
Shifts in the Distribution Range and Niche Dynamics of the Globally Threatened Western Tragopan (Tragopan melanocephalus) Due to Climate Change and Human Population Pressure

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
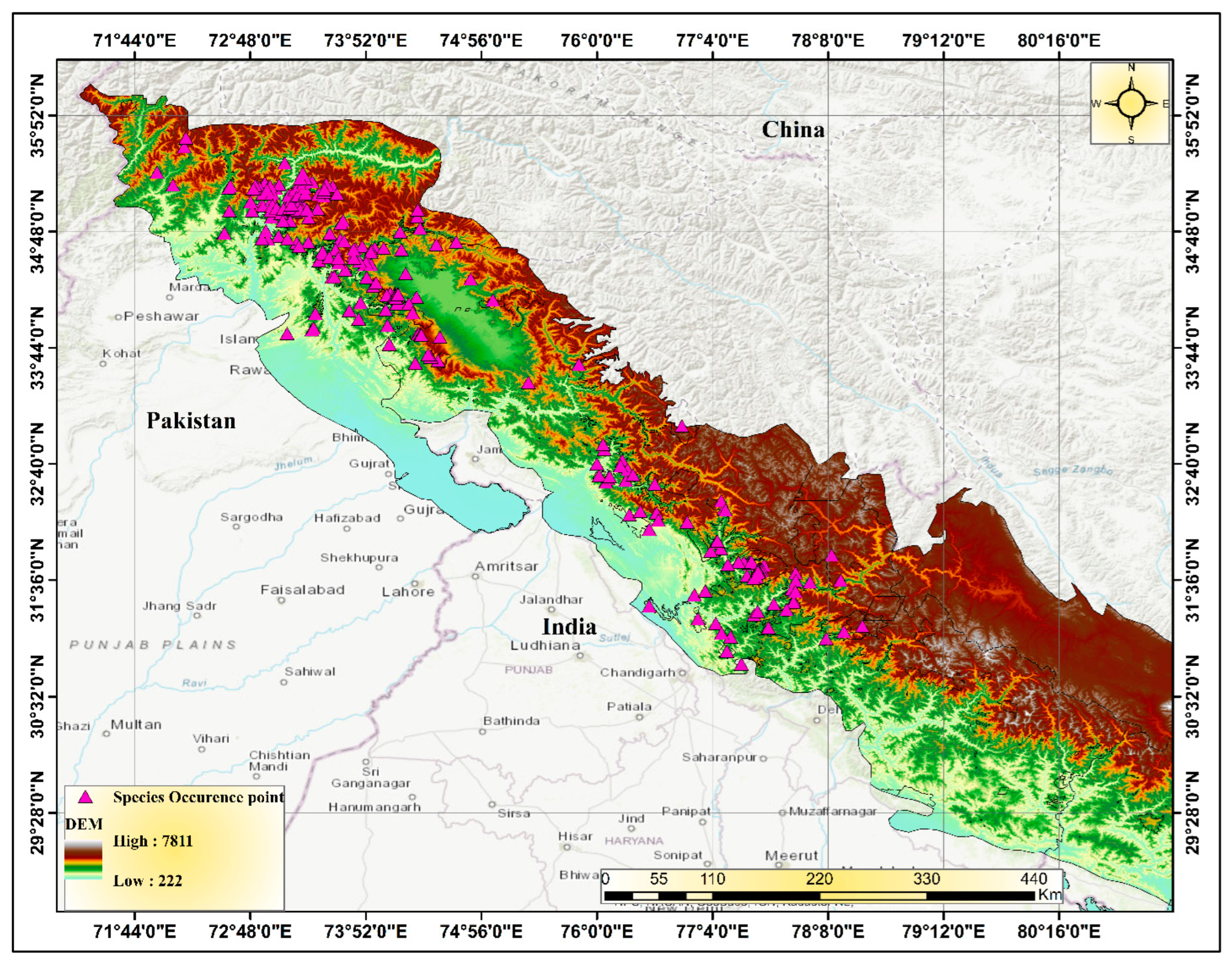
2.1. Study Area
2.2. Species Presence Data
2.3. Environmental Data Collection and Variable Selection
2.4. Preliminary Variables Processing
2.5. Model Calibration and Optimization
2.6. Reclassification of Predictions and Model Evaluation
3. Results
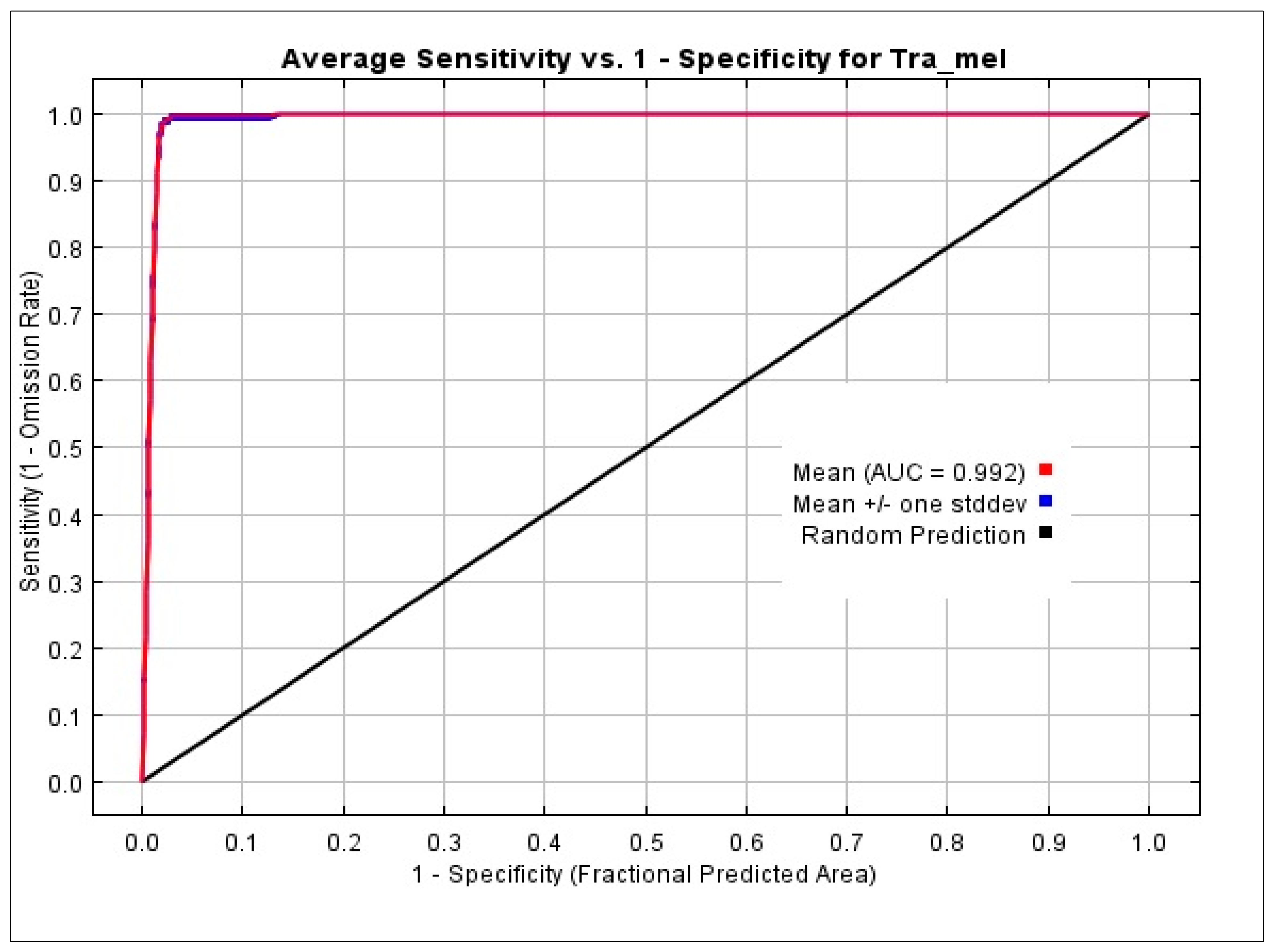
3.1. Model Evaluation
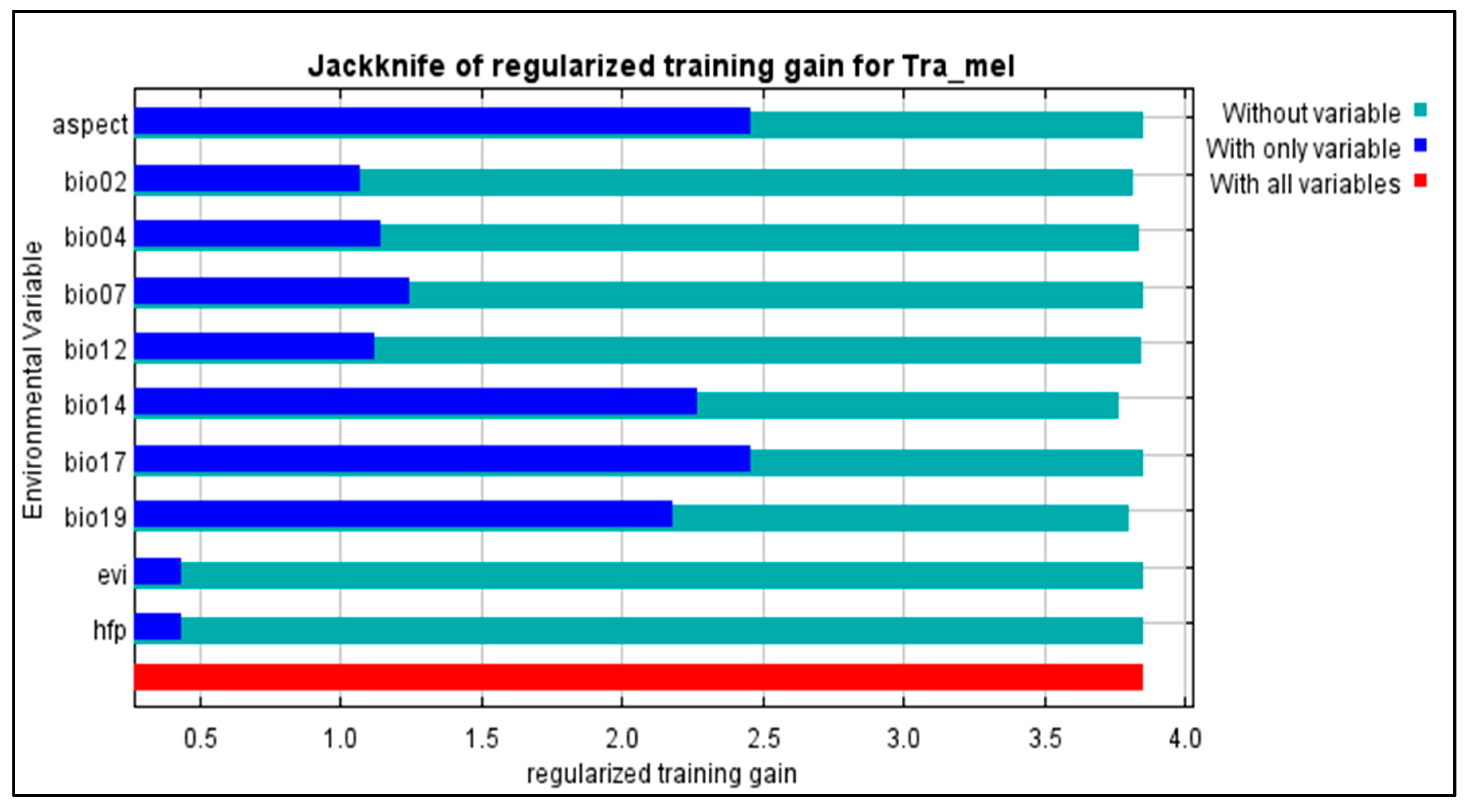
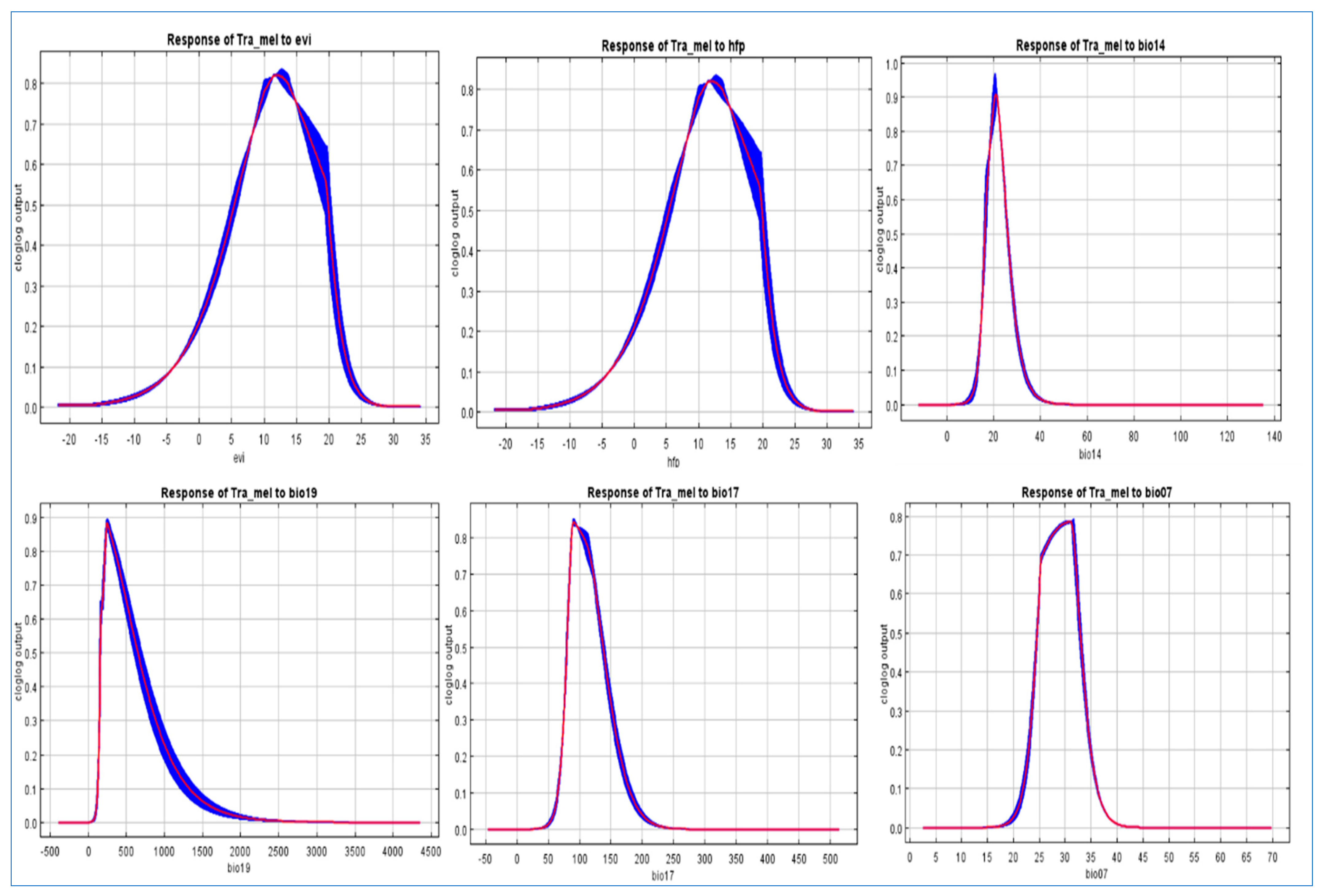
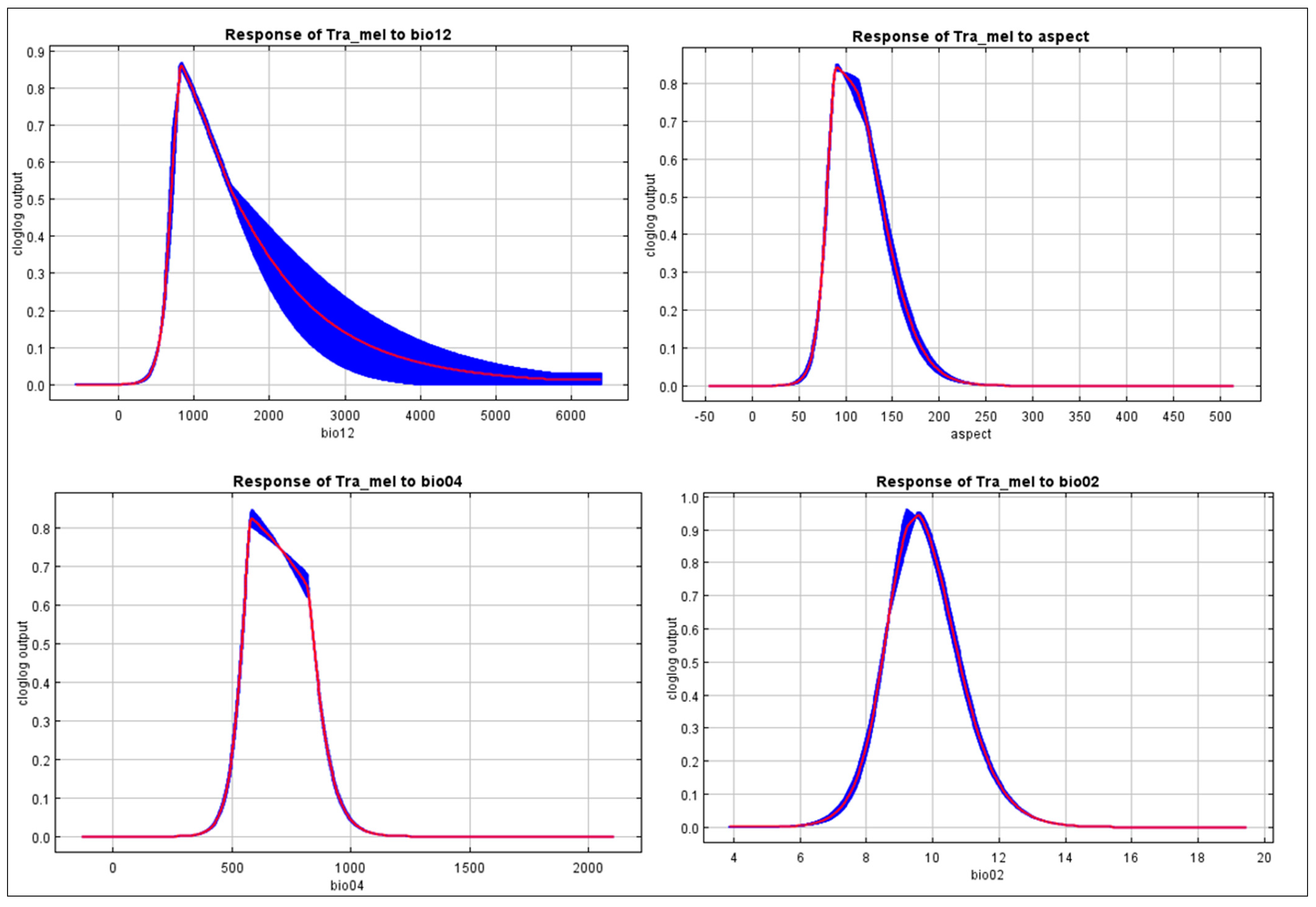
3.2. Elucidating the Ecological Factors Underlying the Spatial Distribution Patterns
3.3. Current Distribution
3.4. Suitability of Habitats under Future Scenarios of Climate Change
4. Discussion
Implications for Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.P.; Jackson, S.T.; House, J.I.; Prentice, I.C.; Mace, G.M. Beyond predictions: Biodiversity conservation in a changing climate. Science 2011, 332, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Cowie, R.H.; Bouchet, P.; Fontaine, B. The Sixth Mass Extinction: Fact, fiction or speculation? Biol. Rev. 2022, 97, 640–663. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.K.; Mishra, I. United Nations Sustainable Development Goals 2030 and environmental sustainability: Race against time. Environ. Sustain. 2019, 2, 339–342. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Cazalis, V.; Dudley, N.; Hoffmann, M.; Rodrigues, A.S.; Stolton, S.; Visconti, P.; Woodley, S.; Kingston, N.; Lewis, E.; et al. Area-based conservation in the twenty-first century. Nature 2020, 586, 217–227. [Google Scholar] [CrossRef]
- Haq, S.M.; Calixto, E.S.; Yaqoob, U.; Ahmed, R.; Mahmoud, A.H.; Bussmann, R.W.; Mohammed, O.B.; Ahmad, K.; Abbasi, A.M. Traditional usage of wild fauna among the local inhabitants of Ladakh, Trans-Himalayan Region. Animals 2020, 10, 2317. [Google Scholar] [CrossRef]
- Romero-Muñoz, A.; Benítez-López, A.; Zurell, D.; Baumann, M.; Camino, M.; Decarre, J.; del Castillo, H.; Giordano, A.J.; Gómez-Valencia, B.; Levers, C.; et al. Increasing synergistic effects of habitat destruction and hunting on mammals over three decades in the Gran Chaco. Ecography 2020, 43, 954–966. [Google Scholar] [CrossRef]
- Jump, A.S.; Peñuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Haq, S.M.; Waheed, M.; Ahmad, R.; Bussmann, R.W.; Arshad, F.; Khan, A.M.; Casini, R.; Alataway, A.; Dewidar, A.Z.; Elansary, H.O. Climate Change and Human Activities, the Significant Dynamic Drivers of Himalayan Goral Distribution (Naemorhedus goral). Biology 2023, 12, 610. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Pandey, P.C.; Sharma, J.K.; Triantakonstantis, D.; Srivastava, P.K. Climate Change and Its Impact on Forest of Indian Himalayan Region: A Review. In Climate Change. Springer Climate; Springer: Cham, Switzerland, 2022; pp. 207–222. [Google Scholar] [CrossRef]
- Lamprecht, A.; Pauli, H.; FernándezCalzado, M.R.; Lorite, J.; Molero Mesa, J.; Steinbauer, K.; Winkler, M. Changes in plant diversity in a water-limited and isolated high-mountain range (Sierra Nevada, Spain). Alp. Bot. 2021, 131, 27–39. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, M.; Harwood, T.D.; Hoskins, A.J.; Ware, C.; Hill, S.L.; Ferrier, S. Projecting impacts of global climate and land-use scenarios on plant biodiversity using compositional-turnover modelling. Glob. Chang. Biol. 2019, 25, 2763–2778. [Google Scholar] [CrossRef]
- Huang, C.; He, H.S.; Liang, Y.; Hawbaker, T.J.; Henne, P.D.; Xu, W.; Gong, P.; Zhu, Z. The changes in species composition mediate direct effects of climate change on future fire regimes of boreal forests in northeastern China. J. Appl. Ecol. 2021, 58, 1336–1345. [Google Scholar] [CrossRef]
- Schmeller, D.S.; Courchamp, F.; Killeen, G. Biodiversity loss, emerging pathogens and human health risks. Biodivers. Conserv. 2020, 29, 3095–3102. [Google Scholar] [CrossRef] [PubMed]
- Prober, S.M.; Doerr, V.A.; Broadhurst, L.M.; Williams, K.J.; Dickson, F. Shifting the conservation paradigm: A synthesis of options for renovating nature under climate change. Ecol. Monogr. 2019, 89, 01333. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.; Breman, E.; CecilioRebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G.; et al. Ten golden rules for reforestation to optimize carbon sequestration, biodiversity recovery and livelihood benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Seddon, N.; Smith, A.; Smith, P.; Key, I.; Chausson, A.; Girardin, C.; House, J.; Srivastava, S.; Turner, B. Getting the message right on nature-based solutions to climate change. Glob. Chang. Biol. 2021, 27, 1518–1546. [Google Scholar] [CrossRef]
- Waheed, M.; Arshad, F.; Majeed, M.; Haq, S.M.; Aziz, R.; Bussmann, R.W.; Ali, K.; Subhan, F.; Jones, D.A.; Zaitouny, A. Potential distribution of a noxious weed (Solanum viarum Dunal), current status, and future invasion risk based on MaxEnt modeling. Geol. Ecol. Landsc. 2023, 1–16. [Google Scholar] [CrossRef]
- Schroeder, B. Challenges of species distribution modeling belowground. J. Plant Nutr. Soil Sci. 2008, 171, 325–337. [Google Scholar] [CrossRef]
- Rather, Z.A.; Ahmad, R.; Khuroo, A.A. Ensemble modelling enables identification of suitable sites for habitat restoration of threatened biodiversity under climate change: A case study of Himalayan Trillium. Ecol. Eng. 2022, 176, 106534. [Google Scholar] [CrossRef]
- Awan, M.N.; Buner, F.; Kingdon, N. A review of published and unpublished surveys of a red-listed ‘flagship species’, the Western Tragopan melanocephalus in Azad Jammu and Kashmir, Pakistan. Bird Conserv. Int. 2016, 26, 380–395. [Google Scholar] [CrossRef]
- Shah, A.; Kayani, A.R.; Ihlow, F.; Nadeem, M.S.; Mahmood, T.; Islam, S.; Hausmann, A.E.; Päckert, M. Range-wide and regional distribution of the Western Tragopan melanocephalus and effects of disturbance on local abundances. Bird Conserv. Int. 2023, 33, e17. [Google Scholar] [CrossRef]
- Jameel, M.A.; Nadeem, M.S.; Aslam, S.; Ullah, W.; Ahmad, D.; Awan, M.N.; Masroor, W.; Mahmood, T.; Ullah, R.; Anjum, M.Z.; et al. Impact of human imposed pressure on pheasants of western Himalayas, Pakistan: Implication for monitoring and conservation. Diversity 2022, 14, 752. [Google Scholar] [CrossRef]
- Davis, A.E.; Gamble, R.; Roche, G.; Gawne, L. International relations and the Himalaya: Connecting ecologies, cultures and geopolitics. Aust. J. Int. Aff. 2021, 75, 15–35. Available online: https://search.informit.org/doi/10.3316/agispt.20210121042564 (accessed on 4 April 2022). [CrossRef]
- Dimri, A.P.; Yasunari, T.; Wiltshire, A.; Kumar, P.; Mathison, C.; Ridley, J.; Jacob, D. Application of regional climate models to the Indian winter monsoon over the western Himalayas. Sci. Total Environ. 2013, 468, S36–S47. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.; Srivastava, R.K. South Asia region and its urban agglomerates: The risk characterization. In Managing Urbanization, Climate Change and Disasters in South Asia; Springer: Singapore, 2020; pp. 23–78. [Google Scholar] [CrossRef]
- Hassan, M.; Haq, S.M.; Ahmad, R.; Majeed, M.; Sahito, H.A.; Shirani, M.; Mubeen, I.; Aziz, M.A.; Pieroni, A.; Bussmann, R.W.; et al. Traditional Use of Wild and Domestic Fauna among Different Ethnic Groups in the Western Himalayas—A Cross Cultural Analysis. Animals 2022, 12, 2276. [Google Scholar] [CrossRef]
- Gao, X.; Liu, J.; Huang, Z. The impact of climate change on the distribution of rare and endangered tree Firmianakwangsiensis using the Maxent modeling. Ecol. Evol. 2022, 12, 9165. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, T.; Xu, W.; Ouyang, Z. Assessment of habitat fragmentation caused by traffic networks and identifying key affected areas to facilitate rare wildlife conservation in China. Wildl. Res. 2015, 42, 266–279. [Google Scholar] [CrossRef]
- Arshad, F.; Waheed, M.; Fatima, K.; Harun, N.; Iqbal, M.; Fatima, K.; Umbreen, S. Predicting the suitable current and future potential distribution of the native endangered tree Tecomellaundulata (Sm.) Seem. in Pakistan. Sustainability 2022, 14, 7215. [Google Scholar] [CrossRef]
- Bosso, L.; Di Febbraro, M.; Cristinzio, G.; Zoina, A.; Russo, D. Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean Basin. Biol. Invasions 2016, 18, 1759–1768. [Google Scholar] [CrossRef]
- Hassan, T.; Hamid, M.; Wani, S.A.; Malik, A.H.; Waza, S.A.; Khuroo, A.A. Substantial shifts in flowering phenology of Sternbergiavernalis in the Himalaya: Supplementing decadal field records with historical and experimental evidences. Sci. Total Environ. 2021, 795, 148811. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Khattak, R.H.; Teng, L.; Ahmad, S.; Bari, F.; Rehman, E.U.; Shah, A.A.; Liu, Z. In pursuit of new spaces for threatened mammals: Assessing habitat suitability for Kashmir Markhor (Capra falconeri cashmeriensis) in the Hindukush Range. Sustainability 2022, 14, 1544. [Google Scholar] [CrossRef]
- Yin, A.; Harrison, T.M. Geologic evolution of the Himalayan-Tibetan orogen. Annu. Rev. Earth Planet. Sci. 2000, 28, 211–280. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Eyring, V.; Cox, P.M.; Flato, G.M.; Gleckler, P.J.; Abramowitz, G.; Caldwell, P.; Collins, W.D.; Gier, B.K.; Hall, A.D.; Hoffman, F.M.; et al. Taking climate model evaluation to the next level. Nat. Clim. Chang. 2019, 9, 102–110. [Google Scholar] [CrossRef]
- Awan, M.N.; Saqib, Z.; Buner, F.; Lee, D.C.; Pervez, A. Using ensemble modeling to predict breeding habitat of the red-listed Western Tragopan (Tragopan melanocephalus) in the Western Himalayas of Pakistan. Glob. Ecol. Conserv. 2021, 31, 01864. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Singh, H.; Kumar, N.; Kumar, M.; Singh, R. Modelling habitat suitability of western tragopan (Tragopan melanocephalus) a range-restricted vulnerable bird species of the Himalayan region, in response to climate change. Clim. Risk Manag. 2020, 29, 100241. [Google Scholar] [CrossRef]
- Bai, D.-F.; Chen, P.-J.; Atzeni, L.; Cering, L.; Li, Q.; Shi, K. Assessment of habitat Suitability of the Snow Leopard (Panthera uncia) in Qomolangma National Nature Reserve Based on MaxEnt Modeling. Zool. Res. 2018, 39, 373. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A Performance Assessment of Methods for Correcting Sampling Bias. PLoS ONE. 2014, 9, 97122. [Google Scholar] [CrossRef] [PubMed]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A. novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Summers, D.M.; Bryan, B.A.; Crossman, N.D.; Meyer, W.S. Species vulnerability to climate change: Impacts on spatial conservation priorities and species representation. Glob. Chang. Biol. 2012, 18, 2335–2348. [Google Scholar] [CrossRef]
- Díaz, S.; Malhi, Y. Biodiversity: Concepts, patterns, trends, and perspectives. Ann. Rev. Env. Resour. 2022, 47, 31–63. [Google Scholar] [CrossRef]
- Haq, S.M.; Yaqoob, U.; Calixto, E.S.; Kumar, M.; Rahman, I.U.; Hashem, A.; Abd_Allah, E.F.; Alakeel, M.A.; Alqarawi, A.A.; Abdalla, M.; et al. Long-term impact of transhumance pastoralism and associated disturbances in high-altitude forests of Indian Western Himalaya. Sustainability 2021, 13, 12497. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Ahmad, S.; Yang, L.; Khan, T.U.; Wanghe, K.; Li, M.; Luan, X. Using an ensemble modelling approach to predict the potential distribution of Himalayan gray goral (Naemorhedus goral bedfordi) in Pakistan. Glob. Ecol. Conserv. 2020, 21, 00845. [Google Scholar] [CrossRef]
- Zhao, D.; He, H.; Wang, W.; Wang, L.; Du, H.; Liu, K.; Zong, S. Predicting wetland distribution changes under climate change and human activities in a Mid- and High-Latitude Region. Sustainability 2018, 10, 863. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G. Habitat Suitability Modelling and Niche Theory. J. Appl. Ecol. 2008, 45, 1372–1381. [Google Scholar] [CrossRef]
- Jameel, M.A.; Khan, M.F.; Awan, M.N.; Nadeem, M.S.; Aslam, S.; Mehmood, S.; Ahmad, D.; Wali, R.; Rehman, Q.; Mahmood, T. Population and risk assessment of sympatric pheasant species in Palas Valley, Pakistan. Braz. J. Biol. 2022, 84. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.M.; Khuroo, A.A.; Malik, A.H.; Rashid, I.; Ahmad, R.; Hamid, M.; Dar, G.H. Forest ecosystems of Jammu and Kashmir state. In Biodiversity of the Himalaya: Jammu and Kashmir State; Springer: Singapore, 2020; pp. 191–208. [Google Scholar] [CrossRef]
- Domíguez-Vega, H.; Monroy-Vilchis, O.; Balderas-Valdivia, C.J.; Gienger, C.M.; Ariano-Sánchez, D. Predicting the potential distribution of the beaded lizard and identification of priority areas for conservation. J. Nat. Conserv. 2012, 20, 247–253. [Google Scholar] [CrossRef]
- Smeraldo, S.; Di Febbraro, M.; Ćirović, D.; Bosso, L.; Trbojević, I.; Russo, D. Species distribution models as a tool to predict range expansion after reintroduction: A case study on Eurasian beavers (Castor fiber). J. Nat. Conserv. 2017, 37, 12–20. [Google Scholar] [CrossRef]
- Feeley, K.J.; Silman, M.R.; Bush, M.B.; Farfan, W.; Cabrera, K.G.; Malhi, Y.; Saatchi, S. Upslope migration of Andean trees. J. Biogeogr. 2011, 38, 783–791. [Google Scholar] [CrossRef]
- Forero-Medina, G.; Terborgh, J.; Socolar, S.J.; Pimm, S.L. Elevational ranges of birds on a tropical montane gradient lag behind warming temperatures. PLoS ONE 2011, 6, 28535. [Google Scholar] [CrossRef] [PubMed]
- Zuckerberg, B.; Woods, A.M.; Porter, W.F. Poleward shifts in breeding bird distributions in New York State. Glob. Chang. Biol. 2009, 15, 1866–1883. [Google Scholar] [CrossRef]
- Chhatre, A.; Saberwal, V. Democracy, development and (Re-) visions of nature: Rural conflicts in the western Himalayas. J. Peasant. Stud. 2006, 33, 678–706. [Google Scholar] [CrossRef]
- Inskipp, C.; Baral, H.S.; Inskipp, T.; Khatiwada, A.P.; Khatiwada, M.P.; Poudyal, L.P.; Amin, R. Nepal’s National Red List of Birds. J. Threat. Taxa 2017, 9, 9700–9722. [Google Scholar] [CrossRef]
- Quintero, I.; Wiens, J.J. Rates of projected climate change dramatically exceed past rates of climatic niche evolution among vertebrate species. Ecol. Lett. 2013, 16, 1095–1103. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution: Int. J. Org. Evol. 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, M.; Liu, Y. Avian diversity and distributions and their evolution through space and time. Bird Species: How They Arise, Modify and Vanish. Life Sci. 2018, 129–145. [Google Scholar] [CrossRef]
- Muñoz-Mendoza, C.; D’Elía, G.; Panzera, A.; Villalobos-Leiva, A.; Sites, J.W., Jr.; Victoriano, P.F. Geography and past climate changes have shaped the evolution of a widespread lizard from the Chilean hotspot. Mol. Phylogenetics Evol. 2017, 116, 157–171. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Foden, W.B.; Butchart, S.H.; Stuart, S.N.; Vié, J.C.; Akçakaya, H.R.; Angulo, A.; Mace, G.M. Identifying the world’s most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE 2013, 8, 65427. [Google Scholar] [CrossRef] [PubMed]
- Abolmaali, S.M.R.; Tarkesh, M.; Bashari, H. MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inf. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecol. 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Li, B.; Liang, C.; Song, P.; Liu, D.; Qin, W.; Jiang, F.; Zhang, T. Threatened birds face new distribution under future climate change on the Qinghai-Tibet Plateau (QTP). Ecol. Indic. 2023, 150, 110217. [Google Scholar] [CrossRef]
- Kadmon, R.; Farber, O.; Danin, A. Effect of roadside bias on the accuracy of predictive maps produced by bioclimatic models. Ecol. Appl. 2004, 14, 401–413. [Google Scholar] [CrossRef]
- Segurado, P.; Araujo, M.B.; Kunin, W.E. Consequences of spatial autocorrelation for niche-based models. J. Appl. Ecol. 2006, 43, 433–444. [Google Scholar] [CrossRef]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Jameel, M.A.; Tabassum, S.; Mehmood, S.; Shah, T.; Khan, M.F.; Kabir, M.; Awan, M.N. Does Trophy Hunting of Kashmir Markhor Really Contributing in Its Conservation? Int. J. Conserv. Sci. 2019, 10, 525–532. [Google Scholar] [CrossRef]
- Yamaura, Y.; Higa, M.; Senzaki, M.; Koizumi, I. Can charismatic megafauna be surrogate species for biodiversity conservation? Mechanisms and a test using citizen data and a hierarchical community model. In Biodiversity Conservation Using Umbrella Species: Blakiston’s Fish Owl and the Red-Crowned Crane; Springer: Singapore, 2018; pp. 151–179. [Google Scholar] [CrossRef]
- Rahman, Q.; Nadeem, M.S.; Umair, M.; Altaf, M.; Ni, J.; Abbasi, A.M.; Jameel, M.A.; Pieroni, A.; Hamed, M.H.; Ashraf, S.; et al. Medicinal water birds in the traditional healthcare system: An assessment of biodiversity–cultural linkages in Eastern Khyber Pakhtunkhwa, Pakistan. J. Ethnobiol. Ethnomedicine 2022, 18, 57. [Google Scholar] [CrossRef] [PubMed]
- Tietze, D.T. Bird Species: How They Arise, Modify and Vanish; Springer Nature: Cham, Switzerland, 2018; Available online: http://library.oapen.org/handle/20.500.12657/22941 (accessed on 4 April 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Name of Variable & Description | Code | Resolution | Database |
|---|---|---|---|---|
| Climatic Variables | Annual Mean Temperature | Bio1 | 30 arc s | WorldClim |
| Mean Diurnal Range | Bio2 | 30 arc s | WorldClim | |
| Temperature Seasonality (sd ×100) | Bio4 | 30 arc s | WorldClim | |
| Isothermality (Bio2/Bio7) (×100) | Bio3 | 30 arc s | WorldClim | |
| Mean Temperature of Wettest Quarter | Bio8 | 30 arc s | WorldClim | |
| Mean Temperature of Driest Quarter | Bio9 | 30 arc s | WorldClim | |
| Mean Temperature of Warmest Quarter | Bio10 | 30 arc s | WorldClim | |
| Mean Temperature of Coldest Quarter | Bio11 | 30 arc s | WorldClim | |
| Temperature Annual Range | Bio7 | 30 arc s | WorldClim | |
| Max. Temperature of Warmest Month | Bio5 | 30 arc s | WorldClim | |
| Min. Temperature of Coldest Month | Bio6 | 30 arc s | WorldClim | |
| Annual Precipitation | Bio12 | 30 arc s | WorldClim | |
| Precipitation of Wettest Month | Bio13 | 30 arc s | WorldClim | |
| Precipitation of Driest Month | Bio14 | 30 arc s | WorldClim | |
| Precipitation of Wettest Quarter | Bio16 | 30 arc s | WorldClim | |
| Precipitation of Driest Quarter | Bio17 | 30 arc s | WorldClim | |
| Precipitation Seasonality (CV) | Bio15 | 30 arc s | WorldClim | |
| Precipitation of Warmest Quarter | Bio18 | 30 arc s | WorldClim | |
| Precipitation of Coldest Quarter | Bio19 | 30 arc s | WorldClim | |
| Vegetation | Enhanced Vegetation Index | EVI | 30 arc s | Moderate Resolution Imaging Spectroradiometer MODIS/Terr |
| Topographical Variables | Elevation | Elev | 30 arc s | SRTM DEM Global |
| Aspect | Aspect | 30 arc s | SRTM DEM Global | |
| Slope | Slope | 30 arc s | SRTM DEM Global | |
| Anthropogenic Variables | Human footprint | HFP | 30 arc s | http://sedac.ciesin.columbia.edu (accessed on 2 June 2023) |
| Road proximity | RP | 30 arc s | http://sedac.ciesin.columbia.edu (accessed on 2 June 2023) | |
| Land cover | LC | 30 arc s | http://www-modis.bu.edu/landcover (accessed on 2 June 2023) | |
| Population density | PD | 30 arc s | http://www.ornl.gov/sci/landscan (accessed on 2 June 2023) |
| Description | Code | Percent Contribution |
|---|---|---|
| Aspect | Aspect | 36.5 |
| Precipitation of Coldest Quarter | Bio19 | 34.7 |
| Mean Diurnal Range | Bio02 | 13.3 |
| Enhanced Vegetation Index | Evi | 7.6 |
| Precipitation of Driest Month | Bio14 | 5.2 |
| Temperature Seasonality (sd × 100) | Bio04 | 1.7 |
| Annual Precipitation | Bio12 | 0.4 |
| Human Footprint | Hfp | 0.4 |
| Precipitation of Driest Quarter | Bio17 | 0.1 |
| Temperature Annual Range | Bio07 | 0.1 |
| Climate Change Scenario | The Probability of Western Tragopan Occurrence within the Selected Habitat Suitability Categories Has Been Estimated | |||||
|---|---|---|---|---|---|---|
| Not-Suitable (NS) | Least (LS) | Moderate (MS) | High (HS) | Very-High (VHS) | Total Suitable Land Area (km2) | |
| (p ≤ 0.2) | (p 0.21–0.4) | (p 0.41–0.6) | (p 0.61–0.8) | (p ≥ 0.81) | ||
| Current climate | 434,734 | 16,334 | 18,276 | 15,387 | 9489 | 59,486 |
| SSPs_245_2050 | 446,863 | 12,778 | 13,778 | 12,963 | 7838 | 47,357 |
| Rate of change (%) | 2.45 | −0.72 | −0.91 | −0.49 | −0.33 | −2.45 |
| SSPs_585_2050 | 453,175 | 11,234 | 12,422 | 11,233 | 6156 | 41,045 |
| Rate of change (%) | 3.7 | −1.0 | −1.2 | −0.8 | −0.7 | −3.7 |
| SSPs_245_2070 | 449,346 | 12,345 | 14,267 | 11,995 | 6267 | 44,874 |
| Rate of change (%) | 2.96 | −0.81 | −0.81 | −0.69 | −0.65 | −2.96 |
| SSPs_585_2070 | 460,430 | 9027 | 9138 | 10,267 | 5358 | 33,790 |
| Rate of change (%) | 5.20 | −1.48 | −1.85 | −1.04 | −0.84 | −5.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jameel, M.A.; Nadeem, M.S.; Haq, S.M.; Mubeen, I.; Shabbir, A.; Aslam, S.; Ahmad, R.; Gaafar, A.-R.Z.; Al-Munqedhi, B.M.A.; Bussmann, R.W. Shifts in the Distribution Range and Niche Dynamics of the Globally Threatened Western Tragopan (Tragopan melanocephalus) Due to Climate Change and Human Population Pressure. Biology 2023, 12, 1015. https://doi.org/10.3390/biology12071015
Jameel MA, Nadeem MS, Haq SM, Mubeen I, Shabbir A, Aslam S, Ahmad R, Gaafar A-RZ, Al-Munqedhi BMA, Bussmann RW. Shifts in the Distribution Range and Niche Dynamics of the Globally Threatened Western Tragopan (Tragopan melanocephalus) Due to Climate Change and Human Population Pressure. Biology. 2023; 12(7):1015. https://doi.org/10.3390/biology12071015
Chicago/Turabian StyleJameel, Muhammad Azhar, Muhammad Sajid Nadeem, Shiekh Marifatul Haq, Iqra Mubeen, Arifa Shabbir, Shahzad Aslam, Riyaz Ahmad, Abdel-Rhman Z. Gaafar, Bander M. A. Al-Munqedhi, and Rainer W. Bussmann. 2023. "Shifts in the Distribution Range and Niche Dynamics of the Globally Threatened Western Tragopan (Tragopan melanocephalus) Due to Climate Change and Human Population Pressure" Biology 12, no. 7: 1015. https://doi.org/10.3390/biology12071015
APA StyleJameel, M. A., Nadeem, M. S., Haq, S. M., Mubeen, I., Shabbir, A., Aslam, S., Ahmad, R., Gaafar, A.-R. Z., Al-Munqedhi, B. M. A., & Bussmann, R. W. (2023). Shifts in the Distribution Range and Niche Dynamics of the Globally Threatened Western Tragopan (Tragopan melanocephalus) Due to Climate Change and Human Population Pressure. Biology, 12(7), 1015. https://doi.org/10.3390/biology12071015