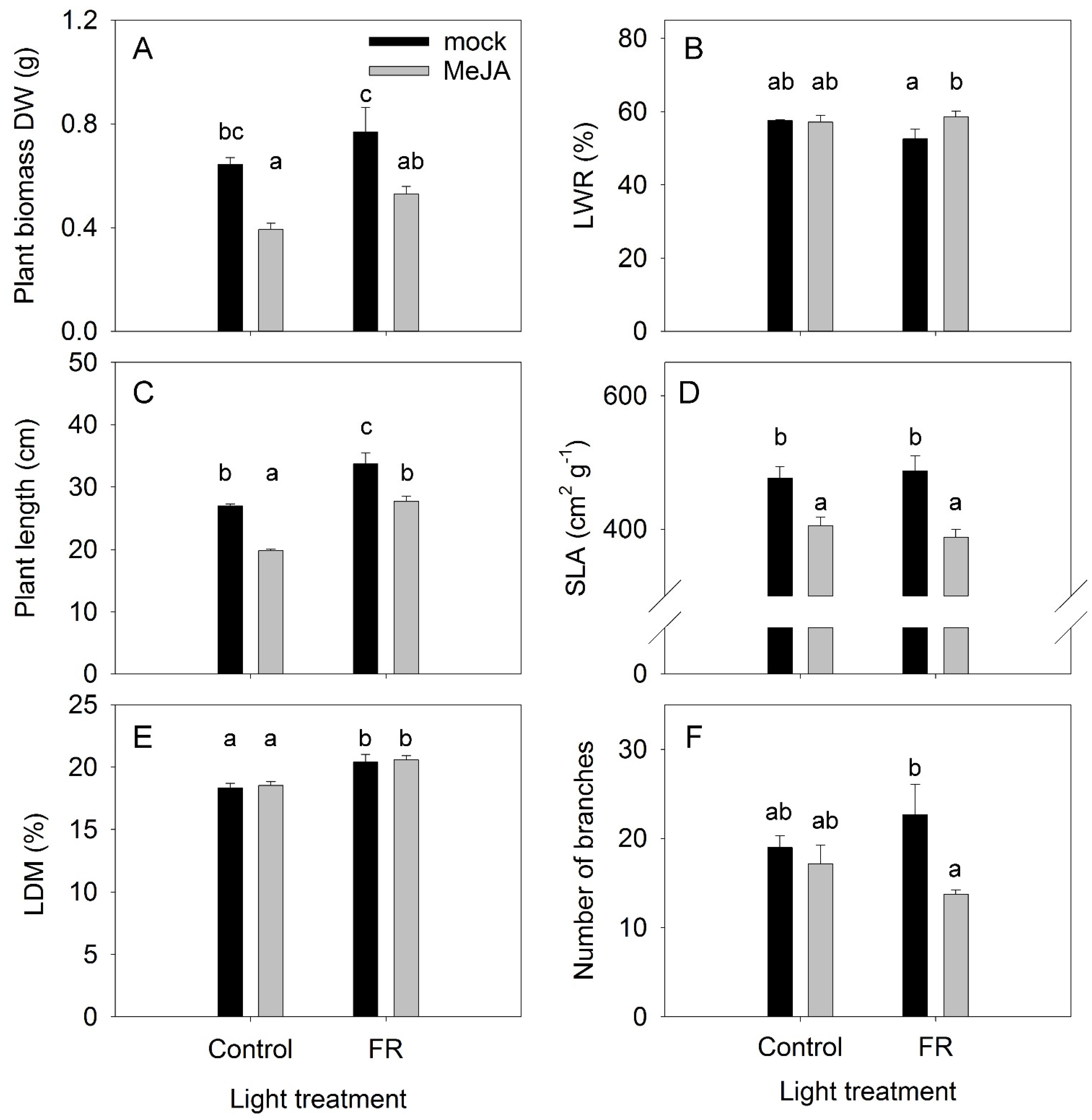
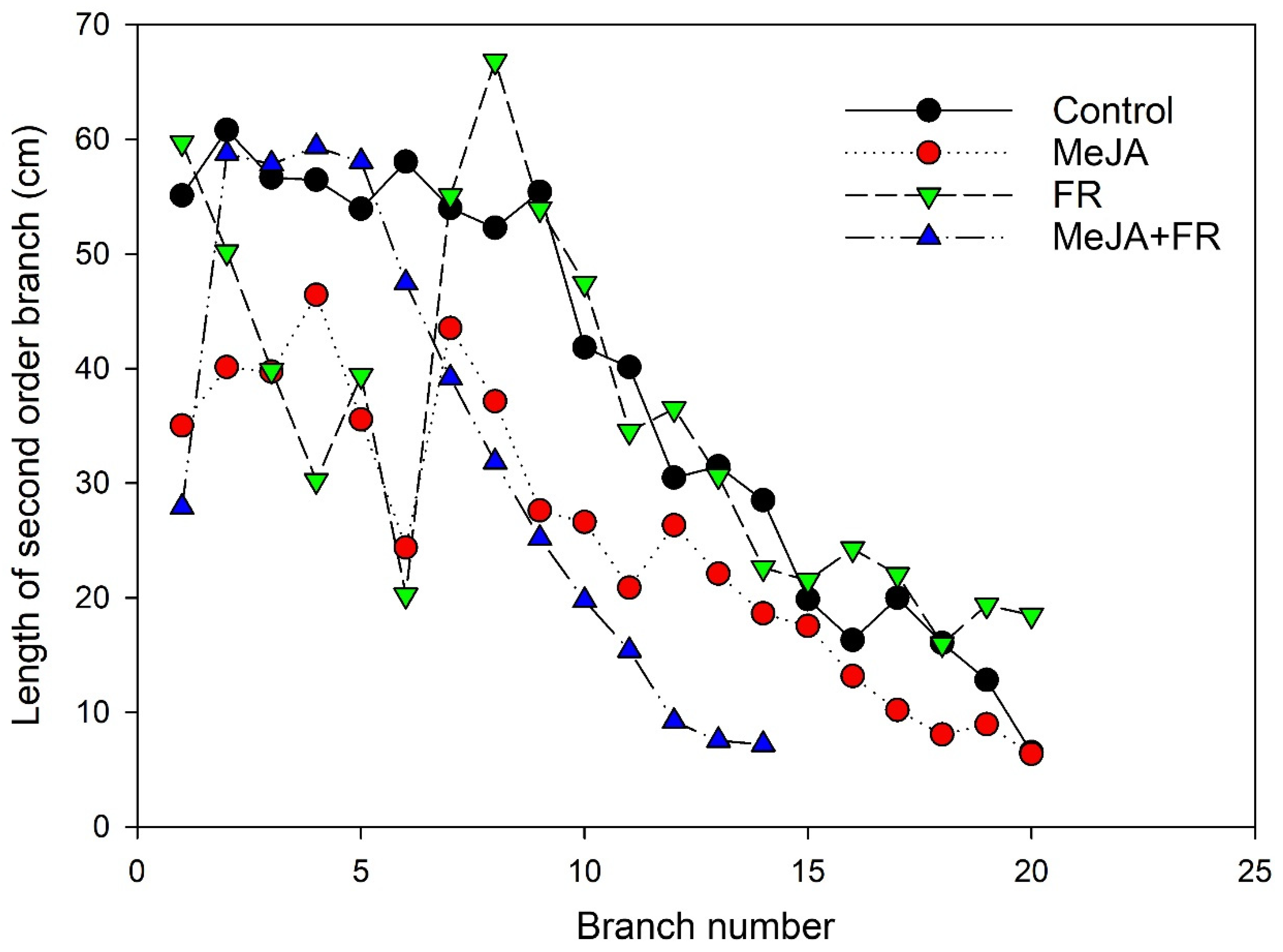
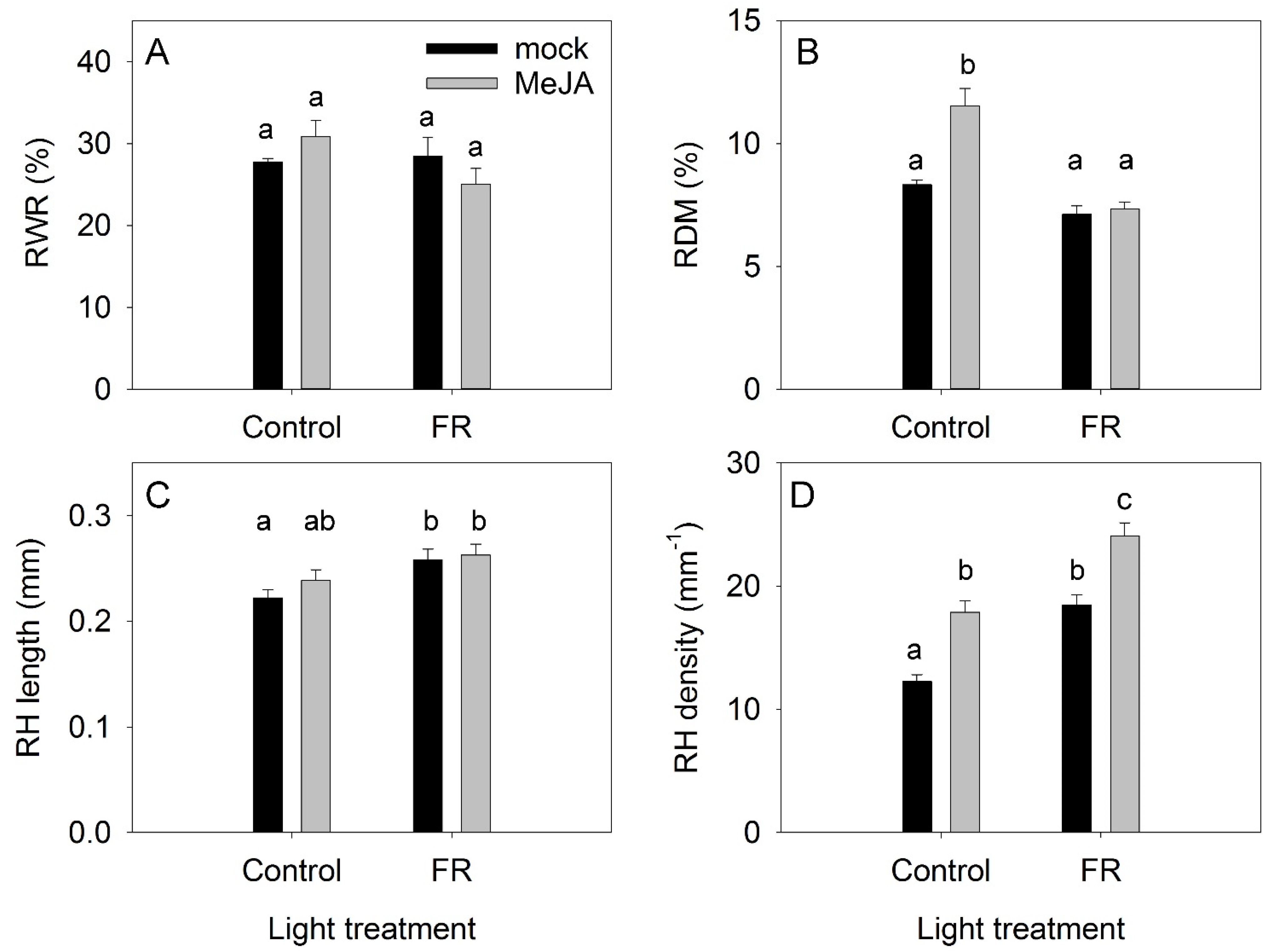
3.1. Plant Growth Traits
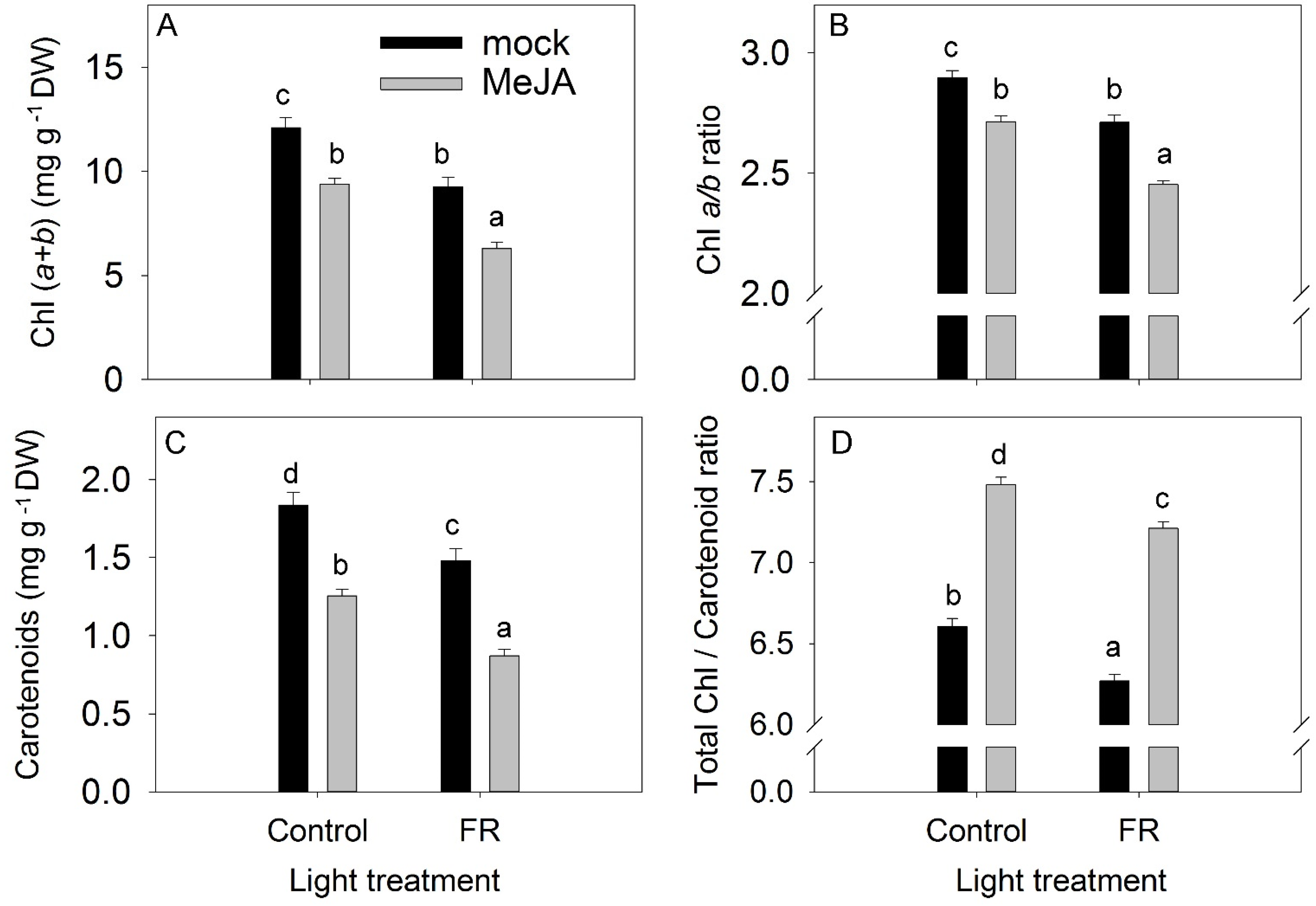
3.2. Pigments
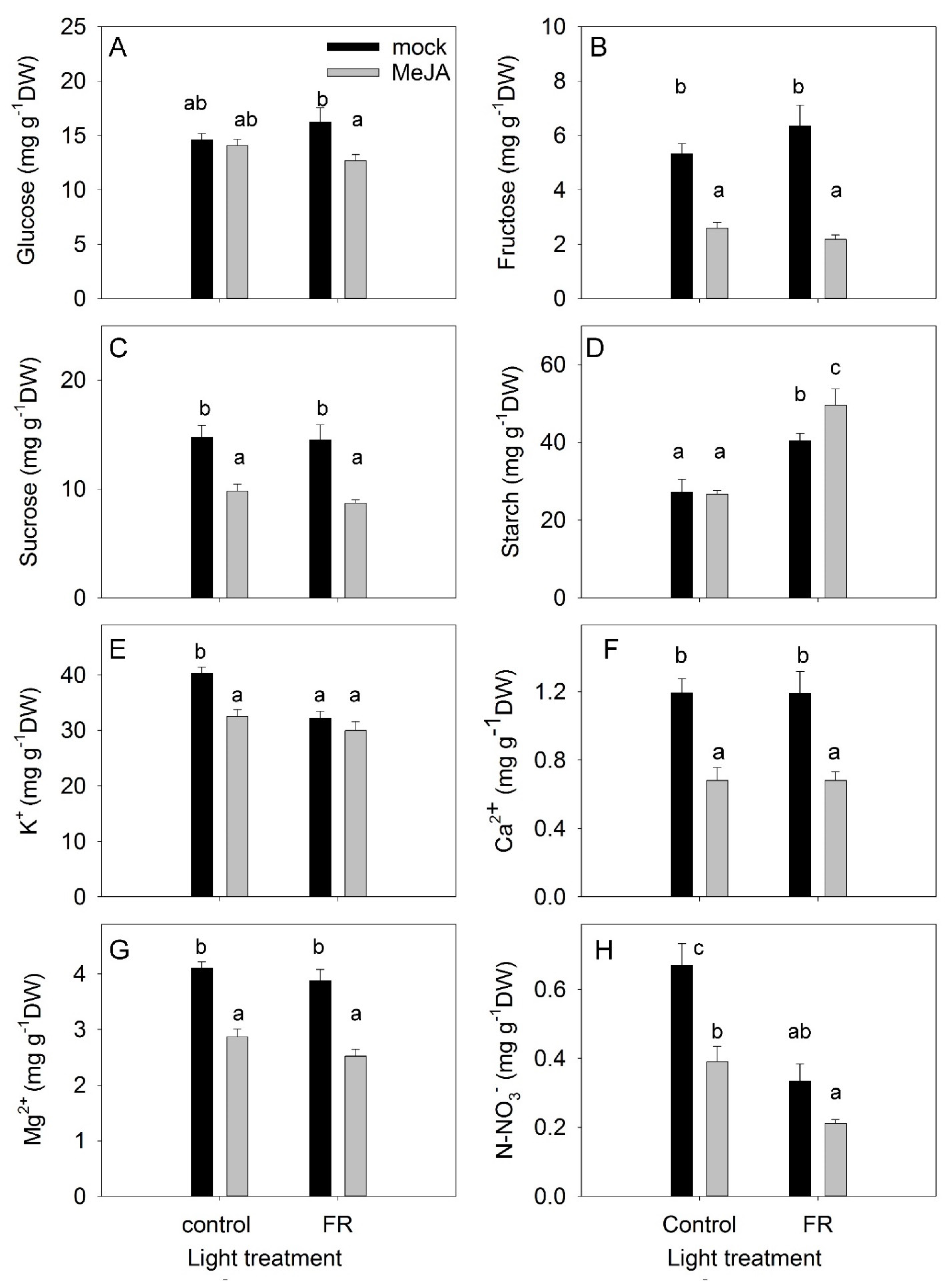
3.3. Nonstructural Carbohydrates
3.4. Ions
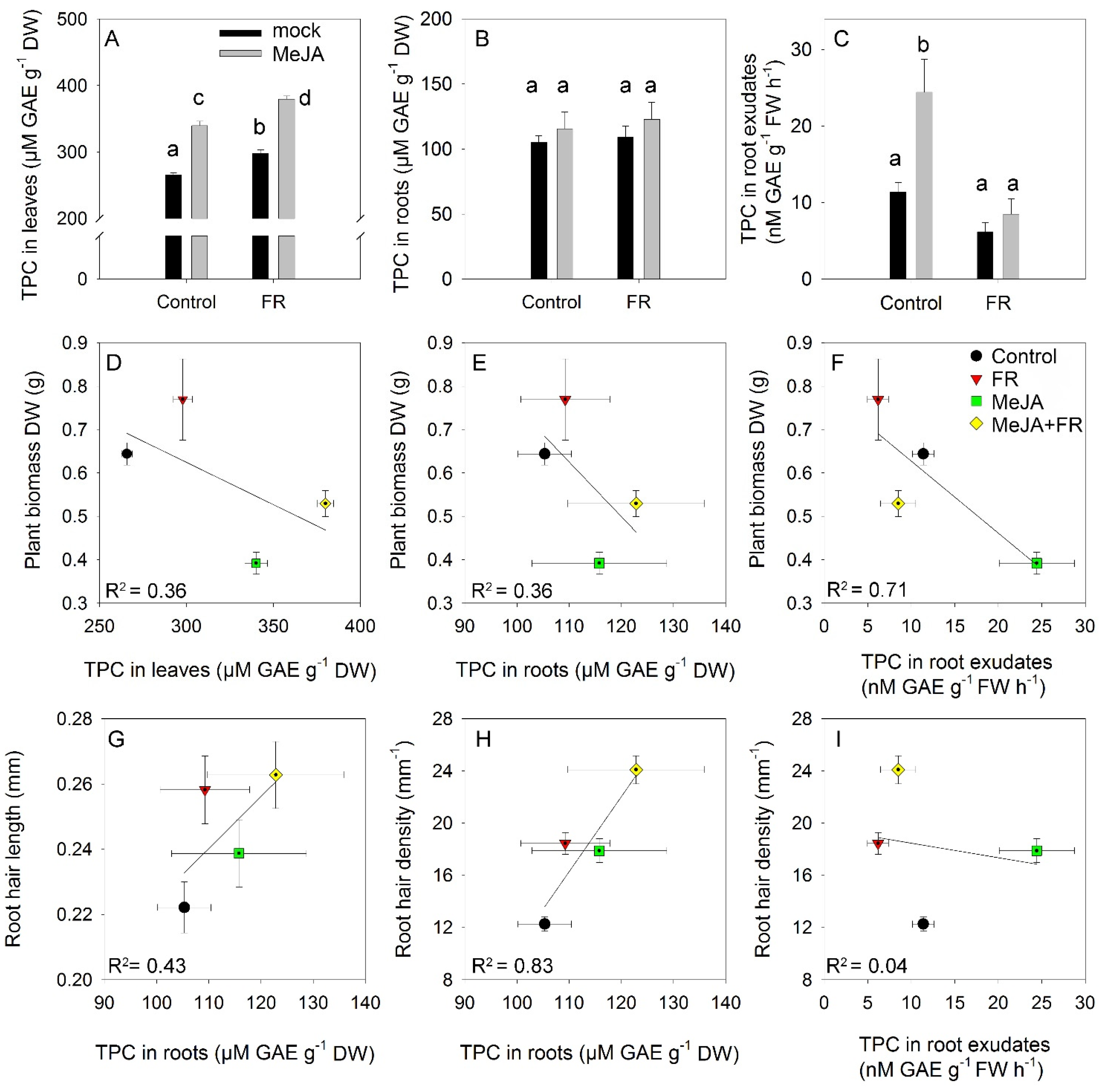
3.5. Total Phenolics
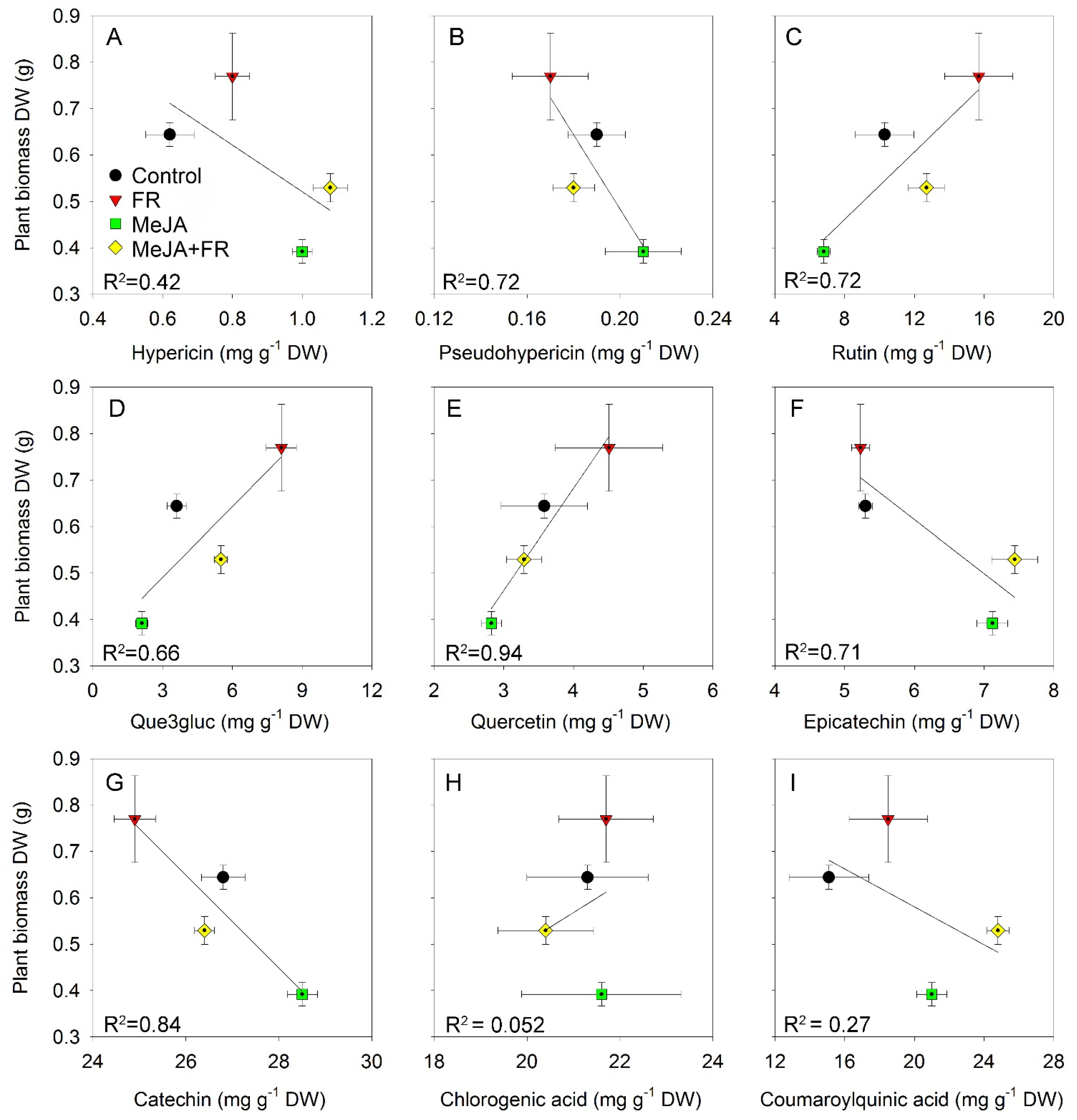
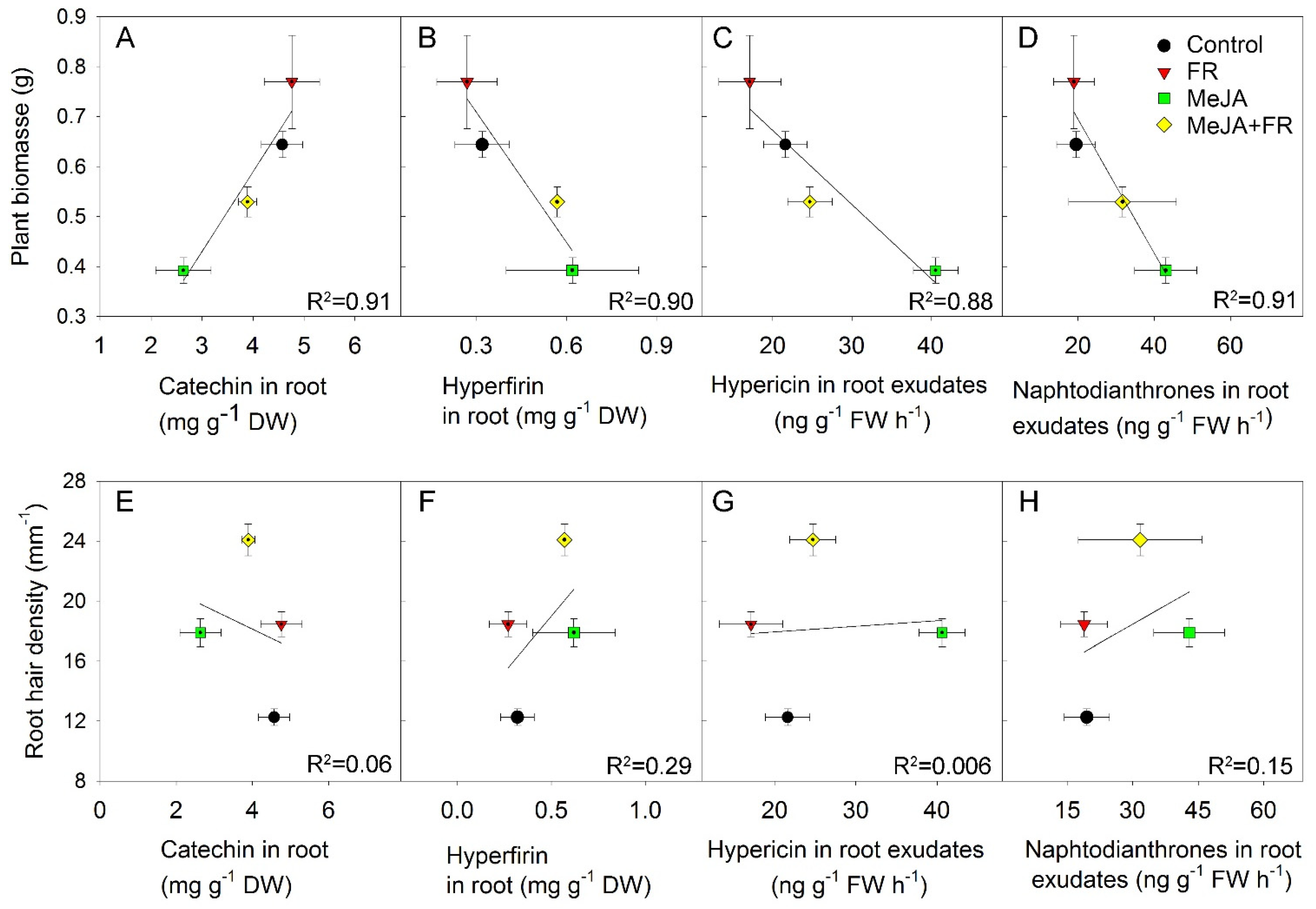
3.6. Relation between Growth Traits and Concentration of Biologically Active Compounds
4. Discussion
4.1. Decoupling of Growth and Accumulation of Secondary Compounds
4.2. Cross-Talk between JA and FR and Effects on Plant Growth
4.3. Cross-Talk between JA and FR and Effects on the Accumulation of Biologically Active Compounds in Plants
5. Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sahoo, N.; Manchikanti, P.; Dey, S. Herbal drugs: Standards and regulation. Fitoterapia 2010, 81, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Cipollini, D.; Walters, D.; Voelckel, C. Costs of Resistance in Plants: From Theory to Evidence. Annu. Plant Rev. Online 2017, 41, 263–307. [Google Scholar] [CrossRef]
- Herms, D.A.; Mattson, W.J. The Dilemma of Plants: To Grow or Defend. Q. Rev. Biol. 1992, 67, 283–335. [Google Scholar] [CrossRef]
- Raskin, I.; Ribnicky, D.M.; Komarnytsky, S.; Ilic, N.; Poulev, A.; Borisjuk, N.; Brinker, A.; Moreno-Fernández, D.; Ripoll, C.; Yakoby, N.; et al. Plants and human health in the twenty-first century. Trends Biotechnol. 2002, 20, 522–531. [Google Scholar] [CrossRef]
- Hall, R.D.; Brouwer, I.; Fitzgerald, M. Plant metabolomics and its potential application for human nutrition. Physiol. Plant. 2007, 132, 162–175. [Google Scholar] [CrossRef]
- Campos, M.; Yoshida, Y.; Major, I.; Ferreira, D.D.O.; Weraduwage, S.M.; Froehlich, J.E.; Johnson, B.F.; Kramer, D.; Jander, G.; Sharkey, T.; et al. Rewiring of jasmonate and phytochrome B signalling uncouples plant growth-defense tradeoffs. Nat. Commun. 2016, 7, 12570. [Google Scholar] [CrossRef]
- Quail, P.H.; Boylan, M.T.; Parks, B.M.; Short, T.W.; Xu, Y.; Wagner, D. Phytochromes: Photosensory perception and signal transduction. Science 1995, 268, 675–680. [Google Scholar] [CrossRef]
- Santner, A.; Estelle, M. The JAZ Proteins Link Jasmonate Perception with Transcriptional Changes. Plant Cell 2007, 19, 3839–3842. [Google Scholar] [CrossRef]
- Zhang, F.; Yao, J.; Ke, J.; Zhang, L.; Lam, V.Q.; Xin, X.-F.; Zhou, X.E.; Chen, J.; Brunzelle, J.; Griffin, P.R.; et al. Structural basis of JAZ repression of MYC transcription factors in jasmonate signalling. Nature 2015, 525, 269–273. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant response to jasmonates: Current developments and their role in changing environment. Bull. Natl. Res. Cent. 2019, 43, 153. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The Crosstalks Between Jasmonic Acid and Other Plant Hormone Signaling Highlight the Involvement of Jasmonic Acid as a Core Component in Plant Response to Biotic and Abiotic Stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-T.; Murthy, H.N.; Park, S.-Y. Methyl Jasmonate Induced Oxidative Stress and Accumulation of Secondary Metabolites in Plant Cell and Organ Cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.-L.; Okamoto, H. Molecular interaction of jasmonate and phytochrome a signalling. J. Exp. Bot. 2014, 65, 2847–2857. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Ballaré, C.L. Control of Plant Growth and Defense by Photoreceptors: From Mechanisms to Opportunities in Agriculture. Mol. Plant 2020, 14, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Whitelam, G.C.; Johnson, E.; Peng, J.; Carol, P.; Anderson, M.L.; Cowl, J.S.; Harberd, N.P. Phytochrome A null mutants of Arabidopsis display a wild-type phenotype in white light. Plant Cell 1993, 5, 757–768. [Google Scholar] [CrossRef]
- Fernández-Milmanda, G.L.; Crocco, C.D.; Reichelt, M.; Mazza, C.A.; Köllner, T.G.; Zhang, T.; Cargnel, M.D.; Lichy, M.Z.; Fiorucci, A.-S.; Fankhauser, C.; et al. A light-dependent molecular link between competition cues and defence responses in plants. Nat. Plants 2020, 6, 223–230. [Google Scholar] [CrossRef]
- Leone, M.; Keller, M.M.; Cerrudo, I.; Ballaré, C.L. To grow or defend? Low red: Far-red ratios reduce jasmonate sensitivity in Arabidopsis seedlings by promoting DELLA degradation and increasing JAZ10 stability. New Phytol. 2014, 204, 355–367. [Google Scholar] [CrossRef]
- Moreno, J.; Tao, Y.; Chory, J.; Ballare, C.L. Ecological modulation of plant defense via phytochrome control of jasmonate sensitivity. Proc. Natl. Acad. Sci. USA 2009, 106, 4935–4940. [Google Scholar] [CrossRef]
- Bell, D.L.; Galloway, L.F. Population differentiation for plasticity to light in an annual herb: Adaptation and cost. Am. J. Bot. 2008, 95, 59–65. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Wu, Y.; Gong, W.; Yang, W. Shade Inhibits Leaf Size by Controlling Cell Proliferation and Enlargement in Soybean. Sci. Rep. 2017, 7, 9259. [Google Scholar] [CrossRef]
- Mir, M.Y.; Hamid, S.; Kamili, A.N.; Hassan, Q.P. Sneak peek of Hypericum perforatum L.: Phytochemistry, phytochemical efficacy and biotechnological interventions. J. Plant Biochem. Biotechnol. 2019, 28, 357–373. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Strehmel, N.; Böttcher, C.; Schmidt, S.; Scheel, D. Profiling of secondary metabolites in root exudates of Arabidopsis thaliana. Phytochemistry 2014, 108, 35–46. [Google Scholar] [CrossRef]
- Carlisle, E.; Myers, S.S.; Raboy, V.; Bloom, A.J. The Effects of Inorganic Nitrogen form and CO2 Concentration on Wheat Yield and Nutrient Accumulation and Distribution. Front. Plant Sci. 2012, 3, 195. [Google Scholar] [CrossRef]
- Butler, W.L.; Norris, K.H.; Siegelman, H.W.; Hendricks, S.B. Detection, assay, and preliminary purification of the pigment controlling photoresponsive development of plants. Proc. Natl. Acad. Sci. USA 1959, 45, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Porzel, A.; Farag, M.A.; Mülbradt, J.; Wessjohann, L.A. Metabolite profiling and fingerprinting of Hypericum species: A comparison of MS and NMR metabolomics. Metabolomics 2013, 10, 574–588. [Google Scholar] [CrossRef]
- Tatsis, E.; Exarchou, V.; Troganis, A.N.; Gerothanassis, I.P. 1H NMR determination of hypericin and pseudohypericin in complex natural mixtures by the use of strongly deshielded OH groups. Anal. Chim. Acta 2008, 607, 219–226. [Google Scholar] [CrossRef]
- Tusevski, O.; Stanoeva, J.P.; Stefova, M.; Kungulovski, D.; Pancevska, N.; Sekulovski, N.; Panov, S.; Simic, S. Hairy roots of Hypericum perforatum L.: A promising system for xanthone production. Open Life Sci. 2013, 8, 1010–1022. [Google Scholar] [CrossRef]
- Salem, M.A.; Jüppner, J.; Bajdzienko, K.; Giavalisco, P. Protocol: A fast, comprehensive and reproducible one-step extraction method for the rapid preparation of polar and semi-polar metabolites, lipids, proteins, starch and cell wall polymers from a single sample. Plant Methods 2016, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Paponov, M.; Arakelyan, A.; Dobrev, P.; Verheul, M.; Paponov, I. Nitrogen Deficiency and Synergism between Continuous Light and Root Ammonium Supply Modulate Distinct but Overlapping Patterns of Phytohormone Composition in Xylem Sap of Tomato Plants. Plants 2021, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Paponov, M.; Kechasov, D.; Lacek, J.; Verheul, M.J.; Paponov, I. Supplemental Light-Emitting Diode Inter-Lighting Increases Tomato Fruit Growth Through Enhanced Photosynthetic Light Use Efficiency and Modulated Root Activity. Front. Plant Sci. 2020, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Whitelam, G.C. Phytochromes and Shade-avoidance Responses in Plants. Ann. Bot. 2005, 96, 169–175. [Google Scholar] [CrossRef]
- Havko, N.E.; Major, I.T.; Jewell, J.B.; Attaran, E.; Browse, J.; Howe, G.A. Control of Carbon Assimilation and Partitioning by Jasmonate: An Accounting of Growth–Defense Tradeoffs. Plants 2016, 5, 7. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Menzies, N.W.; Lombi, E.; Kopittke, P. Effects of methyl jasmonate on plant growth and leaf properties. J. Plant Nutr. Soil Sci. 2018, 181, 409–418. [Google Scholar] [CrossRef]
- Zhen, S.; Bugbee, B. Far-red photons have equivalent efficiency to traditional photosynthetic photons: Implications for redefining photosynthetically active radiation. Plant, Cell Environ. 2020, 43, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.; Beveridge, C.A. Roles for Auxin, Cytokinin, and Strigolactone in Regulating Shoot Branching. Plant Physiol. 2009, 149, 1929–1944. [Google Scholar] [CrossRef]
- Casal, J.J. Light quality effects on the appearance of tillers of different order in wheat (Triticum aestivum). Ann. Appl. Biol. 1988, 112, 167–173. [Google Scholar] [CrossRef]
- Hendry, G.A.; Grime, J.P. (Eds.) Methods in Comparative Plant Ecology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1993. [Google Scholar] [CrossRef]
- Poorter, H.; Van Berkel, Y.; Baxter, R.; Hertog, J.D.; Dijkstra, P.U.; Gifford, R.M.; Griffin, K.L.; Roumet, C.; Roy, J.C.; Wong, S.C. The effect of elevated CO2 on the chemical composition and construction costs of leaves of 27 C3 species. Plant, Cell Environ. 1997, 20, 472–482. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Elemoine, R.; Camera, S.E.; Eatanassova, R.; Edédaldéchamp, F.; Eallario, T.; Epourtau, N.; Ebonnemain, J.-L.; Elaloi, M.; Ecoutos-Thévenot, P.; Emaurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef]
- Cometti, N.N.; Martins, M.Q.; Bremenkamp, C.A.; Nunes, J.A. Nitrate concentration in lettuce leaves depending on photosynthetic photon flux and nitrate concentration in the nutrient solution. Hortic. Bras. 2011, 29, 548–553. [Google Scholar] [CrossRef][Green Version]
- Antioxidant activity and total phenolic compound content of certain medicinal plants. Int. J. Biosci. 2014, 5, 213–222. [CrossRef]
- Jung, N.C.; Tamai, Y. Anatomical observation of polyphenol changes in epidermal cells during the development of Quercus acutissima–Scleroderma verrucosum ectomycorrhizae. Trees 2011, 26, 301–310. [Google Scholar] [CrossRef]
- Chaiwanon, J.; Wang, W.; Zhu, J.-Y.; Oh, E.; Wang, Z.-Y. Information Integration and Communication in Plant Growth Regulation. Cell 2016, 164, 1257–1268. [Google Scholar] [CrossRef]
- Chini, A.; Gimenez-Ibanez, S.; Goossens, A.; Solano, R. Redundancy and specificity in jasmonate signalling. Curr. Opin. Plant Biol. 2016, 33, 147–156. [Google Scholar] [CrossRef]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.H.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCFCO11 complex during jasmonate signalling. Nature 2007, 448, 661–662. [Google Scholar] [CrossRef]
- Gimenez-Ibanez, S.; Boter, M.; Ortigosa, A.; García-Casado, G.; Chini, A.; Lewsey, M.G.; Ecker, J.; Ntoukakis, V.; Solano, R. JAZ 2 controls stomata dynamics during bacterial invasion. New Phytol. 2016, 213, 1378–1392. [Google Scholar] [CrossRef]
- Li, R.; Wang, M.; Wang, Y.; Schuman, M.C.; Weinhold, A.; Schäfer, M.; Jimenez-Aleman, G.; Barthel, A.; Baldwin, I.T. Flower-specific jasmonate signaling regulates constitutive floral defenses in wild tobacco. Proc. Natl. Acad. Sci. USA 2017, 114, E7205–E7214. [Google Scholar] [CrossRef]
- Takaoka, Y.; Iwahashi, M.; Chini, A.; Saito, H.; Ishimaru, Y.; Egoshi, S.; Kato, N.; Tanaka, M.; Bashir, K.; Seki, M.; et al. A rationally designed JAZ subtype-selective agonist of jasmonate perception. Nat. Commun. 2018, 9, 3654. [Google Scholar] [CrossRef]
- Jimenez-Aleman, G.H.; Machado, R.A.R.; Baldwin, I.; Boland, W. JA-Ile-macrolactones uncouple growth and defense in wild tobacco. Org. Biomol. Chem. 2017, 15, 3391–3395. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Baldwin, I.T.; Erb, M. Herbivory-induced jasmonates constrain plant sugar accumulation and growth by antagonizing gibberellin signaling and not by promoting secondary metabolite production. New Phytol. 2017, 215, 803–812. [Google Scholar] [CrossRef]
- Heinrich, M.; Hettenhausen, C.; Lange, T.; Wünsche, H.; Fang, J.; Baldwin, I.; Wu, J. High levels of jasmonic acid antagonize the biosynthesis of gibberellins and inhibit the growth ofNicotiana attenuatastems. Plant J. 2012, 73, 591–606. [Google Scholar] [CrossRef]
- Hisamatsu, T.; King, R.W.; Helliwell, C.; Koshioka, M. The Involvement of Gibberellin 20-Oxidase Genes in Phytochrome-Regulated Petiole Elongation of Arabidopsis. Plant Physiol. 2005, 138, 1106–1116. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.-L.; Yao, J.; Mei, C.-S.; Tong, X.-H.; Zeng, L.-J.; Li, Q.; Xiao, L.-T.; Sun, T.-P.; Li, J.; Deng, X.-W.; et al. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc. Natl. Acad. Sci. USA 2012, 109, E1192–E1200. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Shim, J.S.; Jung, C.; Song, J.T.; Lee, H.Y.; Chung, P.J.; Kim, J.-K.; Choi, Y.D. Volatile methyl jasmonate is a transmissible form of jasmonate and its biosynthesis is involved in systemic jasmonate response in wounding. Plant Biotechnol. Rep. 2014, 8, 409–419. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernandez, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; Garciacasado, G.; Lopezvidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Kramell, R.; Miersch, O.; Atzorn, R.; Parthier, B.; Wasternack, C. Octadecanoid-Derived Alteration of Gene Expression and the “Oxylipin Signature” in Stressed Barley Leaves. Implications for Different Signaling Pathways. Plant Physiol. 2000, 123, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Staswick, P.E.; Tiryaki, I. The Oxylipin Signal Jasmonic Acid Is Activated by an Enzyme That Conjugates It to Isoleucine in Arabidopsis. Plant Cell 2004, 16, 2117–2127. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Lee, L.Y.C.; Xia, K.; Yan, Y.; Yu, H. DELLAs Modulate Jasmonate Signaling via Competitive Binding to JAZs. Dev. Cell 2010, 19, 884–894. [Google Scholar] [CrossRef]
- Djakovic-Petrovic, T.; De Wit, M.; Voesenek, L.; Pierik, R. DELLA protein function in growth responses to canopy signals. Plant J. 2007, 51, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Gidda, S.K.; Miersch, O.; Levitin, A.; Schmidt, J.; Wasternack, C.; Varin, L. Biochemical and Molecular Characterization of a Hydroxyjasmonate Sulfotransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 17895–17900. [Google Scholar] [CrossRef] [PubMed]
- Chico, J.-M.; Fernández-Barbero, G.; Chini, A.; Calvo, P.F.; Diez-Diaz, M.; Solano, R. Repression of Jasmonate-Dependent Defenses by Shade Involves Differential Regulation of Protein Stability of MYC Transcription Factors and Their JAZ Repressors in Arabidopsis. Plant Cell 2014, 26, 1967–1980. [Google Scholar] [CrossRef]
- Gundel, P.E.; Pierik, R.; Mommer, L.; Ballare, C. Competing neighbors: Light perception and root function. Oecologia 2014, 176, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jung, S. Effect of chlorophyll reduction in Arabidopsis thaliana by methyl jasmonate or norflurazon on antioxidant systems. Plant Physiol. Biochem. 2004, 42, 225–231. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Sonobe, R.; Yamashita, H.; Mihara, H.; Morita, A.; Ikka, T. Estimation of Leaf Chlorophyll a, b and Carotenoid Contents and Their Ratios Using Hyperspectral Reflectance. Remote Sens. 2020, 12, 3265. [Google Scholar] [CrossRef]
- Kuai, B.; Chen, J.; Hörtensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. J. Exp. Bot. 2017, 69, 751–767. [Google Scholar] [CrossRef]
- Qiu, X.; Xu, Y.; Xiong, B.; Dai, L.; Huang, S.; Dong, T.; Sun, G.; Liao, L.; Deng, Q.; Wang, X.; et al. Effects of exogenous methyl jasmonate on the synthesis of endogenous jasmonates and the regulation of photosynthesis in citrus. Physiol. Plant. 2020, 170, 398–414. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Li, Z.; Gu, W.R.; Li, J.; Xie, T.L.; Wei, S. Methyl jasmonate pretreatment promotes the growth and photosynthesis of maize seedlings under saline conditions by enhancing the antioxidant defense system. Int. J. Agric. Biol. 2018, 20, 1454–1462. [Google Scholar]
- Buckland, S.M.; Price, A.H.; Hendry, G.A.F. The role of ascorbate in drought-treated Cochlearia atlantica Pobed. and Armeria maritima (Mill.) Willd. New Phytol. 1991, 119, 155–160. [Google Scholar] [CrossRef]
- McLaren, J.S.; Smith, H. Phytochrome control of the growth and development of Rumex obtusifolius under simulated canopy light environments*. Plant, Cell Environ. 1978, 1, 61–67. [Google Scholar] [CrossRef]
- Brouwer, B.; Ziolkowska, A.; Bagard, M.; Keech, O.; Gardeström, P.; Brouwer, B.; Ziolkowska, A.; Bagard, M.; Keech, O.; Gardeström, P. The impact of light intensity on shade-induced leaf senescence. Plant, Cell Environ. 2012, 35, 1084–1098. [Google Scholar] [CrossRef] [PubMed]
- Wientjes, E.; Philippi, J.; Borst, J.W.; van Amerongen, H. Imaging the Photosystem I/Photosystem II chlorophyll ratio inside the leaf. Biochim. et Biophys. Acta (BBA) Bioenerg. 2017, 1858, 259–265. [Google Scholar] [CrossRef]
- Caffarri, S.; Tibiletti, T.; Jennings, R.C.; Santabarbara, S. A comparison between plant photosystem I and photosystem II architecture and functioning. Curr. Protein Pept. Sci. 2014, 15, 296–331. [Google Scholar] [CrossRef]
- Katz, L.; Donadio, S. Polyketide Synthesis: Prospects for Hybrid Antibiotics. Annu. Rev. Microbiol. 1993, 47, 875–912. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen, F.; Goto, E.; Kozai, T. Plant–Environment Interactions: Accumulation of Hypericin in Dark Glands of Hypericum perforatum. Ann. Bot. 2006, 98, 793–804. [Google Scholar] [CrossRef]
- Walker, T. Jasmonic acid-induced hypericin production in cell suspension cultures of Hypericum perforatum L. (St. John’s wort). Phytochemistry 2002, 60, 289–293. [Google Scholar] [CrossRef]
- Murthy, H.N.; Kim, Y.-S.; Park, S.-Y.; Paek, K.-Y. Hypericins: Biotechnological production from cell and organ cultures. Appl. Microbiol. Biotechnol. 2014, 98, 9187–9198. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-Q.; Yu, X.-K.; Lian, M.-L.; Park, S.-Y.; Piao, X.-C. Several factors affecting hypericin production of Hypericum perforatum during adventitious root culture in airlift bioreactors. Acta Physiol. Plant. 2014, 36, 975–981. [Google Scholar] [CrossRef]
- Mosaleeyanon, K.; Zobayed, S.; Afreen, F.; Kozai, T. Relationships between net photosynthetic rate and secondary metabolite contents in St. John’s wort. Plant Sci. 2005, 169, 523–531. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Ghasemzadeh, N. Flavonoids and phenolic acids: Role and biochemical activity in plants and human. J. Med. Plants Res. 2011, 5, 6697–6703. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Sa, R.; Feng, K.; Zhao, M.; Yu, J.; Ji, Y.; Hou, M.; et al. Effect of Methyl Jasmonate on Phenolic Accumulation in Wounded Broccoli. Molecules 2019, 24, 3537. [Google Scholar] [CrossRef] [PubMed]
- Demkura, P.V.; Abdala, G.; Baldwin, I.; Ballareݩ, C.L. Jasmonate-Dependent and -Independent Pathways Mediate Specific Effects of Solar Ultraviolet B Radiation on Leaf Phenolics and Antiherbivore Defense. Plant Physiol. 2009, 152, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Kausar, H.; Ambrin, G.; Okla, M.K.; Soufan, W.; Al-Ghamdi, A.A.; Ahmad, A. Metabolic Flux Analysis of Catechin Biosynthesis Pathways Using Nanosensor. Antioxidants 2020, 9, 288. [Google Scholar] [CrossRef]
- Chemler, J.A.; Lock, L.T.; Koffas, M.A.G.; Tzanakakis, E.S. Standardized biosynthesis of flavan-3-ols with effects on pancreatic beta-cell insulin secretion. Appl. Microbiol. Biotechnol. 2007, 77, 797–807. [Google Scholar] [CrossRef]
- Chemler, J.A.; Fowler, Z.L.; McHugh, K.P.; Koffas, M.A. Improving NADPH availability for natural product biosynthesis in Escherichia coli by metabolic engineering. Metab. Eng. 2010, 12, 96–104. [Google Scholar] [CrossRef]
- Nigutová, K.; Kusari, S.; Sezgin, S.; Petijová, L.; Henzelyová, J.; Bálintová, M.; Spiteller, M.; Cellarova, E. Chemometric evaluation of hypericin and related phytochemicals in 17 in vitro cultured Hypericum species, hairy root cultures and hairy root-derived transgenic plants. J. Pharm. Pharmacol. 2017, 71, 46–57. [Google Scholar] [CrossRef]
- Bensaddek, L.; Villarreal, M.L.; Fliniaux, M.A. Induction and growth of hairy roots for the production of medicinal compounds. Electr. J. Integr. Biosci. 2008, 3, 2–9. [Google Scholar]
- Tusevski, O.; Krstikj, M.; Stanoeva, J.; Stefova, M.; Simic, S.G. Phenolic profile and biological activity of Hypericum perforatum L.: Can roots be considered as a new source of natural compounds? S. Afr. J. Bot. 2018, 117, 301–310. [Google Scholar] [CrossRef]
- Montazeri, M.; Azadbakht, M.; Zarrini, H.; Nematzadeh, G.; Pakdin-Parizi, A.; Davoodi, A. Hairy root cultures of Hyper–icum perforatum L.; a promising method for the high scale production of hypericin. J. Med. Plants Res. 2018, 17, 55–67. [Google Scholar]
- Badri, D.V.; Loyola-Vargas, V.M.; Du, J.; Stermitz, F.R.; Broeckling, C.; Iglesias, L.; Vivanco, J.M. Transcriptome analysis of Arabidopsis roots treated with signaling compounds: A focus on signal transduction, metabolic regulation and secretion. New Phytol. 2008, 179, 209–223. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
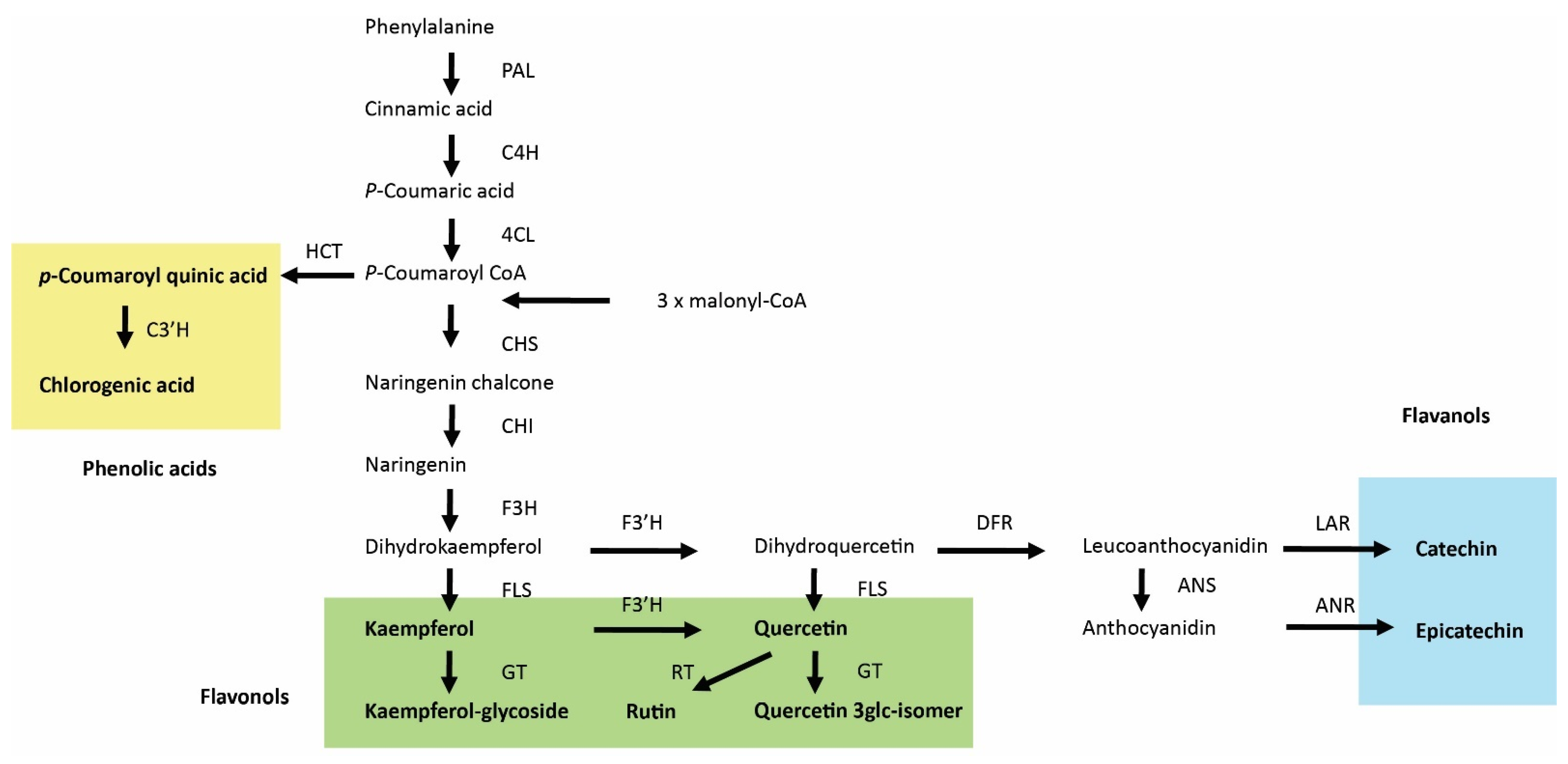{kind=link}
| Control | MeJA | FR | MeJA + FR | MeJA | FR | MeJA × FR | |
|---|---|---|---|---|---|---|---|
| Leaves (mg g−1 DW) | |||||||
| Naphthodi-Anthrones | |||||||
| Hypericin | 0.62 ± 0.07 | 1.00 ± 0.03 | 0.80 ± 0.05 | 1.08 ± 0.05 | *** | * | NS |
| Pseudohypericin | 0.19 ± 0.012 | 0.21 ± 0.016 | 0.17 ± 0.016 | 0.18 ± 0.009 | NS | NS | NS |
| Flavan-3-ols | |||||||
| Epicatechin | 5.30 ± 0.1 | 7.12 ± 0.22 | 5.23 ± 0.13 | 7.44 ± 0.33 | *** | NS | NS |
| Catechin | 26.8 ± 0.47 | 28.5 ± 0.32 | 24.9 ± 0.44 | 26.4 ± 0.21 | ** | *** | NS |
| Procyanidin dimer | 5.06 ± 0.1 | 7.67 ± 0.24 | 5.15 ± 0.27 | 7.33 ± 0.38 | *** | NS | NS |
| Phenolic Acids | |||||||
| Chlorogenic | 21.3 ± 1.3 | 21.6 ± 1.7 | 21.7 ± 1.0 | 20.4 ± 1.0 | NS | NS | NS |
| Coumaroylquinic | 15.1 ± 2.3 | 21.0 ± 0.85 | 18.5 ± 2.2 | 24.8 ± 0.6 | ** | NS | NS |
| Flavonols | |||||||
| Kaempferol-glycoside | 8.3 ± 1.47 | 4.8 ± 0.49 | 11.4 ± 1.84 | 9.8 ± 0.36 | NS | ** | NS |
| Rutin | 10.3 ± 1.7 | 6.8 ± 0.37 | 15.7 ± 1.96 | 12.7 ± 1.03 | * | *** | NS |
| Que-der 1 | 6.48 ± 0.4 | 4.16 ± 0.18 | 11.59 ± 0.74 | 8.70 ± 0.3 | *** | *** | NS |
| Que3glc | 3.6 ± 0.41 | 2.1 ± 0.26 | 8.1 ± 0.65 | 5.5 ± 0.28 | *** | *** | NS |
| Quercetin | 3.58 ± 0.62 | 2.82 ± 0.15 | 4.51 ± 0.77 | 3.29 ± 0.25 | NS | NS | NS |
| Que-der 2 | 6.42 ± 0.87 | 6.44 ± 0.56 | 7.35 ± 0.57 | 7.95 ± 0.49 | NS | NS | NS |
| Roots (mg g−1 DW) | |||||||
| Hyperfirin | 0.32 ± 0.09 | 0.62 ± 0.22 | 0.27 ± 0.10 | 0.57 ± 0.06 | *** | NS | NS |
| Other APGs | 1.05 ± 0.07 | 2.05 ± 0.28 | 1.11 ± 0.10 | 1.93 ± 0.18 | *** | NS | NS |
| Catechin | 4.57 ± 0.4 | 2.63 ± 0.54 | 4.76 ± 0.54 | 3.89 ± 0.17 | * | NS | NS |
| Root exudates (ng g−1 FW(root) h−1) | |||||||
| Hypericin | 21.6 ± 2.7 | 40.6 ± 3.9 | 17.1 ± 2.8 | 24.7 ± 2.8 | *** | ** | NS |
| Other NDA | 19.5 ± 5.2 | 43.0 ± 8.1 | 18.9 ± 5.3 | 31.7 ± 14.1 | * | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paponov, M.; Antonyan, M.; Slimestad, R.; Paponov, I.A. Decoupling of Plant Growth and Accumulation of Biologically Active Compounds in Leaves, Roots, and Root Exudates of Hypericum perforatum L. by the Combination of Jasmonate and Far-Red Lighting. Biomolecules 2021, 11, 1283. https://doi.org/10.3390/biom11091283
Paponov M, Antonyan M, Slimestad R, Paponov IA. Decoupling of Plant Growth and Accumulation of Biologically Active Compounds in Leaves, Roots, and Root Exudates of Hypericum perforatum L. by the Combination of Jasmonate and Far-Red Lighting. Biomolecules. 2021; 11(9):1283. https://doi.org/10.3390/biom11091283
Chicago/Turabian StylePaponov, Martina, Manya Antonyan, Rune Slimestad, and Ivan A. Paponov. 2021. "Decoupling of Plant Growth and Accumulation of Biologically Active Compounds in Leaves, Roots, and Root Exudates of Hypericum perforatum L. by the Combination of Jasmonate and Far-Red Lighting" Biomolecules 11, no. 9: 1283. https://doi.org/10.3390/biom11091283
APA StylePaponov, M., Antonyan, M., Slimestad, R., & Paponov, I. A. (2021). Decoupling of Plant Growth and Accumulation of Biologically Active Compounds in Leaves, Roots, and Root Exudates of Hypericum perforatum L. by the Combination of Jasmonate and Far-Red Lighting. Biomolecules, 11(9), 1283. https://doi.org/10.3390/biom11091283