Regulated Iron Siderophore Production of the Halophilic Archaeon Haloferax volcanii


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
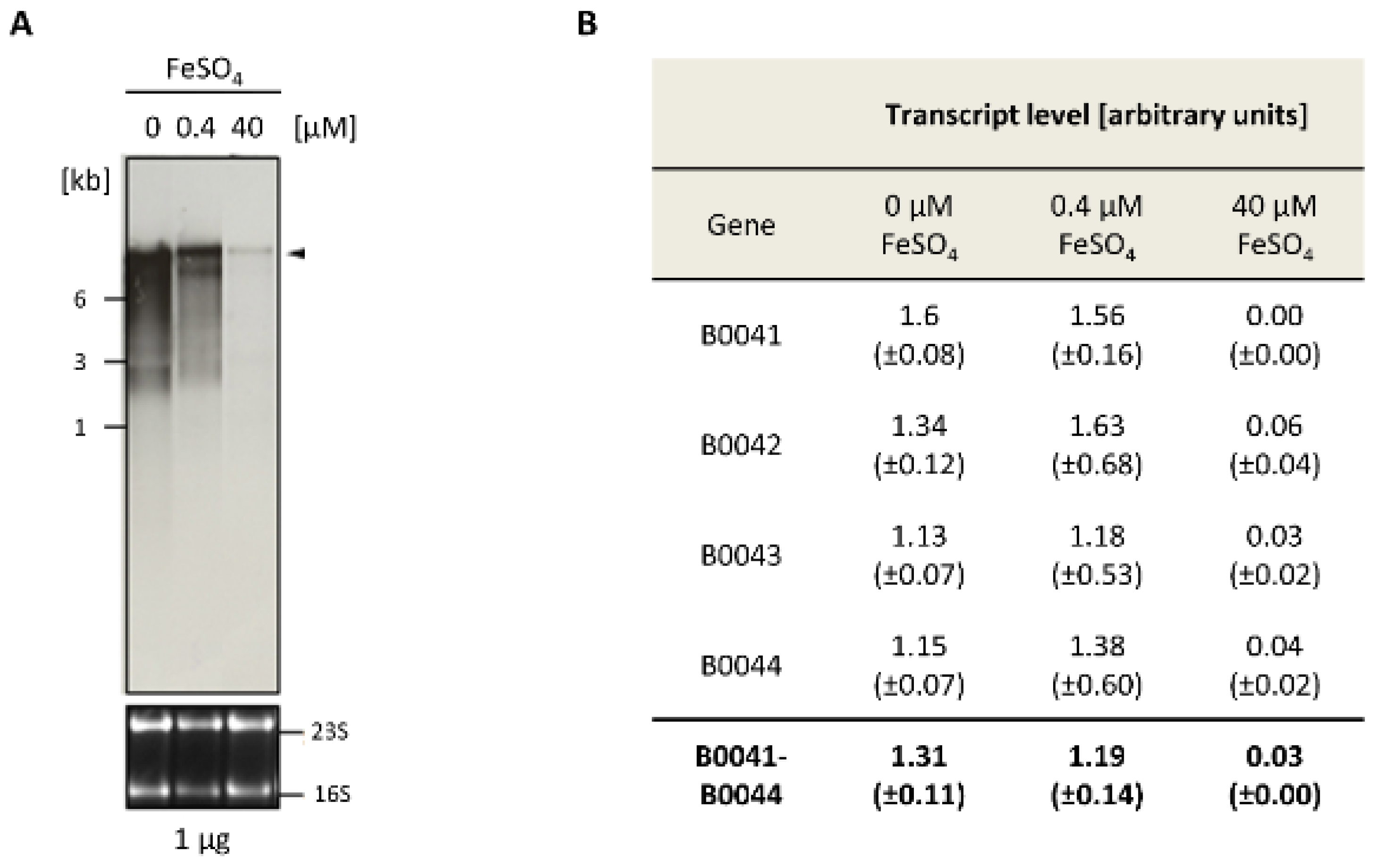
2.2. Transcript Analysis
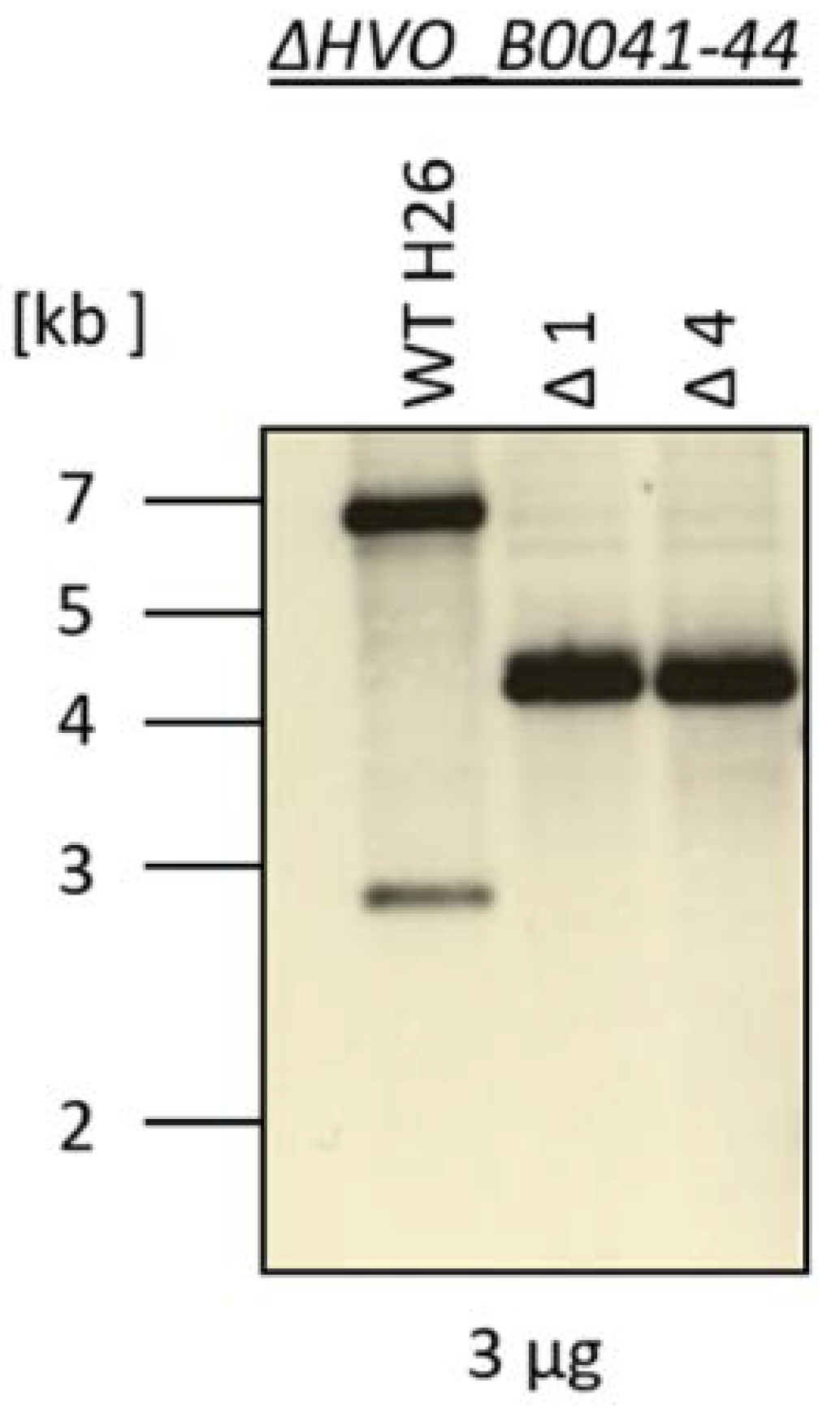
2.3. Construction of a Deletion Strain
2.4. Analysis of Fe-Dependent Growth
2.5. Siderophore Assays
- (1)
- FeCl3 assay [16]. Cultures were grown in liquid synthetic medium with extra pure NaCl and 0.5% (w/v) glucose. Precultures were grown in the absence of FeCl3 overnight. Test cultures were grown in the presence of 20 µM FeCl3. After five days of incubation, cells were pelleted and the supernatant was used for the assay. Then, 0.5 mL of supernatant was added to 2.5 mL of FeCl3 solution (2% (w/v)). Generation of a brown color was indicative for the presence of a siderophore.
- (2)
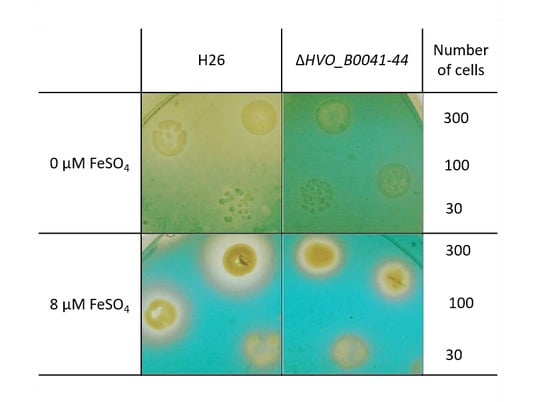
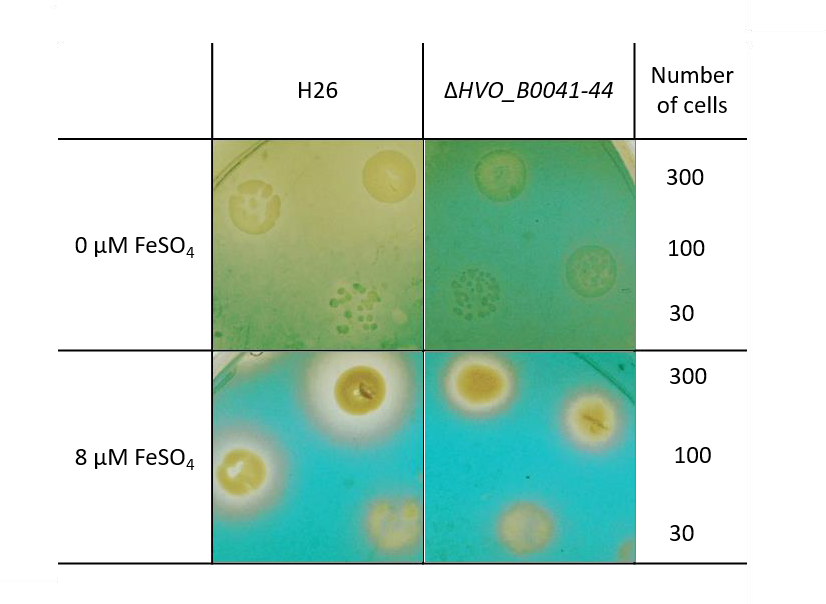
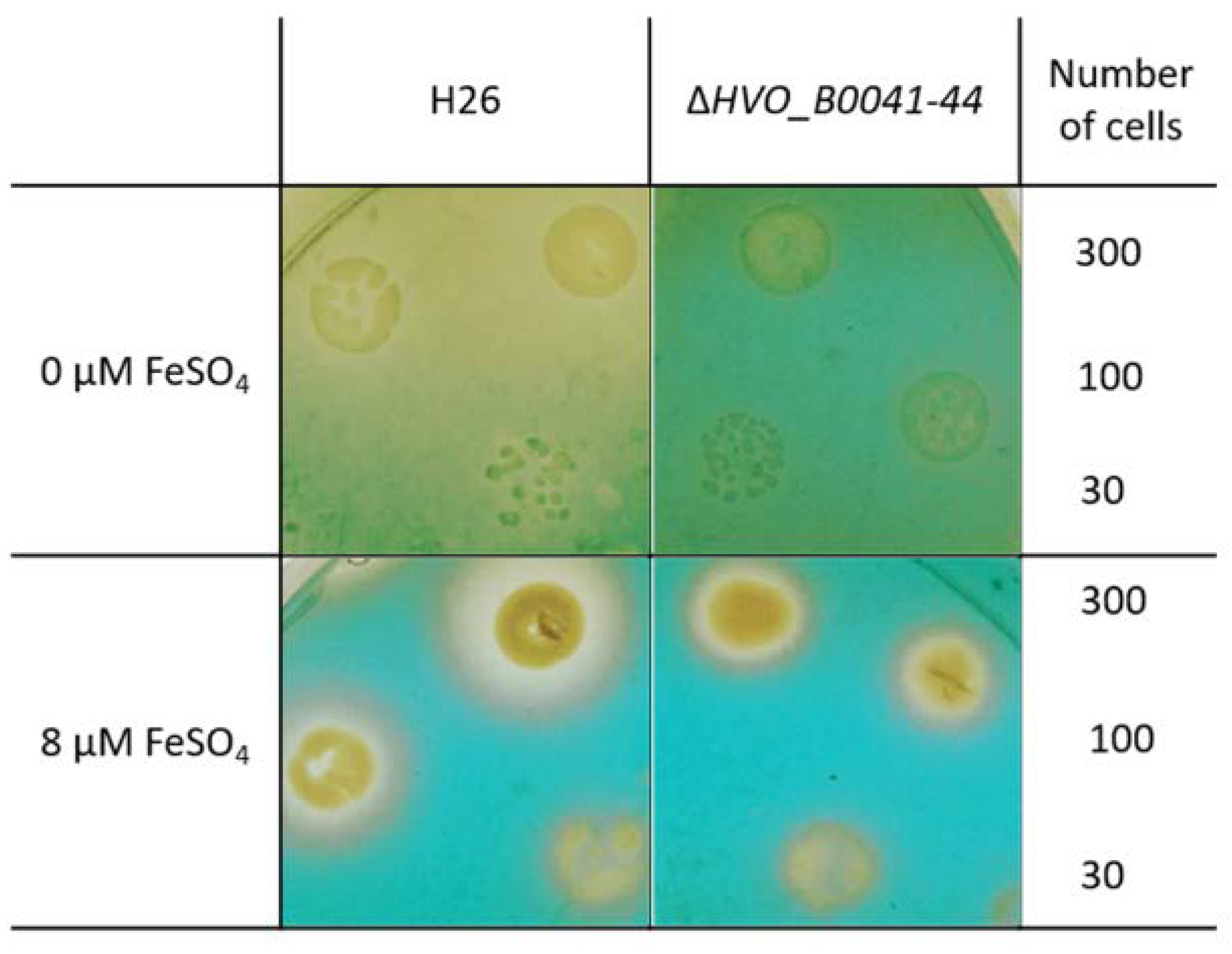
- The O-CAS assay (overlay chromeazurol S assay) [17] was developed to enable the analyses of siderophore production for species that are sensitive to the CAS reagent. It consists of a prior growth phase and a subsequent analysis with the CAS reagent. Precultures were grown in the absence of FeCl3 in liquid synthetic medium with extra pure NaCl and 0.5% (w/v) glucose. Cells numbers were counted using a Neubauer counting chamber, and suitable dilutions were generated to yield the cell numbers indicated in Section 3. Next, 5 µl of the dilutions were spotted on solid medium (extra pure NaCl, 0.5% glucose (w/v), 1% (w/v) agar) containing the indicated FeCl3 concentrations. The agar plates were incubated for four days at 42 °C to allow the formation of small colonies. The plates were overlaid with 1× CAS solution with 0.9% (w/v) agar and incubated at room temperature (10× CAS reagent: 0.1 mM FeCl3, 1 mM HCl, 2 mM HDTMA (Sigma-Aldrich, Taufkirchen, Germany; H52366; hexadecyltrimethylammonium bromide), 1 mM CAS = chromeazurol S, Sigma 72687). In the absence of a siderophore, a blue-green color develops in the plate. The presence of uncolored halos around colonies is indicative for the production and export of a FeCl3 siderophore.
2.6. Databases and Bioinformatic Analyses
3. Results
3.1. A Gene Cluster Possibly Involved in Siderophore Biosynthesis
3.2. Transcript Analyses and Differential Expression
3.3. Generation of a Deletion Mutant
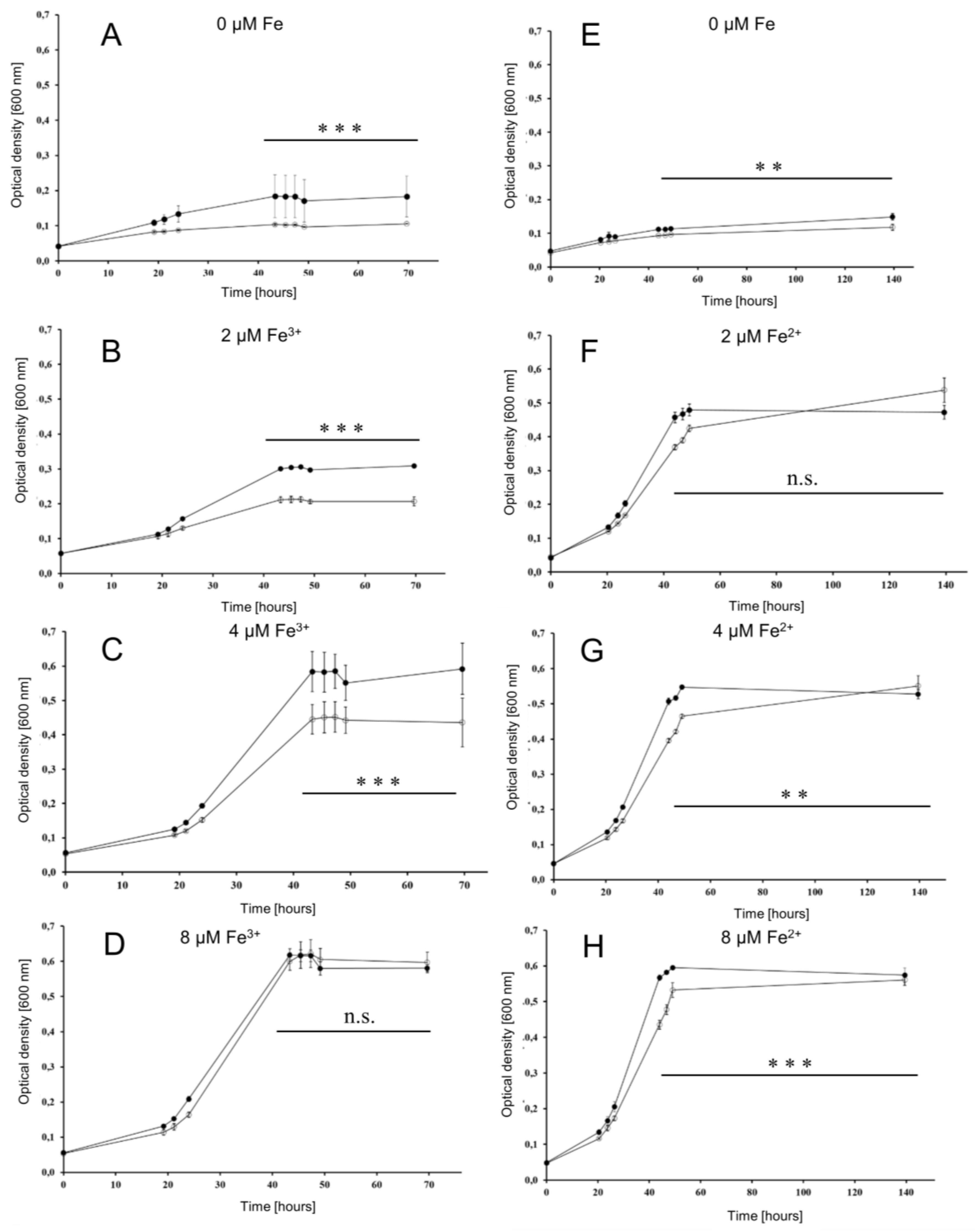
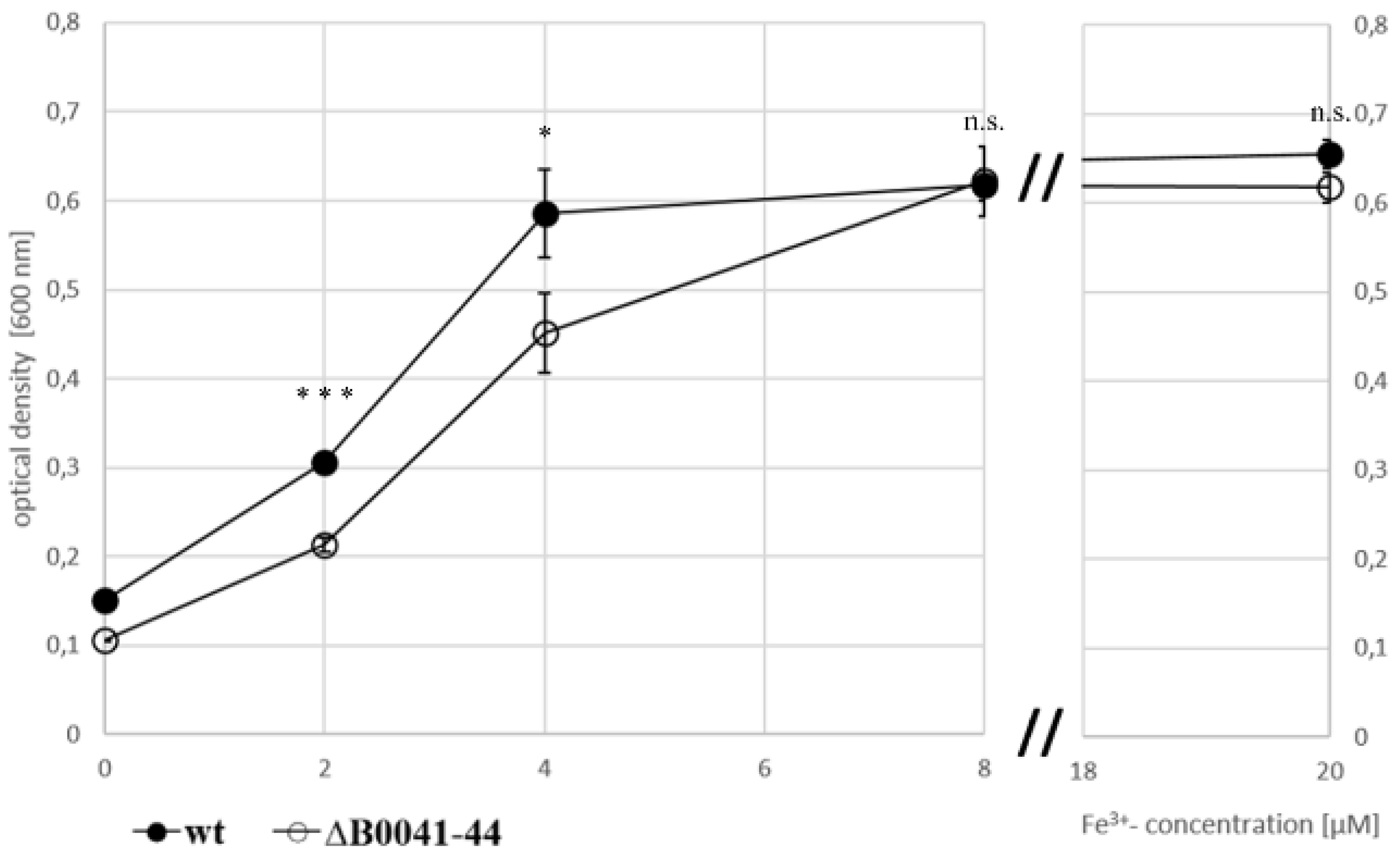
3.4. Fe-Dependent Growth of Wild Type and Deletion Mutant
3.5. Verification of Siderophore Synthesis and Export
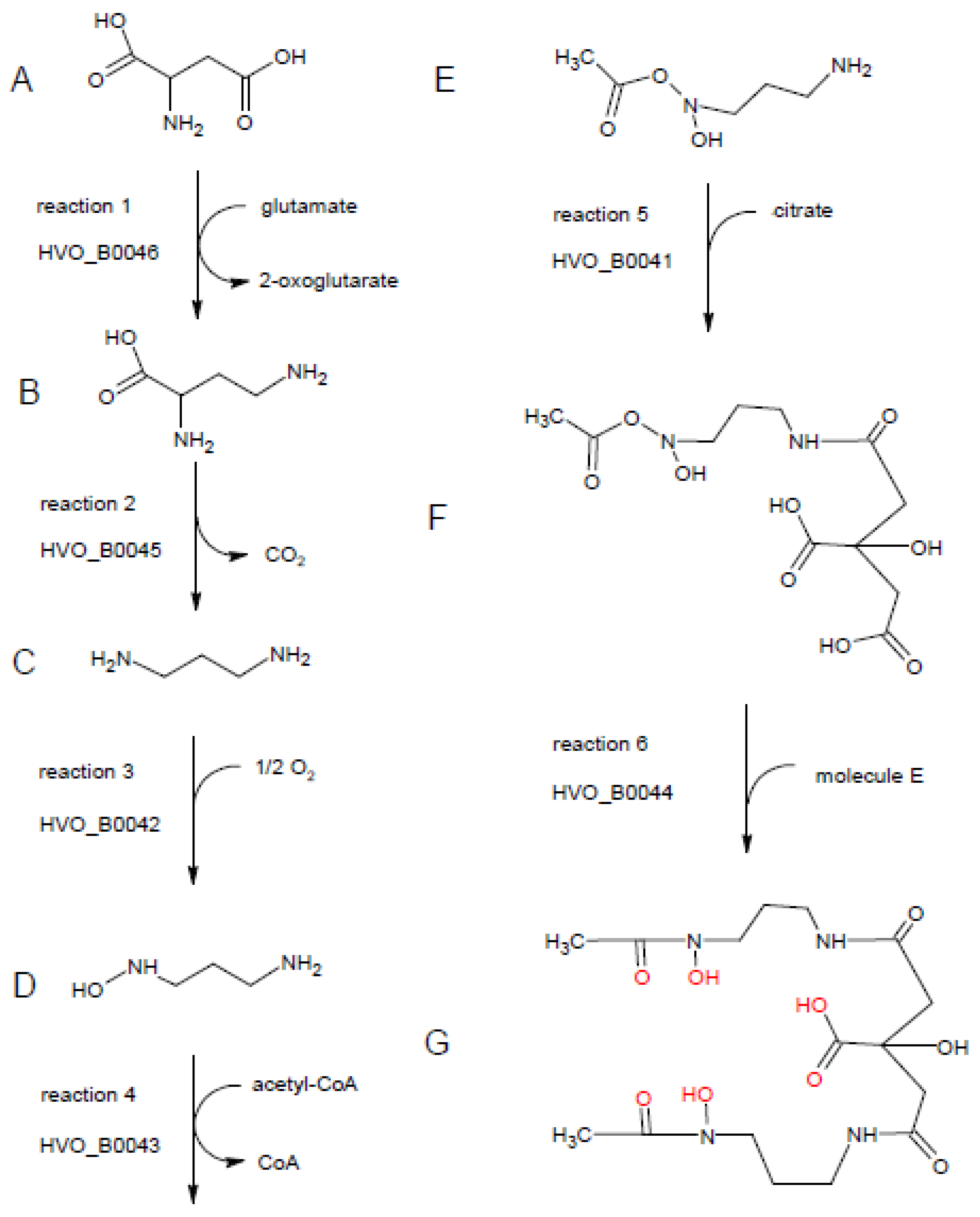
3.6. Bioinformatic Analysis of the Six Iuc Operon-Encoded Proteins
3.7. Phylogenetic Distribution of Iuc and Related Genes, Putative Enzyme Activities, and Proposal of a Siderophore Biosynthesis Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kramer, J.; Özkaya, Ö.; Kümmerli, R. Bacterial siderophores in community and host interactions. Nat. Rev. Microbiol. 2020, 18, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Carver, P.L. The Battle for Iron between Humans and Microbes. Curr. Med. Chem. 2018, 25, 85–96. [Google Scholar] [CrossRef]
- Barry, S.M.; Challis, G.L. Recent advances in siderophore biosynthesis. Curr. Opin. Chem. Biol. 2009, 13, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Årstøl, E.; Hohmann-Marriott, M.F. Cyanobacterial Siderophores-Physiology, Structure, Biosynthesis, and Applications. Mar. Drugs 2019, 17, 281. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Marahiel, M.A. Siderophore-based iron acquisition and pathogen control. Microbiol. Mol. Biol. Rev. 2007, 71, 413–451. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, R.T.; Snow, J.T.; Zhang, Q.; Rickaby, R.E.M. Iron requirements and uptake strategies of the globally abundant marine ammonia-oxidising archaeon, Nitrosopumilus maritimus SCM1. ISME J. 2019, 13, 2295–2305. [Google Scholar] [CrossRef]
- Dave, B.P.; Anshuman, K.; Hajela, P. Siderophores of halophilic archaea and their chemical characterization. Indian J. Exp. Biol. 2006, 44, 340–344. [Google Scholar]
- Hubmacher, D.; Matzanke, B.F.; Anemüller, S. Iron-uptake in the Euryarchaeon Halobacterium salinarum. Biometals 2007, 20, 539–547. [Google Scholar] [CrossRef]
- Hartman, A.L.; Norais, C.; Badger, J.H.; Delmas, S.; Haldenby, S.; Madupu, R.; Robinson, J.; Khouri, H.; Ren, Q.; Lowe, T.M.; et al. The complete genome sequence of Haloferax volcanii DS2, a model archaeon. PLoS ONE 2010, 5, e9605. [Google Scholar] [CrossRef]
- Allers, T.; Ngo, H.-P.; Mevarech, M.; Lloyd, R.G. Development of additional selectable markers for the halophilic archaeon Haloferax volcanii based on the leuB and trpA genes. Appl. Environ. Microbiol. 2004, 70, 943–953. [Google Scholar] [CrossRef]
- Babski, J.; Haas, K.A.; Näther-Schindler, D.; Pfeiffer, F.; Förstner, K.U.; Hammelmann, M.; Hilker, R.; Becker, A.; Sharma, C.M.; Marchfelder, A.; et al. Genome-wide identification of transcriptional start sites in the haloarchaeon Haloferax volcanii based on differential RNA-Seq (dRNA-Seq). BMC Genom. 2016, 17, 629. [Google Scholar] [CrossRef] [PubMed]
- Laass, S.; Monzon, V.A.; Kliemt, J.; Hammelmann, M.; Pfeiffer, F.; Förstner, K.U.; Soppa, J. Characterization of the transcriptome of Haloferax volcanii, grown under four different conditions, with mixed RNA-Seq. PLoS ONE 2019, 14, e0215986. [Google Scholar] [CrossRef] [PubMed]
- Nicol, J.W.; Helt, G.A.; Blanchard, S.G.; Raja, A.; Loraine, A.E. The Integrated Genome Browser: Free software for distribution and exploration of genome-scale datasets. Bioinformatics 2009, 25, 2730–2731. [Google Scholar] [CrossRef] [PubMed]
- Hammelmann, M.; Soppa, J. Optimized generation of vectors for the construction of Haloferax volcanii deletion mutants. J. Microbiol. Methods 2008, 75, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Jantzer, K.; Zerulla, K.; Soppa, J. Phenotyping in the archaea: Optimization of growth parameters and analysis of mutants of Haloferax volcanii. FEMS Microbiol. Lett. 2011, 322, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Atkin, C.L.; Neilands, J.B.; Phaff, H.J. Rhodotorulic acid from species of Leucosporidium, Rhodosporidium, Rhodotorula, Sporidiobolus, and Sporobolomyces, and a new alanine-containing ferrichrome from Cryptococcus melibiosum. J. Bacteriol. 1970, 103, 722–733. [Google Scholar] [CrossRef]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.S.; Fernández, F.J. O-CAS, a fast and universal method for siderophore detection. J. Microbiol. Methods 2007, 70, 127–131. [Google Scholar] [CrossRef]
- Pfeiffer, F.; Broicher, A.; Gillich, T.; Klee, K.; Mejía, J.; Rampp, M.; Oesterhelt, D. Genome information management and integrated data analysis with HaloLex. Arch. Microbiol. 2008, 190, 281–299. [Google Scholar] [CrossRef]
- Huber, M.; Faure, G.; Laass, S.; Kolbe, E.; Seitz, K.; Wehrheim, C.; Wolf, Y.I.; Koonin, E.V.; Soppa, J. Translational coupling via termination-reinitiation in archaea and bacteria. Nat. Commun. 2019, 10, 4006. [Google Scholar] [CrossRef]
- Ludt, K.; Soppa, J. Polyploidy in halophilic archaea: Regulation, evolutionary advantages, and gene conversion. Biochem. Soc. Trans. 2019, 47, 933–944. [Google Scholar] [CrossRef]
- Breuert, S.; Allers, T.; Spohn, G.; Soppa, J. Regulated polyploidy in halophilic archaea. PLoS ONE 2006, 1, e92. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, F.; Oesterhelt, D. A manual curation strategy to improve genome annotation: Application to a set of haloarchael genomes. Life (Basel) 2015, 5, 1427–1444. [Google Scholar] [CrossRef] [PubMed]
- Date, S.V. Estimating protein function using protein-protein relationships. Methods Mol. Biol. 2007, 408, 109–127. [Google Scholar] [CrossRef]
- Lankford, C.E.; Walker, J.R.; Reeves, J.B.; Nabbut, N.H.; Byers, B.R.; Jones, R.J. Inoculum-dependent division lag of Bacillus cultures and its relation to an endogenous factor(s) (“schizokinen”). J. Bacteriol. 1966, 91, 1070–1079. [Google Scholar] [CrossRef]
- Mullis, K.B.; Pollack, J.R.; Neilands, J.B. Structure of schizokinen, an iron-transport compound from Bacillus megaterium. Biochemistry 1971, 10, 4894–4898. [Google Scholar] [CrossRef]
- Maier, L.-K.; Benz, J.; Fischer, S.; Alstetter, M.; Jaschinski, K.; Hilker, R.; Becker, A.; Allers, T.; Soppa, J.; Marchfelder, A. Deletion of the Sm1 encoding motif in the lsm gene results in distinct changes in the transcriptome and enhanced swarming activity of Haloferax cells. Biochimie 2015, 117, 129–137. [Google Scholar] [CrossRef][Green Version]
- Koide, T.; Reiss, D.J.; Bare, J.C.; Pang, W.L.; Facciotti, M.T.; Schmid, A.K.; Pan, M.; Marzolf, B.; Van, P.T.; Lo, F.-Y.; et al. Prevalence of transcription promoters within archaeal operons and coding sequences. Mol. Syst. Biol. 2009, 5, 285. [Google Scholar] [CrossRef]
- Shieh, Y.-W.; Minguez, P.; Bork, P.; Auburger, J.J.; Guilbride, D.L.; Kramer, G.; Bukau, B. Operon structure and cotranslational subunit association direct protein assembly in bacteria. Science 2015, 350, 678–680. [Google Scholar] [CrossRef]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron homeostasis and oxidative stress: An intimate relationship. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118535. [Google Scholar] [CrossRef]
- Morgan, B.; Lahav, O. The effect of pH on the kinetics of spontaneous Fe(II) oxidation by O2 in aqueous solution--basic principles and a simple heuristic description. Chemosphere 2007, 68, 2080–2084. [Google Scholar] [CrossRef] [PubMed]
- Ludt, K.; Soppa, J. Influence of Origin Recognition Complex Proteins on the Copy Numbers of Three Chromosomes in Haloferax volcanii. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef]
- Seong, C.N.; Kang, J.W.; Lee, J.H.; Seo, S.Y.; Woo, J.J.; Park, C.; Bae, K.S.; Kim, M.S. Taxonomic hierarchy of the phylum Firmicutes and novel Firmicutes species originated from various environments in Korea. J. Microbiol. 2018, 56, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Márquez, M.C.; Garabito, M.J.; Arahal, D.R. Moderately halophilic gram-positive bacterial diversity in hypersaline environments. Extremophiles 1998, 2, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Menes, R.J.; Machin, E.V.; Iriarte, A.; Langleib, M. Bacillus natronophilus sp. nov., an alkaliphilic bacterium isolated from a soda lake. Int. J. Syst. Evol. Microbiol. 2020, 70, 562–568. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Bagheri, M.; Makhdoumi-Kakhki, A.; Didari, M.; Schumann, P.; Spröer, C.; Sánchez-Porro, C.; Ventosa, A. Oceanobacillus limi sp. nov., a moderately halophilic bacterium from a salt lake. Int. J. Syst. Evol. Microbiol. 2014, 64, 1284–1289. [Google Scholar] [CrossRef]
- Storey, E.P.; Boghozian, R.; Little, J.L.; Lowman, D.W.; Chakraborty, R. Characterization of ‘Schizokinen’; a dihydroxamate-type siderophore produced by Rhizobium leguminosarum IARI 917. Biometals 2006, 19, 637–649. [Google Scholar] [CrossRef]
- Chuljerm, H.; Chen, Y.-L.; Srichairatanakool, S.; Hider, R.C.; Cilibrizzi, A. Synthesis and iron coordination properties of schizokinen and its imide derivative. Dalton Trans. 2019, 48, 17395–17401. [Google Scholar] [CrossRef]
- Cuív, P.O.; Clarke, P.; O’Connell, M. Identification and characterization of an iron-regulated gene, chtA, required for the utilization of the xenosiderophores aerobactin, rhizobactin 1021 and schizokinen by Pseudomonas aeruginosa. Microbiology 2006, 152, 945–954. [Google Scholar] [CrossRef]
- Zawadzka, A.M.; Abergel, R.J.; Nichiporuk, R.; Andersen, U.N.; Raymond, K.N. Siderophore-mediated iron acquisition systems in Bacillus cereus: Identification of receptors for anthrax virulence-associated petrobactin. Biochemistry 2009, 48, 3645–3657. [Google Scholar] [CrossRef]
- Oberto, J. SyntTax: A web server linking synteny to prokaryotic taxonomy. BMC Bioinform. 2013, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Wolters, M.; Borst, A.; Pfeiffer, F.; Soppa, J. Bioinformatic and genetic characterization of three genes localized adjacent to the major replication origin of Haloferax volcanii. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, O.; Gronau, S.; Pfeiffer, F.; Mendoza, E.; Zimmer, R.; Oesterhelt, D. Systems analysis of bioenergetics and growth of the extreme halophile Halobacterium salinarum. PLoS Comput. Biol. 2009, 5, e1000332. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.J. Polyamine function in archaea and bacteria. J. Biol. Chem. 2018, 293, 18693–18701. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niessen, N.; Soppa, J. Regulated Iron Siderophore Production of the Halophilic Archaeon Haloferax volcanii. Biomolecules 2020, 10, 1072. https://doi.org/10.3390/biom10071072
Niessen N, Soppa J. Regulated Iron Siderophore Production of the Halophilic Archaeon Haloferax volcanii. Biomolecules. 2020; 10(7):1072. https://doi.org/10.3390/biom10071072
Chicago/Turabian StyleNiessen, Natalie, and Jörg Soppa. 2020. "Regulated Iron Siderophore Production of the Halophilic Archaeon Haloferax volcanii" Biomolecules 10, no. 7: 1072. https://doi.org/10.3390/biom10071072
APA StyleNiessen, N., & Soppa, J. (2020). Regulated Iron Siderophore Production of the Halophilic Archaeon Haloferax volcanii. Biomolecules, 10(7), 1072. https://doi.org/10.3390/biom10071072