
Modulation of Fecal Metabolites by Heat Stress and Diet, and Their Association with Inflammation and Leaky Gut Markers in Dairy Cows


Abstract
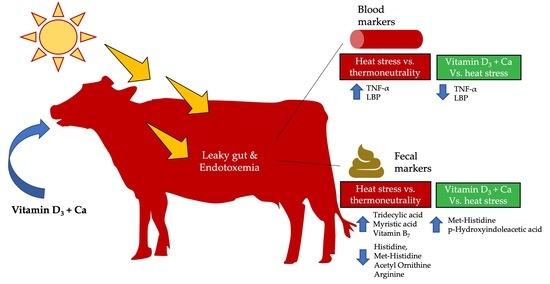
:
1. Introduction
2. Results
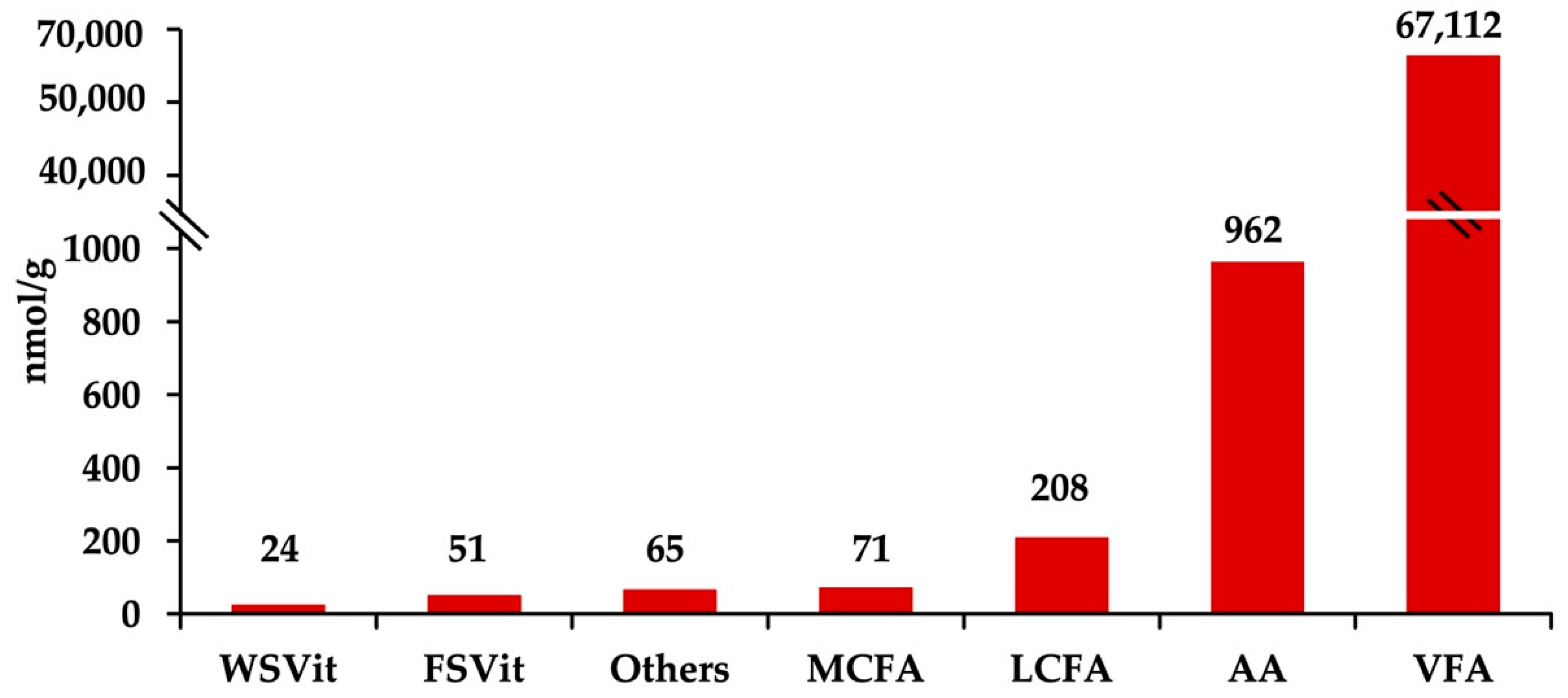
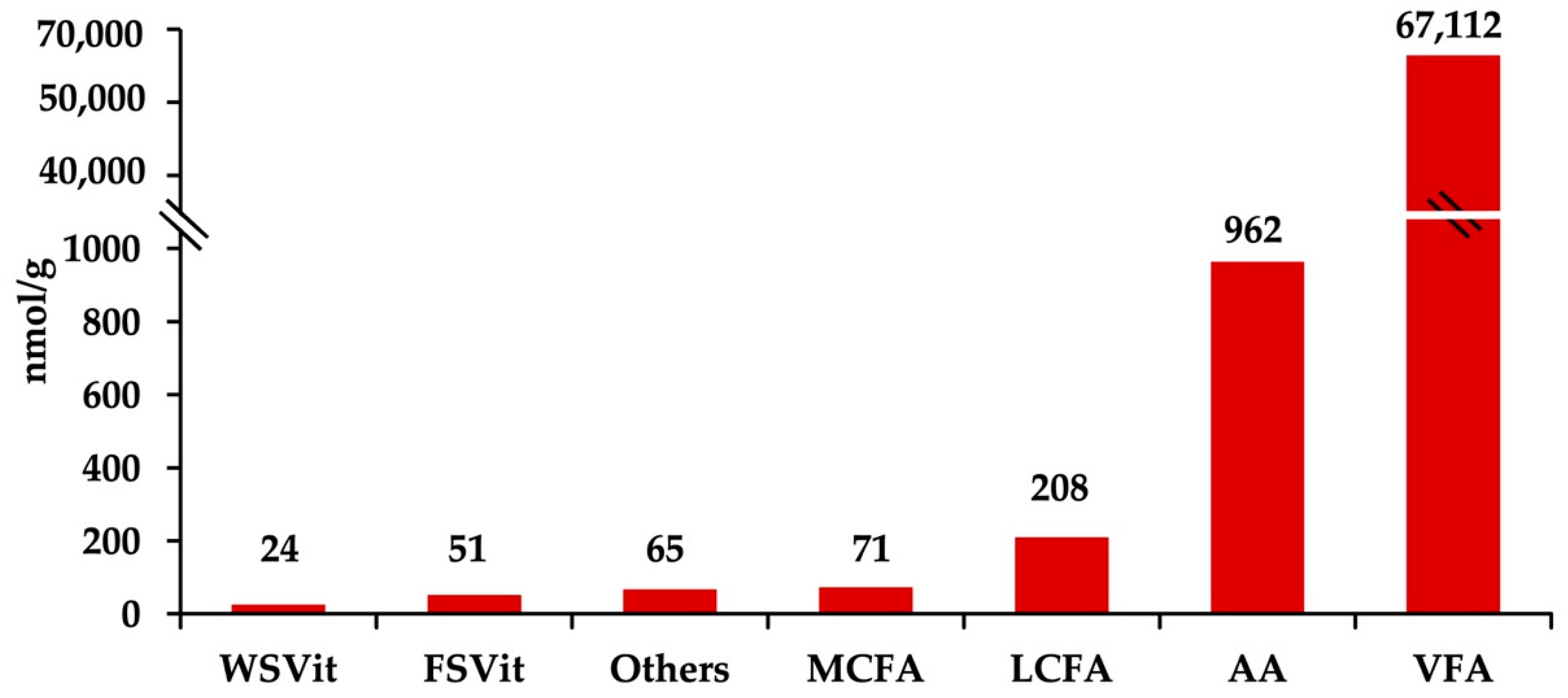
2.1. Fecal Metabolites Concentration
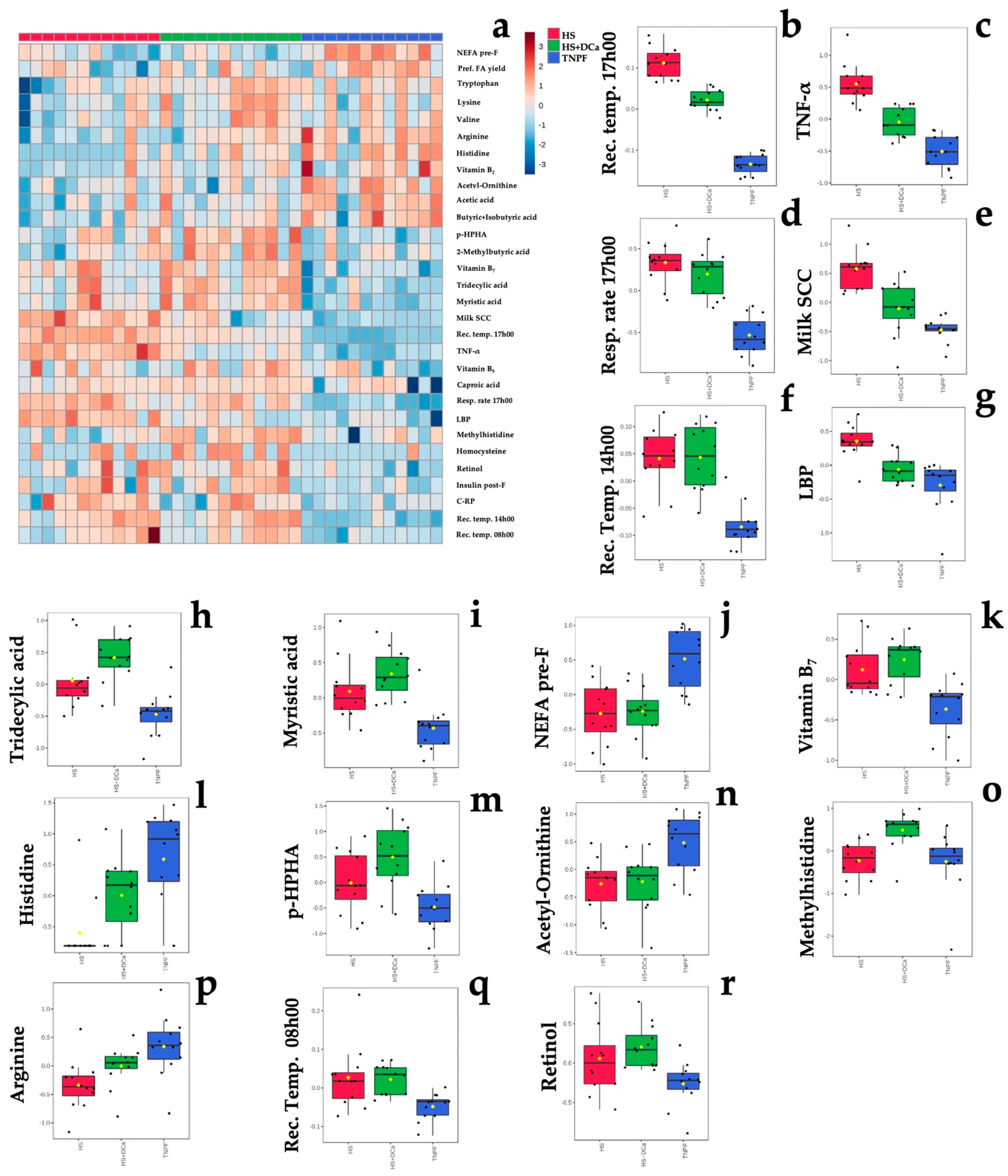
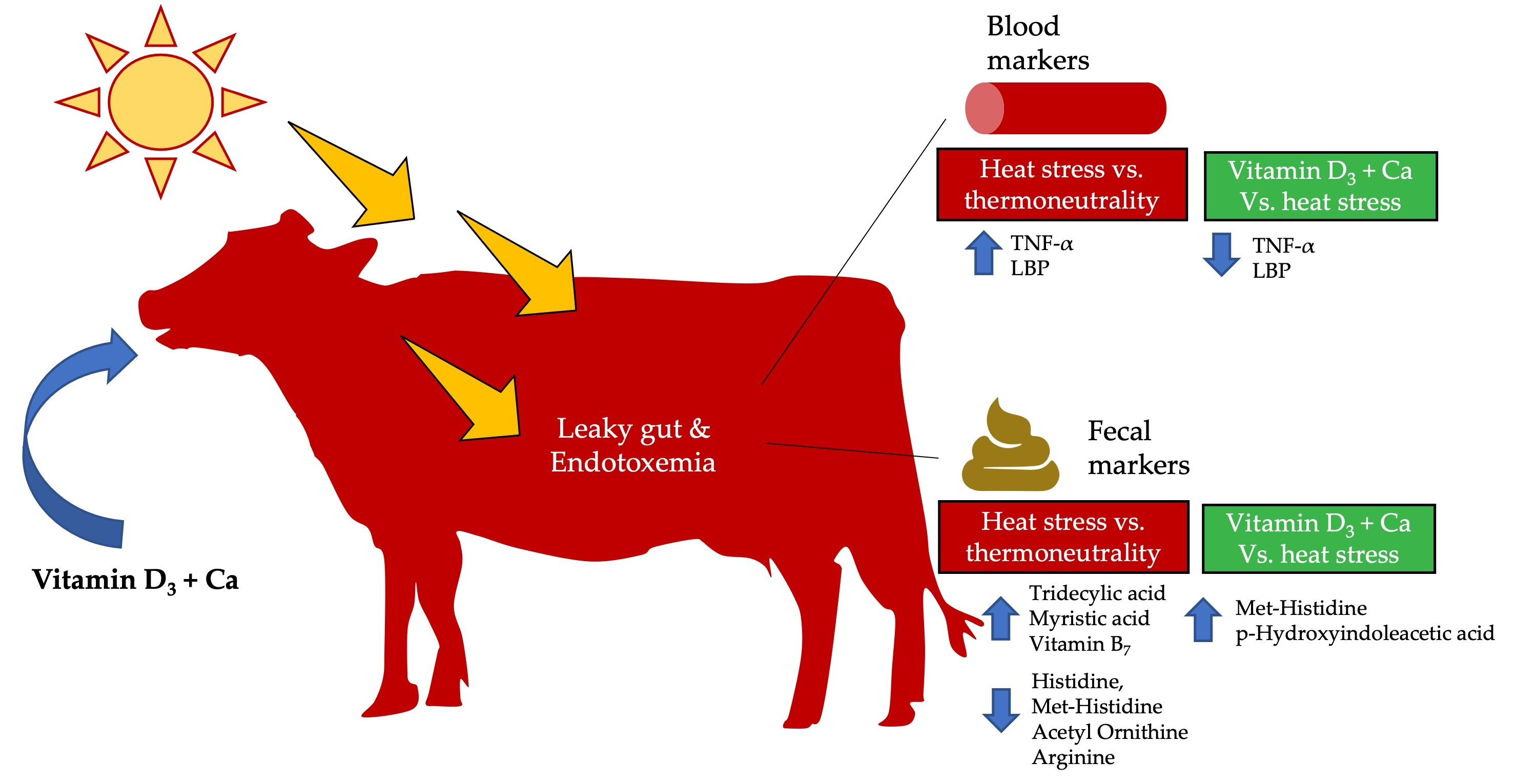
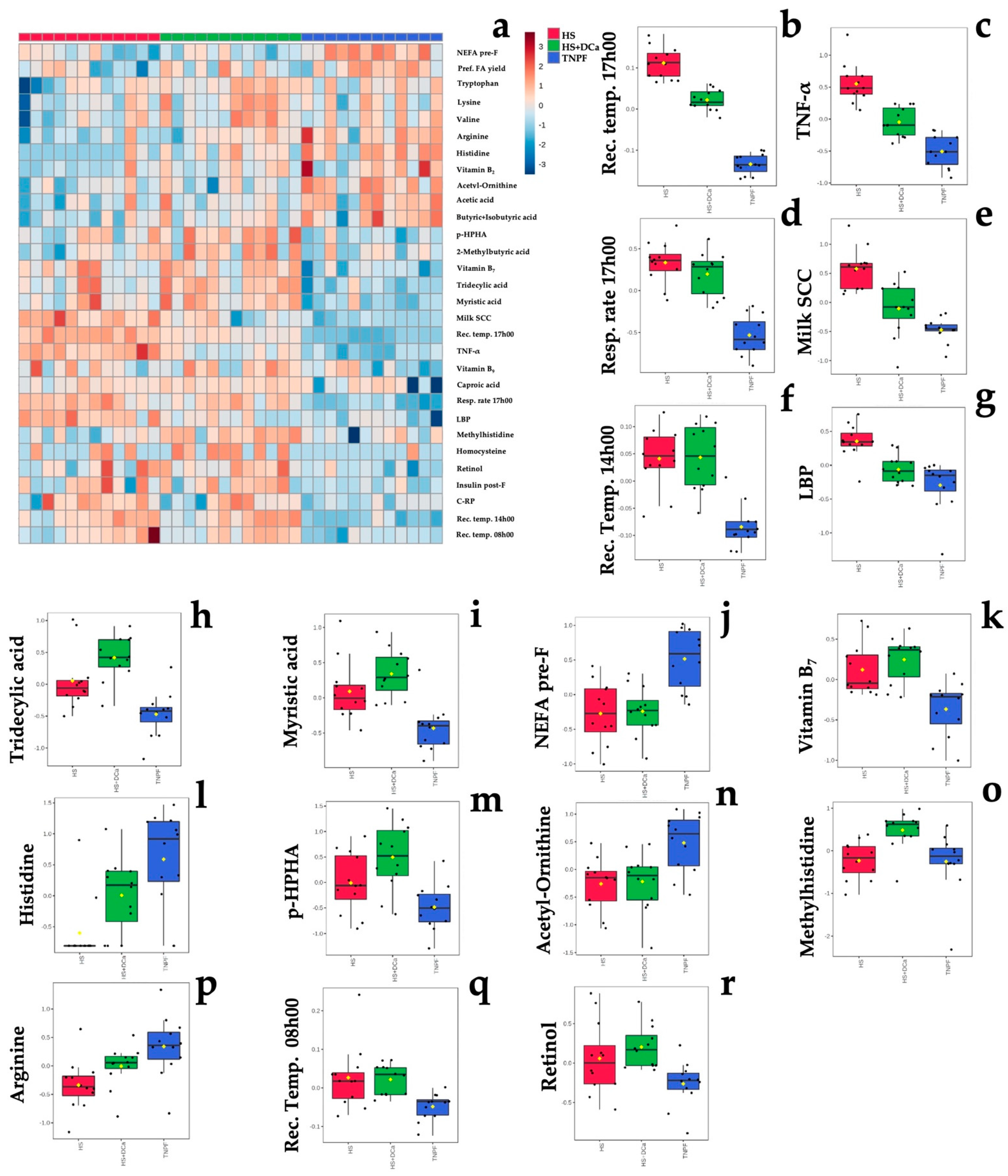
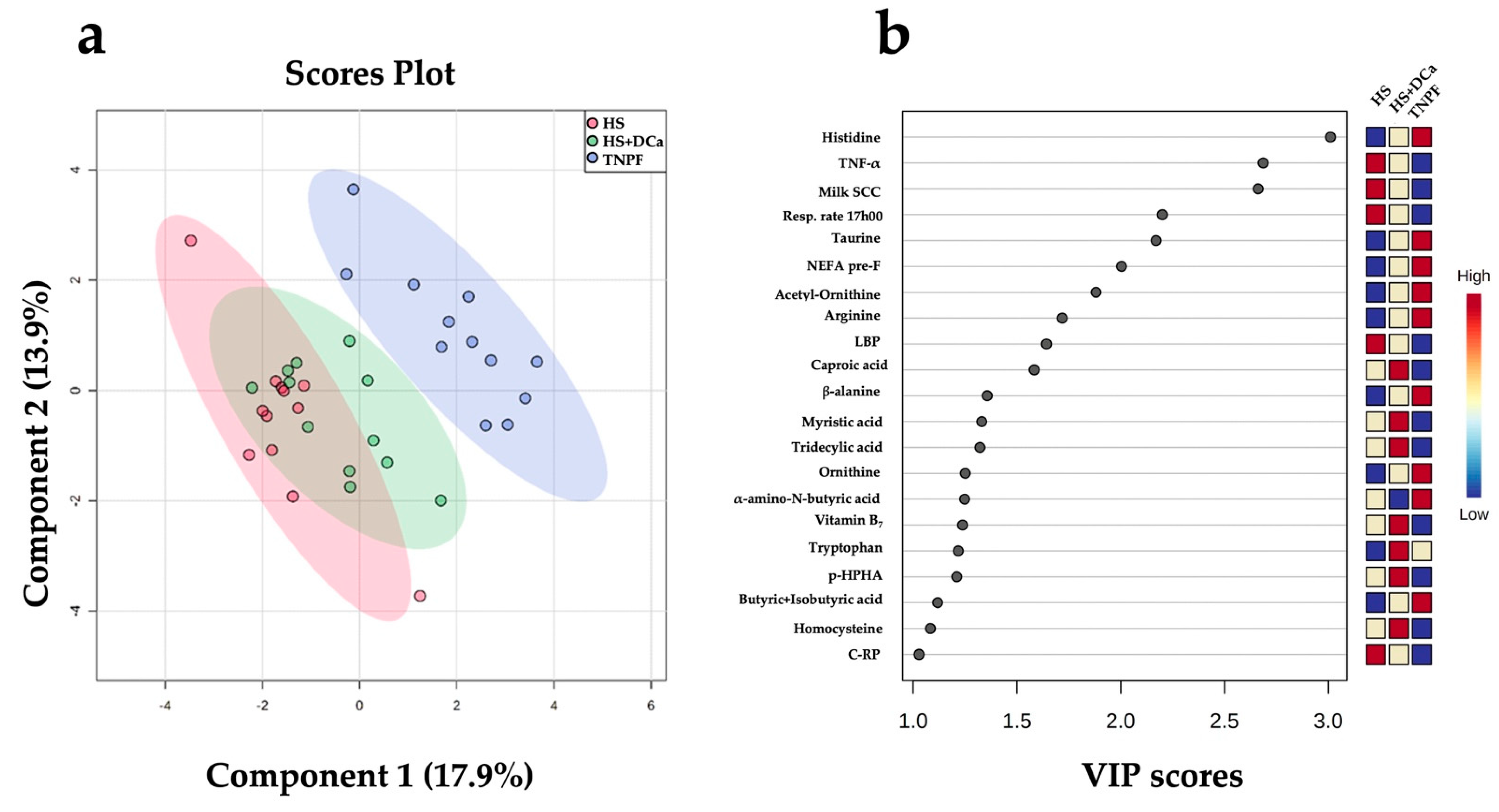
2.2. Effects of Heat Stress and Vitamin D3 and Ca Supplementation during Heat Stress on Fecal Metabolites and Physiological Parameters
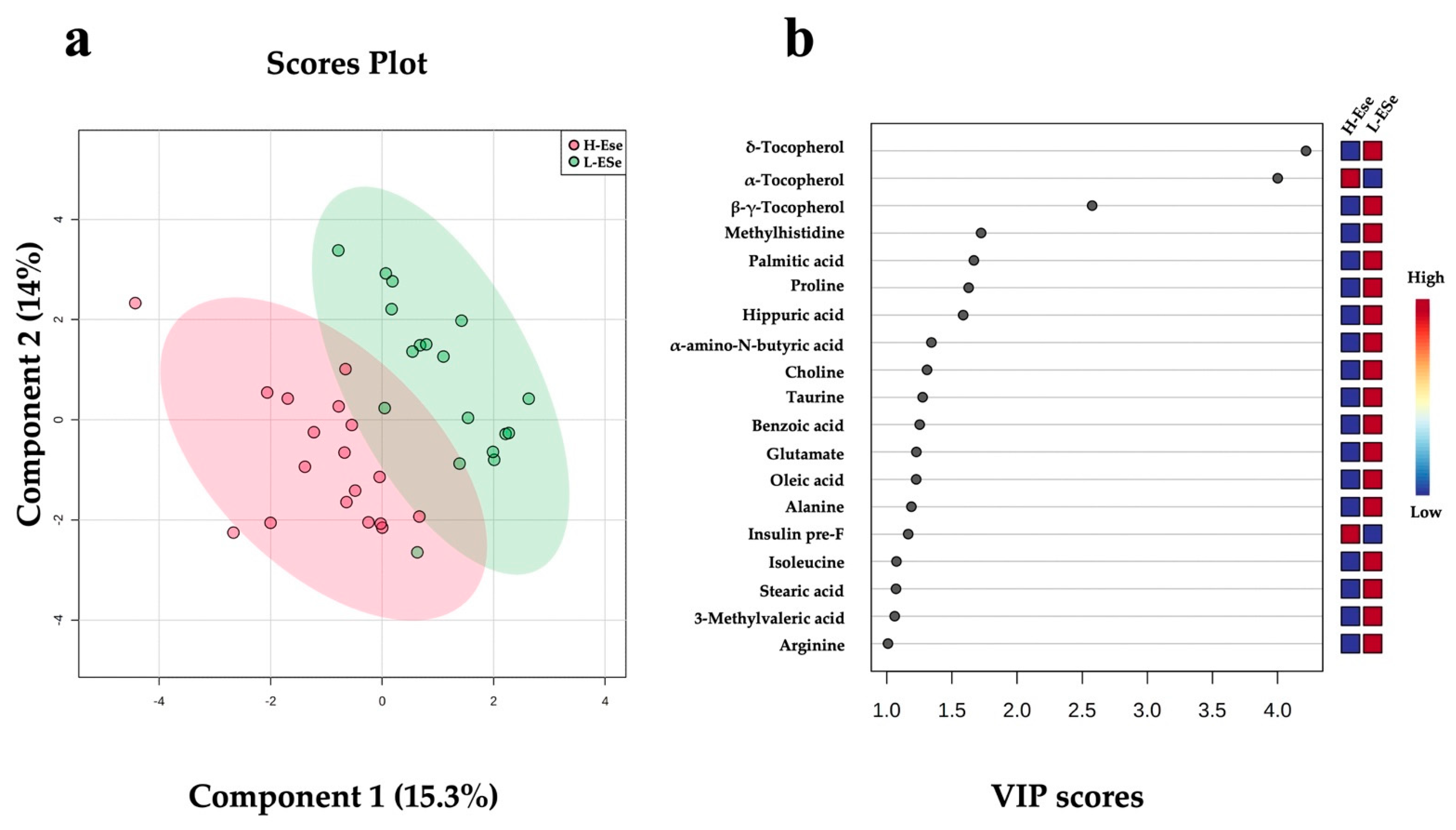
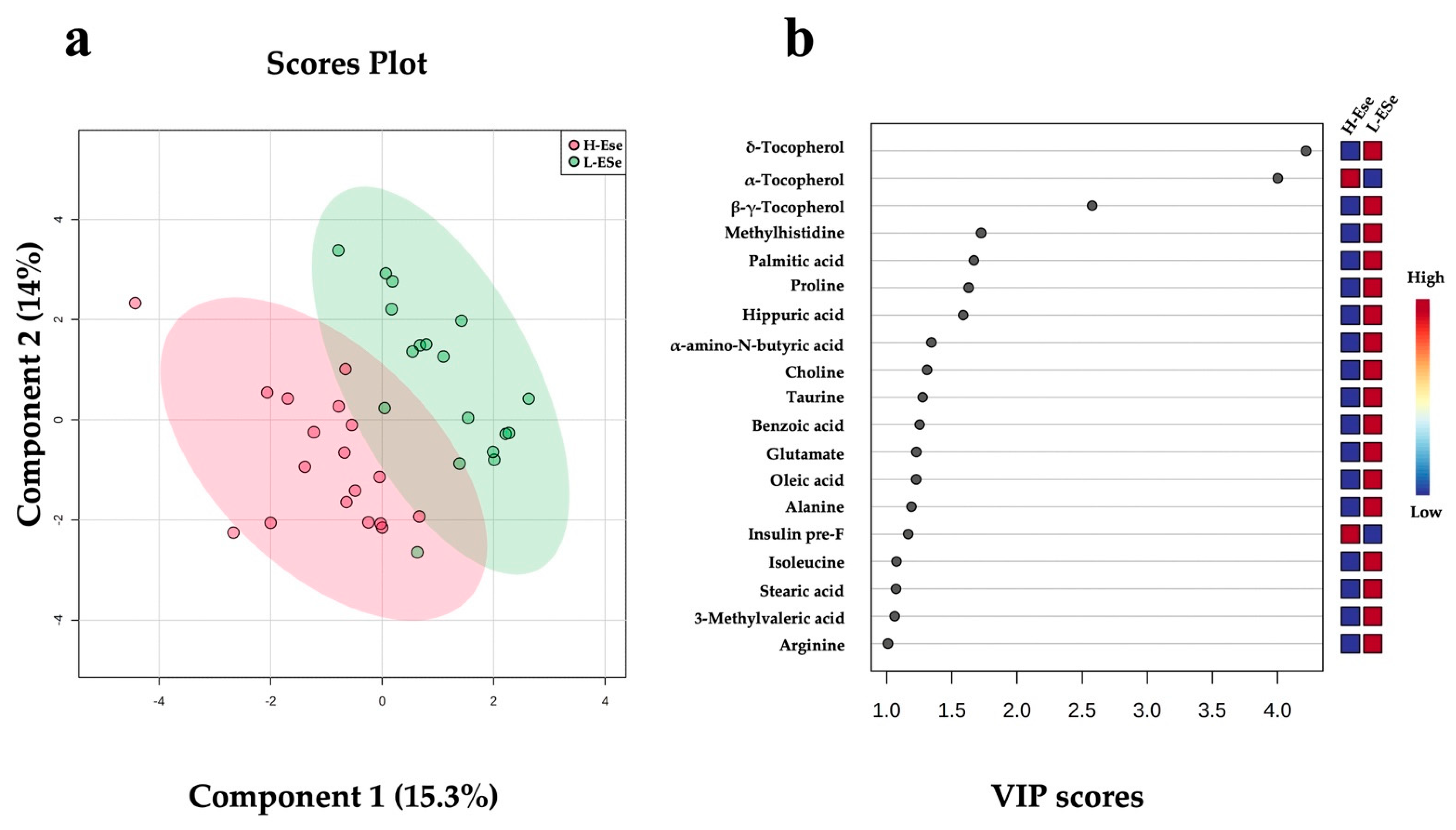
2.3. Changes in Fecal Metabolites Related to the Level of Vitamin E and Selenium of Dairy Cows
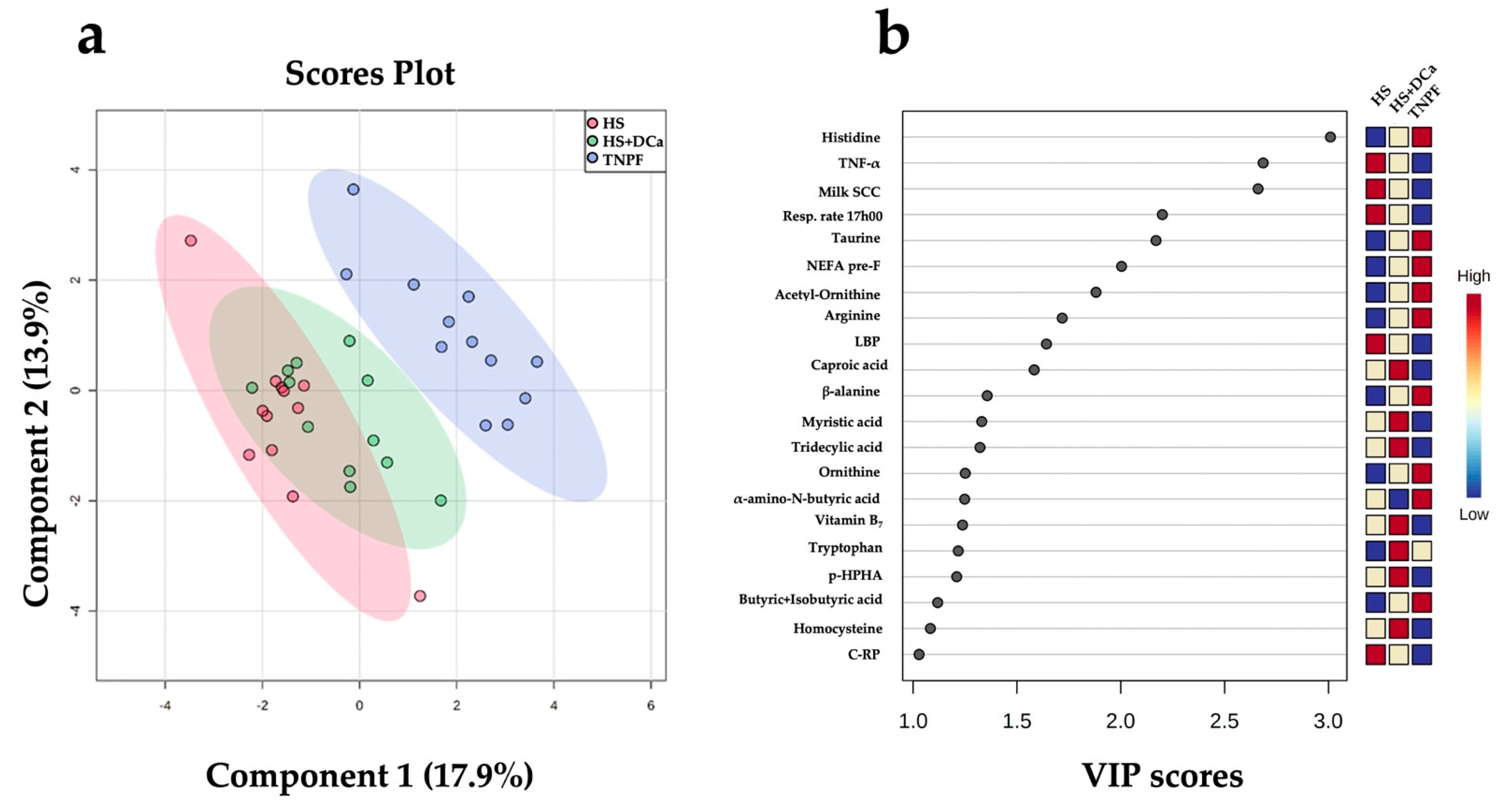
2.4. Correlation of Selected Fecal Metabolites with Leaky Gut and Inflammation Markers, Lactation Performance and Clinical Parameters
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Treatments
4.2. Environmental and Physiological Parameter Measurements
4.3. Sample Collection and Analysis
4.3.1. Targeted Quantitative Metabolomics
4.3.2. Vitamin Analysis
4.3.3. Milk Composition Analyses
4.4. Multivariate Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernabucci, U.; Bi-ani, S.; Buggiotti, L.; Vitali, A.; Lacetera, N.; Nardone, A. The effects of heat stress in Italian Holstein dairy cattle. J. Dairy Sci. 2014, 97, 471–486. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, N.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, 52–77. [Google Scholar] [CrossRef] [Green Version]
- Nardone, A.; Ronchi, B.; Lacetera, N.; Ranieri, M.; Bernabucci, U.E. Effects of climate changes on animal production and sustainability of livestock systems. Livest. Sci. 2010, 130, 57–69. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheelock, J.B.; Rhoads, R.; Vanbaale, M.; Sanders, S.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Wallin, R.P.A.; Lundqvist, A.; Moré, S.H.; von Bonin, A.; Kiessling, R.; Ljunggren, H.G. Heat-shock proteins as activators of the innate inmune system. Trends Immunol. 2002, 23, 130–135. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.J.; Fernandez, M.S.; Baumgard, L.H. Glucose requirements of an activated immune system in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2360–2374. [Google Scholar] [CrossRef] [Green Version]
- Rea, K.; Dinan, T.G.; Cryan, J.F. The microbiome: A key regulator of stress and neuroinflammation. Neurobiol. Stress 2016, 4, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Lambert, G.P. Role of gastrointestinal permeability in exertional heatstroke. Exerc. Sport Sci. Rev. 2004, 2, 185–190. [Google Scholar] [CrossRef]
- Lambert, G.P. Stress-induced gastrointestinal barrier dysfunction and its inflammatory effects. J. Anim. Sci. 2009, 87, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Gabler, N.K. Heat stress reduces intestinal barrier integrity and favors intestinal glucose transport in growing pigs. PLoS ONE 2013, 8, e700215. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, X.H.; Yang, L.; Chen, X.Y.; Jiang, R.S.; Jin, S.H.; Geng, Z.Y. Resveratrol alleviates heat stress-induced impairment of intestinal morphology, microflora, and barrier integrity in broilers. Poult. Sci. 2017, 96, 4325–4332. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Mahon, B.D.; Froicu, M.; Cantorna, M.T. Calcium and 1α, 25-dihydroxyvitamin D3 target the TNF-α pathway to suppress experimental inflammatory bowel disease. Eur. J. Immunol. 2005, 35, 217–224. [Google Scholar] [CrossRef]
- Liu, F.; Cottrell, J.J.; Furness, J.B.; Rivera, L.; Kelly, F.W.; Wijesiriwardana, U.; Pustovit, R.V.; Fothergill, L.J.; Bravo, D.M.; Celi, P.; et al. Selenium and vitamin E together improve intestinal epithelial barrier function and alleviate oxidative stress in heat-stressed pigs. Exp. Physiol. 2016, 101, 801–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waage, S.; Mørk, T.; Røros, A.; Aasland, D.; Hunshamar, A.; Odegaard, A.S. Bacteria associated with clinical mastitis in dairy heifers. J. Dairy Sci. 1998, 82, 712–719. [Google Scholar] [CrossRef]
- Sun, H.Z.; Wang, D.M.; Wang, B.; Wang, J.K.; Liu, H.Y.; Guan, L.L.; Liu, J.X. Metabolomics of Four Biofluids from Dairy Cows: Potential Biomarkers for Milk Production and Quality. J. Proteome Res. 2015, 14, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J.Q. Identification of diagnostic biomarkers and metabolic pathway shifts of heat-stressed lactating dairy cows. J. Proteom. 2015, 125, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Rochus, K.; Janssens, G.P.; Hesta, M. Dietary fibre and the importance of the gut microbiota in feline nutrition: A review. Nutr. Res. Rev. 2014, 7, 295–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stacy, A.; Andrade-Oliveira, V.; McCulloch, J.A.; Hild, B.; Oh, J.H.; Perez-Chaparro, P.J.; Sim, C.K.; Lim, A.I.; Link, V.M.; Enamorado, M.; et al. Infection trains the host for microbiota-enhanced resistance to pathogens. Cell 2021, 184, 615–627. [Google Scholar] [CrossRef]
- Ghaly, S.; Kaakoush, N.O.; Lloyd, F.; McGonigle, T.; Mok, D.; Baird, A.; Klopcic, B.; Gordon, L.; Gorman, S.; Forest, C.; et al. High dose vitamin D supplementation alters faecal microbiome and predisposes mice to more severe colitis. Sci. Rep. 2018, 8, 11511. [Google Scholar] [CrossRef] [PubMed]
- Haffner, J.J.; Katemauswa, M.; Kagone, T.S.; Hossain, E.; Jacobson, D.; Flores, K.; Parab, A.R.; Obregon-Tito, A.J.; Tito, R.Y.; Reyes, L.M.; et al. The Core Human Fecal Metabolome. bioRxiv 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.05.08.442269v1.full (accessed on 1 October 2021).
- Zhao, Y.; Wu, J.; Li, J.V.; Zhou, N.Y.; Tang, H.; Wang, Y. Gut microbiota composition modifies fecal metabolic profiles in mice. J. Proteome Res. 2013, 12, 2987–2999. [Google Scholar] [CrossRef] [PubMed]
- Kirk, R.E. Experimental design. In Handbook of Psychology, 2nd ed.; Schinka, J., Velicer, W.F., Eds.; Wiley: New York, NY, USA, 2012. [Google Scholar]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [Green Version]
- Siegerstetter, S.C.; Petri, R.M.; Magowan, E.; Lawlor, P.G.; Zebeli, Q.; O’Connell, N.E.; Metzler-Zebeli, B.U. Feed restriction modulates the fecal microbiota composition, nutrient retention, and feed efficiency in chickens di-vergent in residual feed intake. Front. Microbiol. 2018, 9, 2698. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.E.; Argenzio, R.A.; Clemens, E.T. Microbial digestion: Rumen versus large intestine. In Digestive Physiology and Metabolism in Ruminants; Springer: Dordrecht, The Netherlands, 1980; pp. 685–706. [Google Scholar]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Mao, S.; Zhang, R.; Wang, D.; Zhu, W. The diversity of the fecal bacterial community and its relationship with the concentration of volatile fatty acids in the feces during subacute rumen acidosis in dairy cows. BMC Vet. Res. 2012, 8, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Min, L.; Zheng, N.; Wang, J. Effect of heat stress on bacterial composition and metabolism in the rumen of lactating dairy cows. Animals 2019, 9, 925. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Kim, E.T.; Eom, J.S.; Choi, Y.Y.; Lee, S.J.; Lee, S.S.; Chung, C.D.; Lee, S.S. Exploration of metabolite profiles in the biofluids of dairy cows by proton nuclear magnetic resonance analysis. PLoS ONE 2021, 16, e0246290. [Google Scholar] [CrossRef]
- Yadav, B.; Singh, G.; Verma, A.K.; Dutta, N.; Sejian, V. Impact of heat stress on rumen functions. Vet. World 2013, 6, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Yi, H.; Wu, Q.; Jiang, Z.; Wang, L. Effects of acute heat stress on intestinal microbiota in grow-finishing pigs, and associations with feed intake and serum profile. J. Appl. Microbiol. 2020, 128, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Wang, W.; Yang, H.; Wang, D.; Ling, W. Influence of intestinal microbiota on the catabolism of flavonoids in mice. J. Food Sci. 2016, 81, H3026–H3034. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Metabolism of dietary and microbial vitamin B family in the regulation of host immunity. Front. Nutr. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.S.; Liang, S.; Zong, M.H.; Yang, J.G.; Lou, W.Y. Microbial synthesis of functional odd-chain fatty acids: A review. World J. Microbiol. Biotechnol. 2020, 36, 35. [Google Scholar] [CrossRef]
- Xu, Y.; Labedan, B.; Glansdorff, N. Surprising arginine biosynthesis: A reappraisal of the enzymology and evolution of the pathway in microorganisms. Microbiol. Mol. Biol. Rev. 2007, 71, 36–47. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Guo, H.; Zheng, W.; Xue, Y.; Zhao, R.; Yao, W. Heat stress affects fecal microbial and metabolic alterations of primiparous sows during late gestation. J. Anim. Sci. Biotechnol. 2019, 10, 84. [Google Scholar] [CrossRef]
- Kamiya, M.; Kamiya, Y.; Tanaka, M.; Oki, T.; Nishiba, Y.; Shioya, S. Effects of high ambient temper-ature and restricted feed intake on urinary and plasma 3-methylhistidine in lactating Holstein cows. Anim. Sci. J. 2006, 77, 201–207. [Google Scholar] [CrossRef]
- Thompson, M.G.; Palmer, R.M.; Thom, A.; Mackie, S.C.; Morrison, K.S.; Harris, C.I. Measurement of protein degradation by release of labelled 3-methylhistidine from skeletal muscle and non-muscle cells. J. Cell. Physiol. 1996, 166, 506–511. [Google Scholar] [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Blatteis, C.M. Endotoxic fever: New concepts of its regulation suggest new approaches to its management. Pharmacol. Ther. 2006, 111, 194–223. [Google Scholar] [CrossRef]
- Colotta, F.; Jansson, B.; Bonelli, F. Modulation of inflammatory and immune responses by vitamin D. J. Autoimmun. 2007, 85, 78–97. [Google Scholar] [CrossRef]
- Schaffer, S.W.; Azuma, J.; Mozaffari, M. Role of antioxidant activity of taurine in diabetes. Can. J. Physiol. Pharmacol. 2009, 87, 91–99. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, N.; Zhang, F.; Yue, W.; Liang, M. Effect of taurine on leukocyte function. Eur. J. Pharmacol. 2009, 616, 275–280. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canadian Council on Animal Care. Guidelines to the Care and Use of Experimental Animals, 2nd ed.; CCAC: Ottawa, ON, Canada, 1993; Volume 1.
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Shwartz, G.; Rhoads, M.L.; Vanbaale, M.J.; Rhoads, R.P.; Baumgard, L.H. Effects of a supplemental yeast culture on heat-stressed lactating Holstein cows. J. Dairy Sci. 2009, 92, 935–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foroutan, A.; Guo, A.C.; Vazquez-Fresno, R.; Lipfert, M.; Zhang, L.; Zheng, J.; Badran, H.; Budinski, Z.; Mandal, R.; Ametaj, B.N.; et al. Chemical composition of commercial cow’s milk. J. Agric. Food Chem. 2019, 67, 4897–4914. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; Morais, D.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.E.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucl. Acids Res. 2021, 11, 6601. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Metabolic Pathway | FC HS vs. TNPF a | FC HS vs. HS+DCa b | ANOVA | PFDR HS vs. TNPF d | PFDR HS vs. HS+Dca e |
|---|---|---|---|---|---|---|
| PFDRc | ||||||
| Rectal temperature 17h00 TNF-α | Heat stress Inflammation | 1.06 3.55 | −1.02 −2.08 | <0.001 <0.001 | <0.001 | <0.001 |
| <0.001 | 0.02 | |||||
| Respiratory rate 17h | Heat stress | 2.48 | −1.15 | <0.001 | <0.001 | ns |
| Milk SCC | Inflammation | 4.1 | −2.31 | <0.001 | <0.001 | 0.03 |
| Rectal temperature 14h00 | Heat stress | 1.02 | −0.99 | <0.001 | <0.001 | ns |
| LBP | Heat stress | 1.74 | −1.45 | <0.001 | <0.001 | 0.006 |
| Tridecylic acid | Microbial metabolism | 2.06 | 0.71 | <0.001 | 0.03 | ns |
| Myristic acid | Fatty acid metabolism | 1.87 | 0.79 | <0.001 | 0.02 | ns |
| NEFA pre-F | Fatty acid metabolism | −0.92 | 1.01 | <0.001 | <0.001 | ns |
| Vitamin B7 | Vitamin metabolism | 1.58 | 0.9 | <0.001 | <0.001 | ns |
| Histidine | Amino acid synthesis | −0.12 | 0.42 | <0.001 | <0.01 | ns |
| p-HPHA | Microbial metabolism | 2.44 | 0.43 | <0.001 | <0.01 | <0.01 |
| Acetyl ornithine | Amino acid synthesis | −0.34 | 0.91 | <0.001 | 0.01 | ns |
| Methyl histidine | Amino acid synthesis | −0.89 | 0.36 | 0.02 | ns | 0.01 |
| Arginine | Amino acid synthesis | −0.42 | 0.71 | 0.02 | 0.02 | ns |
| Rectal temperature 08h00 | Heat stress | 1.01 | −1.00 | 0.02 | 0.04 | ns |
| Retinol | Metabolism of vitamins | 1.41 | −0.92 | 0.04 | ns | ns |
| Fecal Metabolite | FC a | log2(FC) | p-Value b |
|---|---|---|---|
| α-Tocopherol | +3.20 | 1.67 | <0.001 |
| δ-Tocopherol | −0.23 | −2.12 | 0.011 |
| β + γ-Tocopherol | −0.51 | −0.96 | 0.011 |
| Response Variable | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolite | LBP | Calp | TNF | C−RP | DMI | MY | MFY | MPY | MLY | dNFA | mFA | pFA | NEFA | Ins | RT | RR | MSCC |
| Vitamin B1 | −0.19 | −0.51 * | −0.16 | 0.40 * | −0.06 | −0.10 | −0.14 | −0.10 | −0.09 | −0.16 | −0.05 | −0.18 | −0.18 | 0.04 | −0.03 | −0.13 | −0.12 |
| Vitamin B2 | −0.27 | −0.41 * | −0.31 | −0.20 | 0.10 | 0.09 | 0.05 | 0.09 | 0.08 | 0.05 | 0.06 | 0.02 | 0.13 | −0.28 | −0.36 | −0.51 * | −0.29 |
| Vitamin B5 | −0.27 | −0.51 * | −0.22 | −0.06 | −0.04 | −0.01 | 0.06 | 0.03 | −0.02 | 0.03 | 0.09 | 0.06 | 0.00 | −0.39 | −0.27 | −0.40 * | −0.29 * |
| Vitamin B7 | 0.55 ** | 0.10 | 0.41 * | 0.56 * | −0.19 | −0.09 | −0.03 | −0.04 | −0.08 | −0.01 | −0.03 | −0.07 | −0.16 | 0.27 | 0.61 ** | 0.66 ** | 0.41 |
| Vitamin B9 | 0.42 * | 0.40 * | 0.11 | −0.29 | 0.19 | 0.12 | −0.02 | 0.09 | 0.15 | 0.06 | 0.01 | −0.13 | −0.07 | 0.16 | 0.27 | 0.48 * | 0.28 |
| Retinol | 0.24 | 0.20 | 0.49 * | 0.43 * | −0.52 ** | −0.56 * | −0.55 ** | −0.62 ** | −0.56 ** | −0.56 ** | −0.47 * | −0.55 * | −0.35 | 0.49 * | 0.37 * | 0.37 | 0.35 |
| α-Tocopherol | 0.02 | 0.08 | −0.24 | −0.19 | −0.02 | 0.13 | 0.26 | 0.19 | 0.11 | 0.25 | 0.15 | 0.35 | 0.31 | 0.09 | −0.30 | −0.27 | −0.33 |
| β+γ-Tocopherol | −0.01 | 0.05 | −0.30 | 0.02 | 0.09 | 0.21 | 0.14 | 0.19 | 0.22 | 0.13 | 0.12 | 0.20 | 0.38 | −0.10 | −0.01 | 0.14 | −0.23 |
| δ-Tocopherol | −0.23 | 0.10 | 0.00 | −0.09 | 0.19 | 0.06 | −0.02 | 0.05 | 0.07 | −0.03 | 0.06 | −0.08 | −0.10 | −0.43 | 0.08 | −0.07 | −0.09 |
| Alanine | −0.23 | −0.37 | −0.16 | −0.03 | −0.25 | −0.23 | −0.10 | −0.18 | −0.22 | −0.12 | −0.08 | −0.09 | −0.10 | −0.35 | −0.18 | −0.20 | −0.35 |
| Arginine | −0.51 * | −0.24 | −0.46 * | −0.33 | 0.18 | 0.11 | 0.09 | 0.10 | 0.10 | 0.08 | 0.15 | 0.02 | 0.10 | −0.52 | −0.48 * | −0.64 ** | −0.50 * |
| Asparagine | 0.04 | −0.16 | 0.31 | 0.41 * | −0.31 | −0.32 | −0.07 | −0.20 | −0.31 | −0.12 | −0.06 | −0.01 | −0.26 | −0.05 | 0.27 | 0.17 | 0.00 |
| Aspartate | −0.19 | −0.06 | 0.02 | 0.12 | 0.05 | −0.08 | −0.08 | −0.07 | −0.10 | −0.09 | −0.05 | −0.10 | −0.11 | −0.37 * | −0.05 | −0.26 | −0.13 |
| Citrulline | −0.10 | −0.46 * | 0.22 | 0.31 | −0.23 | −0.34 | −0.07 | −0.26 | −0.33 | −0.12 | −0.02 | −0.08 | −0.34 | −0.14 | 0.09 | −0.02 | −0.04 |
| Glutamate | −0.32 | −0.32 | −0.15 | −0.03 | −0.08 | −0.11 | 0.02 | −0.08 | −0.12 | −0.02 | 0.07 | 0.00 | −0.08 | −0.42 * | −0.17 | −0.34 | −0.33 |
| Glutamine | 0.04 | −0.23 | 0.31 | 0.17 | −0.51 * | −0.56 * | −0.34 | −0.49 * | −0.57 ** | −0.39 * | −0.34 | −0.24 | −0.26 | 0.04 | 0.11 | 0.06 | 0.07 |
| Glycine | −0.15 | −0.41 * | −0.08 | 0.14 | −0.24 | −0.18 | −0.06 | −0.14 | −0.18 | −0.10 | 0.01 | −0.08 | −0.12 | −0.28 | −0.07 | −0.18 | −0.24 |
| Histidine | −0.55 ** | −0.36 * | −0.51 * | −0.29 | 0.02 | 0.07 | 0.07 | 0.07 | 0.05 | 0.04 | 0.06 | 0.11 | 0.28 | −0.47 * | −0.61 ** | −0.73 ** | −0.63 ** |
| Tyrosine | −0.21 | −0.33 * | 0.06 | 0.21 | −0.26 | −0.33 | −0.15 | −0.27 | −0.34 | −0.18 | −0.09 | −0.19 | −0.24 | −0.21 | −0.04 | −0.24 | −0.16 |
| Valine | −0.45 * | −0.51 * | −0.16 | 0.12 | −0.31 | −0.24 | −0.09 | −0.19 | −0.25 | −0.16 | −0.09 | −0.01 | 0.01 | −0.36 * | −0.27 | −0.43 * | −0.43 * |
| Isoleucine | −0.29 | −0.43 * | 0.00 | 0.19 | −0.35 | −0.34 | −0.16 | −0.28 | −0.35 | −0.20 | −0.13 | −0.13 | −0.16 | −0.28 | −0.11 | −0.26 | −0.27 |
| Leucine | −0.31 | −0.48 * | 0.08 | 0.22 | −0.41 * | −0.46 * | −0.27 | −0.42 * | −0.46 * | −0.32 | −0.22 | −0.25 | −0.23 | −0.20 | −0.09 | −0.23 | −0.19 |
| Lysine | −0.26 | −0.36 | −0.15 | 0.04 | −0.26 | −0.21 | −0.09 | −0.17 | −0.21 | −0.13 | −0.06 | −0.07 | −0.05 | −0.35 | −0.20 | −0.26 | −0.34 |
| Methionine | −0.33 | −0.46 * | −0.11 | 0.09 | −0.23 | −0.21 | −0.02 | −0.14 | −0.22 | −0.08 | 0.00 | 0.01 | −0.13 | −0.36 | −0.23 | −0.40 * | −0.31 |
| Ornithine | −0.33 | −0.51 * | −0.23 | −0.04 | −0.21 | −0.15 | −0.03 | −0.09 | −0.15 | −0.07 | −0.05 | 0.04 | 0.01 | −0.38 * | −0.30 | −0.35 * | −0.51 * |
| Phenylalanine | −0.29 | −0.45 * | 0.03 | 0.16 | −0.29 | −0.35 | −0.16 | −0.29 | −0.36 | −0.20 | −0.11 | −0.17 | −0.23 | −0.21 | −0.11 | −0.30 | −0.21 |
| Proline | −0.12 | −0.35 | 0.01 | 0.21 | −0.29 | −0.23 | −0.04 | −0.15 | −0.23 | −0.10 | −0.01 | 0.00 | −0.17 | −0.24 | 0.03 | −0.06 | −0.14 |
| Serine | −0.13 | −0.47 * | 0.07 | 0.20 | −0.34 | −0.37 | −0.17 | −0.30 | −0.37 | −0.20 | −0.12 | −0.19 | −0.27 | −0.14 | −0.03 | −0.11 | −0.13 |
| Threonine | −0.29 | −0.53 * | −0.07 | 0.11 | −0.28 | −0.29 | −0.10 | −0.22 | −0.30 | −0.17 | −0.08 | −0.06 | −0.10 | −0.35 | −0.18 | −0.33 | −0.29 |
| Tryptophan | −0.37 | −0.63 ** | −0.27 | 0.16 | −0.25 | −0.19 | −0.08 | −0.13 | −0.20 | −0.13 | −0.04 | −0.08 | −0.05 | −0.21 | −0.28 | −0.41 * | −0.40 * |
| Methylhistidine | −0.17 | 0.10 | 0.17 | −0.05 | 0.05 | −0.13 | −0.04 | −0.17 | −0.15 | −0.02 | 0.07 | −0.19 | −0.34 | −0.24 | 0.02 | −0.15 | 0.12 |
| Choline | −0.30 | −0.45 * | −0.20 | −0.12 | −0.12 | −0.16 | −0.08 | −0.11 | −0.15 | −0.13 | −0.06 | −0.04 | 0.05 | −0.45 * | −0.23 | −0.20 | −0.33 |
| Acetyl-ornithine | −0.47 * | −0.30 | −0.55 ** | −0.24 | −0.05 | 0.15 | 0.14 | 0.17 | 0.15 | 0.13 | 0.12 | 0.18 | 0.21 | −0.47 * | −0.54 ** | −0.52 ** | −0.62 ** |
| Taurine | −0.35 | −0.23 | −0.23 | −0.19 | −0.09 | −0.16 | −0.12 | −0.16 | −0.18 | −0.17 | −0.08 | −0.11 | 0.00 | −0.19 | −0.33 | −0.36 * | −0.32 * |
| Homocysteine | 0.11 | −0.03 | 0.26 | 0.04 | −0.06 | −0.18 | −0.21 | −0.17 | −0.15 | −0.22 | −0.14 | −0.26 | −0.18 | −0.18 | 0.39 * | 0.30 | 0.14 |
| trans-OH-Proline | 0.04 | −0.14 | −0.23 | 0.17 | 0.10 | 0.26 | 0.37 | 0.31 | 0.26 | 0.31 | 0.32 | 0.43 * | 0.17 | 0.01 | −0.16 | −0.14 | −0.11 |
| β-Alanine | −0.49 | −0.16 | −0.32 | −0.10 | 0.16 | 0.12 | 0.03 | 0.09 | 0.10 | 0.03 | 0.03 | 0.04 | 0.21 | −0.54 * | −0.27 | −0.53 * | −0.55 * |
| Proline betaine | 0.07 | 0.30 | 0.02 | 0.12 | −0.04 | 0.01 | 0.09 | 0.04 | 0.02 | 0.14 | 0.15 | −0.05 | −0.03 | −0.11 | 0.17 | 0.18 | −0.02 |
| α-Amino-N-butyric ac | −0.42 | −0.23 | −0.33 | −0.03 | −0.17 | −0.04 | −0.01 | −0.04 | −0.02 | −0.01 | −0.03 | 0.02 | −0.02 | −0.32 | −0.30 | −0.23 | −0.44 * |
| γ-Aminobutyric | 0.04 | −0.38 | −0.15 | 0.07 | −0.11 | 0.19 | 0.28 | 0.25 | 0.20 | 0.22 | 0.23 | 0.37 | 0.14 | −0.12 | −0.08 | 0.05 | −0.17 |
| Acetic acid | −0.42 * | −0.42 * | −0.42 * | −0.15 | 0.08 | 0.14 | 0.22 | 0.16 | 0.14 | 0.21 | 0.24 | 0.20 | 0.09 | −0.50 * | −0.45 * | −0.57 ** | −0.57 ** |
| Propionic acid | −0.26 | −0.51 * | −0.29 | 0.03 | 0.02 | 0.11 | 0.23 | 0.14 | 0.11 | 0.21 | 0.22 | 0.24 | 0.04 | −0.38 * | −0.30 | −0.37 * | −0.43 * |
| Butyric+isobutyric acid | −0.41 * | −0.40 * | −0.34 | −0.10 | −0.08 | 0.08 | 0.19 | 0.09 | 0.07 | 0.13 | 0.21 | 0.21 | 0.12 | −0.39 * | −0.43 * | −0.53 * | −0.41 * |
| 2-Methylbutyric acid | −0.02 | −0.40 * | 0.05 | 0.14 | −0.59 * | −0.44 * | −0.41 * | −0.43 | −0.43 * | −0.41 * | −0.40 * | −0.34 | 0.05 | 0.12 | −0.01 | 0.01 | −0.14 |
| Valeric+Isovaleric acid | −0.11 | −0.22 | 0.04 | 0.17 | −0.01 | 0.10 | 0.28 | 0.14 | 0.11 | 0.27 | 0.34 | 0.16 | −0.19 | −0.30 | 0.02 | −0.11 | 0.01 |
| 3-Methylvaleric acid | 0.03 | 0.21 | 0.17 | −0.20 | −0.05 | −0.23 | −0.28 | −0.26 | −0.23 | −0.26 | −0.22 | −0.34 | −0.13 | 0.38 * | 0.08 | 0.19 | 0.07 |
| Caproic acid | 0.67 * | 0.01 | 0.13 | 0.07 | −0.04 | −0.08 | −0.18 | −0.14 | −0.07 | −0.11 | −0.19 | −0.21 | −0.03 | 0.59 * | 0.27 | 0.50 * | 0.34 * |
| Heptanoic acid | −0.05 | −0.53 ** | 0.00 | 0.07 | −0.36 | −0.07 | 0.04 | −0.03 | −0.05 | 0.00 | 0.01 | 0.11 | −0.01 | −0.21 | −0.03 | 0.02 | −0.06 |
| Caprylic acid | −0.06 | −0.76 ** | 0.18 | 0.33 | −0.34 | −0.38 | −0.30 | −0.34 | −0.35 | −0.33 | −0.26 | −0.29 | −0.30 | 0.06 | 0.13 | 0.09 | 0.03 |
| Pelargonic acid | 0.00 | −0.31 | −0.04 | 0.36 | −0.45 * | −0.13 | −0.01 | −0.09 | −0.11 | −0.08 | −0.04 | 0.09 | 0.06 | 0.14 | 0.06 | 0.21 | 0.08 |
| Capric acid | −0.10 | −0.68 ** | 0.29 | 0.33 | −0.40 * | −0.44 * | −0.34 | −0.42 | −0.42 * | −0.39 * | −0.29 | −0.30 | −0.31 | 0.05 | 0.16 | 0.04 | 0.12 |
| Undecylic acid | 0.02 | −0.68 ** | 0.26 | 0.31 | −0.24 | −0.26 | −0.13 | −0.21 | −0.25 | −0.19 | −0.13 | −0.05 | −0.28 | 0.11 | 0.14 | 0.07 | 0.12 |
| Lauric acid | −0.07 | −0.73 ** | 0.12 | 0.26 | −0.19 | −0.18 | −0.09 | −0.14 | −0.17 | −0.17 | −0.08 | 0.01 | −0.18 | 0.10 | 0.01 | −0.08 | −0.05 |
| Tridecylic acid | 0.34 | −0.07 | 0.38 * | 0.52 * | −0.33 | −0.24 | −0.26 | −0.21 | −0.23 | −0.27 | −0.31 | −0.17 | −0.08 | 0.37 * | 0.51 * | 0.59 * | 0.30 * |
| Myristic acid | 0.34 | −0.04 | 0.43 * | 0.43 * | −0.34 | −0.36 | −0.45 * | −0.35 | −0.35 | −0.42 * | −0.46 * | −0.40 * | −0.12 | 0.28 | 0.55 ** | 0.61 * | 0.30 * |
| Palmitic acid | −0.15 | 0.28 | −0.24 | −0.10 | −0.03 | −0.01 | 0.02 | 0.00 | 0.01 | 0.02 | −0.05 | 0.09 | 0.25 | 0.14 | −0.19 | 0.19 | −0.14 |
| Stearic acid | 0.22 | 0.40 | −0.08 | −0.14 | 0.50 * | 0.39 | 0.16 | 0.33 | 0.38 | 0.21 | 0.12 | 0.15 | 0.12 | −0.06 | 0.12 | 0.21 | 0.13 |
| Oleic acid | −0.08 | 0.42 | −0.21 | −0.15 | 0.35 | 0.37 | 0.27 | 0.33 | 0.36 | 0.33 | 0.17 | 0.29 | 0.20 | −0.42 * | −0.10 | −0.06 | −0.10 |
| Hippuric acid | 0.02 | 0.27 | −0.02 | 0.07 | 0.25 | 0.03 | −0.07 | −0.05 | 0.02 | 0.00 | −0.10 | −0.11 | 0.09 | 0.03 | 0.06 | 0.17 | 0.03 |
| Benzoic acid | 0.11 | −0.09 | 0.29 | 0.27 | −0.16 | −0.11 | 0.05 | −0.08 | −0.10 | 0.00 | 0.09 | 0.03 | −0.10 | −0.02 | 0.30 | 0.23 | 0.31 * |
| r p-HPHA | 0.36 | −0.06 | 0.32 | 0.27 | −0.45 * | −0.29 | −0.37 | −0.34 | −0.28 | −0.34 | −0.31 | −0.43 * | −0.14 | 0.27 | 0.39 * | 0.38 * | 0.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-González, A.; Rico, D.E.; Rico, J.E. Modulation of Fecal Metabolites by Heat Stress and Diet, and Their Association with Inflammation and Leaky Gut Markers in Dairy Cows. Metabolites 2022, 12, 142. https://doi.org/10.3390/metabo12020142
Ruiz-González A, Rico DE, Rico JE. Modulation of Fecal Metabolites by Heat Stress and Diet, and Their Association with Inflammation and Leaky Gut Markers in Dairy Cows. Metabolites. 2022; 12(2):142. https://doi.org/10.3390/metabo12020142
Chicago/Turabian StyleRuiz-González, Alexis, Daniel Enrique Rico, and Jorge Eduardo Rico. 2022. "Modulation of Fecal Metabolites by Heat Stress and Diet, and Their Association with Inflammation and Leaky Gut Markers in Dairy Cows" Metabolites 12, no. 2: 142. https://doi.org/10.3390/metabo12020142
APA StyleRuiz-González, A., Rico, D. E., & Rico, J. E. (2022). Modulation of Fecal Metabolites by Heat Stress and Diet, and Their Association with Inflammation and Leaky Gut Markers in Dairy Cows. Metabolites, 12(2), 142. https://doi.org/10.3390/metabo12020142