
Dynamic Analysis of Stool Microbiota of Simmental Calves and Effects of Diarrhea on Their Gut Microbiota


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Breeding
2.2. Daily Health Monitoring
2.3. Grouping
2.4. High-Throughput 16S Ribosomal RNA Gene Sequencing
2.5. Bioinformatic Analysis
2.6. Statistical Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
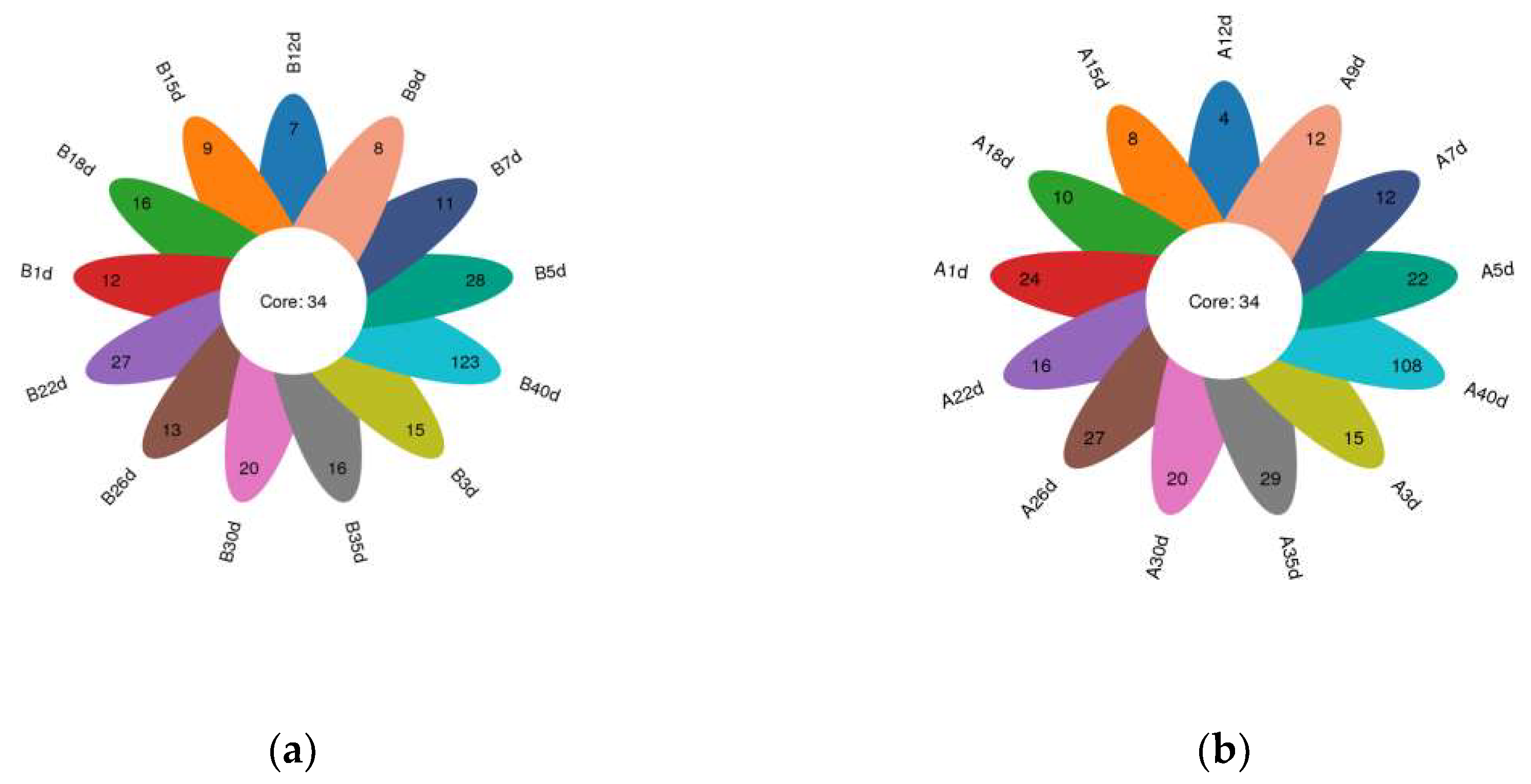
3.1. Data Collection and Diversity of Gut Microbiota
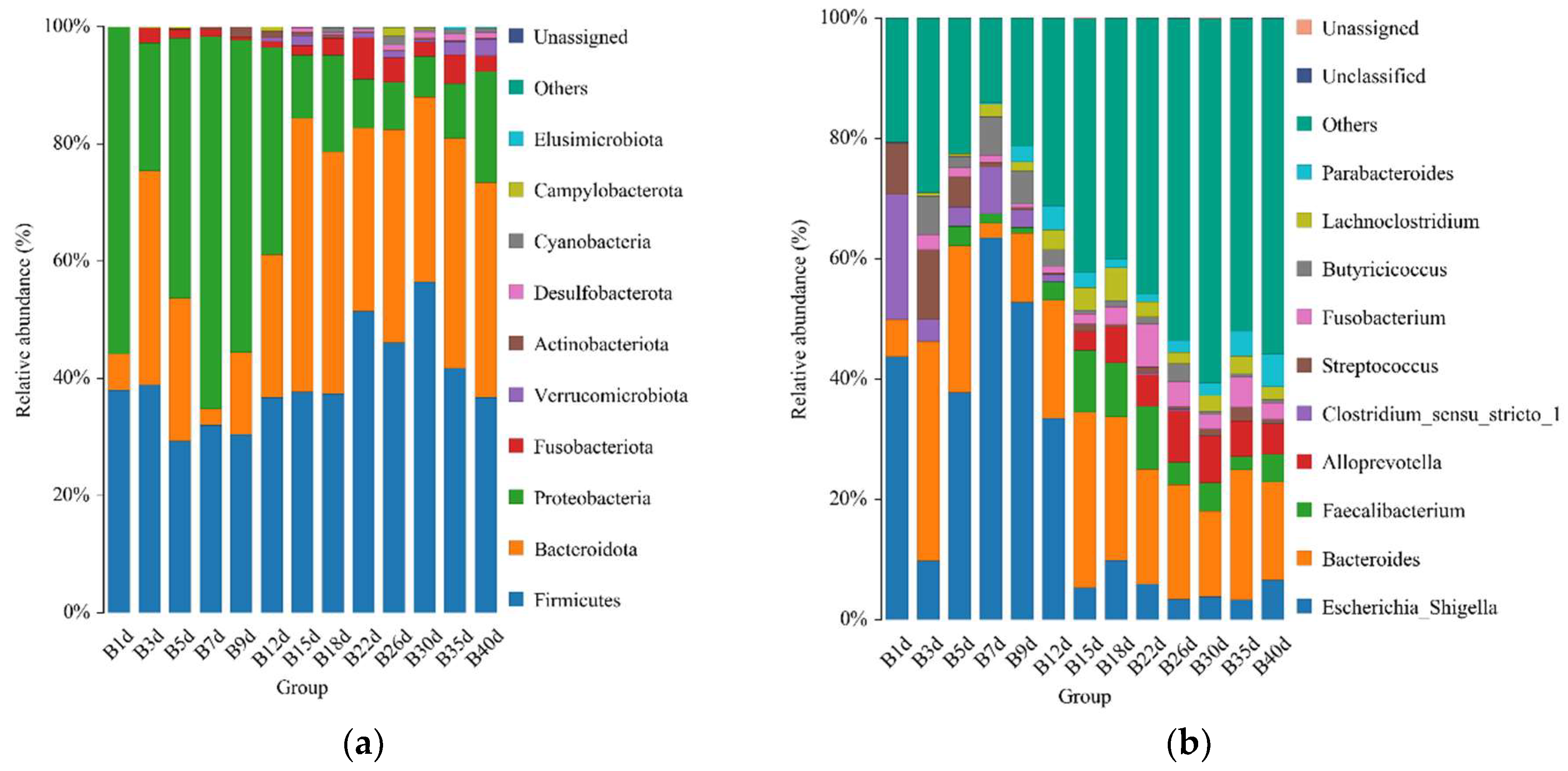
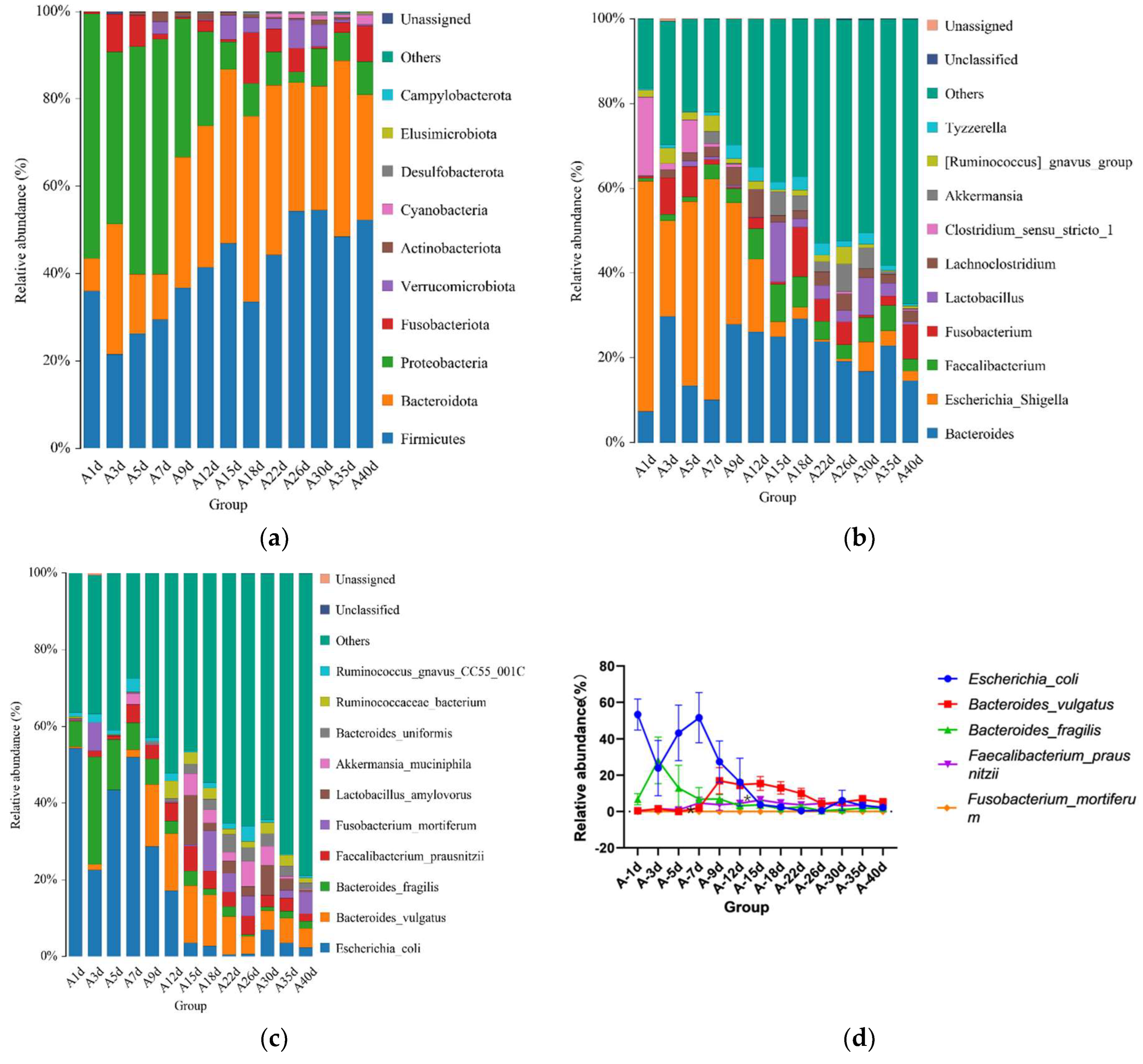
3.2. Taxonomic Composition of Healthy Simmental Calves during Preweaning Period
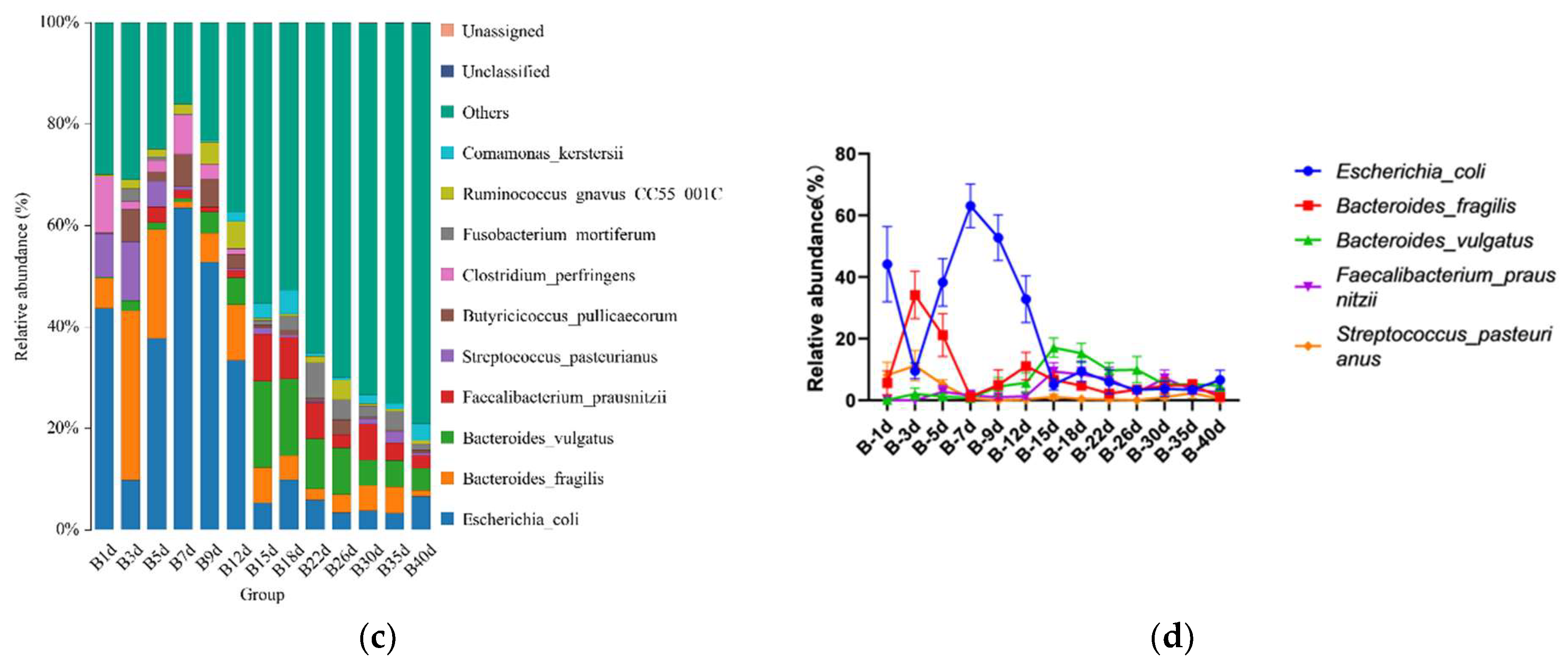
3.3. Taxonomic Composition of Simmental Calves Infected with Diarrhea during Preweaning Period
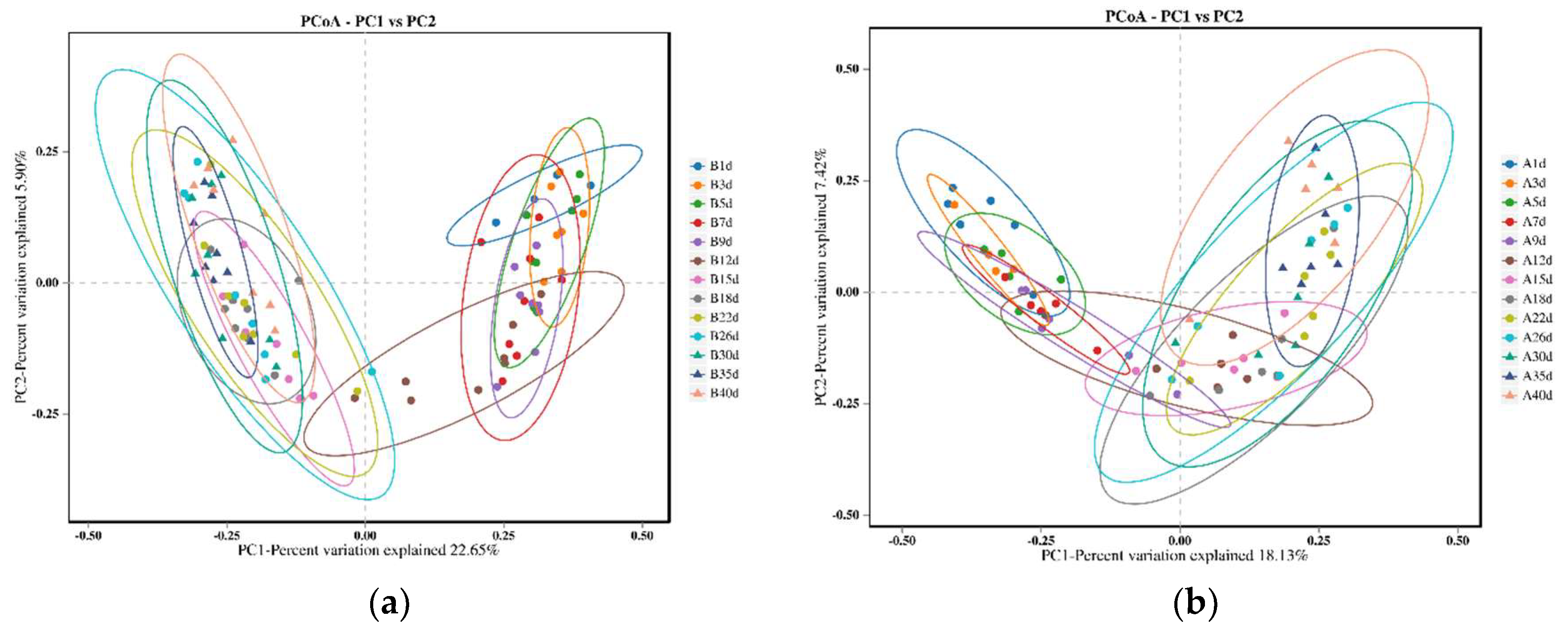
3.4. Similarity Analysis of Gut Microbiota of Simmental Calves during Preweaning Period
3.5. Differential Analysis of Intestinal Microbiota of Simmental Calves between Healthy and Diarrheal Groups
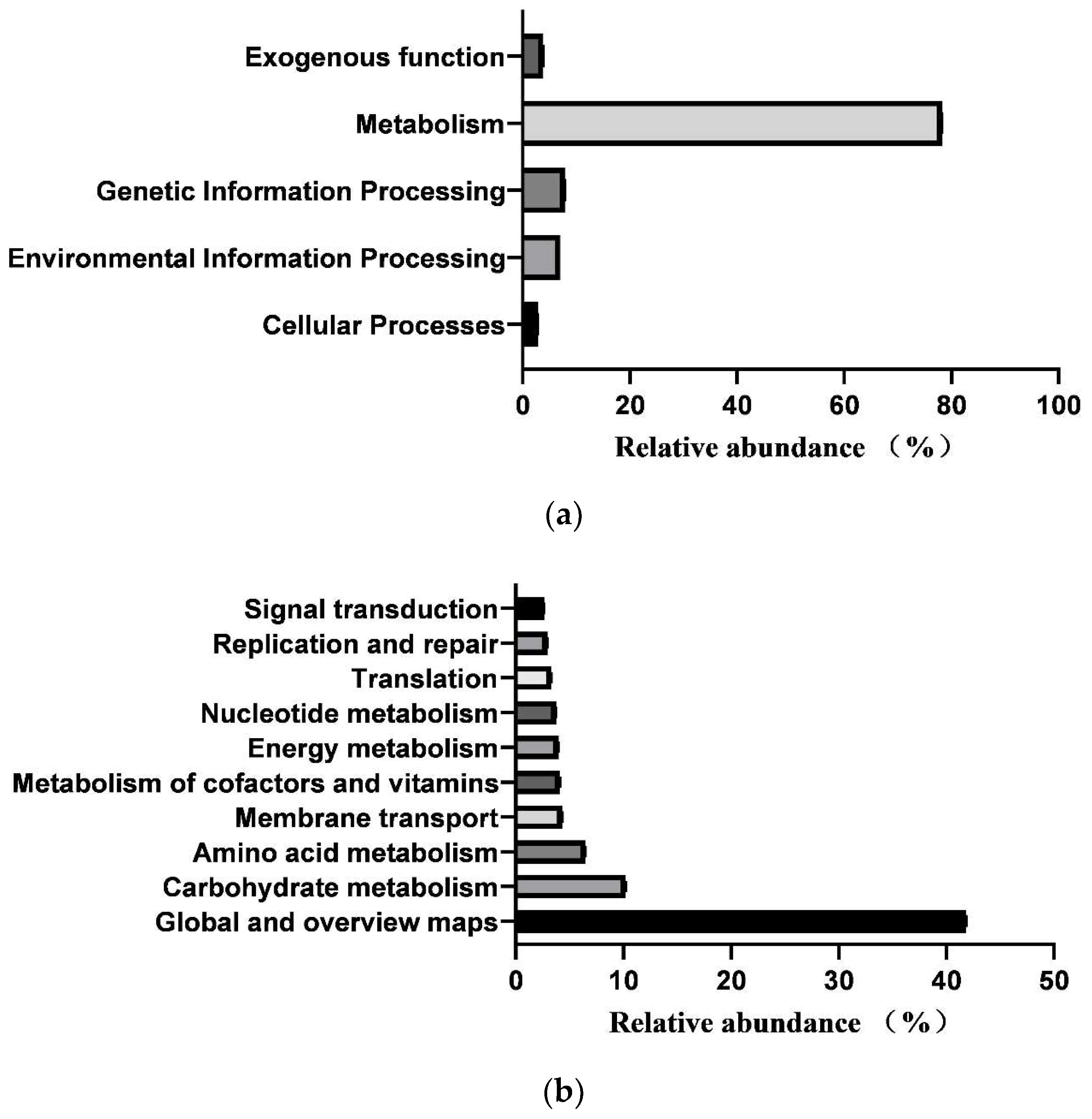
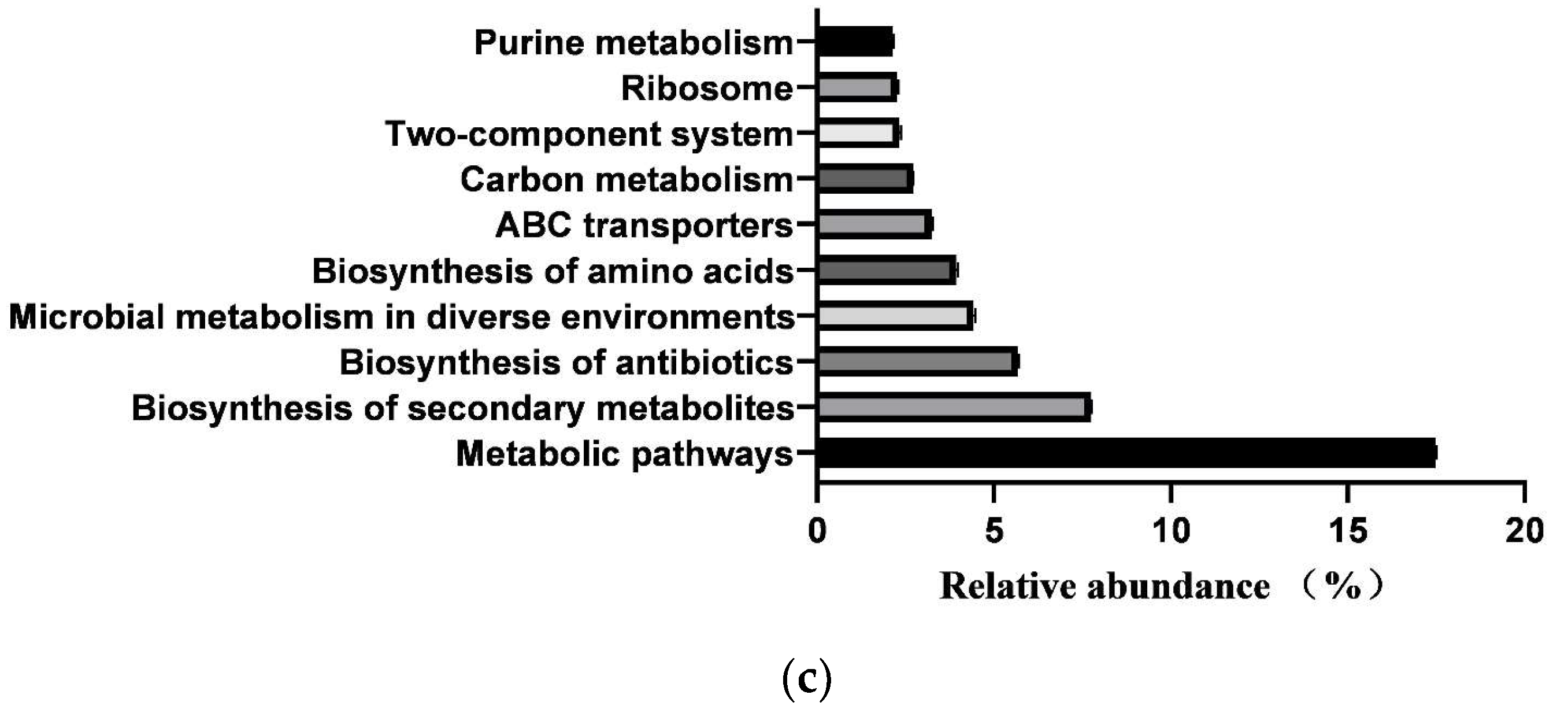
3.6. Prediction of Intestinal Microbial Functions of Simmental Calves
3.7. Differences in Microbial Functions of Calves between Healthy and Diarrheal Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giovanni, B.; Nicolas, L.; Giovanni, N.; Lorenzo, D.; Enrico, D.; Andrea, F.; Giulio, V. The application of Legendre Polynomials to model muscularity and body condition score in primiparous Italian Simmental cattle. VOL 2022, 21, 350–360. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Hu, R.; Peng, Q.; Xue, B.; Wang, L. Comparison of carcass characteristics and meat quality between Simmental crossbred cattle, cattle-yaks and Xuanhan yellow cattle. J. Sci. Food Agric. 2021, 101, 3927–3932. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Shen, Y.; You, L.; Zhang, Y.; Su, Z.; Peng, G.; Deng, J.; Zuo, Z.; Zhong, Z.; Ren, Z.; et al. Pueraria lobata polysaccharides alleviate neonatal calf diarrhea by modulating gut microbiota and metabolites. Front. Vet. Sci. 2023, 9, 1024392. [Google Scholar] [CrossRef] [PubMed]
- Tamrat, H.; Mekonnen, N.; Ferede, Y.; Cassini, R.; Belayneh, N. Epidemiological study on calf diarrhea and coccidiosis in dairy farms in Bahir Dar, North West Ethiopia. Ir. Vet. J. 2020, 73, 14. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Whon, T.W.; Sung, H.; Jeong, Y.S.; Jung, E.S.; Shin, N.R.; Hyun, D.W.; Kim, P.S.; Lee, J.Y.; Lee, C.H.; et al. Longitudinal evaluation of fecal microbiota transplantation for ameliorating calf diarrhea and improving growth performance. Nat. Commun. 2021, 12, 161. [Google Scholar] [CrossRef] [PubMed]
- Inabu, Y.; Fischer, A.; Song, Y.; Guan, L.L.; Oba, M.; Steele, M.A.; Sugino, T. Short communication: The effect of delayed colostrum feeding on plasma concentrations of glucagon-like peptide 1 and 2 in newborn calves. J. Dairy Sci. 2018, 101, 6627–6631. [Google Scholar] [CrossRef] [PubMed]
- Popović, N.; Dinić, M.; Tolinački, M.; Mihajlović, S.; Terzić-Vidojević, A.; Bojić, S.; Djokić, J.; Golić, N.; Veljović, K. New Insight into Biofilm Formation Ability, the Presence of Virulence Genes and Probiotic Potential of Enterococcus sp. Dairy Isolates. Front. Microbiol. 2018, 30, 78. [Google Scholar] [CrossRef]
- Bi, Y.; Tu, Y.; Zhang, N.; Wang, S.; Zhang, F.; Suen, G.; Shao, D.; Li, S.; Diao, Q. Multiomics analysis reveals the presence of a microbiome in the gut of fetal lambs. Gut 2021, 70, 853–864. [Google Scholar] [CrossRef]
- Guzman, C.E.; Wood, J.L.; Egidi, E.; White-Monsant, A.C.; Semenec, L.; Grommen, S.V.H.; Hill-Yardin, E.L.; De Groef, B.; Franks, A.E. A pioneer calf foetus microbiome. Sci. Rep. 2020, 10, 17712. [Google Scholar] [CrossRef]
- Du, Y.; Gao, Y.; Hu, M.; Hou, J.; Yang, L.; Wang, X.; Du, W.; Liu, J.; Xu, Q. Colonization and development of the gut microbiome in calves. J. Anim. Sci. Biotechnol. 2023, 14, 46. [Google Scholar] [CrossRef]
- Uyeno, Y.; Sekiguchi, Y.; Kamagata, Y. rRNA-based analysis to monitor succession of faecal bacterial communities in Holstein calves. Lett. Appl. Microbiol. 2010, 51, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Malmuthuge, N.; Griebel, P.J.; Guan, L.L. Taxonomic identification of commensal bacteria associated with the mucosa and digesta throughout the gastrointestinal tracts of preweaned calves. Appl. Environ. Microbiol. 2014, 80, 2021–2028. [Google Scholar] [CrossRef] [PubMed]
- Vlková, E.; Trojanová, I.; Rada, V. Distribution of bifidobacteria in the gastrointestinal tract of calves. Folia Microbiol. 2006, 51, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, D.; Raspa, F.; Marliani, G.; Nannoni, E.; Martelli, G.; Sardi, L.; Valle, E.; Pollesel, M.; Tassinari, M.; Buonaiuto, G. Growth Performance and Feed Intake Assessment of Italian Holstein Calves Fed a Hay-Based Total Mixed Ration: Preliminary Steps towards a Prediction Model. Vet. Sci. 2023, 10, 554. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Kim, S.; Kwon, M.S.; Lee, J.; Yu, D.H.; Song, R.H.; Choi, H.J.; Park, J. Rotavirus-mediated alteration of gut microbiota and its correlation with physiological characteristics in neonatal calves. J. Microbiol. 2019, 57, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Winder, C.B.; Kelton, D.F.; Duffield, T.F. Mortality risk factors for calves entering a multi-location white veal farm in Ontario, Canada. J. Dairy Sci. 2016, 99, 10174–10181. [Google Scholar] [CrossRef] [PubMed]
- Urie, N.J.; Lombard, J.E.; Shivley, C.B.; Kopral, C.A.; Adams, A.E.; Earleywine, T.J.; Olson, J.D.; Garry, F.B. Preweaned heifer management on US dairy operations: Part V. Factors associated with morbidity and mortality in preweaned dairy heifer calves. J. Dairy Sci. 2018, 101, 9229–9244. [Google Scholar] [CrossRef]
- Gulliksen, S.M.; Jor, E.; Lie, K.I.; Hamnes, I.S.; Løken, T.; Akerstedt, J.; Osterås, O. Enteropathogens and risk factors for diarrhea in Norwegian dairy calves. J. Dairy Sci. 2009, 92, 5057–5066. [Google Scholar] [CrossRef]
- Lopez, A.J.; Heinrichs, A.J. Invited review: The importance of colostrum in the newborn dairy calf. J. Dairy Sci. 2022, 105, 2733–2749. [Google Scholar] [CrossRef]
- Aghakeshmiri, F.; Azizzadeh, M.; Farzaneh, N.; Gorjidooz, M. Effects of neonatal diarrhea and other conditions on subsequent productive and reproductive performance of heifer calves. Vet. Res. Commun. 2017, 41, 107–112. [Google Scholar] [CrossRef]
- Xie, G.; Duff, G.C.; Hall, L.W.; Allen, J.D.; Burrows, C.D.; Bernal-Rigoli, J.C.; Dowd, S.E.; Guerriero, V.; Yeoman, C.J. Alteration of digestive tract microbiome in neonatal Holstein bull calves by bacitracin methylene disalicylate treatment and scours. J. Anim. Sci. 2013, 91, 4984–4990. [Google Scholar] [CrossRef] [PubMed]
- Penati, M.; Sala, G.; Biscarini, F.; Boccardo, A.; Bronzo, V.; Castiglioni, B.; Cremonesi, P.; Moroni, P.; Pravettoni, D.; Addis, M.F. Feeding Pre-weaned Calves With Waste Milk Containing Antibiotic Residues Is Related to a Higher Incidence of Diarrhea and Alterations in the Fecal Microbiota. Front. Vet. Sci. 2021, 8, 650150. [Google Scholar] [CrossRef] [PubMed]
- Lépine, A.F.P.; de Wit, N.; Oosterink, E.; Wichers, H.; Mes, J.; De Vos, P. Lactobacillus acidophilus Attenuates Salmonella-Induced Stress of Epithelial Cells by Modulating Tight-Junction Genes and Cytokine Responses. Front. Microbiol. 2018, 9, 1439. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Mustapha, A.; Lin, M.; Zheng, G. Biocontrol of the internalization of Salmonella enterica and Enterohaemorrhagic Escherichia coli in mung bean sprouts with an endophytic Bacillus subtilis. Int. J. Food Microbiol. 2017, 250, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Memon, F.U.; Yang, Y.; Leghari, I.H.; Lv, F.; Soliman, A.M.; Zhang, W.; Si, H. Transcriptome Analysis Revealed Ameliorative Effects of Bacillus Based Probiotic on Immunity, Gut Barrier System, and Metabolism of Chicken under an Experimentally Induced Eimeria tenella Infection. Genes 2021, 12, 536. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Nie, C.; Luo, R.; Qi, F.; Bai, X.; Chen, H.; Niu, J.; Chen, C.; Zhang, W. Effects of Multispecies Probiotic on Intestinal Microbiota and Mucosal Barrier Function of Neonatal Calves Infected with E. coli K99. Front. Microbiol. 2022, 12, 813245. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Stöber, M. Kennzeichen, Anamnese, Grundregeln der Untersuchungstechnik, Allgemeine Untersuchung. In Die Klinische Untersuchung des Rindes; Dirksen, G., Grunder, H.-D., Stöber, M., Eds.; Enke Verlag: Stuttgart, Germany; Erlangen, Germany, 2012; pp. 75–141. [Google Scholar]
- Lesmeister, K.E.; Heinrichs, A.J. Effects of corn processing on growth characteristics, rumen development, and rumen parameters in neonatal dairy calves. J. Dairy Sci. 2004, 87, 3439–3450. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Ferlizza, E.; Fasoli, S.; Cavallini, D.; Bolcato, M.; Andreani, G.; Isani, G. Preliminary Study on Urine Chemistry and Protein Profile in Cows and Heifers. Pak. Vet. J. 2020, 40, 413–418. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Dingkao, R.; Du, M.; Ahmad, A.A.; Liang, Z.; Zheng, J.; Shen, J.; Yan, P.; Ding, X. Fecal Microbiota Dynamics Reveal the Feasibility of Early Weaning of Yak Calves under Conventional Grazing System. Biology 2021, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Malmuthuge, N.; Steele, M.A.; Guan, L.L. Shift of hindgut microbiota and microbial short chain fatty acids profiles in dairy calves from birth to pre-weaning. FEMS Microbiol. Ecol. 2018, 94, fix179. [Google Scholar] [CrossRef] [PubMed]
- Klein-Jöbstl, D.; Schornsteiner, E.; Mann, E.; Wagner, M.; Drillich, M.; Schmitz-Esser, S. Pyrosequencing reveals diverse fecal microbiota in Simmental calves during early development. Front. Microbiol. 2014, 5, 622. [Google Scholar] [CrossRef]
- Chen, H.; Liu, Y.; Huang, K.; Yang, B.; Zhang, Y.; Yu, Z.; Wang, J. Fecal microbiota dynamics and its relationship to diarrhea and health in dairy calves. J. Anim. Sci. Biotechnol. 2022, 13, 132. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Abenthum, A.; Matthes, J.M.; Kleeberger, D.; Ege, M.J.; Hölzel, C.; Bauer, J.; Schwaiger, K. Development and genetic influence of the rectal bacterial flora of newborn calves. Vet. Microbiol. 2012, 161, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; Collins, M.D. 16S rRNA gene similarities indicate that Hallella seregens (Moore and Moore) and Mitsuokella dentalis (Haapsalo et al.) are genealogically highly related and are members of the genus Prevotella: Emended description of the genus Prevotella (Shah and Collins) and description of Prevotella dentalis comb. nov. Int. J. Syst. Bacteriol. 1995, 45, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Trachsel, J.; Humphrey, S.; Allen, H.K. Butyricicoccus porcorum sp. nov., a butyrate-producing bacterium from swine intestinal tract. Int. J. Syst. Evol. Microbiol. 2018, 68, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Andoh, A. Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn’s Disease. Digestion 2016, 93, 59–65. [Google Scholar] [CrossRef]
- Foditsch, C.; Pereira, R.V.; Ganda, E.K.; Gomez, M.S.; Marques, E.C.; Santin, T.; Bicalho, R.C. Oral Administration of Faecalibacterium prausnitzii Decreased the Incidence of Severe Diarrhea and Related Mortality Rate and Increased Weight Gain in Preweaned Dairy Heifers. PLoS ONE 2015, 10, e0145485. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Nie, C.; Wu, Y.; Luo, R.; Chen, C.; Niu, J.; Zhang, W. Effects of two strains of Lactobacillus isolated from the feces of calves after fecal microbiota transplantation on growth performance, immune capacity, and intestinal barrier function of weaned calves. Front. Microbiol. 2023, 14, 1249628. [Google Scholar] [CrossRef] [PubMed]
- Foditsch, C.; Santos, T.M.; Teixeira, A.G.; Pereira, R.V.; Dias, J.M.; Gaeta, N.; Bicalho, R.C. Isolation and characterization of Faecalibacterium prausnitzii from calves and piglets. PLoS ONE 2014, 9, e116465. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Feng, S.; Wang, Z.; He, J.; Zhang, Z.; Zou, H.; Wu, Z.; Liu, X.; Wei, H.; Tao, S. Limosilactobacillus mucosae-derived extracellular vesicles modulates macrophage phenotype and orchestrates gut homeostasis in a diarrheal piglet model. NPJ Biofilms Microbiomes 2023, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, R.; Burdukiewicz, M.; Schierack, P. A systematic review and meta-analysis of the epidemiology of pathogenic Escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. coli. Front. Cell. Infect. Microbiol. 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Blanton, L.V.; Frese, S.A.; Charbonneau, M.; Mills, D.A.; Gordon, J.I. Cultivating healthy growth and nutrition through the gut microbiota. Cell 2015, 161, 36–48. [Google Scholar] [CrossRef] [PubMed]
- McNeil, N.I. The contribution of the large intestine to energy supplies in man. Am. J. Clin. Nutr. 1984, 39, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Muca, E.; Buonaiuto, G.; Lamanna, M.; Silvestrelli, S.; Ghiaccio, F.; Federiconi, A.; De, M.V.J.; Colleluori, R.; Fusaro, I.; Raspa, F.; et al. Reaching a Wider Audience: Instagram’s Role in Dairy Cow Nutrition Education and Engagement. Animals 2023, 13, 3503. [Google Scholar] [CrossRef]
- Muca, E.; Cavallini, D.; Raspa, F.; Bordin, C.; Bergero, D.; Valle, E. Integrating New Learning Methods into Equine Nutrition Classrooms: The Importance of Students’ Perceptions. J. Equine Vet. Sci. 2023, 126, 104537. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Group | 1 d | 3 d | 5 d | 7 d | 9 d | 12 d | 15 d | 18 d | 22 d | 26 d | 30 d | 35 d | 40 d |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 d | 0 | ||||||||||||
| 3 d | r = 0.398 * | 0 | |||||||||||
| 5 d | r = 0.402 * | r = −0.001 | 0 | ||||||||||
| 7 d | r = 0.340 * | r = 0.363 * | r = 0.328 * | 0 | |||||||||
| 9 d | r = 0.718 * | r = 0.544 * | r = 0.491 * | r = −0.021 | 0 | ||||||||
| 12 d | r = 0.861 * | r = 0.789 * | r = 0.667 * | r = 0.199 * | r = 0.137 * | 0 | |||||||
| 15 d | r = 0.889 * | r = 1 * | r = 0.988 * | r = 0.871 * | r = 0.954 * | r = 0.646 * | 0 | ||||||
| 18 d | r = 1 * | r = 1 * | r = 0.996 * | r = 0.947 * | r = 0.992 * | r = 0.845 * | r = 0.027 | 0 | |||||
| 22 d | r = 1 * | r = 0.985 * | r = 0.978 * | r = 0.925 * | r = 0.951 * | r = 0.801 * | r = 0.388 * | r = 0.122 | 0 | ||||
| 26 d | r = 0.985 * | r = 0.961 * | r = 0.962 * | r = 0.900 * | r = 0.934 * | r = 0.735 * | r = 0.314 * | r = 0.068 | r = −0.109 | 0 | |||
| 30 d | r = 1 * | r = 1 * | r = 0.996 * | r = 0.965 * | r = 0.996 * | r = 0.907 * | r = 0.525 * | r = 0.213 * | r = −0.034 | r = −0.634 | 0 | ||
| 35 d | r = 1 * | r = 1 * | r = 0.999 * | r = 0.996 * | r = 1 * | r = 0.969 * | r = 0.824 * | r = 0.422 * | r = 0.181 * | r = 0.007 | r = −0.047 | 0 | |
| 40 d | r = 1 * | r = 0.998 * | r = 0.991 * | r = 0.980 * | r = 0.996 * | r = 0.906 * | r = 0.611 * | r = 0.413 * | r = 0.252 * | r = 0.014 | r = 0.091 | r = 0.064 | 0 |
| Diarrhea Group | 1 d | 3 d | 5 d | 7 d | 9 d | 12 d | 15 d | 18 d | 22 d | 26 d | 30 d | 35 d | 40 d |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 d | 0 | ||||||||||||
| 3 d | r = 0.017 | 0 | |||||||||||
| 5 d | r = 0.362 * | r = 0.125 | 0 | ||||||||||
| 7 d | r = 0.533 * | r = 0.409 * | r = 0.022 | 0 | |||||||||
| 9 d | r = 0.635 * | r = 0.619 * | r = 0.024 | r = −0.192 | 0 | ||||||||
| 12 d | r = 0.861 * | r = 0.833 * | r = 0.709 * | r = 0.347 * | r = 0.128 | 0 | |||||||
| 15 d | r = 0.938 * | r = 1 * | r = 0.924 * | r = 0.728 * | r = 0.425 * | r = −0.099 | 0 | ||||||
| 18 d | r = 0.942 * | r = 0.976 * | r = 0.887 * | r = 0.728 * | r = 0.474 * | r = 0.039 | r = −0.110 | 0 | |||||
| 22 d | r = 0.983 * | r = 0.996 * | r = 0.979 * | r = 0.867 * | r = 0.648 * | r = 0.109 | r = 0.051 | r = −0.011 | 0 | ||||
| 26 d | r = 0.935 * | r = 0.976 * | r = 0.864 * | r = 0.715 * | r = 0.611 * | r = 0.094 | r = 0.132 | r = −0.057 | r = −0.185 | 0 | |||
| 30 d | r = 0.972 * | r = 0.968 * | r = 0.949 * | r = 0.888 * | r = 0.709 * | r = 0.240 * | r = 0.174 | r = −0.002 | r = −0.067 | r = −0.204 | 0 | ||
| 35 d | r = 0.998 * | r = 1 * | r = 0.997 * | r = 0.992 * | r = 0.898 * | r = 0.583 * | r = 0.462 * | r = 0.302 * | r = −0.019 | r = −0.093 | r = −0.004 | 0 | |
| 40 d | r = 0.959 * | r = 0.968 * | r = 0.947 * | r = 0.912 * | r = 0.863 * | r = 0.561 * | r = 0.581 * | r = 0.435 * | r = 0.146 | r = 0.056 | r = 0.105 | r = −0.098 | 0 |
| Day | Bacteria | Group | p-Value | |
|---|---|---|---|---|
| A | B | |||
| 1 d | L. johnsonii | 0.012 ± 0.010% | 0.030 ± 0.019% | 0.012 |
| Limosilactobacillus | 0 | 0.035 ± 0.021% | 0.046 | |
| P. mirabilis | 0.073 ± 0.071% | 0.043 ± 0.017% | 0.032 | |
| 3 d | P. russellii | 0.023 ± 0.018% | 1.330 ± 0.720% | 0.008 |
| F. prausnitzii | 0.029 ± 0.023% | 1.619 ± 1.461% | 0.014 | |
| E. ramosum | 0.778 ± 0.416% | 2.283 ± 0.611% | 0.023 | |
| L. johnsonii | 0.002 ± 0.002% | 1.208 ± 1.113% | 0.023 | |
| 5 d | F. umbilicata | 0 | 0.269 ± 0.159% | 0.010 |
| G. genomosp._3 | 0.057 ± 0.037% | 2.764 ± 1.556% | 0.019 | |
| C. pharyngocola | 0.309 ± 0.309% | 1.376 ± 0.609% | 0.040 | |
| 7 d | S. mitis | 0.380 ± 0.380% | 0.005 ± 0.008% | 0.013 |
| E. ramosum | 0.638 ± 0.252% | 0.087 ± 0.053% | 0.019 | |
| P. mirabilis | 0.180 ± 0.099% | 0.026 ± 0.017% | 0.019 | |
| A. muciniphila | 2.967 ± 2.760% | 0.004 ± 0.003% | 0.040 | |
| L. amylovorus | 0.011 ± 0.004% | 0.002 ± 0.001% | 0.040 | |
| 9 d | P. russellii | 0.032 ± 0.026% | 6.153 ± 5.149% | 0.004 |
| L. murinus | 8.123 ± 7.134% | 0.128 ± 0.127% | 0.010 | |
| F. necrophorum | 0.071 ± 0.042% | 0.001 ± 0.001% | 0.012 | |
| S. mitis | 0.012 ± 0.018% | 8.893 ± 0.003% | 0.012 | |
| A. muciniphila | 0.159 ± 0.096% | 0.008 ± 0.006% | 0.020 | |
| B. vulgatus | 16.951 ± 7.283% | 4.473 ± 2.922% | 0.020 | |
| L. johnsonii | 0.262 ± 0.253% | 2.409 ± 1.656% | 0.020 | |
| P. dorei | 0.003 ± 0.003% | 0.005 ± 0.005% | 0.020 | |
| K. pneumoniae | 1.384 ± 1.285% | 0.003 ± 0.002% | 0.028 | |
| E. coli | 27.465 ± 11.439% | 52.753 ± 7.410% | 0.039 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Na, M.; Jia, S.; Sun, M.; Gao, S.; Pan, S.; Dong, W.; Song, Y.; Yang, J. Dynamic Analysis of Stool Microbiota of Simmental Calves and Effects of Diarrhea on Their Gut Microbiota. Biology 2024, 13, 520. https://doi.org/10.3390/biology13070520
Wang Q, Na M, Jia S, Sun M, Gao S, Pan S, Dong W, Song Y, Yang J. Dynamic Analysis of Stool Microbiota of Simmental Calves and Effects of Diarrhea on Their Gut Microbiota. Biology. 2024; 13(7):520. https://doi.org/10.3390/biology13070520
Chicago/Turabian StyleWang, Qianxun, Mula Na, Shiyu Jia, Miao Sun, Song Gao, Shiwei Pan, Wu Dong, Yang Song, and Jingfeng Yang. 2024. "Dynamic Analysis of Stool Microbiota of Simmental Calves and Effects of Diarrhea on Their Gut Microbiota" Biology 13, no. 7: 520. https://doi.org/10.3390/biology13070520
APA StyleWang, Q., Na, M., Jia, S., Sun, M., Gao, S., Pan, S., Dong, W., Song, Y., & Yang, J. (2024). Dynamic Analysis of Stool Microbiota of Simmental Calves and Effects of Diarrhea on Their Gut Microbiota. Biology, 13(7), 520. https://doi.org/10.3390/biology13070520