Simple Summary
In order to investigate the effects of low oxygen stress on the survival rate, antioxidant, anaerobic metabolic enzymes, and related genes of crayfish during aquaculture. This experiment conducted a 24-h low oxygen stress experiment on juvenile and subadult crayfish, and the results showed that the higher the degree of low oxygen stress, the higher the antioxidant and anaerobic metabolic enzyme activity and gene expression levels of juvenile crayfish and subadult crayfish. Compared with subadult crayfish, the antioxidant system of crayfish larvae is more sensitive to hypoxic environments.
Abstract
The red swamp crayfish Procambarus clarkii is the most reared shrimp in China, but it is often affected by hypoxia stress in the process of seedling culture and adult crayfish culture. The oxygen consumption rate and asphyxiation point of juvenile crayfish (1.17 ± 0.03 g) and subadult crayfish (11.68 ± 0.11 g) at different temperatures (20, 22, 24, 26, and 28 °C) were studied. The survival, glycolysis, and expression of antioxidant genes were compared under 24 h acute hypoxia stress (1, 2, and 3 mg/L) and normal dissolved oxygen (7.5 mg/L). The results showed that the oxygen consumption rate and asphyxiation point of juvenile and subadult crayfish increased with increasing temperatures (20–28 °C). At the same temperature, the oxygen consumption rate and asphyxiation point of juvenile crayfish were significantly higher than those of subadult crayfish (p < 0.05). Within 24 h, the three hypoxia stress environments did not lead to the death of crayfish, indicating that P. clarkii has a strong ability to adapt to hypoxia. Hypoxia stress significantly affected the activities of antioxidant and anaerobic metabolic enzymes and gene expression in juvenile and subadult crayfish. The activities of the superoxide dismutase (SOD), catalase (CAT), and lactate dehydrogenase (LDH) and the content of lactic acid (LD) in the hepatopancreas of juvenile and subadult crayfish in the hypoxia stress groups increased significantly. The expression levels of SOD mRNA, CAT mRNA, Hsp70 mRNA, and crustin 4 mRNA in the hepatopancreas of juvenile and subadult crayfish in the hypoxia stress groups were significantly higher than those in the control group (p < 0.05), and the higher the degree of hypoxia stress, the higher the expression of each gene. The results showed that the antioxidant system of juvenile crayfish was more sensitive to hypoxia environments, and hypoxia stress resulted in increased stress levels in juvenile crayfish and subadult crayfish.
1. Introduction
Red swamp crayfish (Procambarus clarkii) originated in the south-central United States and northeastern Mexico and were introduced into China from Japan in the 1930s [1]. Due to their rapid growth and high nutritional and commercial value, artificial culture of P. clarkii developed rapidly from the 1980s to 2020s, and their distribution rapidly expanded to more than 20 provinces in China, becoming the crustacean with the highest yield in China and the second highest yield in the world [2,3]. In 2021, the total area of crayfish culture in China reached 1.73 million ha, with an output of 2.63 million tons, accounting for 8.27% of China’s total output of freshwater aquaculture. The output value of the aquaculture industry was approximately 11.76 billion USD [4]. At present, the main mode of Procambarus clarkii (P. clarkii) culture in China is integrated rich crayfish. However, problems such as high mortality and slow growth often occur in the process of crayfish culture in rice fields, affecting the economic benefits to farmers and restricting the sustainable development of crayfish culture.
There are a variety of factors affecting the mortality and growth rate of aquatic animals, including biological and abiotic factors. Biological factors mainly include predators, competitive species, and food resources. Abiotic factors include feed feeding, stocking density, and culture environment, among which the culture environment is considered to be one of the important factors affecting the survival and growth of crustaceans [5,6]. Previous studies have shown that the deterioration of the quality of the aquaculture environment poses a great challenge to the survival of crustaceans, including temperature stress [7], pH stress [8], ammonia stress [9], hypoxia stress [10], and other factors that often lead to disease and death in crustaceans.
In an integrated rice and crayfish cultivation system, the use of fertilizer in the process of rice planting leads to the enrichment of nutrients in the sediment. The decomposition of straw returned to the field after a rice harvest also leads to an increase in nutrients in the water and a rapid change in the physical and chemical indices of water quality. Therefore, the phenomena of low pH, high nitrite, and low dissolved oxygen often occur during the culture of P. clarkii in rice fields [6,11]. France found that the tolerance of juvenile Orconectes virilis to pH changes was lower than that of adult individuals, which led to the death of juvenile crayfish when the pH was lower than 5.5 [12]. Excessive nitrite nitrogen will reduce the oxygen carrying capacity of blood, resulting in hypoxia and even pathological changes in tissues and organs [6]. Nitrite stress can decrease the activity of SOD and CAT, decrease antioxidant capacity, and affect the immune function of P. clarkii [13]. Hypoxia stress can significantly affect the growth, metabolism, and antioxidant systems of Litopenaeus vannamei (L. vannamei) [14], Penaeus vannamei [15], and Macrobrachium rosenbergii [16]. However, there are few studies on the adaptability to hypoxia of P. clarkii and the effects of hypoxia stress on the immune antioxidant system and related gene expression.
Crustaceans mainly show innate immune responses to hypoxia stress, including physical defense and humoral immunity [17,18]. Previous studies have shown that hypoxia stress can increase the content of superoxide anions in the hepatopancreas and the activity of SOD in the gills and hepatopancreas of L. vannamei [14,19]. Moreover, the expression levels of LvGAPDH mRNA and glyceraldehyde-3-phosphate dehydrogenase in gill tissue and phosphofructokinase and fructose-1,6-diphosphatase in the hepatopancreas of L. vannamei were significantly increased [14]. Short-term (≤7 days) hypoxia stress can increase the expression of HIF-1a and enhance the antioxidant response in the hepatopancreas and midgut of L. vannamei, while long-term (≥14 days) cyclic hypoxia stress can destroy the cellular antioxidant mechanism in the hepatopancreas and midgut, inhibit the antioxidant response, and aggravate cell apoptosis and histopathological lesions in the hepatopancreas and midgut [20]. Under hypoxic conditions, the intestinal flora and mucosal morphology of Macrobrachium nipponense (M. nipponense) changed, and the intestinal immune enzyme activity decreased [21]. The enzyme activity related to glucose anaerobic glycolysis increased, the mRNA expression level of aerobic respiration decreased, and the normal metabolic activity changed from aerobic respiration to anaerobic respiration, which disturbed the energy metabolism of M. nipponense and affected the defense regulation of the antioxidant system under a hypoxia environment [13,16,22]. These studies show that hypoxia stress can significantly affect the immune and antioxidant capacity of crustaceans and ultimately affect their normal survival and growth.
Thus, it was hypothesized that the activities of antioxidant enzymes, those of anaerobic respiratory enzymes, and the heat stress gene expression levels of P. clarkii increased under anoxic environments, and there were differences between juvenile and subadult P. clarkii. In this study, to explore the hypoxia adaptability of P. clarkii and to understand the changes in the antioxidant system and anaerobic metabolic enzymes under hypoxic conditions, we measured the respiratory rate and asphyxiation point of juvenile and subadult P. clarkii at different temperatures. The expression levels of antioxidant enzymes, anaerobic metabolic enzymes, and antioxidant and stress genes in the hepatopancreas of juvenile and subadult P. clarkii under different hypoxia stresses were studied. The differences in the antioxidant systems and anaerobic metabolic enzymes between juvenile crayfish and subadult crayfish under the same hypoxic stress were compared. The results of this investigation help to reveal the physiological and biochemical reactions of P. clarkii under hypoxic conditions, providing a theoretical basis for improving the survival rate of P. clarkii in farming.
2. Materials and Methods
2.1. Experimental Animals and Ethics
Cultured juvenile crayfish (1.05 ± 0.05 g) and subadult crayfish (11.21 ± 0.15 g) were obtained from the crayfish breeding farm of Yangtze University in Jingzhou city, Hubei Province, China. After capture, crayfish were immediately transferred to the laboratory. P. clarkii were raised separately in six glass tanks (130 cm × 54 cm × 30 cm) for 14 d to acclimate to laboratory conditions before the experiment. During acclimation, the six glass tanks were filled with 20 cm aerated tap water, and PVC pipes and artificial macrophytes (resembling Hydrilla verticillata) were used as refuges for crayfish. The initial water temperature was the same as that of the farm, and after 7 d of adaptation, it was then increased by 0.5–1 °C every day to reach the experimental temperature of 24 °C. Crayfish were fed a commercial diet (crude protein content 34% and crude fat content 3%) twice at 8:00 h and 18:00 h every day. The food residuals were removed two hours after feeding. The temperature, pH, and dissolved oxygen were measured twice a day with a YSI ProPlus handheld water quality meter (Thermo Fisher Scientific Company, Waltham, MA, USA), with average values of 24.2 ± 1.0 °C, 7.52 ± 0.21, and 7.51 ± 0.63 mg/L during the acclimation, respectively. The photoperiod followed the natural light and dark cycle. The handling of crayfish in this study followed the ethical regulations of the Institutional Animal Care and Use Committee of the Institute of Hydrobiology, Chinese Academy of Sciences (Approval number: Keshuizhuan 08529).
2.2. Oxygen Consumption Rate and Asphyxiation Point Experiment
The oxygen consumption rate and asphyxiation point of juvenile P. clarkii and subadult P. clarkii (juvenile: 1.17 ± 0.03 g, subadult: 11.68 ± 0.11 g) were measured at five different temperatures (20, 22, 24, 26, and 28 °C) before the hypoxia stress test. For the determination of the oxygen consumption rate, single juvenile crayfish or subadult crayfish were placed into a glass tank of 22 L water (35 cm × 23 cm × 30 cm) sealed with 2 mm liquid paraffin wax, with 30 replicates per temperature level. A dissolved oxygen tester (YSI Pro ODO) was used to determine the variation in dissolved oxygen in water within 24 h. The oxygen consumption rates of juvenile and subadult crayfish were calculated [23,24]. The same method was used for the determination of the asphyxiation point. The dissolved oxygen in the water was determined when the juvenile crayfish and subadult crayfish were in asphyxiation coma (the crayfish lay on its side and did not respond to touching its feet with a glass rod), and the dissolved oxygen in the water was the asphyxiation point of P. clarkii at this temperature [23].
2.3. Hypoxia Stress Experiment
Three hypoxia stress groups (1, 2, and 3 mg/L) and a normal dissolved oxygen control group (7.5 mg/L) were set for juvenile P. clarkii and subadult crayfish, respectively. There were 3 replicas for each treatment group. The whole experiment was carried out in 24 glass tanks (35 cm × 23 cm × 30 cm), and each tank contained 12 crayfish. Before starting the experiment, healthy crayfish (juvenile crayfish: 1.17 ± 0.03 g, subadult crayfish: 11.68 ± 0.11 g) were selected and placed into tanks for 24 h of acclimation. During the acclimation period, the water temperature was 24.3 °C, pH 7.52 ± 0.21, photoperiod L:D = 12 h:12 h. After acclimation, each experimental tank was sealed with 2 mm liquid paraffin. Considering that the content of dissolved oxygen in water would be reduced due to the consumption of experimental animals, nitrogen and oxygen control devices were placed in the tank during sealing to monitor the stability of dissolved oxygen in the water of each experimental group and to ensure that crayfish could not carry out climbing interference experiments [25]. The experiment was carried out for 24 h, and the dissolved oxygen was measured every two hours to adjust the nitrogen to oxygen filling ratio and to maintain the stability of the dissolved oxygen in the water. During the experiment, the actual dissolved oxygen of the three hypoxia stress groups and the control group were 1.05 ± 0.07 mg/L, 2.06 ± 0.10 mg/L, 3.03 ± 0.08 mg/L, and 7.52 ± 0.57 mg/L, respectively.
2.4. Sample Collection
Three crayfish were taken from each replicate tankage, for a total of nine shrimp per treatment group, at 3 h, 6 h, 12 h, and 24 h after the beginning of the hypoxia stress experiment, and the hepatopancreases were dissected and preserved in frozen storage tubes. All samples were frozen in liquid nitrogen and then stored at −80 °C to determine the activity of antioxidant enzymes and the expression levels of stress genes.
2.5. Enzyme Activity Assay
The hepatopancreas samples were homogenized on ice with 1:9 normal saline volume (sample weight/normal saline volume) and centrifuged at 2500 RPM for 10 min. Supernatant was used for enzyme activity determination. Enzyme-linked immunosorbent assay (ELISA) was used to determine the activities of superoxide dismutase (SOD), catalase (CAT), and lactate dehydrogenase (LDH) and the content of lactic acid (LD) [20,21]. The above operations were carried out in strict accordance with the instructions of the kit. The kit was purchased from Jiancheng Institute of Biological Engineering, Nanjing. The results were read using an enzyme label instrument (SpectraMax@iD3, Molecular Devices, San Jose, CA, USA).
2.6. RNA Isolation and Quantitative RT–PCR
We washed the liver and hepatopancreas samples twice with PBS (pH 7.4) and placed the samples in TriPure reagents (Roche, Indianapolis, IN, USA) to extract total RNA. Purity and quantity of isolated RNA were determined by NanoDrop ND-1000 (Thermo Fisher Scientific, Waltham, MA, USA). Random hexamer primers and Moroni mouse leukemia virus reverse transcriptase (Promega, Madison, WI, USA) were used for RNA reverse transcription with OD260/OD280 values between 1.8 and 2.1. Real-time PCR analysis of cDNA samples was conducted in a LightCycler 480 Real Time PCR System (Switzerland, Germany) using FastStart Universal SYBR Green Master Mix (Roche Diagnostics GmbH, Mannheim, Germany). 18S was used as a reference for internal reference genes, and the relative gene expression calculation method was the 2−ΔΔct method [26]. The primer pair sequence is shown in Table 1.

Table 1.
Reference sources for qPCR primers.
2.7. Data Analysis
The oxygen consumption rates, asphyxiation points, and expression levels of antioxidant enzymes, anaerobic metabolic enzymes, and antioxidant genes of P. clarkii were expressed as the mean ± standard error (SE). After testing for variance normality and homogeneity, a two-way ANOVA was used to test for differences in oxygen consumption rate and asphyxiation point between juvenile and subadult crayfish at five temperature levels. The activities of antioxidant enzymes (SOD and CAT), the contents of anaerobic metabolizing enzymes (LD and LDH), and the expression levels of the SOD mRNA, CAT mRNA, Hsp70 mRNA, and crustin 4 mRNA of the four different groups at four different sampling time points were tested for differences using two-way ANOVA. If the difference was significant, an LSD test was used to compare the difference between the two pairs. Differences were considered to be significant if p < 0.05. Statistical analysis was performed using SPSS 24.0 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Oxygen Consumption Rate, Asphyxiation Point, and Mortality
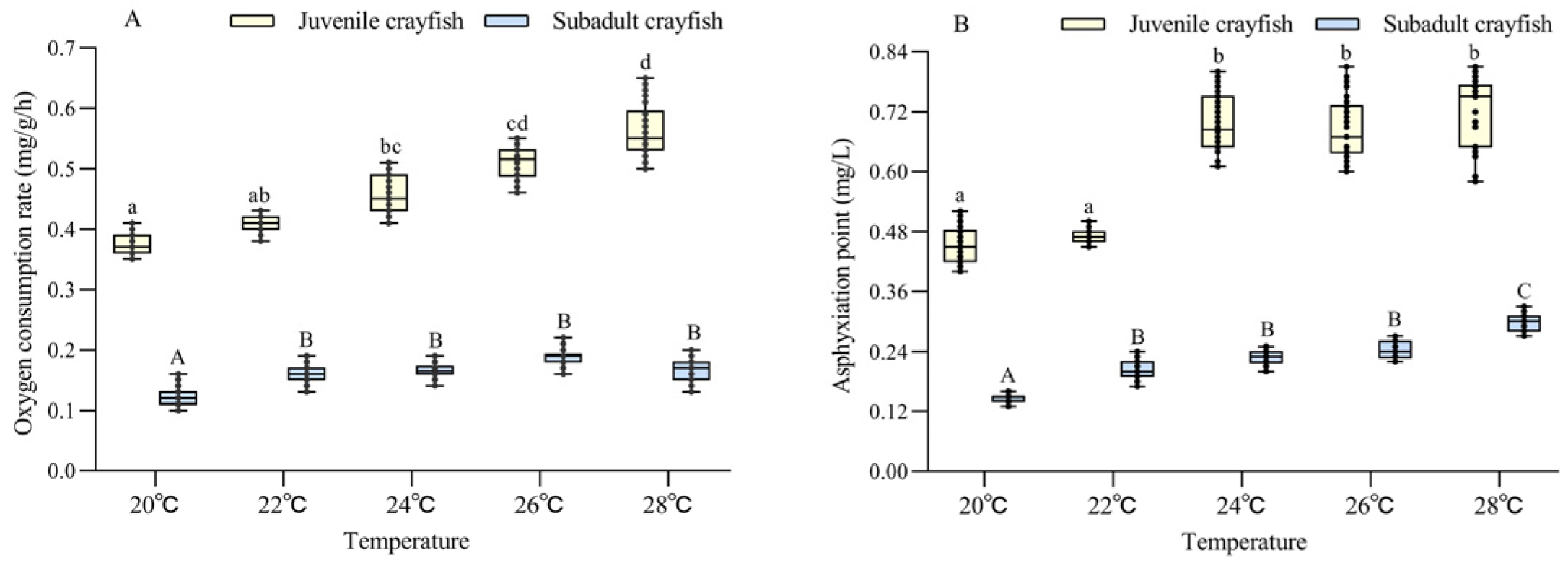
The oxygen consumption rate of P. clarkii was significantly affected by temperature and size, and there was a significant interaction between them (Table 2). The oxygen consumption rate of juvenile and subadult P. clarkii increased with increasing temperatures (20–28 °C). At the same temperature, the oxygen consumption rate of juvenile crayfish was 2.8–2.9 times higher than that of subadult crayfish (all p < 0.05) (Figure 1A).
Table 2.
Two-way ANOVA for oxygen consumption rate and asphyxiation point of juvenile and subadult Procambarus clarkii at five temperature (20, 22, 24, 26, 28 °C) levels.
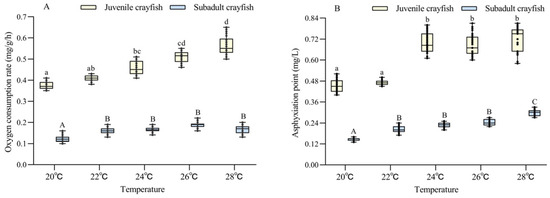
Figure 1.
Analysis of the difference in oxygen consumption rate (A) and asphyxiation point (B) between juvenile and subadult Procambarus clarkii at different temperature levels. There were 30 shrimp per temperature level, and each point showed the individual’s oxygen consumption rate and asphyxiation point. In the figure, there were no significant differences in oxygen consumption rate and asphyxiation point between the treatment groups with the same letters, but there were significant differences between the treatment groups with different letters (p < 0.05).
The asphyxiation point was significantly affected by temperature and size, but there was no significant interaction between them (Table 2). The asphyxiation points of juvenile and subadult P. clarkii also increased with increasing temperatures (20–28 °C). At the same temperature, the oxygen consumption rate of juvenile crayfish was 2.4–3.0 times higher than that of subadult crayfish (all p < 0.05) (Figure 1B).
During the 24 h hypoxia stress experiment, there was no death among the experimental crayfish.
3.2. Activities of Antioxidant Enzymes and Anaerobic Metabolic Enzymes in Juvenile P. clarkii
The SOD activity, LD content, and LDH activity of juvenile P. clarkii were significantly affected by dissolved oxygen and sampling time, and there was a significant interaction between them (Table 3). The CAT activity of juvenile P. clarkii was significantly affected by dissolved oxygen but not by sampling time, and there was a significant interaction between them (Table 3).
Table 3.
Two-way ANOVA for the SOD, CAT, LD, and LDH of juvenile Procambarus clarkii among four dissolved oxygen treatments (1, 2, 3, 7.5 mg/L) at four sampling times (3, 6, 12, 24 h).
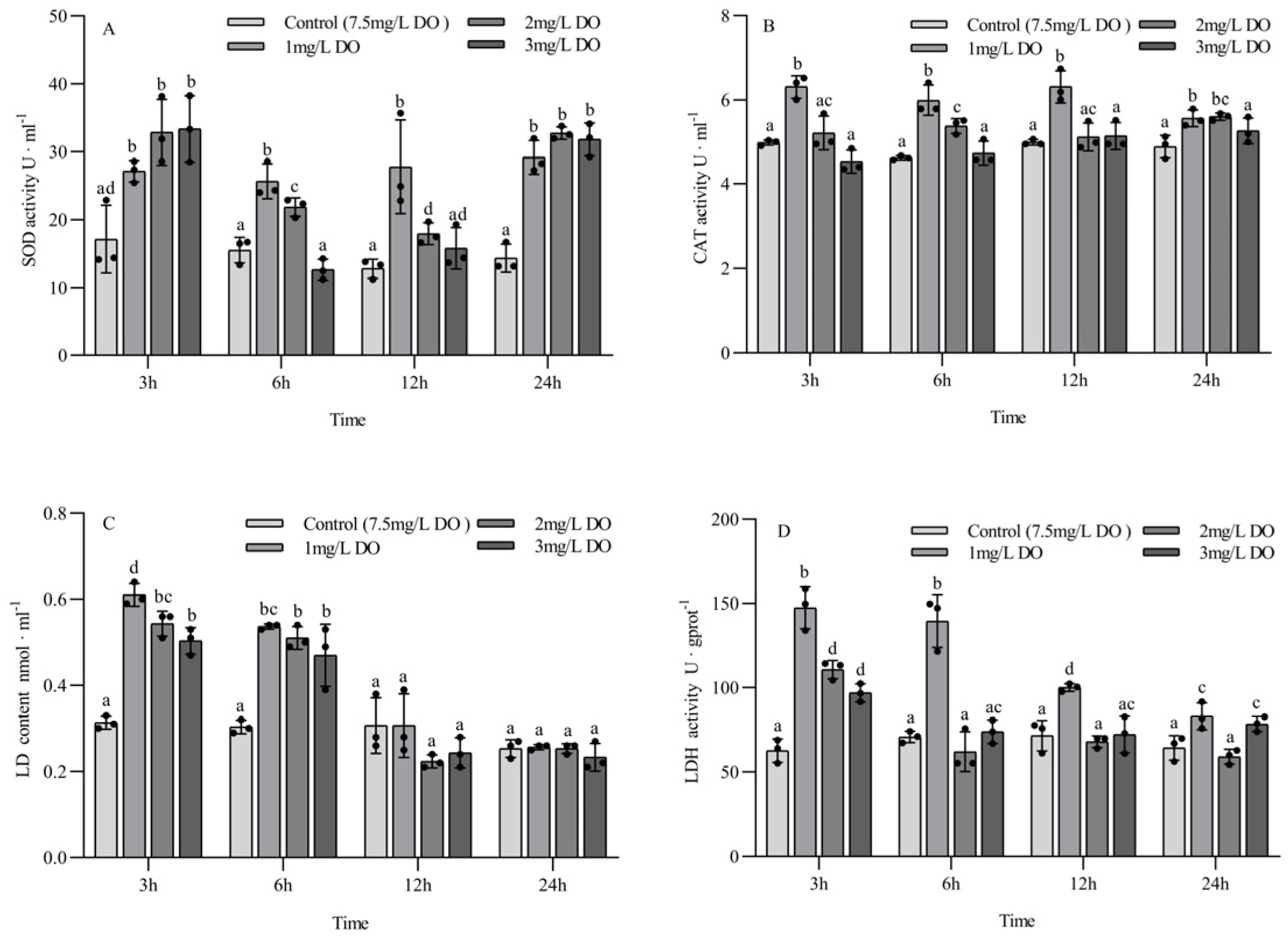
There were no significant differences in SOD activity among the three hypoxia groups at 3 h and 24 h (all p > 0.05), but all of them were significantly higher than those in the control group (all p < 0.05). The activity of SOD in the control group at 6 h and 12 h was significantly lower than that in the 1 mg/L and 2 mg/L groups (all p < 0.05), but there was no significant difference between the control group and the 3 mg/L group (all p > 0.05). Among the different sampling time points of the same group, there were significant differences in SOD activity in the control group and the 1 mg/L group at the four sampling time points (all p < 0.05). The SOD activity of the 2 mg/L and 3 mg/L groups at 3 h and 24 h was significantly higher than that at 6 h and 12 h (all p < 0.05), and the SOD activity of the 2 mg/L group at 6 h was significantly higher than that at 12 h (p < 0.05) (Figure 2A).
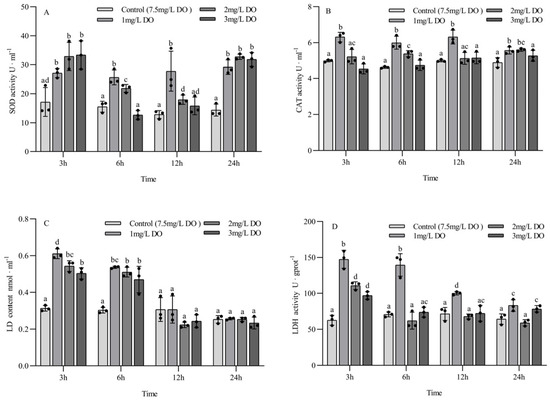
Figure 2.
Changes in antioxidant enzyme and respiratory enzyme activities of juvenile Procambarus clarkii under different dissolved oxygen levels and different sampling times. Superoxide dismutase (A), Catalase (B), Lactate content (C), Lactate dehydrogenase (D). There were three repeat groups for each dissolved oxygen level, replicate groups of 12 shrimp, with water temperatures of 24.0 °C, and each point showed the average enzyme activity of the three shrimp under the current conditions. In the figure, there were no significant differences in enzyme activity between the groups with the same letters, but there were significant differences between the groups with different letters (p < 0.05).
The activity of CAT in the 1 mg/L group was significantly higher than that in the 2 mg/L and 3 mg/L groups and the control group at 3 h, 6 h, and 12 h (p < 0.05), and the CAT activity in the 2 mg/L group was significantly higher than that in the control group at 6 h and 24 h (p < 0.05). There were no significant differences in CAT activity between the 3 mg/L group and the control group at the four sampling time points (all p > 0.05) (Figure 2B).
The LD content of the three hypoxia stress groups was significantly higher than that of the control group at 3 h and 6 h (p < 0.05), and the LD content of the 1 mg/L group was significantly higher than that of the 3 mg/L group at 3 h (p < 0.05), but there were no significant differences among the four groups at 12 h and 24 h (all p > 0.05). Among the different sampling time points of the same group, the LD content of the three hypoxia stress groups at 3 h and 6 h was significantly higher than that at 12 h and 24 h (all p < 0.05), while the LD content of the control group had no significant differences at the four sampling time points (all p > 0.05) (Figure 2C).
The content of LDH in the 1 mg/L group at 3 h, 6 h, and 12 h was significantly higher than that in the 2 mg/L and 3 mg/L groups and the control group (all p < 0.05). The LDH activity in the 2 mg/L and 3 mg/L groups was significantly higher than that in the control group at 3 h (all p < 0.05), but there was no significant difference at 6 h and 12 h. Among the different sampling time points of the same group, the LDH activity at 3 h and 6 h in the 1 mg/L group was significantly higher than that at 12 h and 24 h (p < 0.05), the LDH activity at 12 h was significantly higher than that at 24 h (p < 0.05), and the LDH activity in the 2 mg/L and 3 mg/L groups at 3 h was significantly higher than that at 6 h, 12 h, and 24 h (p < 0.05). There were no significant differences in LDH activity in the control group at the four sampling time points (all p > 0.05) (Figure 2D).
3.3. Activities of Antioxidant Enzymes and Anaerobic Metabolic Enzymes in Subadult P. clarkii
The activities of SOD, CAT, and LDH and the LD content of subadult P. clarkii were significantly affected by dissolved oxygen and sampling time, and there were significant differences (Table 4).
Table 4.
Two-way ANOVA for the SOD, CAT, LD, and LDH of subadult Procambarus clarkii among four dissolved oxygen treatments (1, 2, 3, 7.5 mg/L) at four sampling times (3, 6, 12, 24 h).
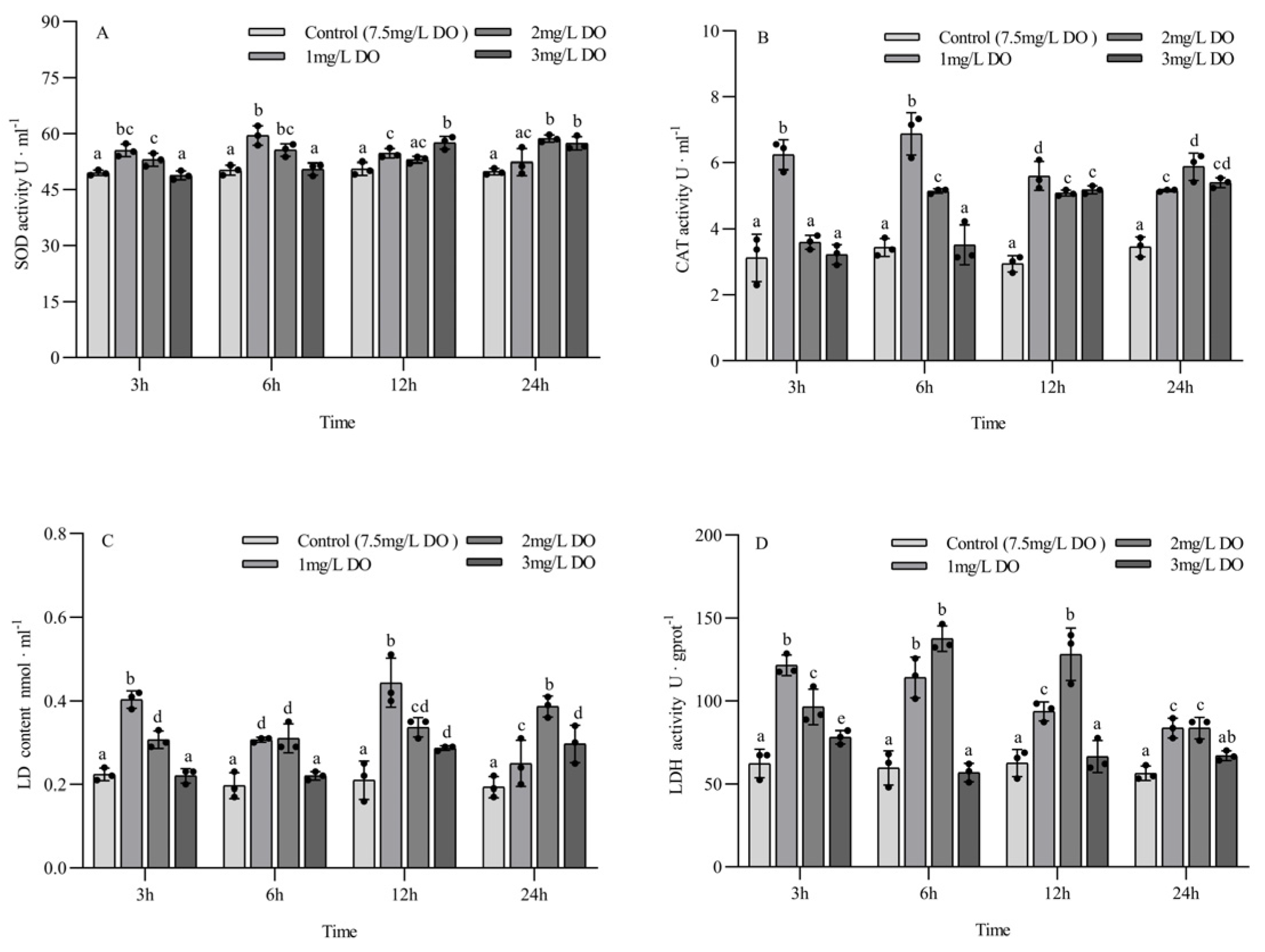
The SOD activity of the 1 mg/L and 2 mg/L groups at 3 h and 6 h was significantly higher than that of the 3 mg/L group and the control group (p < 0.05), but there was no significant difference between the latter two groups. The SOD activity of the 1 mg/L and 3 mg/L groups was significantly higher than that of the control group at 12 h, while the SOD activity of the 2 mg/L and 3 mg/L groups was significantly higher than that of the control group at 24 h. Among the different sampling time points of the same group, the SOD activity of the 1 mg/L group at 6 h was significantly higher than that at 3 h, 12 h, and 24 h (p < 0.05); the SOD activity of the 2 mg/L group at 24 h was significantly higher than that at 3 h and 12 h; and the SOD activity of the 3 mg/L group at 12 h and 24 h was significantly higher than that at 3 h and 6 h. There were no significant differences in SOD content in the control group at the four sampling time points (all p > 0.05) (Figure 3A).
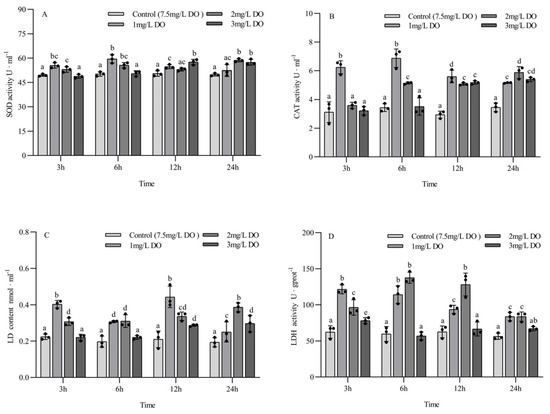
Figure 3.
Changes in antioxidant enzyme and respiratory enzyme activities of subadult Procambarus clarkii under different dissolved oxygen levels and different sampling times. Superoxide dismutase (A), Catalase (B), Lactate content (C), Lactate dehydrogenase (D). There were three repeat groups for each dissolved oxygen level, replicate groups of 12 shrimp, with water temperatures of 24.0 °C, and each point showed the average enzyme activity of the three shrimp under the current conditions. In the figure, there were no significant differences in enzyme activity between the groups with the same letters, but there were significant differences between the groups with different letters (p < 0.05).
The CAT activity of the 1 mg/L group at 3 h was significantly higher than that of the other three groups (p < 0.05), and there were no significant differences among the latter three groups. The CAT activity of the 1 mg/L and 2 mg/L groups at 6 h was significantly higher than that of the 3 mg/L group and control group (p < 0.05), while the CAT activity of the three hypoxia groups at 12 h and 24 h was significantly higher than that of the control group (all p < 0.05). Among the different sampling time points of the same group, the CAT activity of the 1 mg/L group at 3 h and 6 h was significantly higher than that at 12 h and 24 h (all p < 0.05), while the CAT activity of the 2 mg/L group at 24 h was significantly higher than that at 3 h, 6 h, and 12 h (all p < 0.05). The CAT activity of the 3 mg/L group increased with the increase in sampling time. There were no significant differences in CAT content in the control group at the four sampling time points (all p > 0.05) (Figure 3B).
The LD content of the 1 mg/L and 2 mg/L treatment groups was significantly higher than that of the control group at the four sampling time points (all p < 0.05). At 3 h and 6 h, the LD content of the 3 mg/L group was significantly lower than that of the 1 mg/L and 2 mg/L groups, but no significant difference was found between the 3 mg/L group and the control group. The LD content of the 3 mg/L group at 12 h and 24 h was significantly higher than that of the control group (p < 0.05). There were significant differences in LD content among the three hypoxia groups at different sampling time points in the same treatment group, and the LD content in the 2 mg/L and 3 mg/L groups increased with time. There were no significant differences in LD content in the control group at the four sampling time points (all p > 0.05) (Figure 3C).
The LDH activity of the 1 mg/L and 2 mg/L groups was significantly higher than that of the 3 mg/L group and control group at the four sampling time points (all p < 0.05). The LDH activity of the 3 mg/L group was significantly higher than that of control group at 3 h (p < 0.05), but there were no significant differences between the 3 mg/L group and control group at 6 h, 12 h, and 24 h. There were significant differences in LD content among the three hypoxia groups at different sampling time points in the same treatment group, and the LD content in the 1 mg/L group decreased with the time of sampling. There were no significant differences in LDH activity in the control group at the four sampling time points (all p > 0.05) (Figure 3D).
3.4. Antioxidant and Stress Gene Expression of Juvenile P. clarkii
The expression levels of SOD mRNA, Hsp70 mRNA, and crustin 4 mRNA in juvenile P. clarkii were significantly affected by dissolved oxygen and sampling time, and there was a significant interaction between them (Table 5). The CAT mRNA expression level of juvenile P. clarkii was significantly affected by dissolved oxygen but not by sampling time, and there was no significant interaction between them (Table 4).
Table 5.
Two-way ANOVA for the SOD mRNA, CAT mRNA, Hsp70 mRNA, and crustin 4 mRNA of juvenile Procambarus clarkii among four dissolved oxygen treatments (1, 2, 3, 7.5 mg/L) at four sampling times (3, 6, 12, 24 h).
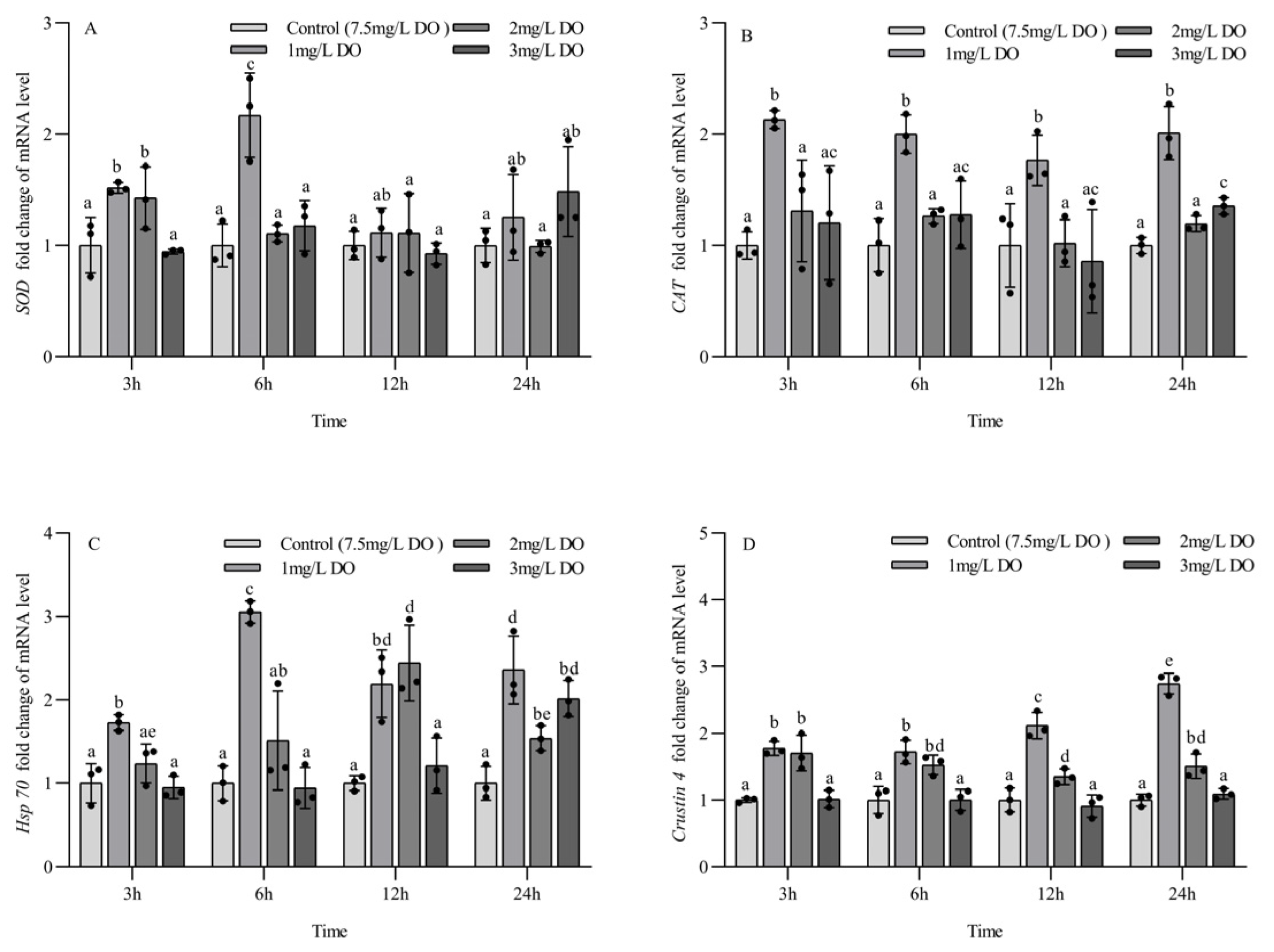
The SOD mRNA expression level of the 1 mg/L and 2 mg/L groups at 3 h was significantly higher than that of the 3 mg/L group and control group (p < 0.05), while the SOD mRNA expression level of the 1 mg/L group was significantly higher than that of the other three groups at 6 h (p < 0.05). There were no significant differences in the SOD mRNA expression level among the four groups at 12 h and 24 h. Among the different sampling time points of the same group, the SOD mRNA expression level at 6 h in the 1 mg/L group was significantly higher than that at 3 h, 12 h, and 24 h (p < 0.05), and the SOD mRNA expression level in the 2 mg/L group at 3 h was significantly higher than that at 6 h, 12 h, and 24 h (p < 0.05). There were no significant differences in the SOD mRNA expression level of the 3 mg/L group and the control group at the four sampling time points (all p > 0.05) (Figure 4A).
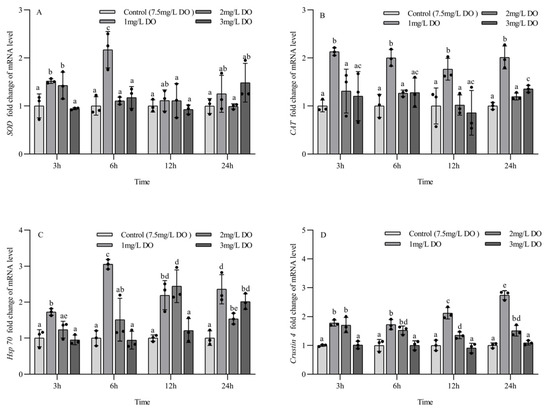
Figure 4.
Changes in antioxidant and stress genes in juvenile Procambarus clarkii under different dissolved oxygen levels and different sampling times. Superoxide dismutase mRNA (A), Catalase mRNA (B), Heat shock protein 70 mRNA (C), Crustin 4 mRNA (D). There were three repeat groups for each dissolved oxygen level, replicate groups of 12 shrimp, with water temperatures of 24.0 °C, and each point showed the average of the gene expression levels of the three shrimp under the current conditions. In the figure, there were no significant differences in gene expression between the treatment groups with the same letters, but there were significant differences between the treatment groups with different letters (p < 0.05).
At the four sampling time points, the CAT mRNA expression level of the 1 mg/L group was significantly higher than that of the other three groups (p < 0.05), and the 2 mg/L group had no significant differences from the control group. The CAT mRNA expression level of the 3 mg/L group was significantly higher than that of the control group only at 24 h (p < 0.05). There were no significant differences in the expression of CAT mRNA at the four sampling time points for all groups (all p > 0.05) (Figure 4B).
The expression of Hsp70 mRNA in the 1 mg/L group was significantly higher than that in the other three groups at 3 h and 6 h (p < 0.05), and the expression level of Hsp70 mRNA in the 1 mg/L and 2 mg/L groups was significantly higher than that in the 3 mg/L group and control group at 12 h (p < 0.05). At 24 h, the expression level of Hsp70 mRNA in the three hypoxia stress groups was significantly higher than that in the control group (all p < 0.05). There were significant differences in the expression of Hsp70 mRNA at the four sampling time points for the three hypoxia stress groups (all p < 0.05), but there were no significant differences in the expression of Hsp70 mRNA in the control group at the four sampling time points (all p > 0.05) (Figure 4C).
At the four sampling time points, the expression of crustin 4 mRNA in the 1 mg/L and 2 mg/L groups was significantly higher than that in the 3 mg/L group and control group (all p < 0.05), but there was no significant difference between the 3 mg/L group and control group (p < 0.05). There were significant differences in crustin 4 mRNA expression at different sampling time points for the 1 mg/L and 2 mg/L groups (all p < 0.05), but no significant differences were found for the 3 mg/L group and control group at the four sampling time points (all p > 0.05) (Figure 4D).
3.5. Antioxidant and Stress Gene Expression of Subadult P. clarkii
The mRNA expression levels of SOD, CAT, Hsp70, and crustin 4 in subadult P. clarkii were significantly affected by dissolved oxygen and sampling time. There was a significant interaction between them for SOD mRNA and CAT mRNA and no significant interaction between them for Hsp70 mRNA and crustin 4 mRNA (Table 6).
Table 6.
Two-way ANOVA for the SOD mRNA, CAT mRNA, Hsp70 mRNA, and crustin 4 mRNA of subadult Procambarus clarkii among four dissolved oxygen treatments (1, 2, 3, 7.5 mg/L) at four sampling times (3, 6, 12, 24 h).
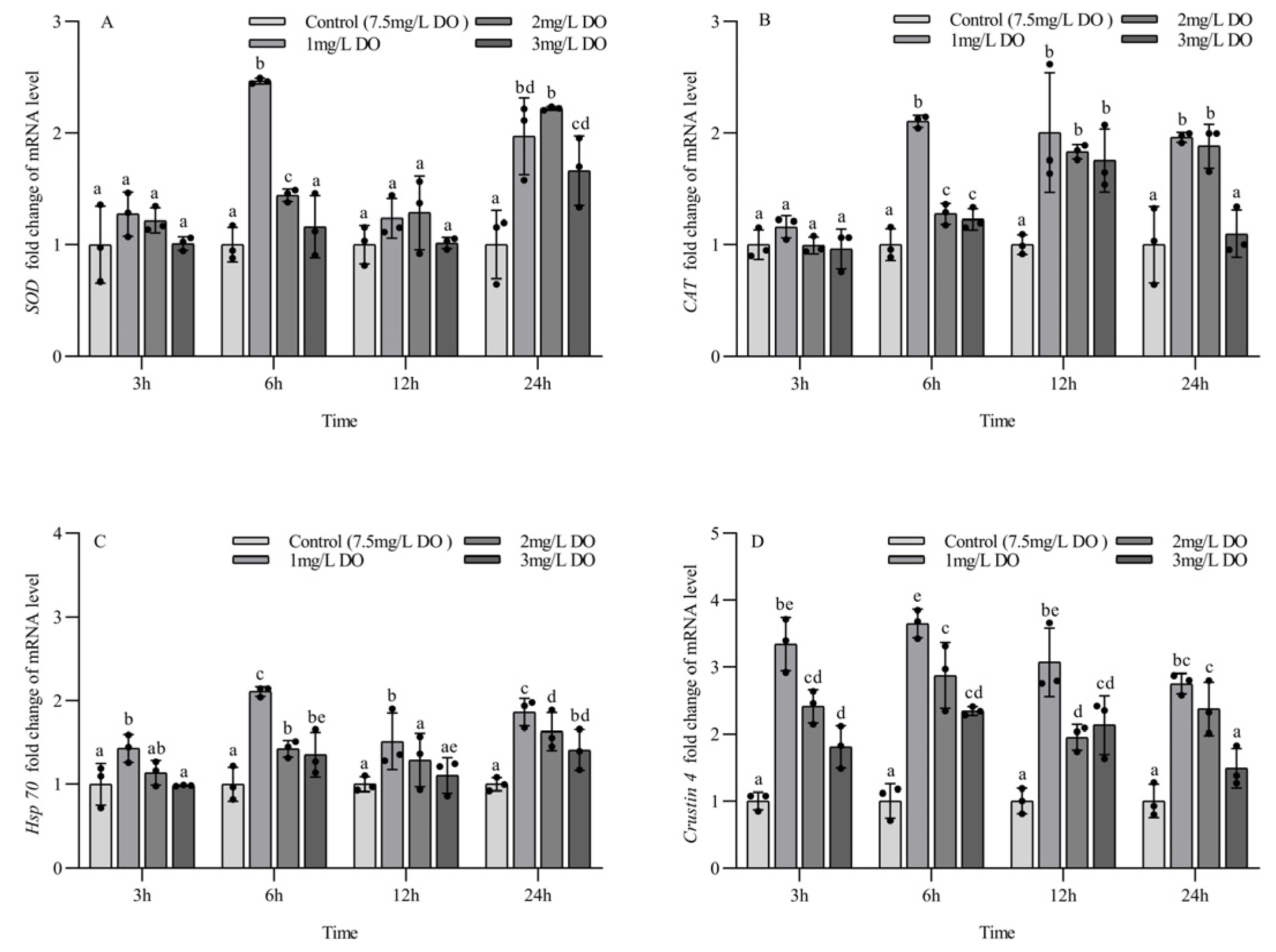
There were no significant differences in the expression of SOD mRNA for the four groups at 3 h and 12 h, but the expression of SOD mRNA in the 1 mg/L and 2 mg/L groups was significantly higher than that in the 3 mg/L group and control group at 6 h (p < 0.05). The SOD mRNA expression level in three hypoxia stress groups was significantly higher than that in control group at 24 h (all p < 0.05). There were significant differences in SOD mRNA expression levels at 6 and 24 h for the three hypoxia stress groups, respectively (all p < 0.05), but no significant differences were found in the control group at the four sampling time points (all p > 0.05) (Figure 5A).
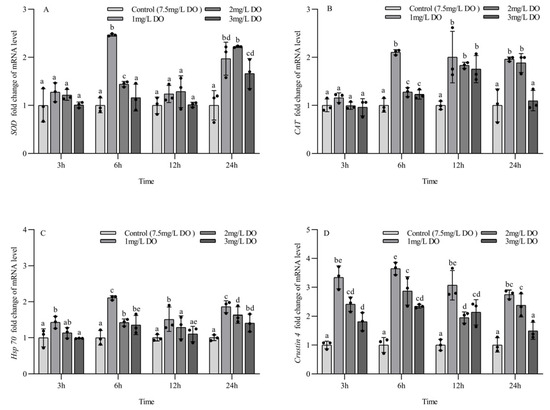
Figure 5.
Changes in antioxidant and stress genes in subadult Procambarus clarkii under different dissolved oxygen levels and different sampling times. Superoxide dismutase mRNA (A), Catalase mRNA (B), Heat shock protein 70 mRNA (C), Crustin 4 mRNA (D). There were three repeat groups for each dissolved oxygen level, replicate groups of 12 shrimp, with water temperatures of 24.0 °C, and each point showed the average of the gene expression levels of the three shrimp under the current conditions. In the figure, there were no significant differences in gene expression between the treatment groups with the same letters, but there were significant differences between the treatment groups with different letters (p < 0.05).
There were no significant differences in the expression of CAT mRNA among the four groups at 3 h, but the expression level of CAT mRNA in three hypoxia stress groups was significantly higher than that in the control group at 6 h and 12 h (all p < 0.05), and the expression level of CAT mRNA in the 1 mg/L and 2 mg/L treatment groups was significantly higher than that in the 3 mg/L group and control group at 24 h (p < 0.05). There were significant differences in CAT mRNA expression at 6 and 24 h for the three hypoxia stress groups, respectively (all p < 0.05). The CAT mRNA expression level in the 1 mg/L and 3 mg/L groups increased firstly and then decreased with the increase in sampling time, and the CAT mRNA expression level in the 2 mg/L group increased gradually with the increase in sampling time. There were no significant differences in CAT mRNA expression levels in the control group at the four sampling time points (all p > 0.05) (Figure 5B).
The expression level of Hsp70 mRNA in the 1 mg/L group was significantly higher than that in the other three groups at 3 h and 12 h (p < 0.05), and there were no significant differences among the latter three groups. The expression level of Hsp70 mRNA in three hypoxia stress groups was significantly higher than that in the control group at 6 h and 24 h (all p < 0.05), and the expression level of Hsp70 mRNA in the 1 mg/L group was significantly higher than that in the 2 mg/L and 3 mg/L groups (p < 0.05). There were significant differences in the expression of Hsp70 mRNA at the four sampling time points for the three hypoxia stress groups, respectively (all p < 0.05), but no significant differences were found in the control group at the four sampling time points (all p > 0.05) (Figure 5C).
At the four sampling time points, the expression level of crustin 4 mRNA in three hypoxia stress groups was significantly higher than that in the control group (all p < 0.05), and the expression level of crustin 4 mRNA in the 1 mg/L group was significantly higher than that in the 3 mg/L group (all p < 0.05). There were significant differences in crustin 4 mRNA expression at the four sampling time points for the three hypoxia stress groups, respectively (all p < 0.05), but no significant differences were found in the control group at the four sampling time points (all p > 0.05) (Figure 5D).
4. Discussion
4.1. Effect of Temperature on the Oxygen Consumption Rate and Asphyxiation Point of P. clarkii
Temperature is the most important environmental factor affecting the oxygen consumption rate and asphyxiation point of shrimps and crabs. In the range of 20 °C to 28 °C, the oxygen consumption rate and asphyxiation point of juvenile and subadult P. clarkii increased with increasing temperature in the study; the results are consistent with those of L. vannamei and Eriocheir sinensis (E. sinensis) [22,23]. This is because in an appropriate temperature range, the enzyme activity in the tissues of aquatic animals will be enhanced with increasing temperatures, thus accelerating the rate of metabolism and increasing the oxygen consumption and asphyxiation point [27,28]. In addition, individual size will also affect the oxygen consumption level of aquatic animals, and usually, the oxygen consumption rate will decrease with the increasing size of aquatic animals [29]. The results in this experiment are consistent with the above conclusions, which indicated that subadult crayfish had stronger hypoxia tolerance than juvenile crayfish. Under the same conditions, compared with existing large-scale cultured crustaceans, the oxygen consumption rate and asphyxiation point of P. clarkii were lower than those of E. sinensis [23,30], Penaeus japonucus [31], and L. vannamei [24]. This shows that P. clarkii has a strong ability to adapt to hypoxia.
4.2. Effect of Hypoxia on the Activity of Antioxidant and Anaerobic Metabolic Enzymes in P. clarkii
Hypoxia stress can cause an oxidative stress response in P. clarkii, disturb the dynamic balance of reactive oxygen species (ROS) in the body, and significantly increase the ROS content in the hepatopancreas, while uncleared ROS will accelerate the formation of intracellular lipid peroxides and cause oxidative damage [32,33]. SOD and CAT are important components of intracellular antioxidant enzymes that scavenge oxygen free radicals. SOD and CAT can convert superoxide free radicals into oxygen molecules and water that are nontoxic to organisms, thus maintaining the normal physiological activities of cells and bodies [34]. In this study, the activities of SOD and CAT in the hypoxia stress groups were higher than those in the control group for both juvenile crayfish and subadult crayfish, indicating that hypoxia stress could induce an increase in SOD and CAT activities in the hepatopancreas of both groups to address the oxidative damage caused by ROS. The results are consistent with the results of L. vannamei [20]. Under the same conditions, the oxygen consumption rate of juvenile shrimps was significantly higher than that of subadult shrimps, resulting in significantly increased SOD and CAT enzyme activities in the hepatopancreas of juvenile shrimps and advanced oxidative stress reactions [20,29].
Previous studies have shown that when P. clarkii is exposed to hypoxia, the pathway of energy metabolism changes from aerobic respiration to anaerobic respiration to maintain physiological balance [35,36]. The main end product of anaerobic respiration in crustaceans is lactic acid (LD), which can enhance the binding ability of hemocyanin and oxygen molecules in hypoxia [37,38,39], while LDH is one of the important enzymes in anaerobic glycolysis and gluconeogenesis, which can catalyze the redox reaction between LD and pyruvate [40]. In this study, the LD content and LDH activity in the hepatopancreas of juvenile crayfish showed significant fluctuations in a short period of time, while the fluctuations in LD content and LDH activity in the hepatopancreas of subadult crayfish were weaker than those of juvenile crayfish. The reason for this difference is that under the same conditions, the brain, hepatopancreas, and other organs and tissues that maintain life activities in juvenile shrimp account for a larger proportion of body mass [41]. These organs consume more energy for metabolic activities, increase anaerobic respiration, and result in higher LD content and LDH activity in the hepatopancreas of juvenile shrimp compared to subadult shrimp.
Under hypoxia stress, juvenile crayfish will rapidly increase the anaerobic respiration rate in a short time (6 h) to generate energy and maintain body balance, which will lead to a rapid increase in LD content and a significant increase in LDH activity, while excessive LDH activity will cause liver damage, and the higher the activity of LDH, the greater the degree of hepatopancreas injury [39]. When the activity of LDH is too high, the degree of liver injury increases. To protect the liver from injury, the body inhibits anaerobic respiration, thus reducing the LD content and LDH activity to protect the body. In summary, juvenile crayfish are more susceptible to hypoxic conditions than subadult crayfish at the same temperature, and anaerobic respiration causes more serious damage to juvenile crayfish.
4.3. Effect of Hypoxia on the Expression of Antioxidant and Stress Genes in P. clarkii
When the metabolic balance of ROS in aquatic animals is disrupted, oxidative stress reactions are induced. Excessive accumulation of ROS leads to damage to cell proteins and lipid membranes, while SOD and CAT are the first line of defense against oxidative stress in biological organisms [42]. Compared with the control group, the expression of SOD mRNA and CAT mRNA in the hypoxia stress groups was significantly increased. The expression of SOD mRNA and CAT mRNA in the 1 mg/L group was higher than that in the 2 mg/L and 3 mg/L groups at 3 h and 6 h, and the expression levels of SOD mRNA and CAT mRNA in juvenile crayfish in the 1 mg/L group were higher than those in subadult crayfish. The results showed that the stronger the degree of hypoxia stress, the higher the expression levels of SOD mRNA and CAT mRNA of P. clarkii in a short time (6 h). Under the same hypoxia stress, the oxidative stress response of juvenile crayfish was stronger than that of subadult crayfish.
The expression level of Hsp70 mRNA can reflect the effects of environmental stress on the body. Hsp70 mRNA can play an important biological function under stress conditions and is an activator of innate immunity, playing a key role in various stress responses and alleviating stress-induced injury [43,44,45]. In this study, the Hsp70 mRNA expression levels of juvenile and subadult crayfish in the hypoxia stress groups were significantly increased compared to those in the control group. The Hsp70 mRNA expression levels in juvenile and subadult crayfish in the 1 mg/L group were higher than those in the 2 mg/L and 3 mg/L groups, and the Hsp70 mRNA expression level of juvenile crayfish was higher than that of subadult crayfish under the same hypoxia stress. The results showed that the expression level of Hsp70 mRNA in juvenile and subadult crayfish was positively correlated with the degree of hypoxia stress, with the latter showing lower stress levels under the same hypoxia stress conditions.
Crustin 4 is an important immune effector molecule that is an important component of pathogen resistance and plays a crucial role in crayfish resistance to bacterial infection [46,47]. In this study, the expression level of crustin 4 mRNA in the 1 mg/L group was higher than that in the 2 mg/L and 3 mg/L groups, and the crustin 4 mRNA expression level of subadult crayfish was generally higher than that of juvenile crayfish, indicating that the stronger the degree of hypoxia stress level, the higher the expression level of crustin 4 mRNA. In the same hypoxic environment, juvenile crayfish have weaker resistance to bacterial infection than subadult crayfish and are more susceptible to disease.
For a short period of time (24 h), whether juvenile crayfish or subadult crayfish are exposed to a low dissolved oxygen environment, P. clarkii will always be in a state of stress, thus inducing the organism to increase the activities of SOD and CAT antioxidant enzymes and gene expression levels and improve the immune capacity of the organism to combat the oxidative damage caused by stress and infection by pathogenic bacteria. However, the enhancement of antioxidant capacity and immune capacity leads to an increase in energy consumption. When the energy generated by aerobic and anaerobic respiration is not sufficient to maintain a high level of immunity, the probability of pathogens from the environment invading the organism increases, resulting in an increase in the incidence or probability of death in crayfish [47]. In addition, when the organism is in a hypoxic environment for a long time, the accumulated oxidation products in the body beyond the tolerance range of the organism will lead to damage to the immune and antioxidant system and eventually cause disease and economic losses [32].
5. Conclusions
Red swamp crayfish P. clarkii had a low oxygen consumption rate and asphyxiation point and no death in the 24 h 1 mg/L hypoxia stress experiment, which indicated that P. clarkii had strong hypoxia adaptability, and the hypoxia adaptability of subadult crayfish was stronger than that of larvae. Hypoxia stress enhanced the antioxidant responses of crayfish and induced the upregulation of antioxidant genes to adapt to hypoxia. Compared to subadult crayfish, the antioxidant system of juvenile crayfish is more sensitive to hypoxic environments and is more likely to produce an oxidative stress response, resulting in increased probability of pathogen infection. The present results are from a 24 h acute hypoxia stress experiment; however, further studies on the effects of intermittent hypoxia stress on the survival, growth, and physiological function of P. clarkii on a longer time scale are needed in the future.
Author Contributions
Conceptualization, Q.Z. and W.L.; methodology, Q.Z. and W.L.; software, Q.Z.; validation, Q.Z., W.L. and L.Q.; formal analysis, Q.Z., M.L., C.G., G.F. and W.L.; resources, T.Z.; data curation, Q.Z. and W.L.; writing—original draft preparation, Q.Z., W.L., J.L., T.Z. and G.F.; writing—review and editing, W.L., T.Z., J.L. and C.G.; visualization, Q.Z.; supervision, W.L., T.Z. and J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Key Research and Development Program of Hubei Province (No. 2021BBA230), the National Key Research and Development Program of China (Grant No. 2023YFD2401802), the Youth Innovation Promotion Association CAS (2019331), the Technical Innovation Project of the Science and Technology Department of Hubei Province (2018ABA102), and the Innovative and Entrepreneurial Talents Project of Jiangxi Province (jxsq2020102109).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Animal Care and Use Committee of the Institute of Hydrobiology, Chinese Academy of Sciences (Approval number: Keshuizhuan 08529).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shu, X.Y. Problems and industrial development of Procambarus clarkii. China Fish. 2010, 08, 22–25. (In Chinese) [Google Scholar] [CrossRef]
- Guo, L.; Lin, W.; Cao, C.; Li, C. Integrated rice-crayfish farming system does not mitigate the global warming potential during rice season. Sci. Total Environ. 2023, 867, 161520. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Li, X.; Onchari, M.M.; Li, W.; Bu, Y.; Lek, S.; Jin, S. High feeding level alters physiological status but does not improve feed conversion efficiency and growth performance of juvenile red swamp crayfish Procambarus clarkii (Girard, 1852). Aquaculture 2021, 537, 736507. [Google Scholar] [CrossRef]
- Yu, X.J.; Hao, X.J.; Dang, Z.Q.; Yang, L.K.; Zhang, Y.H.; Cai, J.; Zhang, Y. China crayfish industry development report. China Fish. 2022, 6, 47–54. (In Chinese) [Google Scholar] [CrossRef]
- Liu, D.L.; Huang, J.H.; Yang, L.S.; Tan, L.Q. Effects of multiple environmental factors on molting death of Procambarus clarkii and countermeasures. Nanfang Shuichan Kexue 2020, 2, 29–35. (In Chinese) [Google Scholar] [CrossRef]
- Shu, B.; Li, J.T.; Zhang, X.X.; Xian, J.A.; Zhou, X.Y.; Wang, D.M. A Review: Effects of Environmental Factors on Immune Indices of Red Swamp Crayfish Procambarus clarkii. Shuichanxue Zazhi 2020, 33, 75. (In Chinese) [Google Scholar] [CrossRef]
- Estrada-Cárdenas, P.; Cruz-Moreno, D.G.; González-Ruiz, R.; Peregrino-Uriarte, A.B.; Leyva-Carrillo, L.; Camacho-Jiménez, L.; Yepiz-Plascencia, G. Combined hypoxia and high temperature affect differentially the response of antioxidant enzymes, glutathione and hydrogen peroxide in the white shrimp Litopenaeus vannamei. Comp. Biochem. Phys. A 2021, 254, 110909. [Google Scholar] [CrossRef] [PubMed]
- Hadley, K.R.; Paterson, A.M.; Reid, R.A.; Rusak, J.A.; Somers, K.M.; Ingram, R.; Smol, J.P. Altered pH and reduced calcium levels drive near extirpation of native crayfish, Cambarus bartonii, in Algonquin Park, Ontario, Canada. Freshw. Sci. 2015, 34, 918–932. [Google Scholar] [CrossRef]
- Jiang, L.X.; Pan, L.Q.; Xiao, G.Q. Effects of ammonia-N on immune parameters of white shrimp Litopenaeus vannamei. Zhongguo Shuichan Kexue 2004, 11, 537–541. (In Chinese) [Google Scholar] [CrossRef]
- Wang, P.; Liu, H.; Zhao, S.; Yu, S.; Xie, S.; Hua, S.; Gao, H. Hypoxia stress affects the physiological responses, apoptosis and innate immunity of Kuruma shrimp, Marsupenaeus japonicus. Fish. Shellfish. Immunol. 2022, 122, 206–214. [Google Scholar] [CrossRef]
- Liu, Q.J. Effects of Straw Return and Food Feeding on Water Quality in Rice Crawfish System. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2017. [Google Scholar]
- France, R.L. Comparative tolerance to low pH of three life stages of the crayfish Orconectes virilis. Can. J. Zool. 1984, 62, 2360–2363. [Google Scholar] [CrossRef]
- Zhong, J.W.; Zhu, Y.A.; Fu, P.S.; An, L.; Lv, B.Y. Toxicity of nitrite nitrogen on Procambarus clarkii and effects of nitrite nitrogen on immune-related factor. J. Yangtze Univ. (Mat. Sci. Ed.) 2014, 11, 36–40. (In Chinese) [Google Scholar] [CrossRef]
- Camacho-Jiménez, L.; Peregrino-Uriarte, A.B.; Martínez-Quintana, J.A.; Yepiz-Plascencia, G. The glyceraldehyde-3-phosphate dehydrogenase of the shrimp Litopenaeus vannamei: Molecular cloning, characterization and expression during hypoxia. Mar. Environ. Res. 2018, 138, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Zou, E.M.; Du, N.S.; Lai, W. The effects of mass temperature and thermal acclimation on the respiration rate of the Chinese freshwater crab Eriocheir sinensis (Crustacea: Decapod). Zool. Res. 1995, 16, 49–58. [Google Scholar] [CrossRef]
- Giordano, F.J. Oxygen, oxidative stress, hypoxia, and heart failure. J. Clin. Investig. 2005, 115, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.; Krishnan, S.; Anand, D.; Kokkattunivarthil Uthaman, S.; Otta, S.K.; Karunasagar, I.; Kooloth Valappil, R. Immune responses and immune protection in crustaceans with special reference to shrimp. Rev. Aquacult. 2021, 13, 431–459. [Google Scholar] [CrossRef]
- Norouzitallab, P.; Baruah, K.; Vanrompay, D.; Bossier, P. Teaching shrimps self-defense to fight infections. Trends Biotechnol. 2019, 37, 16–19. [Google Scholar] [CrossRef]
- Wilhelm Filho, D.; Torres, M.A.; Zaniboni-Filho, E.; Pedrosa, R.C. Effect of different oxygen tensions on weight gain, feed conversion, and antioxidant status in piapara, Leporinus elongatus (Valenciennes, 1847). Aquaculture 2005, 244, 349–357. [Google Scholar] [CrossRef]
- Han, S.Y.; Wang, M.Q.; Liu, M.; Wang, B.J.; Jiang, K.Y.; Wang, L. Comparative sensitivity of the hepatopancreas and midgut in the white shrimp Litopenaeus vannamei to oxidative stress under cyclic serious/medium hypoxia. Aquaculture 2018, 490, 44–52. [Google Scholar] [CrossRef]
- Sun, S.; Yang, M.; Fu, H.; Ge, X.; Zou, J. Altered intestinal microbiota induced by chronic hypoxia drives the effects on lipid metabolism and the immune response of oriental river prawn Macrobrachium nipponense. Aquaculture 2020, 526, 735431. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, X.; Li, J.; Huang, G. The effects of body weight, temperature, salinity, pH, light intensity and feeding condition on lethal DO levels of whiteleg shrimp, Litopenaeus vannamei (Boone, 1931). Aquaculture 2006, 256, 579–587. [Google Scholar] [CrossRef]
- Wen, X.B.; Chen, L.Q.; Ai, C.X.; Jiang, H.B. Standard Metabolism of the juvenile Crab Eriocheir sicensis. Zool. Res. 2001, 22, 425–428. (In Chinese) [Google Scholar] [CrossRef]
- Chen, Q.; Chen, X.H.; Luo, Y.J.; Huang, J.; Li, G.F.; Ning, L.K. Preliminary determination of oxygen consumption rate and asphyxia point of Litopenaeus vannamei. J. Hydrol. 2001, 21, 14–15. (In Chinese) [Google Scholar] [CrossRef]
- Nelson, S.G.; Armstrong, D.A.; Knight, A.W.; Li, H.W. The effects of temperature and salinity on the metabolic rate of juvenile Macrobrachium rosenbergii (Crustacea: Palaemonidae). Comp. Biochem. Phys. A 1977, 56, 533–537. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Liu, D.; Ru, S. Cu accumulation, detoxification and tolerance in the red swamp crayfish Procambarus clarkii. Ecotoxicol. Environ. Saf. 2019, 175, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Hammen, C.S. Aminotransferase activities and amino acid excretion of bivalve mollusks and brachiopods. Comp. Biochem. Phys. B 1968, 26, 697–705. [Google Scholar] [CrossRef]
- Beaufort, N.C. Biology of the blue crab, Callinectes sapidus rathbun, In the St. Johns River, Florida. Fish. B-Noaa. 1968, 67, 17–33. [Google Scholar]
- Christensen, E.A.; Svendsen, M.B.; Steffensen, J.F. The combined effect of body size and temperature on oxygen consumption rates and the size-dependency of preferred temperature in European perch Perca fluviatilis. J. Fish. Biol. 2020, 97, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Feng, W.R.; Zhou, L.H.; Ding, X.F.; Liang, M.; Tang, Y.K. Effects of temperature and pH on the oxygen consumption rate, ammonia discharge rate and asphyxiation point of juvenile Eriocheir sinensis. J. South. Agric. 2022, 05, 1415–1424. (In Chinese) [Google Scholar] [CrossRef]
- Dai, Q.N.; Wang, J.; Li, Q. On the oxygen consumption of Penaeus japouicus. Jimei Daxue Xuebao Ziran Kexueban 1994, 16, 25–29. (In Chinese) [Google Scholar] [CrossRef]
- Franco, R.; Sánchez-Olea, R.; Reyes-Reyes, E.M.; Panayiotidis, M.I. Environmental toxicity, oxidative stress and apoptosis: Menage a trois. Mutat.Res. Genet. Toxicol. Environ. Mutagen. 2009, 674, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, Y.M.; Xu, W.B.; Chen, D.Y.; Li, B.W.; Cheng, Y.X.; Shu, M.A. The effects of salinities stress on histopathological changes, serum biochemical index, non-specific immune and transcriptome analysis in red swamp crayfish Procambarus clarkii. Sci. Total Environ. 2022, 840, 156502. [Google Scholar] [CrossRef]
- Han, M.; Gao, T.; Liu, G.; Zhu, C.; Zhang, T.; Sun, M.; Jiang, Q. The effect of a polystyrene nanoplastic on the intestinal microbes and oxidative stress defense of the freshwater crayfish, Procambarus clarkii. Sci. Total Environ. 2022, 833, 155722. [Google Scholar] [CrossRef]
- Reiber, C.L.; McMahon, B.R. The effects of progressive hypoxia on the crustacean cardiovascular system: A comparison of the freshwater crayfish, (Procambarus clarkii), and the lobster (Homarus americanus). J. Comp. Physiol. B 1998, 168, 168–176. [Google Scholar] [CrossRef]
- Fujimori, T.; Abe, H. Physiological roles of free D-and L-alanine in the crayfish Procambarus clarkii with special reference to osmotic and anoxic stress responses. Comp. Biochem. Phys. A 2002, 131, 893–900. [Google Scholar] [CrossRef]
- da Silva-Castiglioni, D.; Oliveira, G.T.; Buckup, L. Metabolic responses of Parastacus defossus and Parastacus brasiliensis (Crustacea, Decapoda, Parastacidae) to hypoxia. Comp. Biochem. Phys. A 2010, 156, 436–444. [Google Scholar] [CrossRef] [PubMed]
- McMahon, B.R. Respiratory and circulatory compensation to hypoxia in crustaceans. Respir. Physiol. 2001, 128, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Truchot, J.P. Lactate increases the oxygen affinity of crab hemocyanin. J. Exp. Zool. 1980, 214, 205–208. [Google Scholar] [CrossRef]
- Qiu, R.; Cheng, Y.; Huang, X.; Wu, X.; Yang, X.; Tong, R. Effect of hypoxia on immunological, physiological response, and hepatopancreatic metabolism of juvenile Chinese mitten crab Eriocheir sinensis. Aquac. Int. 2011, 19, 283–299. [Google Scholar] [CrossRef]
- Qiu, D.G.; Huang, D.C.; Zheng, L.Y.; Wu, S.Q.; Liu, Y.H.; Qiu, F.Y.; Zheng, J.P.; Chen, X.M. Effects of temperature on respiration and excretion by different sizes of hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceo latus ♂). Zhongguo Shuichan kexue 2017, 24, 295–305. (In Chinese) [Google Scholar] [CrossRef]
- Eyckmans, M.; Celis, N.; Horemans, N.; Blust, R.; De Boeck, G. Exposure to waterborne copper reveals differences in oxidative stress response in three freshwater fish species. Aquat. Toxicol. 2011, 103, 112–120. [Google Scholar] [CrossRef]
- Benjamin, I.J. 20 Expression and function of stress proteins in the ischemic heart. CSH Monogr. 1994, 26, 533–552. [Google Scholar]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Basu, S.; Binder, R.J.; Suto, R.; Anderson, K.M.; Srivastava, P.K. Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-κB pathway. Int. Immunol. 2000, 12, 1539–1546. [Google Scholar] [CrossRef]
- Zhu, Y.T. The Anti-Microbial Functions of Immune Effectors in the Hemocytes of Chinese Mitten Crab, Eriocheir sinensis. Ph.D. Thesis, East China Normal University, Shanghai, China, 2016. [Google Scholar]
- Du, Z.Q.; Li, B.; Shen, X.L.; Wang, K.; Du, J.; Yu, X.D.; Yuan, J.J. A new antimicrobial peptide isoform, Pc-crustin 4 involved in antibacterial innate immune response in fresh water crayfish, Procambarus clarkii. Fish. Shellfish. Immunol. 2019, 94, 861–870. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).