In Vivo Microdialysis of Endogenous and 13C-labeled TCA Metabolites in Rat Brain: Reversible and Persistent Effects of Mitochondrial Inhibition and Transient Cerebral Ischemia

 ,
, 

Abstract

1. Introduction
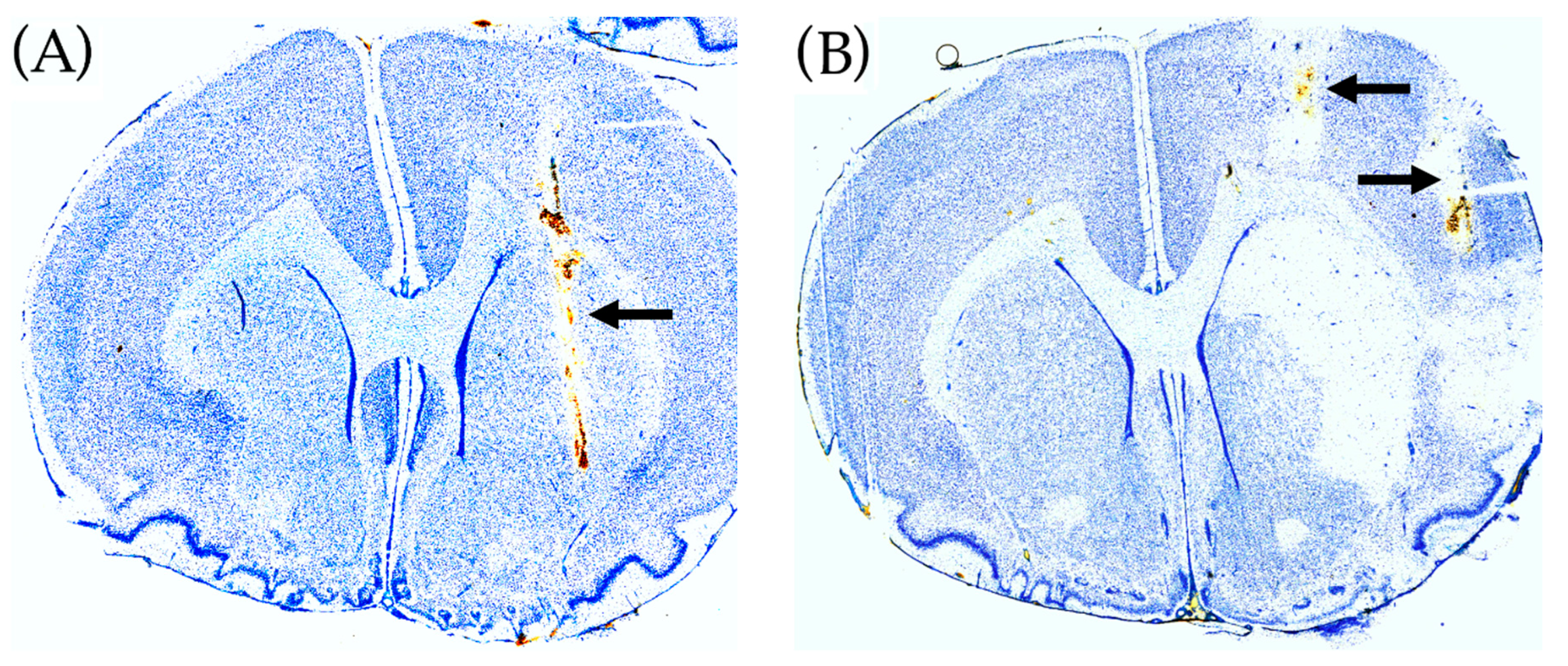
- (a)
- Mitochondrial dysfunction induced by local perfusion with malonate, a reversible inhibitor of SDH.
- (b)
- Transient cerebral ischemia induced by intracerebral application of the potent vasoconstrictor ET-1.
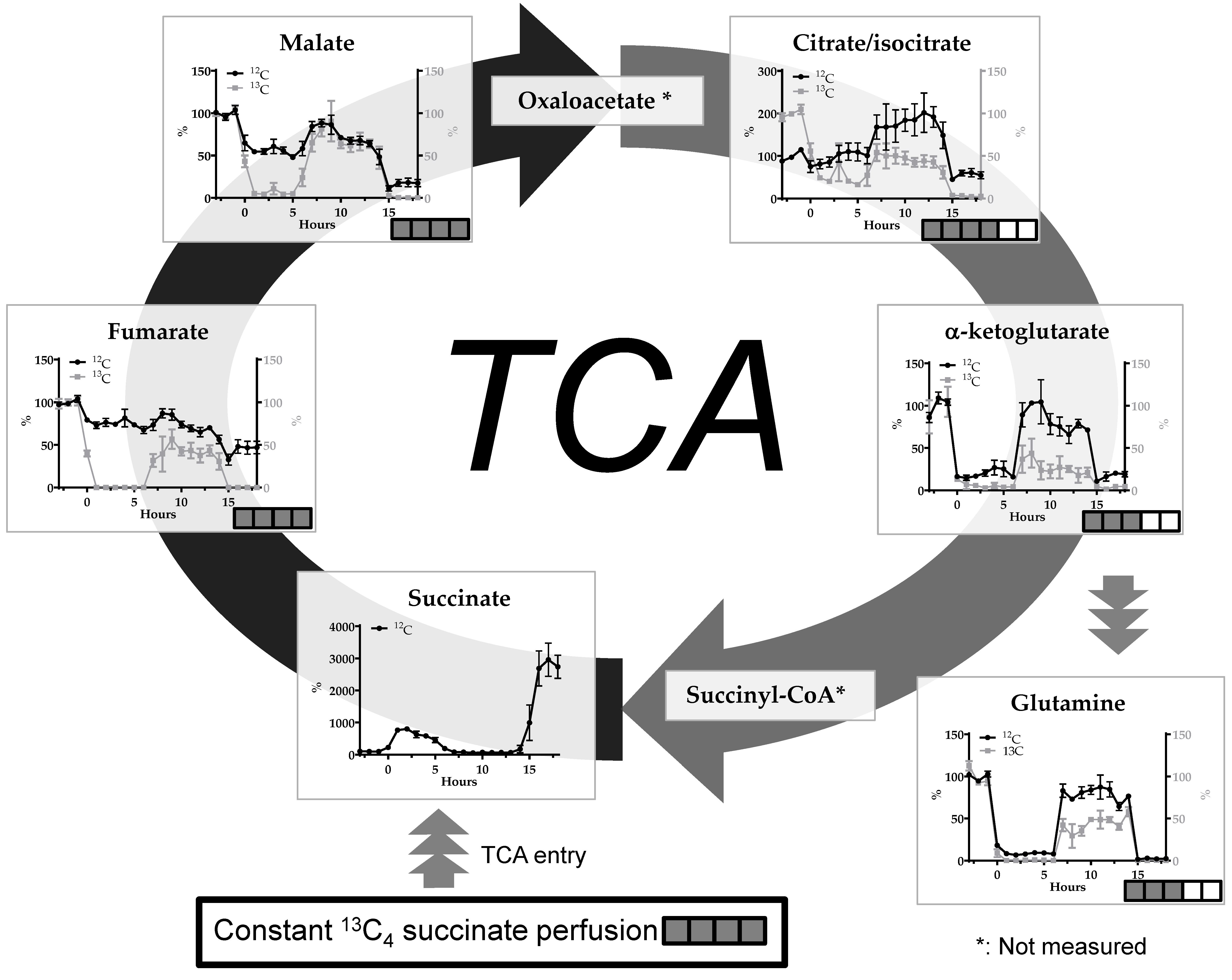
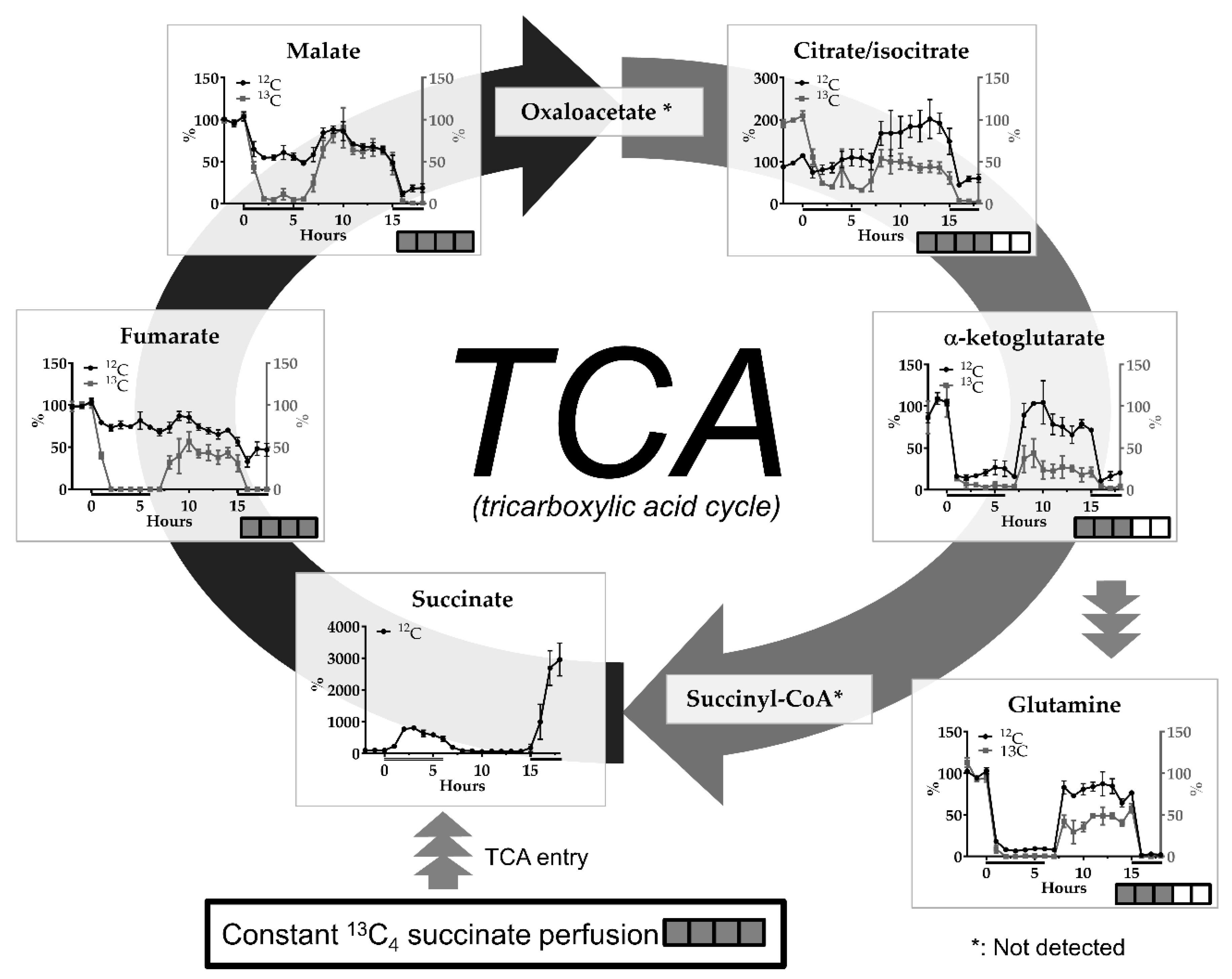
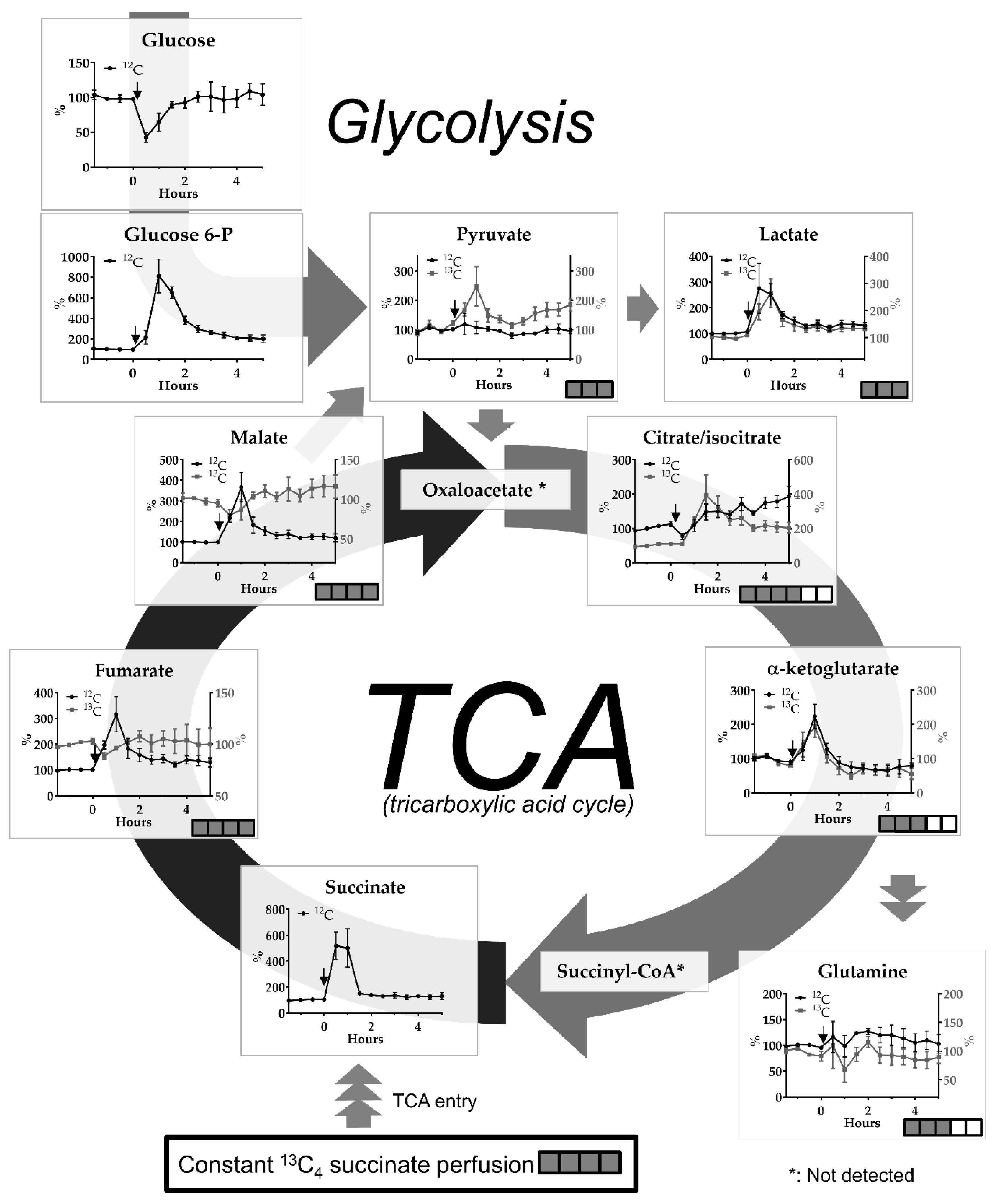
2. Results
2.1. The malonate Model
2.2. Transient Cerebral Ischemia Model
2.3. 13C-labeling %
2.4. Histological Brain Damage
3. Discussion
3.1. General
3.2. The Malonate Model
3.3. The ET-1-induced Transient Ischemia Model
4. Conclusions
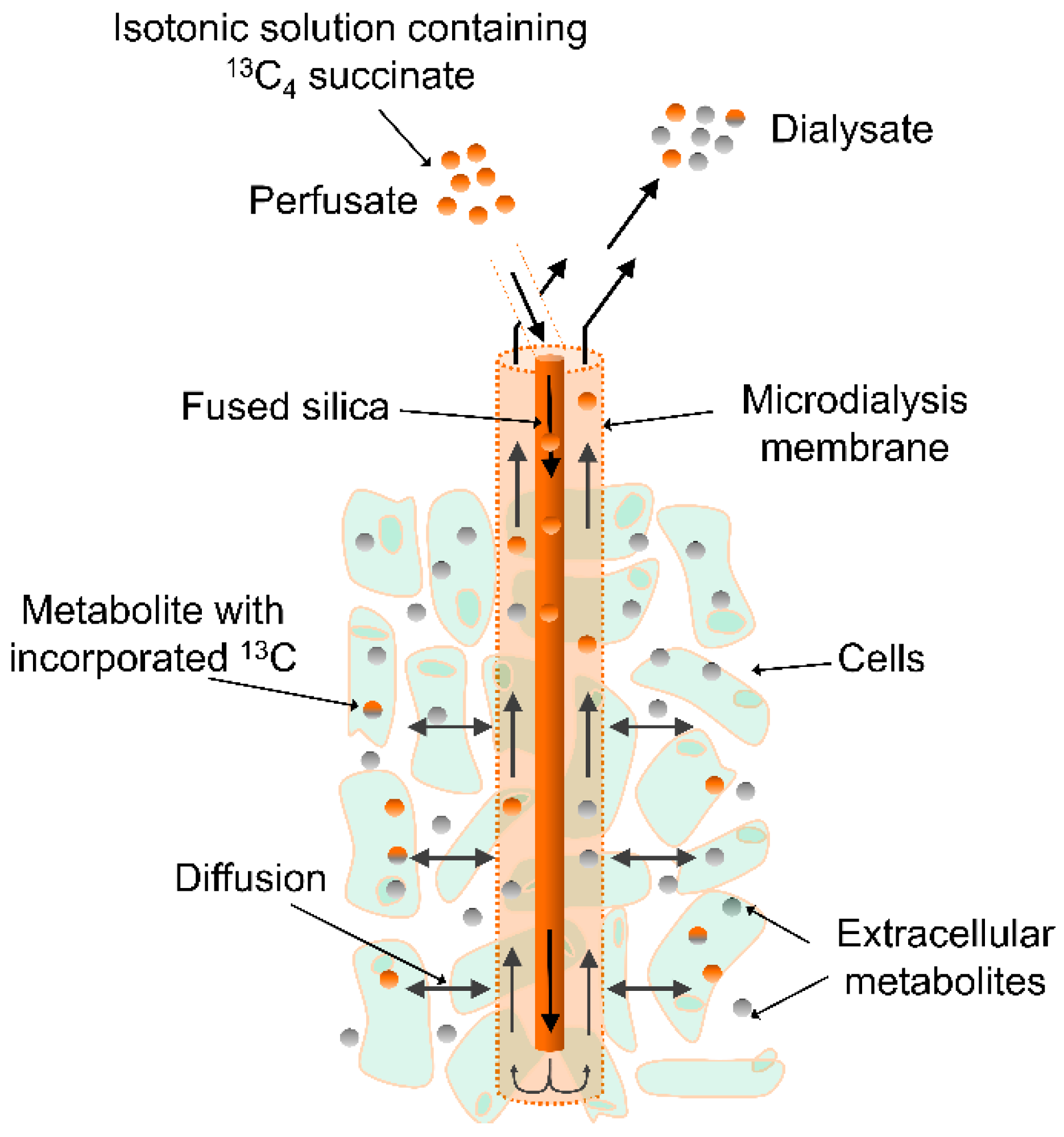
5. Materials and Methods
5.1. Animals
5.2. Stereotaxic Surgery
5.3. Microdialysis Setup for ET-1-Experiments
5.4. Mitochondrial Inhibition by Malonate Perfusion
5.5. ET-1 Induced Transient Cerebral Ischemia
Histology
5.6. Statistical Analysis
5.7. LC-MS Sample Preparation and Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nordstrom, C.H. Cerebral microdialysis in TBI-limitations and possibilities. Acta Neurochir. 2017, 159, 2275–2277. [Google Scholar] [CrossRef] [PubMed]
- Nordstrom, C.H.; Nielsen, T.H.; Schalen, W.; Reinstrup, P.; Ungerstedt, U. Biochemical indications of cerebral ischaemia and mitochondrial dysfunction in severe brain trauma analysed with regard to type of lesion. Acta Neurochir. 2016, 158, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.; Nielsen, T.H.; Nilsson, O.; Schalen, W.; Nordstrom, C.H. Bedside diagnosis of mitochondrial dysfunction in aneurysmal subarachnoid hemorrhage. Acta Neurol. Scand. 2014, 130, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Sarrafzadeh, A.S.; Sakowitz, O.W.; Kiening, K.L.; Benndorf, G.; Lanksch, W.R.; Unterberg, A.W. Bedside microdialysis: A tool to monitor cerebral metabolism in subarachnoid hemorrhage patients? Crit. Care Med. 2002, 30, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Zielke, H.R.; Zielke, C.L.; Baab, P.J. Direct measurement of oxidative metabolism in the living brain by microdialysis: A review. J. Neurochem. 2009, 109 (Suppl. 1), 24–29. [Google Scholar] [CrossRef]
- Carpenter, K.L.; Jalloh, I.; Gallagher, C.N.; Grice, P.; Howe, D.J.; Mason, A.; Timofeev, I.; Helmy, A.; Murphy, M.P.; Menon, D.K.; et al. (13)C-labelled microdialysis studies of cerebral metabolism in TBI patients. Eur. J. Pharm. Sci. 2014, 57, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Jalloh, I.; Helmy, A.; Howe, D.J.; Shannon, R.J.; Grice, P.; Mason, A.; Gallagher, C.N.; Stovell, M.G.; van der Heide, S.; Murphy, M.P.; et al. Focally perfused succinate potentiates brain metabolism in head injury patients. J. Cereb. Blood Flow Metab. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jalloh, I.; Helmy, A.; Howe, D.J.; Shannon, R.J.; Grice, P.; Mason, A.; Gallagher, C.N.; Murphy, M.P.; Pickard, J.D.; Menon, D.K.; et al. A Comparison of Oxidative Lactate Metabolism in Traumatically Injured Brain and Control Brain. J. Neurotrauma 2018, 35, 2025–2035. [Google Scholar] [CrossRef] [PubMed]
- Pajor, A.M. Sodium-coupled dicarboxylate and citrate transporters from the SLC13 family. Pflugers Arch. 2014, 466, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Waagepetersen, H.S.; Qu, H.; Hertz, L.; Sonnewald, U.; Schousboe, A. Demonstration of pyruvate recycling in primary cultures of neocortical astrocytes but not in neurons. Neurochem. Res. 2002, 27, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Olstad, E.; Olsen, G.M.; Qu, H.; Sonnewald, U. Pyruvate recycling in cultured neurons from cerebellum. J. Neurosci. Res. 2007, 85, 3318–3325. [Google Scholar] [CrossRef] [PubMed]
- Huinink, K.; Korf, J.; Gramsbergen, J.B. Microdialysis and microfiltration: Technology and cerebral applications for energy substrates. In Neural Metabolism In Vivo; Gruetter, R., Choi, I.-Y., Eds.; Springer: New York, NY, USA, 2012; Volume 4. [Google Scholar] [CrossRef]
- Walls, A.B.; Bak, L.K.; Sonnewald, U.; Schousboe, A.; Waagepetersen, H.S. Metabolic Mapping of Astrocytes and Neurons in Culture Using Stable Isotopes and Gas Chromatography-Mass Spectrometry (GC-MS). In Brain Energy Metabolism; Hirrlinger, J., Waagepetersen, H.S., Eds.; Humana Press: New York, NY, USA, 2014. [Google Scholar]
- Skjoeth-Rasmussen, J.; Lambertsen, K.; Gramsbergen, J.B. On-line glucose and lactate monitoring in rat striatum: Effect of malonate and correlation with histological damage. J. Neurochem. 2003, 85, 35. [Google Scholar] [CrossRef]
- Fawaz, E.N.; Fawaz, G. Inhibition of glycolysis in rat skeletal muscle by malonate. Biochem. J. 1962, 83, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Siegel, G.J.; Agranoff, B.W. Basic Neurochemistry: Molecular, Cellular, and Medical Aspects; Lippincott Williams Wilkins: Philadelphia, PA, USA, 1999. [Google Scholar]
- Gramsbergen, J.B.; Sandberg, M.; Kornblit, B.; Zimmer, J. Pyruvate protects against 3-nitropropionic acid neurotoxicity in corticostriatal slice cultures. Neuroreport 2000, 11, 2743–2747. [Google Scholar] [CrossRef] [PubMed]
- Sonnewald, U.; Rae, C. Pyruvate carboxylation in different model systems studied by (13)C MRS. Neurochem. Res. 2010, 35, 1916–1921. [Google Scholar] [CrossRef] [PubMed]
- Bak, L.K.; Schousboe, A.; Waagepetersen, H.S. The glutamate/GABA-glutamine cycle: Aspects of transport, neurotransmitter homeostasis and ammonia transfer. J. Neurochem. 2006, 98, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Petzold, G.C.; Einhaupl, K.M.; Dirnagl, U.; Dreier, J.P. Ischemia triggered by spreading neuronal activation is induced by endothelin-1 and hemoglobin in the subarachnoid space. Ann. Neurol. 2003, 54, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Van Hemelrijck, A.; Vermijlen, D.; Hachimi-Idrissi, S.; Sarre, S.; Ebinger, G.; Michotte, Y. Effect of resuscitative mild hypothermia on glutamate and dopamine release, apoptosis and ischaemic brain damage in the endothelin-1 rat model for focal cerebral ischaemia. J. Neurochem. 2003, 87, 66–75. [Google Scholar] [CrossRef]
- Gramsbergen, J.B.; Skjoth-Rasmussen, J.; Rasmussen, C.; Lambertsen, K.L. On-line monitoring of striatum glucose and lactate in the endothelin-1 rat model of transient focal cerebral ischemia using microdialysis and flow-injection analysis with biosensors. J. Neurosci. Methods 2004, 140, 93–101. [Google Scholar] [CrossRef]
- Forsse, A.; Nielsen, T.H.; Nygaard, K.H.; Nordstrom, C.H.; Gramsbergen, J.B.; Poulsen, F.R. Cyclosporin A ameliorates cerebral oxidative metabolism and infarct size in the endothelin-1 rat model of transient cerebral ischaemia. Sci. Rep. 2019, 9, 3702. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijevic, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Tretter, L.; Patocs, A.; Chinopoulos, C. Succinate, an intermediate in metabolism, signal transduction, ROS, hypoxia, and tumorigenesis. Biochim. Biophys. Acta 2016, 1857, 1086–1101. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Lemire, J.; Mailloux, R.J.; Appanna, V.D. A novel strategy involved in [corrected] anti-oxidative defense: The conversion of NADH into NADPH by a metabolic network. PLoS ONE 2008, 3, e2682. [Google Scholar] [CrossRef]
- Samuelsson, C.; Hillered, L.; Zetterling, M.; Enblad, P.; Hesselager, G.; Ryttlefors, M.; Kumlien, E.; Lewen, A.; Marklund, N.; Nilsson, P.; et al. Cerebral glutamine and glutamate levels in relation to compromised energy metabolism: A microdialysis study in subarachnoid hemorrhage patients. J. Cereb. Blood Flow Metab. 2007, 27, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Labeling | Fumaric Acid | Malic Acid | (Iso) Citrate | α-Ketogluarate | Glutamine | Pyruvate | Lactic Acid |
|---|---|---|---|---|---|---|---|
| Baseline | 88.1 (+/− 0.2) | 91.3 (+/− 0.4) | 8.4 (+/− 0.9) | 14.0 (+/− 2.0) | 6.1 (+/− 0.6) | 6.3 (+/− 1.3) | 0.3 (+/− 0.04) |
| Ischemia + 30 min of Reperfusion | 79.2 (+/− 2.1) ** | 75.3 (+/− 2.6) **** | 11.9 (+/− 2.8) | 15.9 (+/− 2.7) | 4.7 (+/− 0.7) | 8.7 (+/− 1.4) | 0.2 (+/− 0.02) |
| After >4 h Reperfusion | 87.5 (+/− 0.5) | 91.1 (0.4) | 9.6 (+/− 1.0) | 14.1 (+/− 3.0) | 5.3 (+/− 0.8) | 8.4 (+/− 1.3) | 0.3 (+/− 0.03) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havelund, J.F.; Nygaard, K.H.; Nielsen, T.H.; Nordström, C.-H.; Poulsen, F.R.; Færgeman, N.J.; Forsse, A.; Gramsbergen, J.B. In Vivo Microdialysis of Endogenous and 13C-labeled TCA Metabolites in Rat Brain: Reversible and Persistent Effects of Mitochondrial Inhibition and Transient Cerebral Ischemia. Metabolites 2019, 9, 204. https://doi.org/10.3390/metabo9100204
Havelund JF, Nygaard KH, Nielsen TH, Nordström C-H, Poulsen FR, Færgeman NJ, Forsse A, Gramsbergen JB. In Vivo Microdialysis of Endogenous and 13C-labeled TCA Metabolites in Rat Brain: Reversible and Persistent Effects of Mitochondrial Inhibition and Transient Cerebral Ischemia. Metabolites. 2019; 9(10):204. https://doi.org/10.3390/metabo9100204
Chicago/Turabian StyleHavelund, Jesper F., Kevin H. Nygaard, Troels H. Nielsen, Carl-Henrik Nordström, Frantz R. Poulsen, Nils. J. Færgeman, Axel Forsse, and Jan Bert Gramsbergen. 2019. "In Vivo Microdialysis of Endogenous and 13C-labeled TCA Metabolites in Rat Brain: Reversible and Persistent Effects of Mitochondrial Inhibition and Transient Cerebral Ischemia" Metabolites 9, no. 10: 204. https://doi.org/10.3390/metabo9100204
APA StyleHavelund, J. F., Nygaard, K. H., Nielsen, T. H., Nordström, C.-H., Poulsen, F. R., Færgeman, N. J., Forsse, A., & Gramsbergen, J. B. (2019). In Vivo Microdialysis of Endogenous and 13C-labeled TCA Metabolites in Rat Brain: Reversible and Persistent Effects of Mitochondrial Inhibition and Transient Cerebral Ischemia. Metabolites, 9(10), 204. https://doi.org/10.3390/metabo9100204