
Paraphoma chrysanthemicola Affects the Carbohydrate and Lobetyolin Metabolism Regulated by Salicylic Acid in the Soilless Cultivation of Codonopsis pilosula


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Treatments and Strain Preservation
2.2. Co-Cultivation between Fungus and Plants
2.3. Determination of Morphological Indicators
2.4. Detection of Chlorophyll and Carbohydrate Content
2.5. Detection of Soluble Protein and Lobetyolin Content
2.6. Analysis of Endogenous Signalling Molecules
2.7. Detection of Viability of the Antioxidant Enzyme Lines and Detection of Metabolite Enzyme Lines Related to Metabolite Synthesis
2.8. Data Processing and Statistical Analysis
3. Results
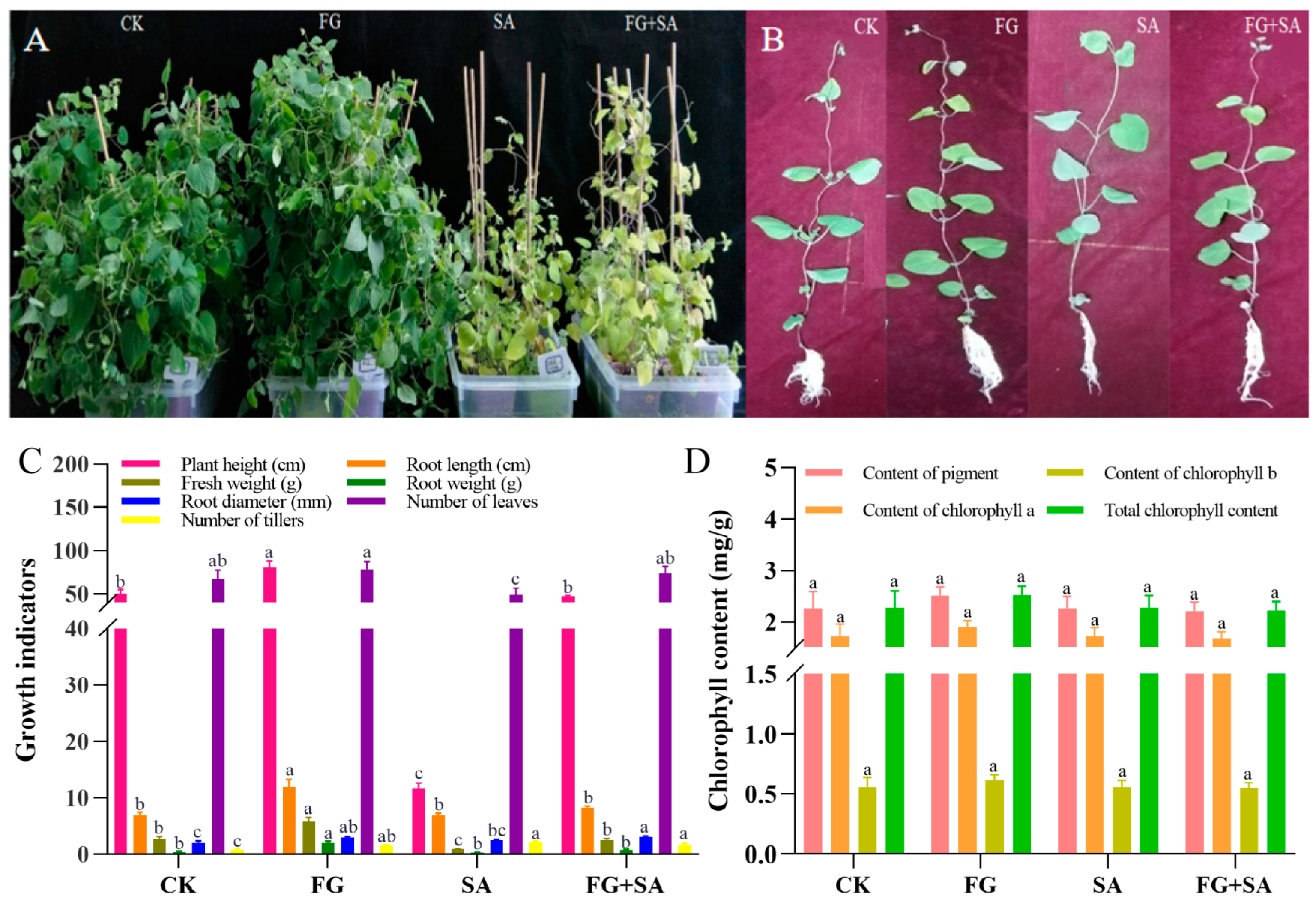
3.1. Effects on C. pilosula Seedling Growth and Total Chlorophyll Concentration
3.2. Effects of Different Treatments on Carbohydrate Content in C. pilosula
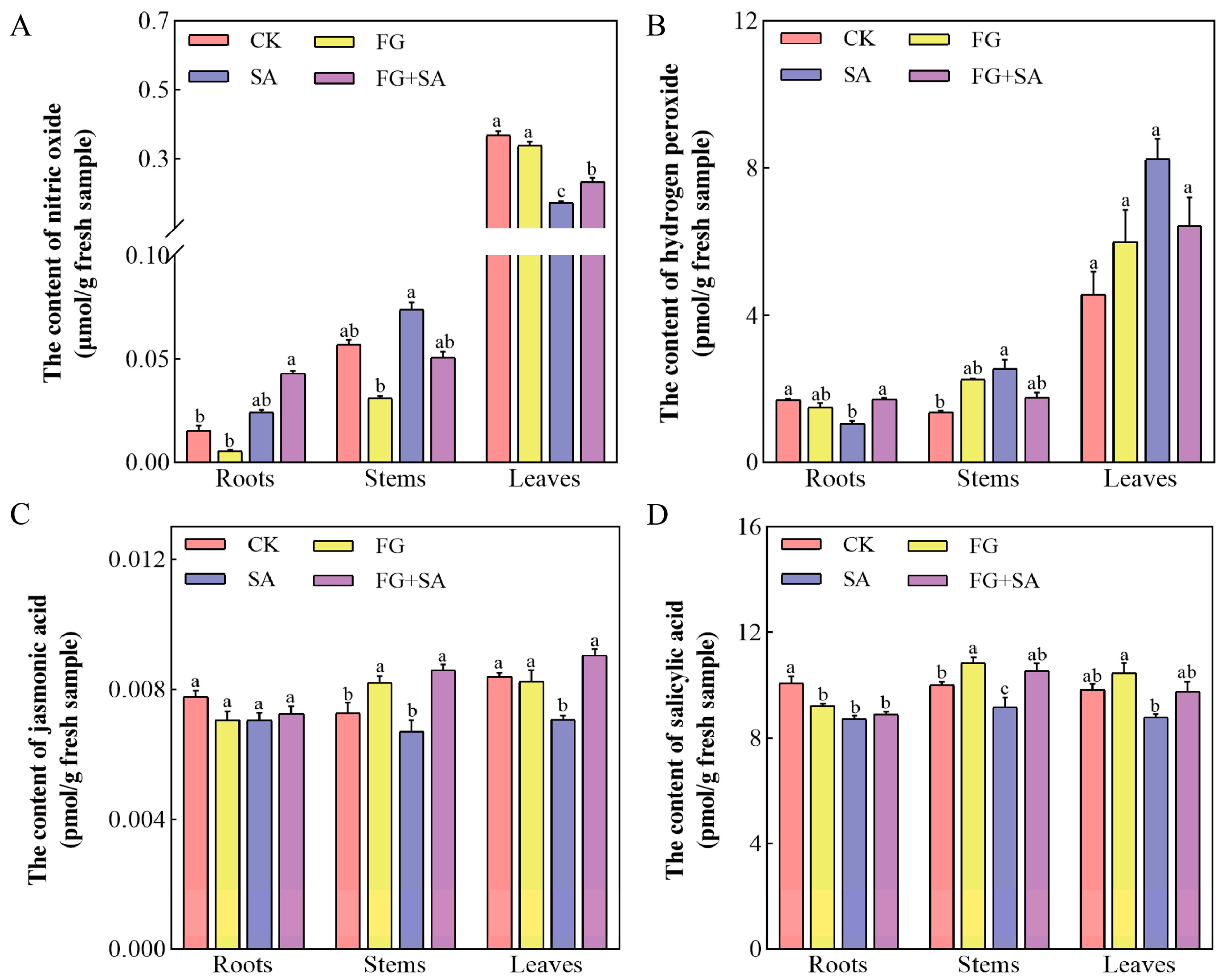
3.3. Effects of Different Treatments on the Accumulation of Signal Molecules in C. pilosula
3.4. Effects on the Accumulation of Soluble Protein and Lobetyolin Content
3.5. Effects of Different Treatments on Antioxidase-like Activity in C. pilosula
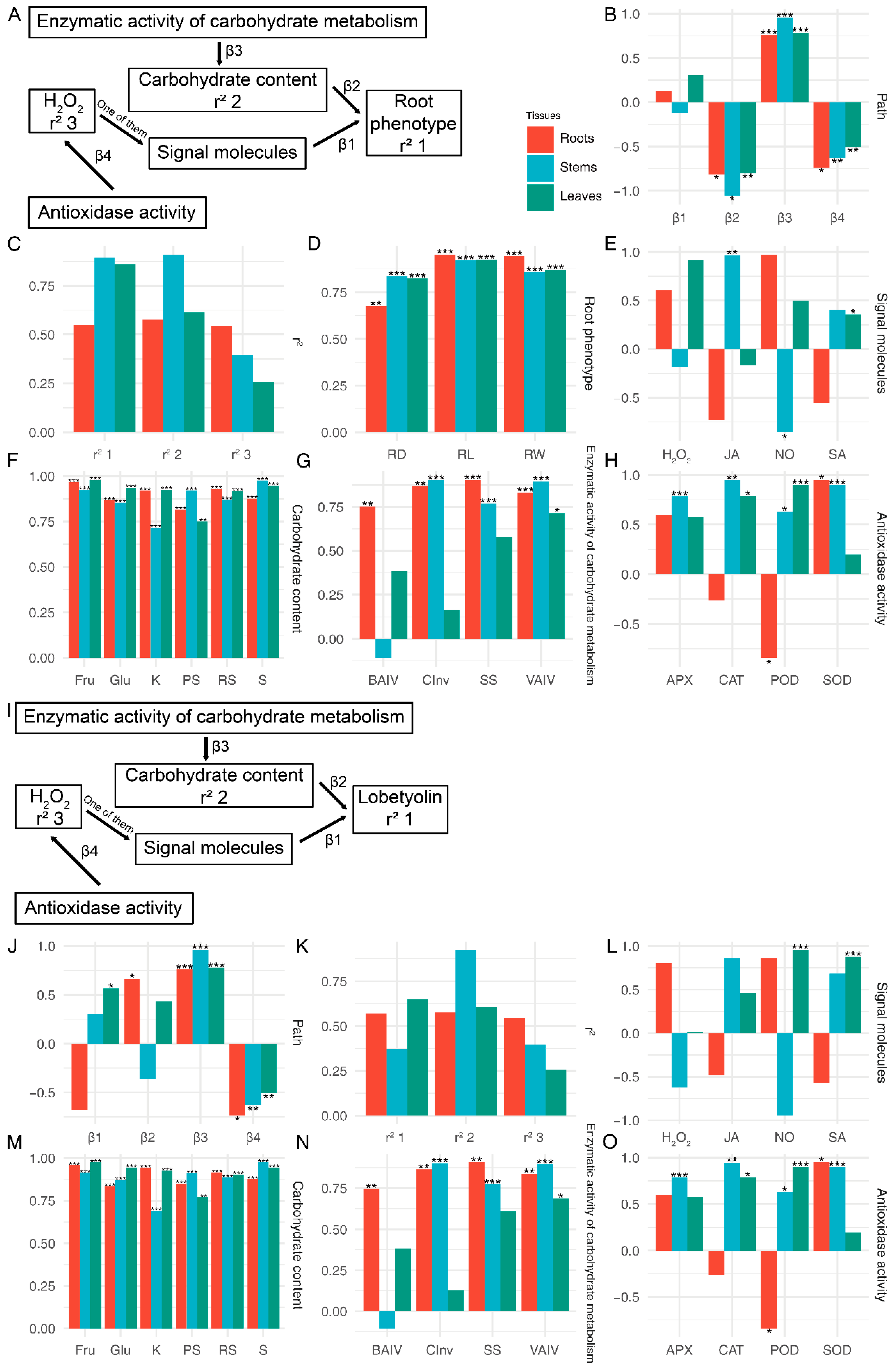
3.6. Effects on Enzymatic Activity Related to Glucose Metabolism in C. pilosula under Treatment of P. chrysanthemicola and Salicylic Acid
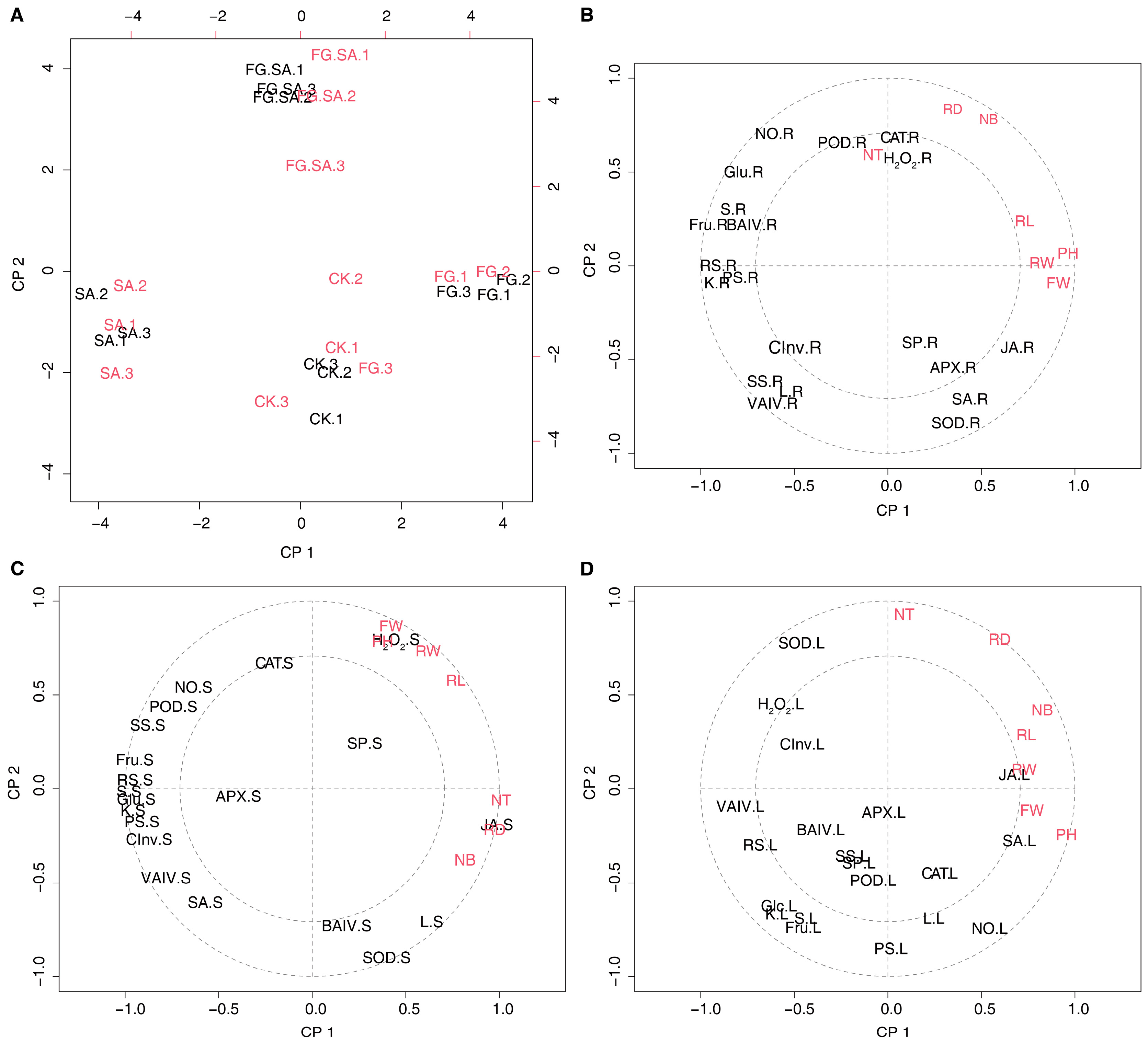
3.7. Correlation Analysis of the Physiological and Biochemical Indexes of C. pilosula
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Chen, L.; Xin, H.; Zheng, C.; Rahman, K.; Han, T.; Qin, L. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef]
- Tian, Y.; Fu, X.; Zhang, G.Y.; Zhang, R.; Kang, Z.; Gao, K.; Mendgen, K.W. Mechanisms in growth-promoting of cucumber by the endophytic fungus Chaetomium globosum strain ND35. J. Fungi 2022, 8, 180. [Google Scholar] [CrossRef] [PubMed]
- González-Guzmán, A.; Rey, M.-D.; Froussart, E.; Quesada-Moraga, E. Elucidating the effect of endophytic entomopathogenic fungi on bread wheat growth through signaling of immune response-related hormones. Appl. Environ. Microbiol. 2022, 88, e0088222. [Google Scholar] [CrossRef] [PubMed]
- Roitsch, T.G.; González, M.C. Function and regulation of plant invertases: Sweet sensations. Trends Plant Sci. 2004, 9, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, L.; Calderon, C.; Medina, L.M.; Bahamon, I.; Cardenas, M.; Bernal, A.J.; Gonzalez, A.; Restrepo, S. Characterization of cellulases of fungal endophytes isolated from Espeletia spp. J. Microbiol. 2012, 50, 1009–1013. [Google Scholar] [CrossRef]
- Zhai, X.; Jia, M.; Chen, L.; Zheng, C.; Rahman, K.; Han, T.; Qin, L. The regulatory mechanism of fungal elicitor-induced secondary metabolite biosynthesis in medical plants. Crit. Rev. Microbiol. 2017, 43, 238–261. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, S.; Liu, H.; Qiu, D. The role of radical burst in plant defense responses to necrotrophic fungi. J. Integr. Agric. 2012, 11, 1305–1312. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, J.; Li, S.; Hui, L.; Li, Y.; Chen, K.; Meng, T.; Yu, C.; Leng, F.; Ma, J. Comparative analysis of carbon and nitrogen metabolism, antioxidant indexes, polysaccharides and lobetyolin changes of different tissues from Codonopsis pilosula co-inoculated with Trichoderma. J. Plant Physiol. 2021, 267, 153546. [Google Scholar] [CrossRef]
- Shields, A.R.; Shivnauth, V.; Castroverde, C.D.M. Salicylic acid and N-hydroxypipecolic acid at the fulcrum of the plant immunity-growth equilibrium. Front. Plant Sci. 2022, 13, 841688. [Google Scholar] [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Dempsey, D.M.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Benediktyová, Z.; Matous, K.; Bonfig, K.B.; Mueller, M.J.; Nedbal, L.; Roitsch, T.G. Visualization of dynamics of plant-pathogen interaction by novel combination of chlorophyll fluorescence imaging and statistical analysis: Differential effects of virulent and avirulent strains of P. syringae and of oxylipins on A. thaliana. J. Exp. Bot. 2007, 58, 797–806. [Google Scholar] [CrossRef]
- Benjamin, G.; Pandharikar, G.; Frendo, P. Salicylic acid in plant symbioses: Beyond plant pathogen interactions. Biology 2022, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- Dastogeer, K.M.G.; Li, H.; Sivasithamparam, K.; Jones, M.G.K.; Du, X.; Ren, Y.; Wylie, S.J. Metabolic responses of endophytic Nicotiana benthamiana plants experiencing water stress. Environ. Exp. Bot. 2017, 143, 59–71. [Google Scholar] [CrossRef]
- De Rocchis, V.; Jammer, A.; Camehl, I.; Franken, P.; Roitsch, T. Tomato growth promotion by the fungal endophytes Serendipita indica and Serendipita herbamans is associated with sucrose de-novo synthesis in roots and differential local and systemic effects on carbohydrate metabolisms and gene expression. J. Plant Physiol. 2022, 276, 153755. [Google Scholar] [CrossRef] [PubMed]
- Vukelić, I.D.; Prokić, L.T.; Racić, G.M.; Pešić, M.B.; Bojović, M.M.; Sierka, E.M.; Panković, D.M. Effects of Trichoderma harzianum on photosynthetic characteristics and fruit quality of tomato plants. Int. J. Mol. Sci. 2021, 22, 6961. [Google Scholar] [CrossRef]
- Dong, C.; Wang, X.; Shang, Q. Salicylic acid regulates sugar metabolism that confers tolerance to salinity stress in cucumber seedlings. Sci. Hortic. 2011, 129, 629–636. [Google Scholar] [CrossRef]
- García-Rodríguez, S.; Azcón-Aguilar, C.; Ferrol, N. Transcriptional regulation of host enzymes involved in the cleavage of sucrose during arbuscular mycorrhizal symbiosis. Physiol. Plant. 2007, 129, 737–746. [Google Scholar] [CrossRef]
- Liang, Y.; Wei, G.; Ning, K.; Zhang, G.; Liu, Y.; Dong, L.; Chen, S. Contents of lobetyolin, syringin, and atractylolide III in Codonopsis pilosula are related to dynamic changes of endophytes under drought stress. Chin. Med. 2021, 16, 122. [Google Scholar] [CrossRef]
- Han, Q.; Wu, Y.; Gao, H.; Xu, R.; Paré, P.W.; Shi, H.; Zhao, Q.; Li, H.R.; Khan, S.A.; Wang, Y.; et al. Improved salt tolerance of medicinal plant Codonopsis pilosula by Bacillus amyloliquefaciens GB03, Acta Physiol. Plant. 2016, 39, 11770534. [Google Scholar]
- Yue, J.; Ni, Y.; Jiang, M.; Chen, H.; Chen, P.; Liu, C. Characterization of Codonopsis pilosula subsp. Tangshen plastome and comparative analysis of Codonopsis species. PLoS ONE 2022, 17, e0271813. [Google Scholar]
- Zou, Y.; Zhang, Y.; Paulsen, B.S.; Fu, Y.; Huang, C.; Feng, B.; Li, L.; Chen, X.; Jia, R.; Song, X.; et al. Prospects of Codonopsis pilosula polysaccharides: Structural features and bioactivities diversity. Trends Food Sci. Technol. 2020, 103, 1–11. [Google Scholar] [CrossRef]
- Gao, S.; Liu, J.; Wang, M.; Cao, T.; Qi, Y.-D.; Zhang, B.; Sun, X.; Liu, H.; Xiao, P. Traditional uses, phytochemistry, pharmacology and toxicology of Codonopsis: A review. J. Ethnopharmacol. 2018, 219, 50–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xia, Y.; Luo, H.; Huang, S.; Wang, Y.; Shentu, Y.; Mahaman, Y.A.R.; Huang, F.; Ke, D.; Wang, Q.; et al. Codonopsis pilosula polysaccharide attenuates tau hyperphosphorylation and cognitive impairments in hTau infected mice. Front. Mol. Neurosci. 2018, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Paidi, M.K.; Agarwal, P.; More, P.; Agarwal, P.K. Chemical derivatization of metabolite mass profiling of the recretohalophyte Aeluropus lagopoides revealing salt stress tolerance mechanism. Mar. Biotechnol. 2017, 19, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Sobhanian, H.; Motamed, N.; Jazii, F.R.; Nakamura, T.; Komatsu, S. Salt stress induced differential proteome and metabolome response in the shoots of Aeluropus lagopoides (poaceae), a halophyte C4 plant. J. Proteome Res. 2010, 9, 2882–2897. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Cheng, J.; Zhao, G.; Liu, H.; Zhang, L.; Yang, C. Natural wax from non-medicinal aerial part of Codonopsis pilosula as a biolubricant. J. Clean. Prod. 2020, 242, 118403. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Y.; Guo, F.; Huang, B.; Okyere, S.A.; Wang, H. Comparative analysis of chemical composition, antioxidant and antimicrobial activities of leaves, leaf tea and root from Codonopsis pilosula. Ind. Crops Prod. 2019, 142, 111844. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, Y.; Paulsen, B.S.; Rise, F.; Chen, Z.; Jia, R.; Li, L.; Song, X.; Feng, B.; Tang, H.; et al. Structural features of pectic polysaccharides from stems of two species of radix Codonopsis and their antioxidant activities. Int. J. Biol. Macromol. 2020, 159, 704–713. [Google Scholar] [CrossRef]
- de Gruyter, J.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Systematic reappraisal of species in Phoma section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 2010, 102, 1066–1081. [Google Scholar] [CrossRef]
- Boerema, G.H.; Gruyter, J.D.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual: Differentiation of Specific and Infra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- Sun, W.; Feng, M.; Zhu, N.; Leng, F.; Yang, M.; Wang, Y. Genomic characteristics and comparative genomics analysis of the endophytic fungus Paraphoma chrysanthemicola DS-84 isolated from Codonopsis pilosula root. J. Fungi 2023, 9, 1022. [Google Scholar] [CrossRef]
- Li, Y.Y. Alleviating effect of exogenous salicylic acid on low temperature stress of Saussurea psammophila; Northwest Normal University: Lanzhou, China, 2017. [Google Scholar]
- Wang, L. Effects of exogenous calcium chloride and salicylic acid on cold resistance of different grape varieties; Xinjiang Agricultural University: Xinjiang, China, 2005. [Google Scholar]
- Liu, Y.Y. Study on mechanism of exogenous salicylic acid and calcium chloride to improve salt tolerance of wetland plant Acorus calamus; Qingdao University: Qingdao, China, 2021. [Google Scholar]
- Ding, M. Effect of Paraphoma chrysanthemicola on the growth and active component accumulation of Codonopsis pilosula (Franch.) Nannf. mediated by salicylic acid; Lanzhou University of Technology: Lanzhou, China, 2020. [Google Scholar]
- Pal, P.K.; Prasad, R.; Singh, R.D. Evaluating the non-destructive method for determining the chlorophyll and nitrogen content in Stevia rebaudiana (bertoni) leaf. Plant Biosyst. Int. J. Deal. Asp. Plant Biosyst. 2013, 149, 131–135. [Google Scholar]
- Kondo, M.; Mulianda, R.; Matamura, M.; Shibata, T.; Mishima, T.; Jayanegara, A.; Isono, N. Validation of a phenol-sulfuric acid method in a microplate format for the quantification of soluble sugars in ruminant feeds. Anim. Sci. J. 2021, 92, e13530. [Google Scholar] [CrossRef]
- Naz, S.; Uroos, M.; Asim, A.M.; Muhammad, N.; Shah, F.U. One-pot deconstruction and conversion of lignocellulose into reducing sugars by pyridinium-based ionic liquid–metal salt system. Front. Chem. 2020, 8, 236. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, J.; Wang, H.; Li, Q.; Wei, S.A.; Wang, D. Effects of metal ions on the hydrolysis of bamboo biomass in 1-butyl-3-methylimidazolium chloride with dilute acid as catalyst. Bioresour. Technol. 2014, 173, 399–405. [Google Scholar] [CrossRef]
- Jalaludin, I.; Kim, J. Comparison of ultraviolet and refractive index detections in the HPLC analysis of sugars. Food Chem. 2021, 365, 130514. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Qi, P.; Guo, L. A new approach for economical pretreatment of corncobs. Appl. Sci. 2019, 9, 504. [Google Scholar] [CrossRef]
- He, J.Y.; Zhu, S.; Komatsu, K. HPLC/UV analysis of polyacetylenes, phenylpropanoid and pyrrolidine alkaloids in medicinally used Codonopsis species. Phytochem. Anal. 2014, 25, 213–219. [Google Scholar] [CrossRef]
- Liu, R.; Shi, L.; Zhu, T.; Yang, T.; Ren, A.; Zhu, J.; Zhao, M.W. Cross talk between nitric oxide and calcium-calmodulin regulates ganoderic acid biosynthesis in Ganoderma lucidum under heat stress. Appl. Environ. Microbiol. 2018, 84, e00043-18. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Wang, X.; Ding, Y. Detection of seven phytohormones in peanut tissues by ultra-high-performance liquid chromatography-triple quadrupole tandem mass spectrometry. J. Integr. Agric. 2020, 19, 700–708. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.; Lin, W.; Tong, J.; Huang, Z.; Xiao, L. Determination of both jasmonic acid and methyl jasmonate in plant samples by liquid chromatography tandem mass spectrometry. Chin. Sci. Bull. 2010, 55, 2231–2235. [Google Scholar] [CrossRef]
- Li, S.; Xue, L.; Xu, S.; Feng, H.; An, L. Hydrogen peroxide acts as a signal molecule in the adventitious root formation of mung bean seedlings. Environ. Exp. Bot. 2009, 65, 63–71. [Google Scholar] [CrossRef]
- Cui, J.; Wang, Y.; Jiao, J.; Gong, Y.; Wang, J.; Wang, M. Fungal endophyte-induced salidroside and tyrosol biosynthesis combined with signal cross-talk and the mechanism of enzyme gene expression in Rhodiola crenulata. Sci. Rep. 2017, 7, 12540. [Google Scholar] [CrossRef]
- Bastías, D.A.; Alejandra Martínez-Ghersa, M.; Newman, J.A.; Card, S.D.; Mace, W.J.; Gundel, P.E. The plant hormone salicylic acid interacts with the mechanism of anti-herbivory conferred by fungal endophytes in grasses. Plant Cell Environ. 2018, 41, 395–405. [Google Scholar] [CrossRef]
- Stamp, N. Out of the quagmire of plant defense hypotheses. Q. Rev. Biol. 2003, 78, 23–55. [Google Scholar] [CrossRef]
- Cheng, F.; Zamski, E.; Guo, W.; Pharr, D.M.; Williamson, J.D. Salicylic acid stimulates secretion of the normally symplastic enzyme mannitol dehydrogenase: A possible defense against mannitol-secreting fungal pathogens. Planta 2009, 230, 1093–1103. [Google Scholar] [CrossRef]
- Dong, C.; Cao, N.; Li, L.; Shang, Q. Quantitative proteomic profiling of early and late responses to salicylic acid in cucumber leaves. PLoS ONE 2016, 11, e0161395. [Google Scholar] [CrossRef]
- Shaki, F.; Maboud, H.E.; Niknam, V. Differential proteomics: Effect of growth regulators on salt stress responses in safflower seedlings. Pestic. Biochem. Physiol. 2020, 164, 149–155. [Google Scholar] [CrossRef]
- Bernard, F.; Baghai, M.; Hadad, K.S. In vitro carbohydrate stress: Salicylic acid increases soluble invertase activity in Pistacia vera L. in vitro plantlets. Iran. J. Plant Physiol. 2012, 2, 355–360. [Google Scholar]
- Zhou, Y.; Liu, C.; Li, G.; Hao, L. Ethylene insensitive mutation increases Arabidopsis tolerance to Cd in NPR1-dependent manner. Plant Soil 2019, 441, 49–69. [Google Scholar] [CrossRef]
- Rasmussen, S.; Liu, Q.; Parsons, A.J.; Xue, H.; Sinclair, B.; Newman, J.A. Grass–endophyte interactions: A note on the role of monosaccharide transport in the Neotyphodium lolii–Lolium perenne symbiosis. New Phytol. 2012, 196, 7–12. [Google Scholar] [CrossRef]
- Sun, K.; Zhang, W.; Yuan, J.; Song, S.L.; Wu, H.; Tang, M.J.; Dai, C.C. Nitrogen fertilizer-regulated plant-fungi interaction is related to root invertase-induced hexose generation. FEMS Microbiol. Ecol. 2020, 96, fiaa139. [Google Scholar] [CrossRef]
- Höke, D.; Dräger, B. Calystegines in Calystegia sepium do not inhibit fungal growth and invertase activity but interact with plant invertase. Plant Biol. 2004, 6, 206–213. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.J. Endophytic fungus Aspergillus japonicus mediates host plant growth under normal and heat stress conditions. BioMed Res. Int. 2018, 2018, 7696831. [Google Scholar]
- Campbell, I.M. Secondary metabolism and microbial physiology. Adv. Microb. Physiol. 1985, 25, 1–60. [Google Scholar]
- Ji, Z.; Yu, M.; Ding, Y.; Li, J.; Zhu, F.; He, J.; Yang, L. Coiled-coil N21 of Hpa1 in Xanthomonas oryzae pv. oryzae promotes plant growth, disease resistance and drought tolerance in non-hosts via eliciting HR and regulation of multiple defense response genes. Int. J. Mol. Sci. 2020, 22, 203. [Google Scholar]
- Praveen, N.; Murthy, H.N. Synthesis of withanolide a depends on carbon source and medium pH in hairy root cultures of Withania somnifera. Ind. Crops Prod. 2012, 35, 241–243. [Google Scholar] [CrossRef]
- Shi, X.; Qin, T.; Liu, H.; Wu, M.; Li, J.; Shi, Y.; Gao, Y.; Ren, A. Endophytic fungi activated similar defense strategies of Achnatherum sibiricum host to different trophic types of pathogens. Front. Microbiol. 2020, 11, 1607. [Google Scholar] [CrossRef]
- Pozo, M.J.; López-Ráez, J.A.; Azcón-Aguilar, C.; García-Garrido, J.M. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Beltrán-Peña, E.; Herrera-Estrella, A.; López-Bucio, J. Trichoderma-induced plant immunity likely involves both hormonal- and camalexin-dependent mechanisms in Arabidopsis thaliana and confers resistance against necrotrophic fungi Botrytis cinerea. Plant Signal. Behav. 2014, 6, 1554–1563. [Google Scholar] [CrossRef] [PubMed]
- Khokon, M.A.R.; Okuma, E.; Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of extracellular oxidative burst in salicylic acid-induced stomatal closure in Arabidopsis. Plant Cell Environ. 2011, 34, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Tada, Y. Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 2014, 5, 70455. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Waqas, M.; Hamayun, M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.J. Co-synergism of endophyte Penicillium resedanum Lk6 with salicylic acid helped Capsicum annuum in biomass recovery and osmotic stress mitigation. BMC Microbiol. 2013, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate peroxidase and catalase activities and their genetic regulation in plants subjected to drought and salinity stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef] [PubMed]
- Ben Hassena, A.; Zouari, M.; Labrousse, P.; Decou, R.; Soua, N.; Khabou, W.; Zouari, N. Effect of arbuscular myccorhizal fungi on soil properties, mineral nutrition and antioxidant enzymes of olive plants under treated wastewater irrigation. S. Afr. J. Bot. 2022, 148, 710–719. [Google Scholar] [CrossRef]
- Chakraborty, N. Salicylic acid and nitric oxide cross-talks to improve innate immunity and plant vigor in tomato against Fusarium oxysporum stress. Plant Cell Rep. 2021, 40, 1415–1427. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Luo, C.; Wu, Y.; Ding, M.; Feng, M.; Leng, F.; Wang, Y. Paraphoma chrysanthemicola Affects the Carbohydrate and Lobetyolin Metabolism Regulated by Salicylic Acid in the Soilless Cultivation of Codonopsis pilosula. Biology 2024, 13, 408. https://doi.org/10.3390/biology13060408
Sun W, Luo C, Wu Y, Ding M, Feng M, Leng F, Wang Y. Paraphoma chrysanthemicola Affects the Carbohydrate and Lobetyolin Metabolism Regulated by Salicylic Acid in the Soilless Cultivation of Codonopsis pilosula. Biology. 2024; 13(6):408. https://doi.org/10.3390/biology13060408
Chicago/Turabian StyleSun, Wenbin, Caiming Luo, Yamiao Wu, Miao Ding, Min Feng, Feifan Leng, and Yonggang Wang. 2024. "Paraphoma chrysanthemicola Affects the Carbohydrate and Lobetyolin Metabolism Regulated by Salicylic Acid in the Soilless Cultivation of Codonopsis pilosula" Biology 13, no. 6: 408. https://doi.org/10.3390/biology13060408
APA StyleSun, W., Luo, C., Wu, Y., Ding, M., Feng, M., Leng, F., & Wang, Y. (2024). Paraphoma chrysanthemicola Affects the Carbohydrate and Lobetyolin Metabolism Regulated by Salicylic Acid in the Soilless Cultivation of Codonopsis pilosula. Biology, 13(6), 408. https://doi.org/10.3390/biology13060408