
Advances in Cell Engineering of the Komagataella phaffii Platform for Recombinant Protein Production

 ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Metabolic Engineering for Improved Metabolism and Energy Supply
2.1. Engineering of the Methanol Catabolic Pathway
2.2. Engineering of Co-Substrate Catabolic Pathways
2.3. Engineering of Turnover Co-Factor Metabolism
2.4. Engineering for Alternative Carbon Source Catabolism
3. Transcription Factor Engineering in K. phaffii for Recombinant Protein Production
4. Improving Protein Folding and Secretion

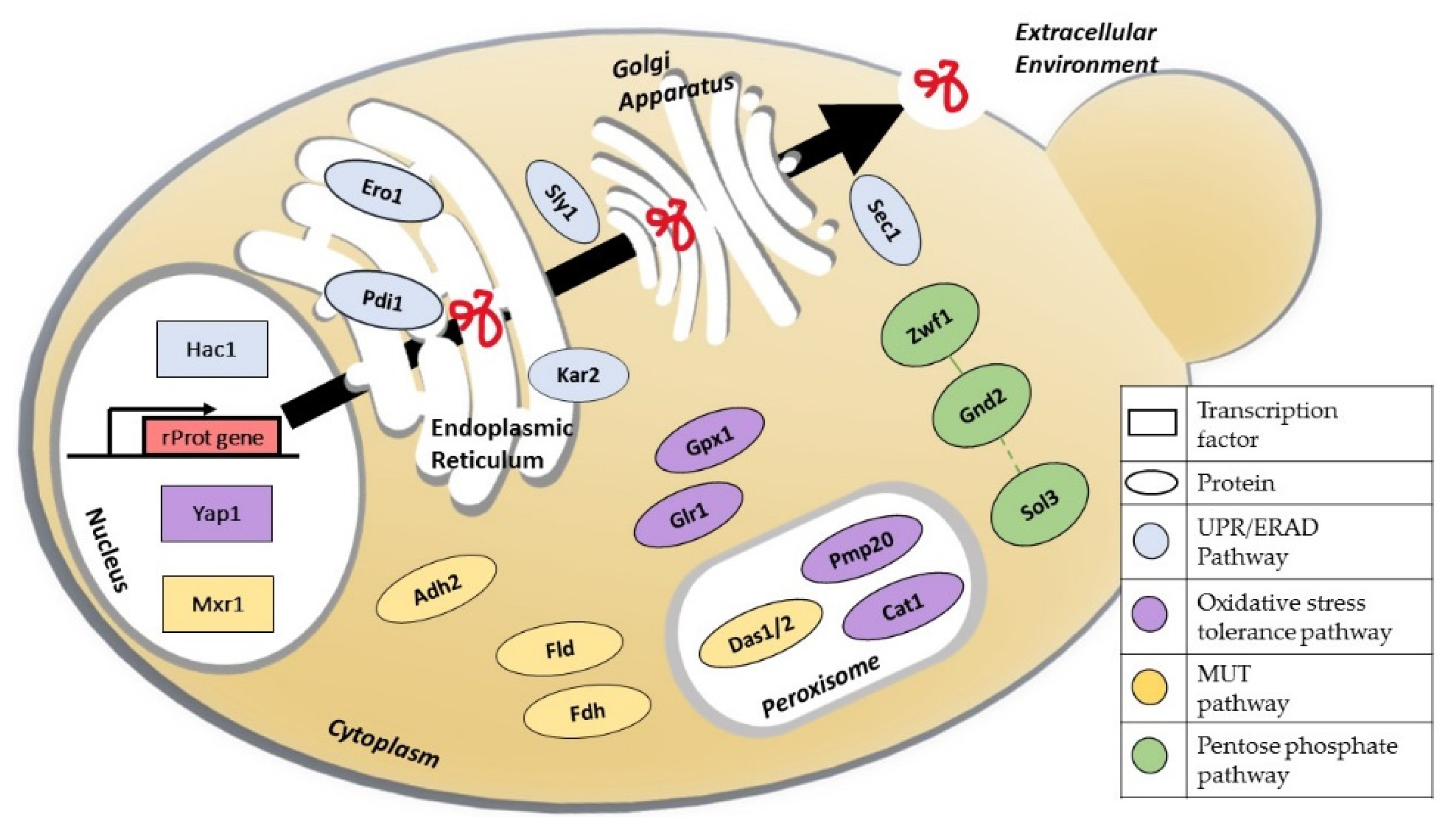{kind=link}
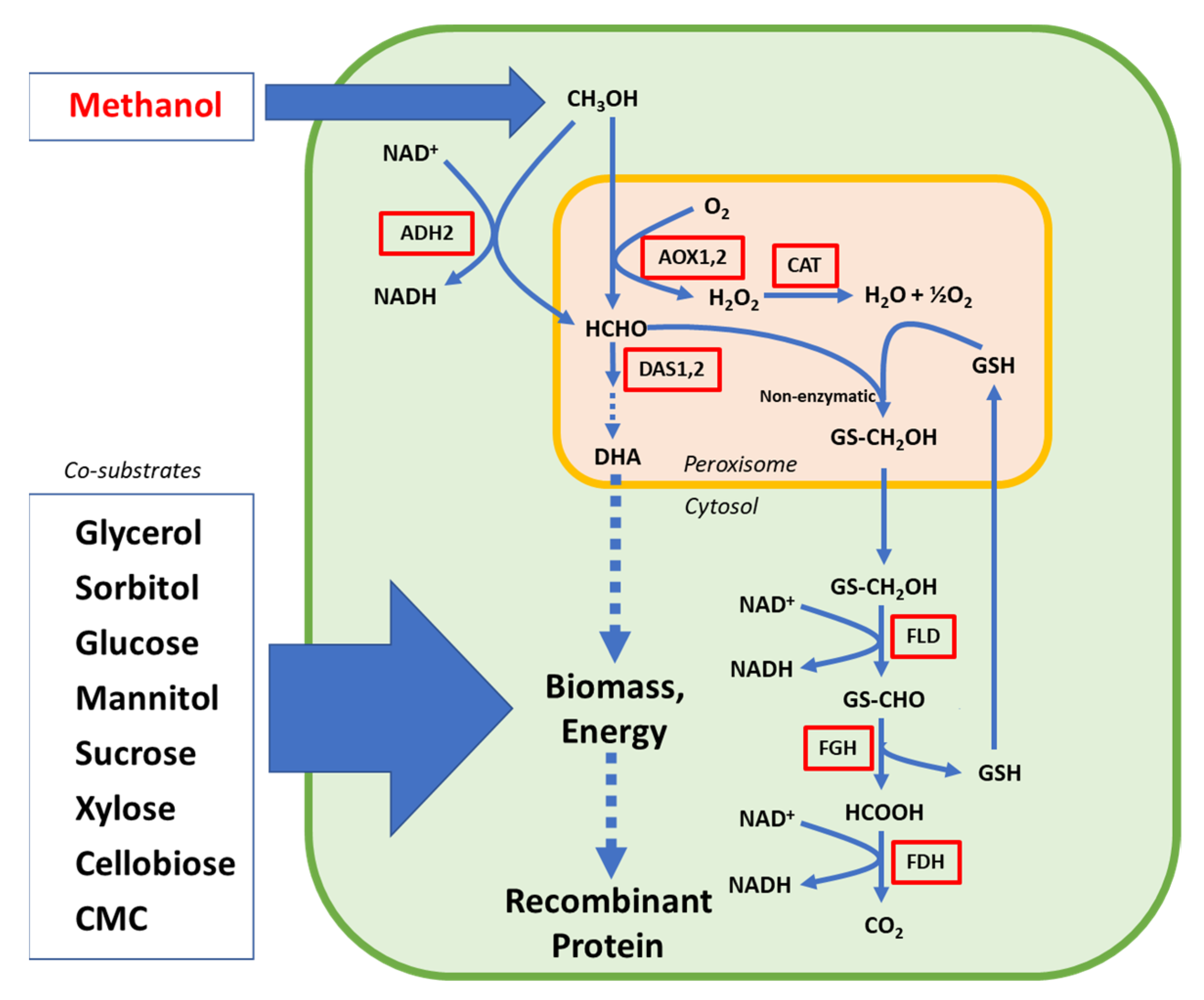{kind=link}
| Auxiliary Gene | Modification | Pathways Involved | Heterologous Product | Production (Fold Change) | Operation Mode | Scale | Strain | Ref. |
|---|---|---|---|---|---|---|---|---|
| PDI1 | OE | Folding | Fab (S) | +1.9 | Batch | Flask | X-33 | [117] |
| PDI1 PDI1 w/ CN | OE | Folding | Na-ASP1 | +3.2 + 7.9 | Batch | Flask | X-33 | [118] |
| KAR2 PDI1 PDI1/KAR2 | OE | Folding | A33scFv (S) | +3 No effect No effect | Batch | 2.5 L Bioreactor | GS200 | [119] |
| PDI1 KAR2 ERO1 SEC1 SLY1 | OE | Folding and Trafficking | IL2-HSA (S) | +2.2 +1.9 +2.3 +2.5 +1.9 | Batch | Flask | GS115 | [111] |
| PDI1 + CN KAR2 + CN ERO1 CN | OE | Folding | Hydrophobin (S) | +7.8 +22 +30 No effect | Batch | Flask | GS115 | [108] |
| PDI1 ERO1 CN | OE | Folding | hLYZ (S) | +2.43 +2.30 +1.57 | Batch | 5 L Bioreactor | GS115 | [7] |
| YDJ1 SSA1 SEC63 KAR2 | OE | Folding and Trafficking | CalB (S) | +1.6 +1.4 +1.4 −0.7 | Batch | Flask | GS115 | [115] |
| PDI1 ERO1 GPX1 GLR1 YAP1 | OE | Folding | RABV-G (S) | +9.5 +3.3 +8.2 +1.2 No effect | Batch | Flask | KM71H/GS115 | [114,116] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kurtzman, C.P. Biotechnological strains of Komagataella (Pichia) pastoris are Komagataella phaffii as determined from multigene sequence analysis. J. Ind. Microbiol. Biotechnol. 2009, 36, 1435–1438. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Sun, H.; Wang, M.; Li, Y. Pichia pastoris as a Versatile Cell Factory for the Production of Industrial Enzymes and Chemicals: Current Status and Future Perspectives. Biotechnol. J. 2019, 14, e1800694. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [PubMed]
- Peña, D.A.; Gasser, B.; Zanghellini, J.; Steiger, M.; Mattanovich, D. Metabolic engineering of Pichia pastoris. Metab. Eng. 2018, 50, 2–15. [Google Scholar] [CrossRef]
- Maccani, A.; Landes, N.; Stadlmayr, G.; Maresch, D.; Leitner, C.; Maurer, M.; Gasser, B.; Ernst, W.; Kunert, R.; Mattanovich, D. Pichia pastoris secretes recombinant proteins less efficiently than Chinese hamster ovary cells but allows higher space-time yields for less complex proteins. Biotechnol. J. 2014, 9, 526–537. [Google Scholar] [CrossRef]
- Raza, A.; Pothula, R.; Abdelgaffar, H.; Bashir, S.; Jurat-Fuentes, J.L. Supplemental Information 3: Gene sequence for beta-glucosidase from Bacillus tequelensis. PeerJ 2020, 8, e8792. [Google Scholar] [CrossRef]
- He, H.; Wu, S.; Mei, M.; Ning, J.; Li, C.; Ma, L.; Zhang, G.; Yi, L. A Combinational Strategy for Effective Heterologous Production of Functional Human Lysozyme in Pichia pastoris. Front. Bioeng. Biotechnol. 2020, 8, 1–12. [Google Scholar] [CrossRef]
- Ata, Ö.; Ergün, B.G.; Fickers, P.; Heistinger, L.; Mattanovich, D.; Rebnegger, C.; Gasser, B. What makes Komagataella phaffii non-conventional? FEMS Yeast Res. 2021, 21, 21. [Google Scholar] [CrossRef]
- Henríquez, M.; Braun-Galleani, S.; Nesbeth, D.N. Whole cell biosynthetic activity of Komagataella phaffii (Pichia pastoris) GS115 strains engineered with transgenes encoding Chromobacterium violaceum ω-transaminase alone or combined with native transketolase. Biotechnol. Prog. 2020, 36, e2893. [Google Scholar] [CrossRef]
- Braun-Galleani, S.; Henríquez, M.-J.; Nesbeth, D.N. Whole cell biosynthesis of 1-methyl-3-phenylpropylamine and 2-amino-1,3,4-butanetriol using Komagataella phaffii (Pichia pastoris) strain BG-10 engineered with a transgene encoding Chromobacterium violaceum ω-transaminase. Heliyon 2019, 5, e02338. [Google Scholar] [CrossRef]
- Heistinger, L.; Gasser, B.; Mattanovich, D. Microbe Profile: Komagataella phaffii: A methanol devouring biotech yeast formerly known as Pichia pastoris. Microbiol. 2020, 166, 614–616. [Google Scholar] [CrossRef] [PubMed]
- Burgard, J.; Grünwald-Gruber, C.; Altmann, F.; Zanghellini, J.; Valli, M.; Mattanovich, D.; Gasser, B.; Gruber-Grünwald, C. The secretome of Pichia pastoris in fed-batch cultivations is largely independent of the carbon source but changes quantitatively over cultivation time. Microb. Biotechnol. 2019, 13, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Raschmanová, H.; Weninger, A.; Knejzlík, Z.; Melzoch, K.; Kovar, K. Engineering of the unfolded protein response pathway in Pichia pastoris: Enhancing production of secreted recombinant proteins. Appl. Microbiol. Biotechnol. 2021, 105, 4397–4414. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, R.; Bergin, A.; Flampouri, E.; Mota, L.M.; Obaidi, I.; Quigley, A.; Xie, Y.; Butler, M. Mammalian cell culture for production of recombinant proteins: A review of the critical steps in their biomanufacturing. Biotechnol. Adv. 2020, 43, 107552. [Google Scholar] [CrossRef]
- Özçelik, A.T.; Yılmaz, S.; Inan, M. Pichia pastoris Promoters. Methods Pharmacol. Toxicol. 2019, 1923, 97–112. [Google Scholar] [CrossRef]
- Jia, L.; Li, T.; Wu, Y.; Wu, C.; Li, H.; Huang, A. Enhanced human lysozyme production by Pichia pastoris via periodic glycerol and dissolved oxygen concentrations control. Appl. Microbiol. Biotechnol. 2021, 105, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.; Haridas, S.; Wolfe, K.H.; Lopes, M.R.; Hittinger, C.T.; Göker, M.; Salamov, A.A.; Wisecaver, J.H.; Long, T.M.; Calvey, C.H.; et al. Comparative genomics of biotechnologically important yeasts. Proc. Natl. Acad. Sci. USA 2016, 113, 9882–9887. [Google Scholar] [CrossRef]
- Heistinger, L.; Gasser, B.; Mattanovich, D. Creation of Stable Heterothallic Strains of Komagataella phaffii Enables Dissection of Mating Gene Regulation. Mol. Cell. Biol. 2018, 38, e00398-17. [Google Scholar] [CrossRef]
- Braun-Galleani, S.; Dias, J.A.; Coughlan, A.Y.; Ryan, A.P.; Byrne, K.P.; Wolfe, K.H. Genomic diversity and meiotic recombination among isolates of the biotech yeast Komagataella phaffii (Pichia pastoris). Microb. Cell Factories 2019, 18, 1–13. [Google Scholar] [CrossRef]
- Nieto-Taype, M.A.; Garcia-Ortega, X.; Albiol, J.; Seguí, J.L.M.; Valero, F. Continuous Cultivation as a Tool Toward the Rational Bioprocess Development With Pichia Pastoris Cell Factory. Front. Bioeng. Biotechnol. 2020, 8, 632. [Google Scholar] [CrossRef]
- Liu, W.-C.; Inwood, S.; Gong, T.; Sharma, A.; Yu, L.-Y.; Zhu, P. Fed-batch high-cell-density fermentation strategies for Pichia pastoris growth and production. Crit. Rev. Biotechnol. 2019, 39, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Bernauer, L.; Radkohl, A.; Lehmayer, L.G.K.; Emmerstorfer-Augustin, A. Komagataella phaffii as Emerging Model Organism in Fundamental Research. Front. Microbiol. 2021, 11, 11. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: A review. Biotechnol. Adv. 2018, 36, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Jiang, L.; Lian, J. Development of synthetic biology tools to engineer Pichia pastoris as a chassis for the production of natural products. Synth. Syst. Biotechnol. 2021, 6, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Schwarzhans, J.-P.; Luttermann, T.; Geier, M.; Kalinowski, J.; Friehs, K. Towards systems metabolic engineering in Pichia pastoris. Biotechnol. Adv. 2017, 35, 681–710. [Google Scholar] [CrossRef]
- Kafri, M.; Metzl-Raz, E.; Jona, G.; Barkai, N. The Cost of Protein Production. Cell Rep. 2015, 14, 22–31. [Google Scholar] [CrossRef]
- Tomàs-Gamisans, M.; Andrade, C.C.P.; Maresca, F.; Monforte, S.; Ferrer, P.; Albiol, J. Redox Engineering by Ectopic Overexpression of NADH Kinase in Recombinant Pichia pastoris (Komagataella phaffii): Impact on Cell Physiology and Recombinant Production of Secreted Proteins. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Jia, L.; Mpofu, E.; Tu, T.; Huai, Q.; Sun, J.; Chen, S.; Ding, J.; Shi, Z. Transcriptional analysis for carbon metabolism and kinetic modeling for heterologous proteins productions by Pichia pastoris in induction process with methanol/sorbitol co-feeding. Process Biochem. 2017, 59, 159–166. [Google Scholar] [CrossRef]
- Cai, H.; Doi, R.; Shimada, M.; Hayakawa, T.; Nakagawa, T. Metabolic regulation adapting to high methanol environment in the methylotrophic yeast Ogataea methanolica. Microb. Biotechnol. 2021, 14, 1512–1524. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Zhao, F.; Zhang, Y.; Yang, X.; Lin, Y.; Han, S. Methanol oxidase from Hansenula polymorpha shows activity in peroxisome-deficient Pichia pastoris. Biochem. Eng. J. 2022, 180, 108369. [Google Scholar] [CrossRef]
- Carly, F.; Niu, H.; Delvigne, F.; Fickers, P. Influence of methanol/sorbitol co-feeding rate on pAOX1 induction in a Pichia pastoris Mut+ strain in bioreactor with limited oxygen transfer rate. J. Ind. Microbiol. Biotechnol. 2016, 43, 517–523. [Google Scholar] [CrossRef]
- Azadi, S.; Mahboubi, A.; Naghdi, N.; Solaimanian, R.; Mortazavi, S.A. Evaluation of Sorbitol-Methanol Co-Feeding Strategy on Production of Recombinant Human Growth Hormone in Pichia Pastoris. Iran. J. Pharm. Res. 2017, 16, 1555–1564. [Google Scholar] [PubMed]
- Berrios, J.; Flores, M.-O.; Díaz-Barrera, A.; Altamirano, C.; Martínez, I.; Cabrera, Z. A comparative study of glycerol and sorbitol as co-substrates in methanol-induced cultures of Pichia pastoris: Temperature effect and scale-up simulation. J. Ind. Microbiol. Biotechnol. 2017, 44, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Zepeda, A.B.; Pessoa, A.; Farías, J.G. Carbon metabolism influenced for promoters and temperature used in the heterologous protein production using Pichia pastoris yeast. Braz. J. Microbiol. 2018, 49, 119–127. [Google Scholar] [CrossRef]
- Niu, H.; Jost, L.; Pirlot, N.; Sassi, H.; Daukandt, M.; Rodriguez, C.; Fickers, P. A quantitative study of methanol/sorbitol co-feeding process of a Pichia pastoris Mut+/pAOX1-lacZ strain. Microb. Cell Factories 2013, 12, 33. [Google Scholar] [CrossRef]
- Liu, W.; Xiang, H.; Zhang, T.; Pang, X.; Su, J.; Liu, H.; Ma, B.; Yu, L. Development of a New High-Cell Density Fermentation Strategy for Enhanced Production of a Fungus β-Glucosidase in Pichia pastoris. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Krainer, F.W.; Dietzsch, C.; Hajek, T.; Herwig, C.; Spadiut, O.; Glieder, A. Recombinant protein expression in Pichia pastoris strains with an engineered methanol utilization pathway. Microb. Cell Factories 2012, 11, 22. [Google Scholar] [CrossRef]
- Geier, M.; Brandner, C.; Strohmeier, G.A.; Hall, M.; Hartner, F.S.; Glieder, A. Engineering Pichia pastoris for improved NADH regeneration: A novel chassis strain for whole-cell catalysis. Beilstein J. Org. Chem. 2015, 11, 1741–1748. [Google Scholar] [CrossRef]
- Zavec, D.; Troyer, C.; Maresch, D.; Altmann, F.; Hann, S.; Gasser, B.; Mattanovich, D. Beyond alcohol oxidase: The methylotrophic yeast Komagataella phaffii utilizes methanol also with its native alcohol dehydrogenase Adh2. FEMS Yeast Res. 2021, 21, 21. [Google Scholar] [CrossRef] [PubMed]
- Tyurin, O.V.; Kozlov, D.G. Deletion of the FLD gene in methylotrophic yeasts Komagataella phaffii and Komagataella kurtzmanii results in enhanced induction of the AOX1 promoter in response to either methanol or formate. Microbiol. 2015, 84, 408–411. [Google Scholar] [CrossRef]
- Liu, T.; Zhao, Y.; Zhang, J.; Zhang, J. Enhancement of xylanase expression by Komagataella phaffii through pexophagy inhibition. Biotechnol. Biotechnol. Equip. 2019, 33, 855–862. [Google Scholar] [CrossRef]
- Nocon, J.; Steiger, M.; Mairinger, T.; Hohlweg, J.; Rußmayer, H.; Hann, S.; Gasser, B.; Mattanovich, D. Increasing pentose phosphate pathway flux enhances recombinant protein production in Pichia pastoris. Appl. Microbiol. Biotechnol. 2016, 100, 5955–5963. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, A.A.; Veeranki, V.D. Metabolic Engineering of Pichia pastoris GS115 for Enhanced Pentose Phosphate Pathway (PPP) Flux toward Recombinant Human Interferon Gamma (HIFN-γ) Production. Mol. Biol. Rep. 2018, 45, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Sun, H.; Chen, Z.; Li, Y.; Zhu, T. Construction of efficient xylose utilizing Pichia pastoris for industrial enzyme production. Microb. Cell Factories 2015, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Cereghino, J.L.; Cregg, J.M. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev. 2000, 24, 45–66. [Google Scholar] [CrossRef]
- Zavec, D.; Gasser, B.; Mattanovich, D. Characterization of methanol utilization negative Pichia pastoris for secreted protein production: New cultivation strategies for current and future applications. Biotechnol. Bioeng. 2020, 117, 1394–1405. [Google Scholar] [CrossRef]
- Chiruvolu, V.; Cregg, J.M.; Meagher, M.M. Recombinant protein production in an alcohol oxidase-defective strain of Pichia pastoris in fedbatch fermentations. Enzym. Microb. Technol. 1997, 21, 277–283. [Google Scholar] [CrossRef]
- Guo, F.; Dai, Z.; Peng, W.; Zhang, S.; Zhou, J.; Ma, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Metabolic engineering of Pichia pastoris for malic acid production from methanol. Biotechnol. Bioeng. 2021, 118, 357–371. [Google Scholar] [CrossRef]
- Liu, T.; Liu, B.; Zhou, H.; Zhang, J. Knockout of the DAS gene increases S-adenosylmethionine production in Komagataella phaffii. Biotechnol. Biotechnol. Equip. 2021, 35, 29–36. [Google Scholar] [CrossRef]
- Inan, M.; Meagher, M.M. Non-repressing carbon sources for alcohol oxidase (AOX1) promoter of Pichia pastoris. J. Biosci. Bioeng. 2001, 92, 585–589. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, W.; Zhou, X.; Bai, P.; Cregg, J.M.; Zhang, Y. Catabolite Repression of Aox in Pichia pastoris Is Dependent on Hexose Transporter PpHxt1 and Pexophagy. Appl. Environ. Microbiol. 2010, 76, 6108–6118. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Mohsin, A.; Chu, J.; Zhuang, Y.; Liu, Y.; Guo, M. Enhanced protein production by sorbitol co-feeding with methanol in recombinant Pichia pastoris strains. Biotechnol. Bioprocess Eng. 2017, 22, 767–773. [Google Scholar] [CrossRef]
- Moreira, L.R.S.; Filho, E.X.F. Insights into the mechanism of enzymatic hydrolysis of xylan. Appl. Microbiol. Biotechnol. 2016, 100, 5205–5214. [Google Scholar] [CrossRef] [PubMed]
- Çalık, P.; Ata, O.; Güneş, H.; Massahi, A.; Boy, E.; Keskin, A.; Öztürk, S.; Zerze, G.H.; Özdamar, T.H. Recombinant protein production in Pichia pastoris under glyceraldehyde-3-phosphate dehydrogenase promoter: From carbon source metabolism to bioreactor operation parameters. Biochem. Eng. J. 2015, 95, 20–36. [Google Scholar] [CrossRef]
- Arias, C.A.D.; Molino, J.V.D.; Marques, D.D.A.V.; Maranhão, A.Q.; Parra, D.A.S.; Junior, A.P.; Converti, A. Influence of carbon source on cell size and production of anti LDL (-) single-chain variable fragment by a recombinant Pichia pastoris strain. Mol. Biol. Rep. 2019, 46, 3257–3264. [Google Scholar] [CrossRef]
- Boehm, T.; Pirie-Shepherd, S.; Trinh, L.-B.; Shiloach, J.; Folkman, J. Disruption of the KEX1 gene in Pichia pastoris allows expression of full-length murine and human endostatin. Yeast 1999, 15, 563–572. [Google Scholar] [CrossRef]
- Arias, C.A.D.; Marques, D.D.A.V.; Malpiedi, L.P.; Maranhão, A.Q.; Parra, D.A.S.; Converti, A.; Junior, A.P. Cultivation of Pichia pastoris carrying the scFv anti LDL (-) antibody fragment. Effect of preculture carbon source. Braz. J. Microbiol. 2017, 48, 419–426. [Google Scholar] [CrossRef]
- Canales, C.; Altamirano, C.; Berrios, J. The growth of Pichia pastoris Mut+ on methanol-glycerol mixtures fits to interactive dual-limited kinetics: Model development and application to optimised fed-batch operation for heterologous protein production. Bioprocess Biosyst. Eng. 2018, 41, 1827–1838. [Google Scholar] [CrossRef]
- Geertman, J.-M.A.; van Maris, A.J.; van Dijken, J.P.; Pronk, J.T. Physiological and genetic engineering of cytosolic redox metabolism in Saccharomyces cerevisiae for improved glycerol production. Metab. Eng. 2006, 8, 532–542. [Google Scholar] [CrossRef]
- Kim, S.; Hahn, J.-S. Efficient production of 2,3-butanediol in Saccharomyces cerevisiae by eliminating ethanol and glycerol production and redox rebalancing. Metab. Eng. 2015, 31, 94–101. [Google Scholar] [CrossRef]
- Kickenweiz, T.; Glieder, A.; Wu, J.C. Construction of a cellulose-metabolizing Komagataella phaffii (Pichia pastoris) by co-expressing glucanases and β-glucosidase. Appl. Microbiol. Biotechnol. 2017, 102, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.R.; Su, X.; Shrestha, S.K.; Yang, S.Y.; Soh, Y. 2E-Decene-4,6-diyn-1-ol-acetate inhibits osteoclastogenesis through mitogen-activated protein kinase-c-Fos-NFATc1 signalling pathways. Clin. Exp. Pharmacol. Physiol. 2021, 49, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Bai, C.; Liu, Y.; Song, L.; Tian, L.; Yan, Y.; Zhou, J.; Zhou, X.; Zhang, Y.; Cai, M. Modulation of acetate utilization in Komagataella phaffii by metabolic engineering of tolerance and metabolism. Biotechnol. Biofuels 2019, 12, 61. [Google Scholar] [CrossRef]
- Gassler, T.; Sauer, M.; Gasser, B.; Egermeier, M.; Troyer, C.; Causon, T.; Hann, S.; Mattanovich, D.; Steiger, M.G. The industrial yeast Pichia pastoris is converted from a heterotroph into an autotroph capable of growth on CO2. Nat. Biotechnol. 2020, 38, 210–216. [Google Scholar] [CrossRef]
- Resina, D.; Bollók, M.; Khatri, N.K.; Valero, F.; Neubauer, P.; Ferrer, P. Transcriptional response of P. pastoris in fed-batch cultivations to Rhizopus oryzae lipase production reveals UPR induction. Microb. Cell Factories 2007, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Glieder, A. Regulation of Pichia pastoris promoters and its consequences for protein production. New Biotechnol. 2012, 30, 385–404. [Google Scholar] [CrossRef] [PubMed]
- Ergün, B.G.; Demir, I.; Özdamar, T.H.; Gasser, B.; Mattanovich, D.; Çalık, P. Engineered Deregulation of Expression in Yeast with Designed Hybrid-Promoter Architectures in Coordination with Discovered Master Regulator Transcription Factor. Adv. Biosyst. 2020, 4, e1900172. [Google Scholar] [CrossRef]
- Gasser, B.; Sauer, M.; Maurer, M.; Stadlmayr, G.; Mattanovich, D. Transcriptomics-Based Identification of Novel Factors Enhancing Heterologous Protein Secretion in Yeasts. Appl. Environ. Microbiol. 2007, 73, 6499–6507. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, Y.; Miao, H.; Tang, X.; Xu, B.; Wu, Q.; Mu, Y.; Huang, Z. Transcriptomic Analysis of Pichia pastoris (Komagataella phaffii) GS115 During Heterologous Protein Production Using a High-Cell-Density Fed-Batch Cultivation Strategy. Front. Microbiol. 2020, 11, 463. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Zhan, C.; Zhang, Z.; Liu, X.; Liu, H.; Bai, Z. Transcriptional analysis of impacts of glycerol transporter 1 on methanol and glycerol metabolism in Pichia pastoris. FEMS Yeast Res. 2017, 18, 18. [Google Scholar] [CrossRef]
- Ata, Ö.; Rebnegger, C.; Tatto, N.E.; Valli, M.; Mairinger, T.; Hann, S.; Steiger, M.G.; Çalık, P.; Mattanovich, D. A single Gal4-like transcription factor activates the Crabtree effect in Komagataella phaffii. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kalender, Ö.; Çalık, P. Transcriptional regulatory proteins in central carbon metabolism of Pichia pastoris and Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2020, 104, 7273–7311. [Google Scholar] [CrossRef] [PubMed]
- Mitsis, T.; Efthimiadou, A.; Bacopoulou, F.; Vlachakis, D.; Chrousos, G.; Eliopoulos, E. Transcription factors and evolution: An integral part of gene expression (Review). World Acad. Sci. J. 2020. [Google Scholar] [CrossRef]
- Bankefa, O.E.; Wang, M.; Zhu, T.; Li, Y. Hac1p homologues from higher eukaryotes can improve the secretion of heterologous proteins in the yeast Pichia pastoris. Biotechnol. Lett. 2018, 40, 1149–1156. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, Y.; Wang, P.; Li, X.; Zhan, C.; Linhardt, R.J.; Zhang, F.; Liu, X.; Zhan, J.; Bai, Z. Characterization and application of a putative transcription factor (SUT2) in Pichia pastoris. Mol. Genet. Genom. 2020, 295, 1295–1304. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, Y.; Zhang, X.; Li, C.; Liu, X.; Lin, Y.; Liang, S. Fhl1p protein, a positive transcription factor in Pichia pastoris, enhances the expression of recombinant proteins. Microb. Cell Factories 2019, 18, 1–12. [Google Scholar] [CrossRef]
- Chang, C.-H.; Hsiung, H.-A.; Hong, K.-L.; Huang, C.-T. Enhancing the efficiency of the Pichia pastoris AOX1 promoter via the synthetic positive feedback circuit of transcription factor Mxr1. BMC Biotechnol. 2018, 18, 81. [Google Scholar] [CrossRef]
- Jiang, B.; Argyros, R.; Bukowski, J.; Nelson, S.; Sharkey, N.; Kim, S.; Copeland, V.; Davidson, R.C.; Chen, R.; Zhuang, J.; et al. Inactivation of a GAL4 -Like Transcription Factor Improves Cell Fitness and Product Yield in Glycoengineered Pichia pastoris Strains. Appl. Environ. Microbiol. 2015, 81, 260–271. [Google Scholar] [CrossRef]
- Delic, M.; Graf, A.B.; Koellensperger, G.; Troyer, C.; Hann, S.; Mattanovich, D.; Gasser, B. Overexpression of the transcription factor Yap1 modifies intracellular redox conditions and enhances recombinant protein secretion. Microb. Cell 2014, 1, 376–386. [Google Scholar] [CrossRef]
- Ruth, C.; Buchetics, M.; Vidimce, V.; Kotz, D.; Naschberger, S.; Mattanovich, D.; Pichler, H.; Gasser, B. Pichia pastoris Aft1—A novel transcription factor, enhancing recombinant protein secretion. Microb. Cell Factories 2014, 13, 1–15. [Google Scholar] [CrossRef]
- Sun, J.; Jiang, J.; Zhai, X.; Zhu, S.; Qu, Z.; Yuan, W.; Wang, Z.; Wei, C. Coexpression of Kex2 Endoproteinase and Hac1 Transcription Factor to Improve the Secretory Expression of Bovine Lactoferrin in Pichia pastoris. Biotechnol. Bioprocess Eng. 2019, 24, 934–941. [Google Scholar] [CrossRef]
- Guerfal, M.; Ryckaert, S.; Jacobs, P.P.; Ameloot, P.; Van Craenenbroeck, K.; Derycke, R.; Callewaert, N. The HAC1 gene from Pichia pastoris: Characterization and effect of its overexpression on the production of secreted, surface displayed and membrane proteins. Microb. Cell Factories 2010, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Vogl, T.; Thallinger, G.G.; Zellnig, G.; Drew, D.; Cregg, J.M.; Glieder, A.; Freigassner, M. Towards improved membrane protein production in Pichia pastoris: General and specific transcriptional response to membrane protein overexpression. New Biotechnol. 2014, 31, 538–552. [Google Scholar] [CrossRef]
- Liu, J.; Han, Q.; Cheng, Q.; Chen, Y.; Wang, R.; Li, X.; Liu, Y.; Yan, D. Efficient Expression of Human Lysozyme Through the Increased Gene Dosage and Co-expression of Transcription Factor Hac1p in Pichia pastoris. Curr. Microbiol. 2020, 77, 846–854. [Google Scholar] [CrossRef] [PubMed]
- De Waele, S.; Vandenberghe, I.; Laukens, B.; Planckaert, S.; Verweire, S.; Van Bogaert, I.; Soetaert, W.; Devreese, B.; Ciesielska, K. Optimized expression of the Starmerella bombicola lactone esterase in Pichia pastoris through temperature adaptation, codon-optimization and co-expression with HAC1. Protein Expr. Purif. 2018, 143, 62–70. [Google Scholar] [CrossRef]
- Han, M.; Wang, W.; Zhou, J.; Gong, X.; Xu, C.; Li, Y.; Li, Q. Activation of the Unfolded Protein Response via Co-expression of the HAC1i Gene Enhances Expression of Recombinant Elastase in Pichia pastoris. Biotechnol. Bioprocess Eng. 2020, 25, 302–307. [Google Scholar] [CrossRef]
- Yano, T.; Yurimoto, H.; Sakai, Y. Activation of the Oxidative Stress Regulator PpYap1 through Conserved Cysteine Residues during Methanol Metabolism in the Yeast Pichia pastoris. Biosci. Biotechnol. Biochem. 2009, 73, 1404–1411. [Google Scholar] [CrossRef]
- Yano, T.; Takigami, E.; Yurimoto, H.; Sakai, Y. Yap1-Regulated Glutathione Redox System Curtails Accumulation of Formaldehyde and Reactive Oxygen Species in Methanol Metabolism of Pichia pastoris. Eukaryot. Cell 2009, 8, 540–549. [Google Scholar] [CrossRef]
- Lin, X.-Q.; Liang, S.-L.; Han, S.-Y.; Zheng, S.-P.; Ye, Y.-R.; Lin, Y. Quantitative iTRAQ LC–MS/MS proteomics reveals the cellular response to heterologous protein overexpression and the regulation of HAC1 in Pichia pastoris. J. Proteom. 2013, 91, 58–72. [Google Scholar] [CrossRef]
- Li, C.; Lin, Y.; Zheng, X.; Pang, N.; Liao, X.; Liu, X.; Huang, Y.; Liang, S. Combined strategies for improving expression of Citrobacter amalonaticus phytase in Pichia pastoris. BMC Biotechnol. 2015, 15, 1–11. [Google Scholar] [CrossRef]
- Hohenblum, H.; Gasser, B.; Maurer, M.; Borth, N.; Mattanovich, D. Effects of gene dosage, promoters, and substrates on unfolded protein stress of recombinant Pichia pastoris. Biotechnol. Bioeng. 2004, 85, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Miao, L.; Huang, H.; Li, Y.; Zhu, T. High-level production of glucose oxidase in Pichia pastoris: Effects of Hac1p overexpression on cell physiology and enzyme expression. Enzym. Microb. Technol. 2020, 141, 109671. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Gao, Y.; Zhou, X.; Zhang, Y.; Cai, M. Regulating unfolded protein response activator HAC1p for production of thermostable raw-starch hydrolyzing α-amylase in Pichia pastoris. Bioprocess Biosyst. Eng. 2016, 40, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Haberhauer-Troyer, C.; Delic, M.; Gasser, B.; Mattanovich, D.; Hann, S.; Koellensperger, G. Accurate quantification of the redox-sensitive GSH/GSSG ratios in the yeast Pichia pastoris by HILIC–MS/MS. Anal. Bioanal. Chem. 2013, 405, 2031–2039. [Google Scholar] [CrossRef]
- Pekarsky, A.; Veiter, L.; Rajamanickam, V.; Herwig, C.; Grünwald-Gruber, C.; Altmann, F.; Spadiut, O. Production of a recombinant peroxidase in different glyco-engineered Pichia pastoris strains: A morphological and physiological comparison. Microb. Cell Factories 2018, 17, 1–15. [Google Scholar] [CrossRef]
- Barnard, G.C.; Kull, A.R.; Sharkey, N.S.; Shaikh, S.S.; Rittenhour, A.M.; Burnina, I.; Jiang, Y.; Li, F.; Lynaugh, H.; Mitchell, T.; et al. High-throughput screening and selection of yeast cell lines expressing monoclonal antibodies. J. Ind. Microbiol. Biotechnol. 2010, 37, 961–971. [Google Scholar] [CrossRef]
- Zha, D. Glycoengineered Pichia-Based Expression of Monoclonal Antibodies. Methods Mol. Biol. 2013, 988, 31–43. [Google Scholar] [CrossRef]
- Ata, Ö.; Prielhofer, R.; Gasser, B.; Mattanovich, D.; Çalık, P. Transcriptional engineering of the glyceraldehyde-3-phosphate dehydrogenase promoter for improved heterologous protein production in Pichia pastoris. Biotechnol. Bioeng. 2017, 114, 2319–2327. [Google Scholar] [CrossRef]
- Vogl, T.; Sturmberger, L.; Fauland, P.C.; Hyden, P.; Fischer, J.E.; Schmid, C.; Thallinger, G.G.; Geier, M.; Glieder, A. Methanol independent induction in Pichia pastoris by simple derepressed overexpression of single transcription factors. Biotechnol. Bioeng. 2018, 115, 1037–1050. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Shi, L.; Qi, F.; Zhang, P.; Zhang, Y.; Zhou, X.; Song, Z.; Cai, M. Methanol-Independent Protein Expression by AOX1 Promoter with trans-Acting Elements Engineering and Glucose-Glycerol-Shift Induction in Pichia pastoris. Sci. Rep. 2017, 7, srep41850. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.; Chen, M.; Yang, M.; Li, L.; Mou, H. Enhancing the expression of recombinant κ-carrageenase in Pichia pastoris using dual promoters, co-expressing chaperones and transcription factors. Biocatal. Biotransformation 2019, 38, 104–113. [Google Scholar] [CrossRef]
- Shirozu, R.; Yashiroda, H.; Murata, S. Identification of minimum Rpn4-responsive elements in genes related to proteasome functions. FEBS Lett. 2015, 589, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Barbay, D.; Mačáková, M.; Sützl, L.; De, S.; Mattanovich, D.; Gasser, B. Two homologs of the Cat8 transcription factor are involved in the regulation of ethanol utilization in Komagataella phaffii. Curr. Genet. 2021, 67, 641–661. [Google Scholar] [CrossRef]
- Delic, M.; Valli, M.; Graf, A.B.; Pfeffer, M.; Mattanovich, D.; Gasser, B. The secretory pathway: Exploring yeast diversity. FEMS Microbiol. Rev. 2013, 37, 872–914. [Google Scholar] [CrossRef] [PubMed]
- Delic, M.; Göngrich, R.; Mattanovich, D.; Gasser, B. Engineering of Protein Folding and Secretion—Strategies to Overcome Bottlenecks for Efficient Production of Recombinant Proteins. Antioxidants Redox Signal. 2014, 21, 414–437. [Google Scholar] [CrossRef] [PubMed]
- Karaoğlan, M.; Erden-Karaoğlan, F. Effect of codon optimization and promoter choice on recombinant endo-polygalacturonase production in Pichia pastoris. Enzym. Microb. Technol. 2020, 139, 109589. [Google Scholar] [CrossRef]
- Che, Z.; Cao, X.; Chen, G.; Liang, Z. An effective combination of codon optimization, gene dosage, and process optimization for high-level production of fibrinolytic enzyme in Komagataella phaffii (Pichia pastoris). BMC Biotechnol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Sallada, N.D.; Harkins, L.E.; Berger, B.W. Effect of gene copy number and chaperone coexpression on recombinant hydrophobin HFBI biosurfactant production in Pichia pastoris. Biotechnol. Bioeng. 2019, 116, 2029–2040. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, T.; Lu, L.; Cai, F.; Lin, J.; Jiang, Y.; Lin, Y. Codon pair optimization (CPO): A software tool for synthetic gene design based on codon pair bias to improve the expression of recombinant proteins in Pichia pastoris. Microb. Cell Factories 2021, 20, 1–10. [Google Scholar] [CrossRef]
- Lan, D.; Qu, M.; Yang, B.; Wang, Y. Enhancing production of lipase MAS1 from marine Streptomyces sp. strain in Pichia pastoris by chaperones co-expression. Electron. J. Biotechnol. 2016, 22, 62–67. [Google Scholar] [CrossRef]
- Guan, B.; Chen, F.; Su, S.; Duan, Z.; Chen, Y.; Li, H.; Jin, J. Effects of co-overexpression of secretion helper factors on the secretion of a HSA fusion protein (IL2-HSA) inpichia pastoris. Yeast 2016, 33, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.; Vanz, A.L.; Lünsdorf, H.; Nimtz, M.; Rinas, U. Fate of the UPR marker protein Kar2/Bip and autophagic processes in fed-batch cultures of secretory insulin precursor producing Pichia pastoris. Microb. Cell Factories 2018, 17, 123. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Ding, L.; Wei, D.; Zhou, H.; Chu, J.; Zhang, S.; Qian, J. Screening endogenous signal peptides and protein folding factors to promote the secretory expression of heterologous proteins in Pichia pastoris. J. Biotechnol. 2019, 306, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Ben Azoun, S.; Ben Zakour, M.; Sghaier, S.; Kallel, H. Expression of rabies virus glycoprotein in the methylotrophic yeast Pichia pastoris. Biotechnol. Appl. Biochem. 2017, 64, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Samuel, P.; Vadhana, A.K.P.; Kamatchi, R.; Antony, A.; Meenakshisundaram, S. Effect of molecular chaperones on the expression of Candida antarctica lipase B in Pichia pastoris. Microbiol. Res. 2013, 168, 615–620. [Google Scholar] [CrossRef]
- Ben Azoun, S.; Belhaj, A.E.; Göngrich, R.; Gasser, B.; Kallel, H. Molecular optimization of rabies virus glycoprotein expression in Pichia pastoris. Microb. Biotechnol. 2016, 9, 355–368. [Google Scholar] [CrossRef]
- Gasser, B.; Maurer, M.; Gach, J.; Kunert, R.; Mattanovich, D. Engineering of Pichia pastoris for improved production of antibody fragments. Biotechnol. Bioeng. 2006, 94, 353–361. [Google Scholar] [CrossRef]
- Inan, M.; Aryasomayajula, D.; Sinha, J.; Meagher, M.M. Enhancement of protein secretion in Pichia pastoris by overexpression of protein disulfide isomerase. Biotechnol. Bioeng. 2006, 93, 771–778. [Google Scholar] [CrossRef]
- Damasceno, L.M.; Anderson, K.A.; Ritter, G.; Cregg, J.M.; Old, L.J.; Batt, C.A. Cooverexpression of chaperones for enhanced secretion of a single-chain antibody fragment in Pichia pastoris. Appl. Microbiol. Biotechnol. 2007, 74, 381–389. [Google Scholar] [CrossRef]
- Gasser, B.; Prielhofer, R.; Marx, H.; Maurer, M.; Nocon, J.; Steiger, M.; Puxbaum, V.; Sauer, M.; Mattanovich, D. Pichia pastoris: Protein production host and model organism for biomedical research. Futur. Microbiol. 2013, 8, 191–208. [Google Scholar] [CrossRef]
- Spohner, S.; Müller, H.; Quitmann, H.; Czermak, P. Expression of enzymes for the usage in food and feed industry with Pichia pastoris. J. Biotechnol. 2015, 202, 118–134. [Google Scholar] [CrossRef] [PubMed]
- Saitua, F.; Torres, P.; Pérez-Correa, J.R.; Agosin, E. Dynamic genome-scale metabolic modeling of the yeast Pichia pastoris. BMC Syst. Biol. 2017, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.K.; Selvarasu, S.; Camattari, A.; Ryu, J.; Lee, H.; Ahn, J.; Lee, H.; Lee, D.-Y. Genome-scale metabolic reconstruction and in silico analysis of methylotrophic yeast Pichia pastoris for strain improvement. Microb. Cell Factories 2010, 9, 50. [Google Scholar] [CrossRef]
- Tomàs-Gamisans, M.; Ferrer, P.; Albiol, J. Integration and Validation of the Genome-Scale Metabolic Models of Pichia pastoris: A Comprehensive Update of Protein Glycosylation Pathways, Lipid and Energy Metabolism. PLoS ONE 2016, 11, e0148031. [Google Scholar] [CrossRef]
- Tomàs-Gamisans, M.; Ferrer, P.; Albiol, J. Fine-tuning the P. pastoris iMT1026 genome-scale metabolic model for improved prediction of growth on methanol or glycerol as sole carbon sources. Microb. Biotechnol. 2018, 11, 224–237. [Google Scholar] [CrossRef]
- Canales, C.; Altamirano, C.; Berrios, J. Effect of dilution rate and methanol-glycerol mixed feeding on heterologous Rhizopus oryzae lipase production with Pichia pastoris Mut+ phenotype in continuous culture. Biotechnol. Prog. 2015, 31, 707–714. [Google Scholar] [CrossRef]
- Kastberg, L.L.B.; Ard, R.; Jensen, M.K.; Workman, C.T. Burden Imposed by Heterologous Protein Production in Two Major Industrial Yeast Cell Factories: Identifying Sources and Mitigation Strategies. Front. Fungal Biol. 2022, 3, 1. [Google Scholar] [CrossRef]
- Torres, P.; Saa, P.A.; Albiol, J.; Ferrer, P.; Agosin, E. Contextualized genome-scale model unveils high-order metabolic effects of the specific growth rate and oxygenation level in recombinant Pichia pastoris. Metab. Eng. Commun. 2019, 9, e00103. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.; Huang, Z.; Qian, M.; Zhang, Q.; Feng, J. Process development of recombinant Aspergillus flavus urate oxidase production in Pichia pastoris intracellularly and its characterization as a potential biosimilar. Process Biochem. 2021, 102, 376–385. [Google Scholar] [CrossRef]


| Auxiliary Gene | Modification | Pathways Involved | Heterologous Product | Production (Fold Change) | Operation Mode | Scale | Strain | Ref. |
|---|---|---|---|---|---|---|---|---|
| A. Engineering of the MUT pathway | ||||||||
| PAOXDAS | OE | MUT | HRP (S) | +0.94 | Batch | 5 L Bioreactor | CBS7435 (MUTs) | [37] |
| CalB (S) | +0.56 | |||||||
| PAOXFLD | OE | MUT | HRP (S) | +1.21 | ||||
| CalB (S) | +1.07 | |||||||
| DAS1, DAS2 DAS1/2 | Del | MUT | eGFP | +1.3 +1.2 +1.15 | Batch | Flask | CBS7435 (MUTs) | [38] |
| PFLD1ADH2 | OE | MUT | cFab-vHH (S) | +2.69 | Fed-batch | 1 L Bioreactor | CBS2612 (MUT−) | [39] |
| FDH | Del | MUT | β-galactosidase | +2 | Batch | Flask | GS115 (MUT+) | [40] |
| B. Engineering of co-substrate catabolic pathways | ||||||||
| ATG30 | Del | MUT | Xylanase (S) | +3 | Batch | Flask | GS115 (MUT+) | [41] |
| C. Engineering of co-factor metabolism | ||||||||
| PGAPPOS5 | OE | PPP | Fab (S) | +2 | Chemostat | 2 L Bioreactor | X-33 (MUT+) | [27] |
| PGAPZWF1-PGAP SOL3 | OE | PPP | hSOD | +3.8 | Batch | Flask | SMD1168H | [42] |
| PAOXSOL3-PAOXGND2 | OE | PPP | hIFN-γ (S) | +2.2 | Fed- batch | 1 L Bioreactor | GS115 (MUT+) | [43] |
| D. Synthetic biology for alternative carbon source catabolism | ||||||||
| PGAPXI | Ex | Xylose path. | β-mannanase (S) | +1.36 | Batch | Flask | GS115 (MUT+) | [44] |
| Auxiliary Gene | Modification | Pathways Involved | Heterologous Product | Production (Fold Change) | Operation Mode | Scale | Strain | Ref. |
|---|---|---|---|---|---|---|---|---|
| PSUT2-MXR1 | OE | MUT and sterol biosynthesis | eGFP | +1.18 | Batch | Flask | GS115 | [75] |
| FHL1p | OE | Ribosome biosynthesis, processing | Phytase (S) Pectinase (S) mRFP | +1.2 +1.35 +1.31 | Batch | Flask | GS115 | [76] |
| PAOX2-MXR1 | OE | MUT | scFv | +2.7 | Batch | Flask | KM71 | [77] |
| ATT1 | Del | Cellular fitness modulation | IgG1 (anti-HER2) | +1.5 | Fed-batch | 15 L Bioreactor | Gly. Eng. | [78] |
| YAP1 | OE | Oxidative stress response | Trypsinogen (S) | +2.0 | Batch | Flask | X-33 | [79] |
| AFT1 | OE | Secretion and carbohydrate metabolism | Carboxylesterase (S) | +2.5 | Fed-batch | 1 L Bioreactor | CBS7435 | [80] |
| HAC1 | OE | UPR | Bovine lactoferrin (S) | +5.0 | Fed-batch | 5 L Bioreactor | GS115 | [81] |
| HAC1 | OE | UPR | Thrombomodulin Adenosine A2A mIL-10 (S) Trans-sialidase (S) | +1.9 +1.18 +2.2 +2.1 | Batch | Flask | GS115 | [82] |
| HAC1 | OE | UPR | HsCstp HsCtr1p OsCstp | +2.1 +1.7 +1.5 | Fed-batch | 1 L Bioreactor | CBS7435 | [83] |
| HAC1 | OE | UPR | Lysozyme (HYL) (S) | +2.13 | Batch | Flask | KM71H | [84] |
| HAC1 | OE | UPR | Lactone esterase (S) | +1.8 | Batch | Flask | NRRL-Y-11430 | [85] |
| HAC1 | OE | UPR | elastase (S) | +1.8 | Batch | Flask | GS115 | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustos, C.; Quezada, J.; Veas, R.; Altamirano, C.; Braun-Galleani, S.; Fickers, P.; Berrios, J. Advances in Cell Engineering of the Komagataella phaffii Platform for Recombinant Protein Production. Metabolites 2022, 12, 346. https://doi.org/10.3390/metabo12040346
Bustos C, Quezada J, Veas R, Altamirano C, Braun-Galleani S, Fickers P, Berrios J. Advances in Cell Engineering of the Komagataella phaffii Platform for Recombinant Protein Production. Metabolites. 2022; 12(4):346. https://doi.org/10.3390/metabo12040346
Chicago/Turabian StyleBustos, Cristina, Johan Quezada, Rhonda Veas, Claudia Altamirano, Stephanie Braun-Galleani, Patrick Fickers, and Julio Berrios. 2022. "Advances in Cell Engineering of the Komagataella phaffii Platform for Recombinant Protein Production" Metabolites 12, no. 4: 346. https://doi.org/10.3390/metabo12040346
APA StyleBustos, C., Quezada, J., Veas, R., Altamirano, C., Braun-Galleani, S., Fickers, P., & Berrios, J. (2022). Advances in Cell Engineering of the Komagataella phaffii Platform for Recombinant Protein Production. Metabolites, 12(4), 346. https://doi.org/10.3390/metabo12040346