An Improved Validated Method for the Determination of Short-Chain Fatty Acids in Human Fecal Samples by Gas Chromatography with Flame Ionization Detection (GC-FID)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Design
2.2. Chemicals and Reagents
2.3. GC-FID Method
2.4. Standard Preparation and Method Validation
2.5. Sample Preparation for Extraction
2.5.1. Tert-Butyl Methyl Ether Extraction (TBME) Method
2.5.2. Aqueous Extraction (AE) Method
2.5.3. Lyophilization Pre-Treatment
2.6. Statistical Analysis
3. Results
3.1. GC-FID Method Validation
3.2. Sample Preparation
3.3. Application of the Validated Method to Fecal Samples Collected from a Previous Clinical Trial
3.3.1. Participant Demographics
3.3.2. Assessment of Metabolic Status on SCFAs
3.3.3. Assessment of Intervention on SCFAs
4. Discussion
4.1. Method Development and Validation
4.2. Understanding Metabolic Status and SCFAs
4.3. Response to Study Intervention
4.4. Metabolic Fate of SCFAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, E.; Baldi, S.; Ricci, F.; Russo, E.; Nannini, G.; Menicatti, M.; Poli, G.; Taddei, A.; Bartolucci, G.; Calabrò, A.S.; et al. Evaluation and comparison of short chain fatty acids composition in gut diseases. World J. Gastroenterol. 2019, 25, 5543–5558. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Rasmussen, H.S.; Holtug, K.; Mortensen, P.B. Degradation of amino acids to short-chain fatty acids in humans. An in vitro study. Scand. J. Gastroenterol. 1988, 23, 178–182. [Google Scholar] [CrossRef]
- Jeon, B.; Choi, O.; Um, Y.; Byoung-In, S. Production of medium-chain carboxylic acids by Megasphaera sp. MH with supplemental electron acceptors. Biotechnol. Biofuels 2016, 9, 129. [Google Scholar] [CrossRef]
- Ferry, J.G. Acetate kinase and phosphotransacetylase. Methods Enzymol. 2011, 494, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Isipato, M.; Dessì, P.; Sánchez, C.; Mills, S.; Ijaz, U.Z.; Asunis, F.; Spiga, D.; De Gioannis, G.; Mascia, M.; Collins, G.; et al. Propionate Production by Bioelectrochemically-Assisted Lactate Fermentation and Simultaneous CO2 Recycling. Front. Microbiol. 2020, 11, 599438. [Google Scholar] [CrossRef]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef]
- Primec, M.; Mičetić-Turk, D.; Langerholc, T. Analysis of short-chain fatty acids in human feces: A scoping review. Anal. Biochem. 2017, 526, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Hoving, L.R.; Heijink, M.; van Harmelen, V.; van Dijk, K.W.; Giera, M. GC-MS Analysis of Short-Chain Fatty Acids in Feces, Cecum Content, and Blood Samples. Methods Mol. Biol. 2018, 1730, 247–256. [Google Scholar] [CrossRef]
- Scortichini, S.; Boarelli, M.C.; Silvi, S.; Fiorini, D. Development and validation of a GC-FID method for the analysis of short chain fatty acids in rat and human faeces and in fermentation fluids. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1143, 121972. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, H.; Zhu, M.J. A sensitive GC/MS detection method for analyzing microbial metabolites short chain fatty acids in fecal and serum samples. Talanta 2019, 196, 249–254. [Google Scholar] [CrossRef]
- Han, J.; Lin, K.; Sequeira, C.; Borchers, C.H. An isotope-labeled chemical derivatization method for the quantitation of short-chain fatty acids in human feces by liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2015, 854, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Ecker, J.; Roth, S.; Schweizer, S.; Öttl, V.; Schött, H.F.; Yoon, H.; Haller, D.; Holler, E.; Burkhardt, R.; et al. Quantification of Fecal Short Chain Fatty Acids by Liquid Chromatography Tandem Mass Spectrometry-Investigation of Pre-Analytic Stability. Biomolecules 2019, 9, 121. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Functional Deficits in Gut Microbiome of Young and Middle-Aged Adults with Prediabetes Apparent in Metabolizing Bioactive (Poly)phenols. Nutrients 2020, 12, 3595. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, A.; Sandhu, A.K.; Edirisinghe, I.; Burton-Freeman, B.M. Red Raspberry and Fructo-Oligosaccharide Supplementation, Metabolic Biomarkers, and the Gut Microbiota in Adults with Prediabetes: A Randomized Crossover Clinical Trial. J. Nutr. 2022, 152, 1438–1449. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Chen, C.C.; Lin, Y.T.; Wu, W.K.; Chang, L.C.; Lai, C.H.; Wu, M.S.; Kuo, C.H. Evaluation and Optimization of Sample Handling Methods for Quantification of Short-Chain Fatty Acids in Human Fecal Samples by GC-MS. J. Proteome Res. 2019, 18, 1948–1957. [Google Scholar] [CrossRef]
- Eberhart, B.L., 2nd; Wilson, A.S.; O’Keefe, S.J.D.; Ramaboli, M.C.; Nesengani, L.T. A simplified method for the quantitation of short-chain fatty acids in human stool. Anal. Biochem. 2021, 612, 114016. [Google Scholar] [CrossRef]
- Zhao, G.; Nyman, M.; Jönsson, J.A. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed. Chromatogr. 2006, 20, 674–682. [Google Scholar] [CrossRef]
- Verbeke, K.A.; Boobis, A.R.; Chiodini, A.; Edwards, C.A.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; van Tol, E.A.; Tuohy, K.M. Towards microbial fermentation metabolites as markers for health benefits of prebiotics. Nutr. Res. Rev. 2015, 28, 42–66. [Google Scholar] [CrossRef]
- Bravo, J.A.; Julio-Pieper, M.; Forsythe, P.; Kunze, W.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Communication between gastrointestinal bacteria and the nervous system. Curr. Opin. Pharmacol. 2012, 12, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.; Wong, F.S.; Szot, G.L.; Bluestone, J.A.; et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef]
- de la Cuesta-Zuluaga, J.; Mueller, N.T.; Álvarez-Quintero, R.; Velásquez-Mejía, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher Fecal Short-Chain Fatty Acid Levels Are Associated with Gut Microbiome Dysbiosis, Obesity, Hypertension and Cardiometabolic Disease Risk Factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef]
- Weitkunat, K.; Schumann, S.; Nickel, D.; Kappo, K.A.; Petzke, K.J.; Kipp, A.P.; Blaut, M.; Klaus, S. Importance of propionate for the repression of hepatic lipogenesis and improvement of insulin sensitivity in high-fat diet-induced obesity. Mol. Nutr. Food Res. 2016, 60, 2611–2621. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Chung, W.S.F.; Meijerink, M.; Zeuner, B.; Holck, J.; Louis, P.; Meyer, A.S.; Wells, J.M.; Flint, H.J.; Duncan, S.H. Prebiotic potential of pectin and pectic oligosaccharides to promote anti-inflammatory commensal bacteria in the human colon. FEMS Microbiol. Ecol. 2017, 93, fix127. [Google Scholar] [CrossRef]
- Zheng, C.; Liu, R.; Xue, B.; Luo, J.; Gao, L.; Wang, Y.; Ou, S.; Li, S.; Peng, X. Impact and consequences of polyphenols and fructooligosaccharide interplay on gut microbiota in rats. Food Funct. 2017, 8, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Alles, M.S.; Hautvast, J.G.; Nagengast, F.M.; Hartemink, R.; Van Laere, K.M.; Jansen, J.B. Fate of fructo-oligosaccharides in the human intestine. Br. J. Nutr. 1996, 76, 211–221. [Google Scholar] [CrossRef]
- Scholtens, P.A.; Alles, M.S.; Willemsen, L.E.; van den Braak, C.; Bindels, J.G.; Boehm, G.; Govers, M.J. Dietary fructo-oligosaccharides in healthy adults do not negatively affect faecal cytotoxicity: A randomised, double-blind, placebo-controlled crossover trial. Br. J. Nutr. 2006, 95, 1143–1149. [Google Scholar] [CrossRef]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Clausen, M.R.; Mortensen, P.B. Kinetic studies on colonocyte metabolism of short chain fatty acids and glucose in ulcerative colitis. Gut 1995, 37, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
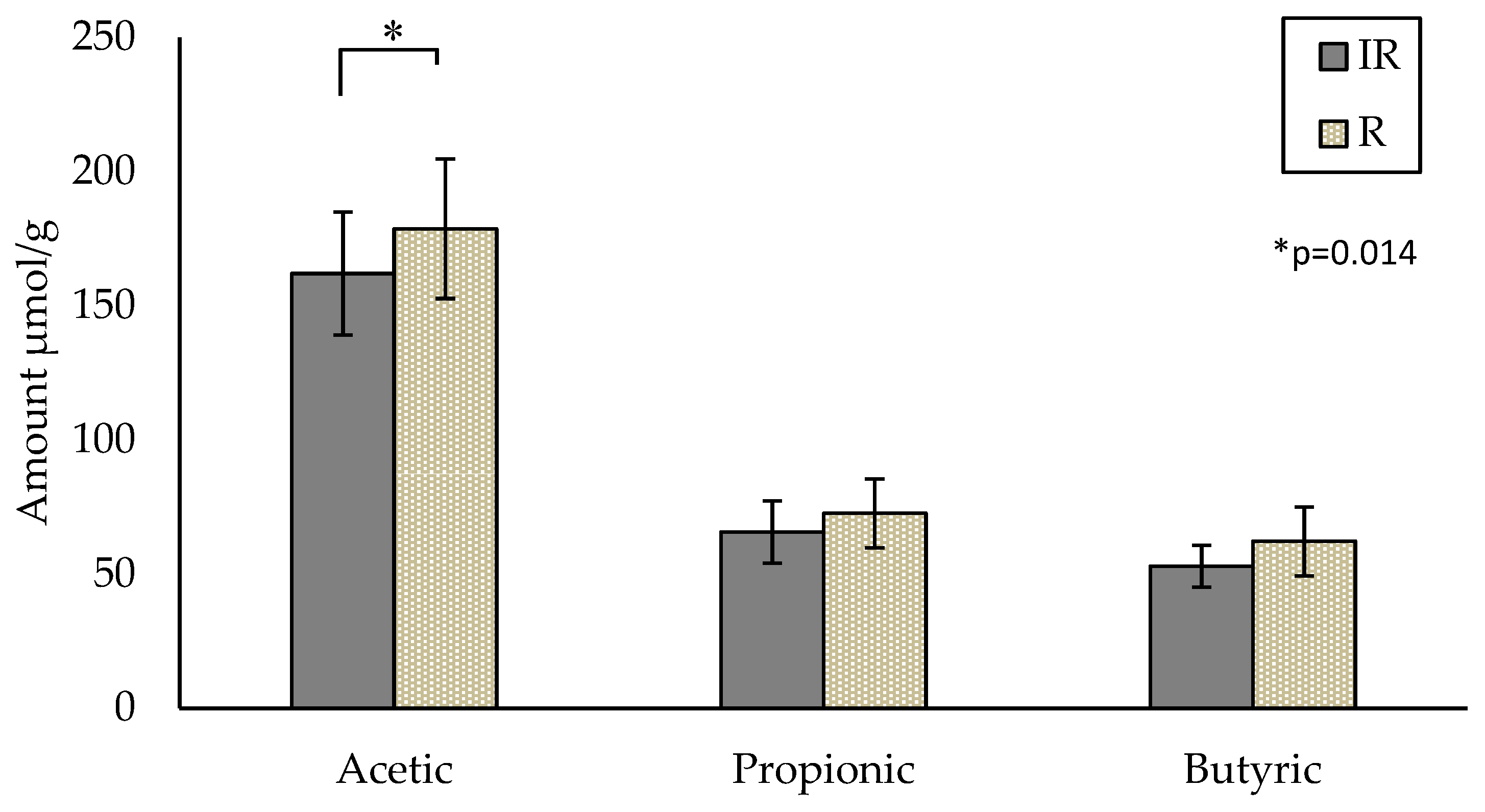

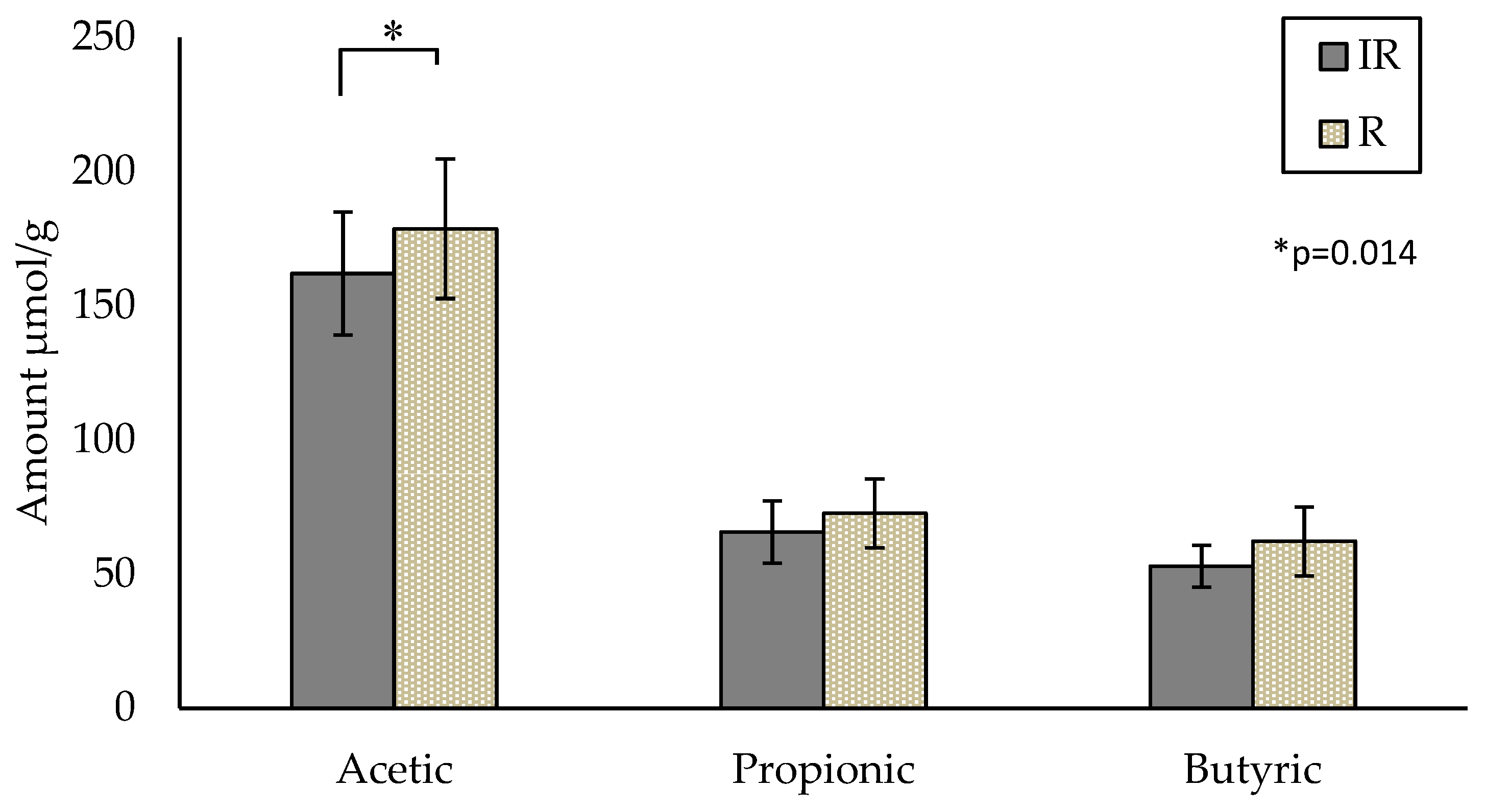{kind=link}
| SCFA | Retention Time (min) | Slope (m) | Intercept (b) | R2 | LOD (µg/mL) | LOQ (µg/mL) | Intra-Day at 2000 ppm (% RSD) | Inter-Day at 50 ppm (% RSD) | Inter-Day at 750 ppm (% RSD) | Inter-Day at 2000 ppm (% RSD) |
|---|---|---|---|---|---|---|---|---|---|---|
| Acetic | 3.32 | 0.04 | 0.002 | 0.99994 | 0.15 | 0.50 | 0.66 | 4.76 | 0.92 | 0.35 |
| Propionic | 3.74 | 0.61 | 0.001 | 0.99996 | 0.17 | 0.56 | 0.64 | 0.85 | 1.52 | 0.93 |
| IsoButyric | 3.91 | 0.08 | 0.002 | 0.99995 | 0.23 | 0.78 | N/A | N/A | N/A | N/A |
| Butyric | 4.20 | 0.08 | 0.001 | 0.99996 | 0.10 | 0.32 | 0.77 | 0.45 | 1.11 | 0.78 |
| IsoValeric | 4.42 | 0.09 | 0.001 | 0.99997 | 0.07 | 0.23 | 0.56 | 0.81 | 0.10 | 0.23 |
| Valeric | 4.78 | 0.91 | 0.000 | 0.99998 | 0.06 | 0.21 | 1.03 | 0.99 | 0.86 | 0.64 |
| 4-methyl valeric | 5.12 | 0.09 | 0.001 | 0.99998 | 0.04 | 0.13 | 0.82 | 0.73 | 0.44 | 0.74 |
| Hexanoic | 5.34 | 1.02 | 0.001 | 0.99998 | 0.04 | 0.12 | 0.82 | 0.80 | 0.35 | 0.18 |
| Heptanoic | 5.89 | 1.05 | 0.003 | 0.99998 | 0.02 | 0.08 | 0.69 | 1.11 | 0.36 | 0.63 |
| Percent (%) Recovery at Respective Spike Levels 1 | ||
|---|---|---|
| 100 ppm | 1000 ppm | |
| Acetic | 57.64 ± 2.89 | 54.24 ± 1.17 |
| Propionic | 87.60 ± 1.64 | 92.82 ± 1.43 |
| IsoButyric | N/A | N/A |
| Butyric | 100.71 ± 2.97 | 106.61 ± 1.54 |
| IsoValeric | 119.96 ± 4.10 | 127.83 ± 1.47 |
| Valeric | 121.41 ± 4.11 | 129.74 ± 1.88 |
| 4-methyl valeric | 107.80 ± 26.30 | 129.52 ± 3.34 |
| Hexanoic | 124.74 ± 4.94 | 129.57 ± 9.63 |
| Heptanoic | 128.97 ± 5.43 | 140.94 ± 2.10 |
| SCFAs | AE Method Amount (µmol/g) 1 | AE Method % RSD | TBME Method Amount (µmol/g) 1 | TBME Method % RSD | p-Value AE vs. TBME 2,3 |
|---|---|---|---|---|---|
| Acetic | 69.42 | 8.63 | 36.80 | 20.94 | <0.0001 |
| Propionic | 20.95 | 8.82 | 12.57 | 19.81 | <0.0001 |
| IsoButyric | 5.89 | 6.82 | 3.53 | 19.45 | <0.0001 |
| Butyric | 22.25 | 2.49 | 13.81 | 19.09 | <0.0001 |
| IsoValeric | 8.48 | 6.05 | 5.24 | 20.21 | <0.0001 |
| Valeric | 5.44 | 2.18 | 3.21 | 19.62 | <0.0001 |
| 4-methyl valeric | ND | N/A | ND | N/A | N/A |
| Hexanoic | 0.57 | 2.15 | 0.25 | 17.98 | <0.0001 |
| Heptanoic | ND | N/A | ND | N/A | N/A |
| SCFAs | FD Amount 1 (µmol/g) | FR Amount 1 (µmol/g) | p-Value FD vs. FR 2 |
|---|---|---|---|
| Acetic | 73.8 ± 7.2 | 69.4 ± 5.2 | 0.504 |
| Propionic | 19.0 ± 1.7 | 21.0 ± 1.6 | 0.270 |
| IsoButyric | 4.8 ± 0.5 | 5.9 ± 0.3 | 0.058 |
| Butyric | 21.0 ± 2.8 | 22.2 ± 0.5 | 0.548 |
| IsoValeric | 7.0 ± 0.8 | 8.4 ± 0.4 | 0.101 |
| Valeric | 4.9 ± 0.7 | 5.4 ± 0.1 | 0.289 |
| 4-methyl valeric | 0.3 ± 0.0 | ND | 1.62 × 10−5 * |
| Hexanoic | 0.6 ± 0.0 | 0.6 ± 0.0 | 0.915 |
| Heptanoic | 0.3 ± 0.1 | ND | 0.011 * |
| Variable | IR-Group (n = 20) | R-Group (n = 9) |
|---|---|---|
| Age (year) 1 | 34.8 ± 1.4 | 30.4 ± 1.7 |
| BMI (kg/m2) 1 | 28.2 ± 0.8 | 23.4 ± 1.0 |
| Female:Male (n) | 9:11 | 6:3 |
| Race (Asian:His:Cau:AA) | 7:2:6:5 | 3:2:2:2 |
| SCFAs | R-Group (n = 9) | IR-Group (n = 20) | ||||||
|---|---|---|---|---|---|---|---|---|
| Baseline (Week 0) Amount (µmol/g) 1 | RRB (Week 4) Amount (µmol/g) 1 | RRB + FOS (Week 4) Amount (µmol/g) 1 | p-Value Interv at Week 4 2 | Baseline (Week 0) Amount (µmol/g) 1 | RRB (Week 4) Amount (µmol/g) 1 | RRB + FOS (Week 4) Amount (µmol/g) 1 | p-Value Interv at Week 4 2 | |
| Acetic | 178.7 ± 26.0 | 159.7 ± 29.2 | 155.3 ± 35.1 | 0.905 | 162.0 ± 22.9 | 142.3 ± 18.4 | 164.6 ± 17.7 | 0.257 |
| Propionic | 72.6 ± 12.9 | 53.7 ± 8.3 | 51.5 ± 9.3 | 0.712 | 65.7 ± 11.7 | 52.9 ± 5.9 | 62.1 ± 10.5 | 0.426 |
| IsoButyric | 6.6 ± 1.7 | 6.2 ± 1.1 | 5.2 ± 0.8 | 0.412 | 6.1 ± 0.7 | 5.8 ± 0.6 | 4.9 ± 0.5 | 0.208 |
| Butyric | 62.2 ± 12.9 | 52.4 ± 14.5 | 48.7 ± 7.9 | 0.781 | 52.9 ± 7.8 | 45.3 ± 4.9 | 42.6 ± 3.9 | 0.640 |
| IsoValeric | 10.5 ± 3.0 | 9.3 ± 1.7 | 7.7 ± 1.2 | 0.394 | 9.0 ± 1.0 | 8.7 ± 0.9 | 6.9 ± 0.7 | 0.097 |
| Valeric | 7.7 ± 1.7 | 7.3 ± 1.7 | 6.1 ± 1.3 | 0.438 | 6.9 ± 1.1 | 5.4 ± 0.7 | 5.3 ± 0.7 | 0.918 |
| 4-Methyl Valeric | 0.5 ± 0.3 | 0.1 ± 0.1 | 0.3 ± 0.1 | 0.040 * | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.827 |
| Hexanoic | 1.8 ± 0.7 | 1.0 ± 0.4 | 2.1 ± 0.8 | 0.224 | 1.5 ± 0.5 | 1.2 ± 0.5 | 1.3 ± 0.4 | 0.886 |
| Heptanoic | 0.7 ± 0.2 | 0.1 ± 0.1 | 0.5 ± 0.2 | 0.073 | 1.0 ± 0.7 | 0.2 ± 0.1 | 0.4 ± 0.1 | 0.352 |
| Total SCFA | 341.4 ± 51.5 | 289.7 ± 53.4 | 277.3 ± 50.6 | 0.827 | 305.3 ± 40.3 | 262.1 ± 28.4 | 288.3 ± 28.9 | 0.442 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, M.; Polite, L.; Christy, A.; Edirisinghe, I.; Burton-Freeman, B.; Sandhu, A. An Improved Validated Method for the Determination of Short-Chain Fatty Acids in Human Fecal Samples by Gas Chromatography with Flame Ionization Detection (GC-FID). Metabolites 2023, 13, 1106. https://doi.org/10.3390/metabo13111106
Smith M, Polite L, Christy A, Edirisinghe I, Burton-Freeman B, Sandhu A. An Improved Validated Method for the Determination of Short-Chain Fatty Acids in Human Fecal Samples by Gas Chromatography with Flame Ionization Detection (GC-FID). Metabolites. 2023; 13(11):1106. https://doi.org/10.3390/metabo13111106
Chicago/Turabian StyleSmith, Morganne, Lee Polite, Andreas Christy, Indika Edirisinghe, Britt Burton-Freeman, and Amandeep Sandhu. 2023. "An Improved Validated Method for the Determination of Short-Chain Fatty Acids in Human Fecal Samples by Gas Chromatography with Flame Ionization Detection (GC-FID)" Metabolites 13, no. 11: 1106. https://doi.org/10.3390/metabo13111106
APA StyleSmith, M., Polite, L., Christy, A., Edirisinghe, I., Burton-Freeman, B., & Sandhu, A. (2023). An Improved Validated Method for the Determination of Short-Chain Fatty Acids in Human Fecal Samples by Gas Chromatography with Flame Ionization Detection (GC-FID). Metabolites, 13(11), 1106. https://doi.org/10.3390/metabo13111106