Simple Summary
Beneficial microorganisms, such as microalgae and bacteria, have a strong ability to restore health and fertility in degraded soils. However, the use of these microorganisms interacting in a mixed consortium has yet to be well explored. Furthermore, most of the current knowledge on the effects of these microorganisms on soil fertility derives from studies focused on the potential of either of these groups as biofertilizers; thus, more information on their real impact on degraded soils is required. This mini-review addresses the current knowledge on using a consortium of microalgae and bacteria for this purpose.
Abstract
Soil restoration is one of the biggest challenges of this century. Besides the negative impacts of climate change, the current increase in food demands has put severe pressure on soil resources, resulting in a significant area of degraded land worldwide. However, beneficial microorganisms, such as microalgae and plant growth-promoting bacteria, have an outstanding ability to restore soil health and fertility. In this mini-review, we summarize state-of-the-art knowledge on these microorganisms as amendments that are used to restore degraded and contaminated soils. Furthermore, the potential of microbial consortia to maximize beneficial effects on soil health and boost the production of plant-growth-promoting compounds within a mutualistic interaction is discussed.
1. Introduction
The ability to meet the continuous increase in food demands as a consequence of population growth is one of the biggest challenges of this century. Climate change and freshwater limitations are realities that we must consider when developing sustainable agricultural systems [1]. The current increase in food demands has put severe pressure on soil resources, resulting in significant areas of degraded soil worldwide due to intensive and poor agricultural land management [2,3]. Similarly, industrial activities, modern agricultural practices, improper waste disposal, and accidental spills of hazardous substances result in soil contamination [4,5]. Moreover, climate change alters fire regimes, significantly affecting soils and ecosystems [6]. Sadly, more than 33% of global land is degraded, and this percentage will grow if no actions are taken to prevent and reverse the degradation [7].
One of the first steps in tackling the increasing food demands is restoring degraded soils (i.e., degraded farmlands, contaminated soils, and post-fire ecosystems). The use of organic amendments, such as microalgae, especially cyanobacteria, is an increasing field of study that aims to restore soil health and fertility [6,8,9]. These microorganisms have the capability to restore soil structure and aggregate stability by releasing exopolysaccharides and forming soil aggregates, providing O2 to the subsurface, solubilizing and mobilizing macro- and micronutrients, mineralizing simpler organics, and serving as a source of organic matter and nutrients [10,11]. Additionally, the fertilizer potential of these microorganisms is well documented. Microalgae, including cyanobacteria, contain some plant-growth-promoting substances, such as phytohormones (auxins, cytokinins, abscisic acid, ethylene, and gibberellins), amino acids, vitamins, polyamines, betaines, protein hydrolysates, and polysaccharides, which can be used as biostimulants [12]. Furthermore, microalgae extracts have a recognized potential to improve soil physical and biological properties by acting as organic slow-release fertilizers that can return nutrients (carbon and macro-elements) and ensure the efficient use of resources [13].
Plant-growth-promoting bacteria (PGPB) are bacterial strains isolated from diverse environments with the potential to positively influence the growth and yield of diverse plants, mostly of agricultural importance [14]. In recent years, the use of PGPB has been explored beyond agricultural practices toward environmental applications, such as the restoration of eroded desert soils [15,16]. Inoculation with PGPB affects the growth and metabolism of plants through a multitude of mechanisms, such as nitrogen fixation, phosphorus, iron solubilization, the production of indoleacetic acid (IAA) and other plant hormones, the production of signal molecules, and the mitigation of environmental stresses [17,18]. Several comprehensive and critical reviews describing the operational mechanisms of PGPB have been published [19,20]. Consequently, a general discussion of the likely mechanisms for promoting plant growth is not described in this review.
The use of microalgae and bacteria for the bioremediation of contaminated soils has also been explored, with promising results. Contaminants, such as heavy metals, pesticides, and hydrocarbons, can have adverse effects on soil quality and ecosystem health [4,5]. These microorganisms offer an environmentally friendly and cost-effective solution for reducing soil pollution by breaking down pollutants and transforming them into less harmful forms [21,22]. The use of these natural agents in soil remediation strategies has the potential to reduce reliance on chemical-based treatments and to promote the development of more sustainable and eco-friendly agricultural practices.
Despite their proven fertilizer and bioremediation potentials, the capacity of these microorganisms to restore degraded soils needs to be better explored. Most studies have focused on the positive effects of microalgae and PGPB on plant growth, and all aspects regarding soil health and fertility still need to be assessed. Moreover, the use of microalgae and bacteria consortia for soil restorations has been poorly studied, even though their synergistic interaction can significantly boost their positive impact on soil fertility. Microalgae and bacteria exhibit positive interactions through substrate exchange, cell-to-cell communication via small signaling molecules, and horizontal gene transfer, conferring adaptive advantages to environmental stressors [23,24].
This mini-review discusses the current knowledge of microalgae (including cyanobacteria) and PGPB as promoters of soil recovery. Furthermore, we explore the use of artificial microalgae–bacteria consortia as a promising organic amendment to restore marginal and degraded soils.
2. Climate Change and Soil Degradation
Soil degradation is an increasing threat of climate change that entails the loss of biodiversity and ecosystem services [25,26]. Soils provide valuable ecosystem services, such as sustainable plant production, the control of water quality, the control of biological pests and diseases, the filtering of nutrients and contaminants, carbon storage, greenhouse gas regulation, waste detoxification and recycling, and flood and climate change mitigation, among others [27,28,29,30]. Besides the obvious negative environmental impacts of dropping such services, there is also an important economic cost to society [31,32]. Additionally, when the soil becomes degraded, it loses its ability to support a diverse range of microorganisms, leading to a decline in soil microbial diversity, complexity, and functionality [33,34,35]. This, in turn, can have far-reaching consequences for the health of ecosystems and for human health and well-being. The loss of soil microbial diversity can disrupt soil ecosystem functioning by altering nutrient cycling, decreasing soil fertility, and increasing the risk of soil erosion and pollution [36,37,38]. Therefore, soil microbial biomass is widely regarded as an indicator of soil fertility and ecosystem productivity, and is a fundamental characteristic that plays a key role in soil restoration [39].
Since population growth is associated with ever-increasing food and water consumption, one of the biggest challenges that humanity is facing in the current climate change scenario is the increased degradation of arable land around the world, which is linked to food and nutritional insecurity [40,41]. Furthermore, exposure to soil contaminants and the adverse effects of their toxicity will be impacted by climate change. Changes in environmental conditions might modify the mobilization, transfer, behavior, concentration, deposition, and fate of pollutants [42,43]. Additionally, increased temperatures and salinity might increase the chemical toxicity of certain soil contaminants by altering the contaminant’s biotransformation into more bioactive metabolites [43].
There is a real threat of human displacement during the next decade because of land degradation, which encroaches on over one-third of global land [44]. In a comprehensive review, Prăvălie [45] reported several pressures around the globe that result in land degradation, such as coastal erosion, biological invasions, aridity, land erosion by water or wind, land subsidence, landslides, permafrost thawing, soil biodiversity loss, soil compaction, soil organic carbon loss, waterlogging, salinization, soil sealing, and vegetation degradation. Aridity, land erosion by water, salinization, soil organic carbon loss, and vegetation degradation were described as major degradation pathways. Land degradation leads to the transformation of grasslands into hyper-arid and desert environments, reaching up to 41.3% of the global land’s surface [46]. Furthermore, globally, the main pressures driving arable land degradation are aridity and soil erosion, which affect 40% and 20% of arable land, respectively [47].
The world’s degraded land mapped by Gibbs and Salmon [2] provides a general idea of the complex situation in many parts of the world. Ironically, global warming not only triggers land degradation but also renders degraded land less resilient and highly vulnerable to climate change [41]. Furthermore, since agricultural production is limited by the increasing scarcity and diminishing quality of land and water resources [48], the required increment in production must align with sustainable agricultural techniques and the conscious use of water resources. Avoiding agricultural expansion at the expense of forestland is mandatory, which only intensifies the environmental threat [2,47]. In this sense, restoration plans should focus on recuperating native ecosystems rather than expanding agricultural land in areas that were once forests or savannas [2].
3. Use of PGPB and Microalgae for Restoration of Degraded Soil
PGPB are beneficial in harsh and limiting environments because of their role in alleviating stress in plants, making them excellent candidates to assist revegetation of eroded zones. For instance, PGPB can help plants tolerate drought stress by improving their water and nutrient uptake [49]. There are several examples of soil restoration with plants inoculated with PGPB; a severely eroded land in the southern Sonoran Desert was restored using native leguminous trees and the giant cardon cactus inoculated with two PGPB (Azospirillum brasilense and Bacillus pumilus), native arbuscular mycorrhizal fungi, and small quantities of compost [16,50]. Over a decade later, highly eroded land, destroyed for 25 years with almost no topsoil and extremely low mineral quantities to support plant growth, was successfully recovered. Likewise, the outdoor nursery cultivation of mesquite tree transplants was evaluated as a way to restore arid zones [51]. The study showed that inoculating the seedlings with PGPB—A. brasilense immobilized in dry alginate microbeads—resulted in the enhancement of all growth parameters of the plants, including biomass, aerial volume, root system, and chlorophyll pigments. Ramachandran and Radhapriya [52] explored a similar approach in a highly degraded forest in the Nanmangalam Reserve Forest in the Eastern Ghats of India. The authors planted 12 native tree species inoculated with a consortium of five native types of PGPB, small amounts of compost, and chemical fertilizer. The results of an experiment that lasted almost three years revealed that the PGPB consortium enhanced plant biomass in all the native plants and improved soil quality in the degraded forest. Schoebitz et al. [53] evaluated the combined effect of A. brasilense, Pantoea dispersa, and an organic olive residue immobilized in clay in the revegetation of semiarid land. The study revealed that PGPB improved soil properties by increasing phosphorus and potassium content availability by up to 100% and 70%, respectively. The inoculant also increased the total carbon and microbial biomass carbon content and enzyme activities, such as dehydrogenase, urease, and protease.
High soil salinity is another undesirable feature that reduces soil fertility [54]. However, salt-tolerant PGPB can significantly enhance salt tolerance in plants through several mechanisms, such as the adjustment of osmosis, protection from free radicals, the excretion of phytohormones that enhance growth parameters, and the release of extracellular polymeric substances (EPSs) that bind with Na+ cations, decreasing its bioavailability for plant uptake [55,56,57]. For instance, the PGPB Bacillus pumilus strain JPVS11, improved the growth performance of rice (Oryza sativa L.), which was negatively impacted by high soil salinity [58]. The study also revealed a significant improvement in soil enzyme activities of up to 56%, 46%, 48%, and 56% in alkaline phosphatase, acid phosphatase, urease, and β-glucosidase, respectively. Likewise, Hafez et al. [59] evaluated the potential of PGPB—Azospirillum brasilense—to restore saline–sodic soils. Following the inoculation of the strain with eco-friendly organic wastes for 150 days, the authors reported that soil fertility was enhanced with increases in soil organic carbon, dehydrogenase, urease enzymes, micronutrients (Fe, Zn, Mn, Cu, and B), and macronutrients (N, P, and K).
The soil restoration potential of microalgae, especially cyanobacteria, has been studied far more extensively because of the protagonist-like role they play in biological soil crust or biocrust. Biocrust corresponds to a cohesive and thin horizontal ground cover composed of photosynthetic organisms, such as lichens, bryophytes, and microalgae, and their associated bacteria, archaea, and fungi, which are of uttermost importance in stabilizing the soil against erosion [6,60]. Cyanobacteria, i.e., the first colonizers, stabilize the topmost layers and facilitate the formation of the soil crust with other microalgae groups and bacteria [61]. This is particularly important in arid or semiarid lands, desertified soils, and soils affected by fire, where cyanobacteria can be a suitable soil amendment that increases nutrient availability and promotes plant growth [61]. Additionally, microalgae act as biostimulants, affecting soil biological activity by enhancing enzymatic activity [62].
Wang et al. [63] reported the suitability of an artificial consortium composed of the cyanobacterial species Microcoleus vaginatus and Scytonema javanicum to recover the biological soil crust of degraded soil in a desert area in Inner Mongolia. After cyanobacterial inoculation, the authors reported a significant increase in total nitrogen, organic carbon, total salt, calcium carbonate, and electrical conductivity. The inoculation of this consortium with the plant Salix mongolica was later evaluated by Lan et al. [64]. Cyanobacteria inoculation quickly formed a biocrust and gradually gave rise to the moss crust, helping vascular plants to regenerate. A similar study reported on the inoculation of a cyanobacterial consortium with the species Anabaena doliolum, Cylindrospermum sphaerica, and Nostoc calcicole in a semiarid clay–loam soil, improved carbon and nitrogen mineralization, increased water-holding capacity, and enhanced hydraulic conductivity. Additionally, in response to the cyanobacterial biofertilizer, pear millet and wheat crops showed an increase in their growth and yield [65]. Another artificial consortium co-formed by the filamentous cyanobacteria Microcoleus vaginatus, Phormidium tenue, Scytonema javanicum, Nostoc spp., and the chlorophycea Desmococcus olivaceus efficiently assisted in the stabilization of fine sands, helping to control erosion in aeolian sandy soil in the south-eastern region of the Tenger Desert [66]. Similarly, Issa et al. [67] evaluated the effect of the cyanobacteria Nostoc spp. on the structural stability of poorly aggregated tropical soil from the Eastern Cape Province of South Africa. Cyanobacterial inoculation increased the resistance of soil aggregates to break down, enhancing soil stability two to four times over the control after six weeks of inoculation. Another study reported the potential of the acid-tolerant microalgae species Desmodesmus spp. and Heterochlorella spp., alone, or in combination, to improve soil health and fertility. The inoculation of strains in two acid soils (Kurosol and Podosol) collected from Queensland, Australia, resulted in the development of algal soil crust. Additionally, the authors reported an increase in the release of exopolysaccharides (more than 200%) which facilitate soil stability, an increase in carbon content (up to a 57%), an increase in dehydrogenase activity (more than 500%), and an increased production of indolacetic acid (between 200 and 500%) [68]. Furthermore, the algalization of acid soils with these species enhanced the richness of ecologically important soil bacteria, such as rhizobacteria and diazotrophs [69]. Muñoz-Rojas et al. [70] also evaluated the potential of a cyanobacteria consortium with Nostoc commune, Tolypothrix distorta, and Scytonema hyalinum to restore mine soil and reported that up to 40% of the soil surface was covered by biocrust after 90 days, as well as a significant increase in soil organic carbon and the promotion of C sequestration.
All of these studies have revealed that the use of PGPB and microalgae, particularly cyanobacteria, is an effective approach in restoring degraded soils, increasing soil fertility, stabilizing the soil against erosion, and promoting plant growth in arid and semiarid regions. Because the inoculation of PGPB and microalgae can be a sustainable and eco-friendly strategy for soil restoration programs, the use of these microorganisms in consortia has also been explored to further enhance the positive effects of these beneficial microorganisms.
4. Microalgae–Bacteria Consortia as Inoculants to Restore Degraded Soils
Microbial consortia have several advantages over individual species, such as strength to environmental fluctuations, the ability to survive nutrient starvation periods by sharing metabolites, and resistance to invasion by other species [71]. The co-inoculation of different species of microalgae or bacteria in microbial consortia has shown the enhanced positive effects that each species alone has on soil fertility. The use of these microbial consortia, especially cyanobacteria species, for the restoration of degraded soils leads to significant enhancements in several traits of soil fertility (Table 1).

Table 1.
Microbial consortia of microalgae or bacteria with proven potential to enhance the characteristics of degraded soils.
Furthermore, a mutualistic consortium between microalgae and bacteria can boost their metabolism [24]. Mutualism is a positive interaction between organisms of two different species in which each benefits from, and is based on, the exchange of resources and services. A mutualistic microalgae–bacteria consortium is based on the exchange of metabolites; the best-described mechanism of bacterial improvement in the growth of microalgae is supplementation with CO2, whereby the growth of microalgae in an environment with reduced O2 tension and enriched CO2 will increase the primary metabolism and promote an increase in their populations [72]. In return, the bacterial growth can be stimulated by the uptake of extracellular polymeric substances (EPSs) released by the microalgae, including organic carbon (myo-inositol or lactate), proteins, and amino acids (such as tryptophan) [73,74].
Alternatively, a microalgae–bacteria mutualism is established by exchanging photosynthates produced by the microalgae and vitamins produced by the bacteria, which are needed as co-factors for enzymes in key metabolic pathways in the microalgae. For example, many microalgae are auxotrophs for vitamin B12 (cobalamin) and vitamin B1 (thiamine). Because vitamin B12 is only produced by prokaryotes, it becomes the source of vitamins for the microalgae. Palacios et al. [75] reported that the production of vitamin B2 (riboflavin) and its degradation compound, lumichrome, using Azospirillum brasilense positively affected the microalga Chlorella sorokiniana. Furthermore, growth-promoting factors produced by bacteria, such as phytohormones IAA, gibberellins [76,77,78], or organic nitrogen compounds (such as uracil) [74], have been proven to have positive effects on microalgae.
This significant enhancement in the bioactivity of both partners may lead to the enhanced capacity of microorganisms as agents used for soil restoration, as proposed in Figure 1. However, most reports of mutualism have focused on eukaryotic microalga and bacteria, and a limited number of reports found in the literature on the use of these consortia for enhancing soil characteristics mostly present the combined use of cyanobacteria and bacteria (Table 2). For example, the co-cultivation of the cyanobacteria Anabaena variabilis and Nostoc calsicola with the green microalgae Chlorella vulgaris and the nitrogen-fixing bacteria Azotobacter spp. in two different consortia (each with one cyanobacterium species) revealed the improved growth of microorganisms with the bacteria, suggesting an increase in the nitrogen-fixing activity of the consortia, which might lead to an enhancement in soil fertility [79]. In another study, Swarnalakshmi et al. [80] evaluated novel biofilm preparations using cyanobacterium Anabaena torulosa as a matrix for diazotrophic and phosphate-solubilizing bacteria. The authors assessed the fertilizer potential of biofilms on wheat crops and reported a significant increase in the available nitrogen, even after 14 weeks of inoculation. Similarly, the inoculation of A. torulosa–Azotobacter chroococcum biofilm on leguminous crops resulted in increases of 80% and 24% of available nitrogen and phosphorous, respectively. In the same study, A. torulosa–Bradyrhizobium spp. biofilm increased N2 fixation, N mobilization, and soil C sequestration [81].
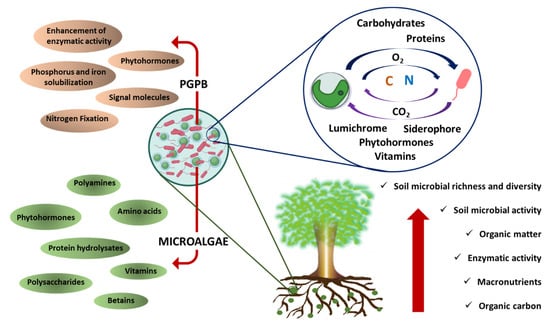
Figure 1.
Schematic diagram of the biostimulant activity of a microalgae–bacteria consortium to restore degraded soil.
Table 2.
Microalgae–bacteria consortia used as inoculants with a proven ability to enhance soil fertility.
Significant enhancements in microbiological and enzymatic activities have also been described following the inoculation of microbial consortia. Manjunath et al. [82] reported that the inoculation of microbial consortia (a mix of proteobacterial and cyanobacterial strains) in wheat crops enhanced dehydrogenase activity and soil microbial activity. Similarly, the inoculation of Anabaena torulosa in consortium with either Rhizobium spp. or Pseudomonas fluorescens in leguminous crops (chickpea, pea, and lentils) led to an increase in soil polysaccharides, dehydrogenase and nitrogenase activity, soil carbon, and available P [87]. Bidyarani et al. [88] evaluated the same cyanobacterial species in consortium with Mesorhizobium cicero inoculated in chickpea crops. The authors reported a significant increase in the soil available N and P, N2 fixation, and dehydrogenase activity. Another study of Anabaena sp. in consortium with the bacteria Providencia spp. revealed that the consortium inoculated in wheat seeds produced significant increases in alkaline phosphatase and dehydrogenase activities and microbial biomass carbon in the soil [86].
Microalgae–bacteria inoculants can also help to control soil erosion induced by rainfall. For example, the direct inoculation of selected strains of cyanobacteria (Lyngbya spp., Nostoc spp., and Oscillatoria spp.), and bacteria (Azotobacter spp. and Bacillus subtilis) in soil collected from an area highly susceptible to erosion effectively increased soil stability, leading to a 99% soil loss reduction rate [85].
In a few reports on the consortia between eukaryotic microalgae and bacteria, Trejo et al. [89] proved the efficiency of Chlorella sorokiniana and Azospirillum brasilense debris as an amendment for infertile soils with low levels of organic matter. After wastewater treatment, the inoculation of dried alginate beads containing C. sorokiniana and A. brasilense significantly increased the organic matter, organic carbon, and microbial carbon of eroded infertile desert soil. Later, Lopez et al. [84] revealed a significant increase in soil microbial richness and diversity following the inoculation of the same microbial consortium. Additionally, the growth of sorghum in the amended soil was greater, and the deep colonization of the root surface using A. brasilense was observed.
In summary, co-inoculating different species of microalgae and bacteria has been shown to enhance the positive effects of each species on soil fertility. Additionally, the mutualistic consortium between microalgae and bacteria can boost their metabolisms through the exchange of metabolites, photosynthates, and vitamins, thus enhancing the capacity of microorganisms as agents for soil restoration.
5. Microalgae and Bacteria for Bioremediation
Besides the natural phenomena inducing soil degradation, several contaminants, such as heavy metals, pesticides, and hydrocarbons in petroleum products, can have detrimental effects on soil quality and ecosystem health. Heavy metals, such as lead, arsenic, and cadmium, can accumulate in soil and affect plant growth and microbial activity, leading to reduced soil fertility and crop yield [90]. Similarly, polycyclic aromatic hydrocarbons (PAHs) from petroleum products can contaminate soil and cause toxic effects on soil organisms, impairing their ability to decompose organic matter and carry out nutrient cycling [91,92]. Pesticides can also have toxic effects on soil biota, including beneficial microbes and insects, leading to reduced soil biodiversity and enzymatic activity. This can result in biological and physicochemical transformations that negatively impact ecosystem function and crop productivity [93]. These contaminants can persist in the soil for long periods of time, not only affecting the immediate ecosystem but also potentially contaminating groundwater and nearby water bodies. Bioremediation with PGPB and microalgae has also been explored for the removal of these contaminants, with promising results. Microalgae and bacteria can play a crucial role in reducing contaminants from soils by breaking down pollutants and transforming them into less harmful forms [21,22]. This process helps to reduce the overall toxicity of the soil and prevents further contamination.
PGPB in phytoremediation have shown great potential by enhancing plant growth and biomass, especially under stressed conditions, resulting in faster and more efficient processes [94]. For example, Silambarasan et al. [95] reported an enhancement in the phytoremediation efficiency of Helianthus annus with the inoculation of Pseudomonas citronellolis strain SLP6. The Cu- and salinity-tolerant bacterial strains efficiently assisted in the phytostabilization of Cu-contaminated saline soils by enhancing plant growth and the Cu accumulation potential. Similarly, Rajkumar and Freitas [96] evaluated the potential of the PGPB Pseudomonas spp. and Pseudomonas jessenii as growth-promoting bioinoculants for plants and metal sequestration in soil contaminated with Ni, Cu, and Zn. The inoculation of Ricinus communis with PGPB increased the efficiency of phytoextraction by increasing metal solubilization in contaminated soil. Additionally, the biosorption and bioaccumulation capacities of the PGPB reduced the phytotoxic effects of the metals. In another study, the PGPB Bacillus altitudinis strain KP-14 successfully assisted in the phytostabilization potential of Miscanthus x giganteus in aged soil contaminated with several trace elements [97]. Similarly, Franchi et al. [98] reported that the combined effect of thiosulfate and a bacterial consortium increased the phytoaccumulation efficacy of Brassica juncea by up to 85% for arsenic and 45% for mercury in contaminated soil from a disused industrial area in northern Italy.
The use of PGPB in assisting in the bioremediation of pesticide residues in agricultural soils has also been explored, with promising results [99]. For instance, the potential of Bacillus aryabhattai to mitigate paraquat residues in drought soil was evaluated in pot experiments with cowpea seeds. The authors reported that the strain remediated the paraquat-contaminated soil and significantly improved the growth of cowpea compared to soil without bacterial inoculation [100]. Another study revealed that several species of the PGPB genus Bacillus were able to degrade pesticides, including acibenzolar-S-methyl, metribuzin, napropamide, propamocarb hydrochloride, and thiamethoxam, within a 72 h incubation period [101]. Similarly, the PGPB Pseudomonas rhizophila S211 was identified as a promising strain for the bioremediation of soils contaminated with pentachlorophenol [102], while the bacterial strains Acinetobacter calcoaceticus MCm5, Brevibacillus parabrevis FCm9, and Sphingomonas spp. were highly efficient in degrading cypermethrin and other pyrethroids, such as RCm6. These strains also exhibited plant-growth-promoting traits, such as phosphate solubilization and indole acetic acid and ammonia production [103].
Environmental pollution from petroleum spills is another problematic issue when restoring soil quality, and PGPB has exhibited great potential in their degradation. For example, the inoculation of PGPB increased the phytoremediation potential of Scirpus triqueter growing in a co-contaminated soil with Ni and pyrene, increasing the bioavailability of Ni and promoting the degradation of pyrene [104]. Similarly, Sampaio et al. [105] evaluated the biodegradation of PAHs in a diesel oil-contaminated mangrove at the Paraguassu River in Brazil. The authors monitored the ability of Rhizophora mangle L. to degrade PAHs in contaminated soil inoculated with PGPB Bacillus spp. and Pseudomonas aeruginosa and reported a removal rate of up to 80%. The interaction between R. mangle and the bacterial strains revealed the potential of the bacteria to assist in the phytoremediation of soils contaminated with diesel oil.
Furthermore, numerous studies have shown the potential of microalgae to reduce contaminant levels in soil. For example, Iliev et al. [106] evaluated the seed germination and plant growth of Tribulus terrestris following inoculation with a mixed algal suspension (mainly Scenedesmus spp. and Nostoc spp.) in soil contaminated with a mineral oil spill. The authors found a similar percentage of germination of seeds grown in non-polluted conditions and those grown in contaminated soil after watering with the mixed algal suspension, which suggests an efficient removal of the pollutant. Similarly, two species of green algae and five cyanobacteria were evaluated as inoculants of Oryza sativa in arsenic (As)-contaminated paddy soils. The inoculation, especially of the cyanobacteria Anabaena azotica, improved soil nutrient bioavailability, greatly enhanced rice growth, and reduced As translocation from roots to rice grains [107]. The removal of up to 78% and 48% of fluoranthene and pyrene, respectively, in contaminated soil was reported by Lei et al. [108] with the inoculation of four different microalgae species (Chlorella vulgaris, Scenedesmus platydiscus, Scenedesmus quadricauda, and Selenastrum capricornutum). The authors found that the removal rate was species-specific and toxicant-dependent. In another study, Decesaro et al. [109] compared the degradation potential of diesel and biodiesel using phycocyanin from Spirulina platensis, the inactive biomass of S. platensis, and ammonium sulfate. Maximal degradation in biodiesel-contaminated soil was achieved with the addition of phycocyanin (88.75%) and in diesel-contaminated soil with the addition of inactive S. platensis (63.89%), confirming the advantages of using natural compounds over chemicals. Similarly, a few studies have reported the successful degradation of pesticides, such as fenanmiphos, tricyclazole, and R-endosulfan, by some species of green algae and cyanobacteria [110,111,112].
Despite the proven potential of microalgae and bacteria for the bioremediation of contaminated soils, few studies have explored their use as combined consortia. One of these studies reported that the biological degradation of high-molecular-weight PAHs was successfully achieved on long-term contaminated soil with the inoculation of the microbial consortium co-formed by the microalga Chlorella spp. and the bacterium Rhodococcus wratislaviensis. The consortium efficiently degraded phenanthrene, pyrene, and benzo[a]pyrene (BaP) to below detection levels in soil slurry within 30 days [113]. The synergy between microalgae and bacteria produces a combined effect greater than that produced by each microorganism alone. Luo et al. [114] demonstrated this when evaluating the potential of a microalga–bacterium consortium (Selenastrum capricornutum and Mycobacterium spp.) to degrade recalcitrant PAHs. In addition to a complete degradation of pyrene in 10 days, the authors reported that the bacterial degradation of pyrene mitigated its toxicity for microalgae, whose growth was substantially inhibited when growing alone. Additionally, microalgae and bacterial growth were mutually enhanced with the co-culture.
6. Concluding Remarks
The enormous area of degraded land worldwide demands sustainable solutions to climate change and food scarcity. However, despite the several advantages of using microbial consortia for soil restoration, there are few studies on microbial microalgae–bacteria consortia inoculants. Additionally, although the biodiversity of microalgae is huge, most studies include only filamentous cyanobacteria (mostly Anabaena spp.) and unicellular green microalgae (mostly Chlorella spp.). Furthermore, a better understanding of the synergistic interaction between cyanobacteria and bacteria is imperative for the success of restoration programs using these consortia.
Finally, most of the published information regarding the effect of microbial inoculants on soil fertility derives from studies focused on the fertilizer potential of the inoculants and their growth-promoting effect on plants; therefore, more information regarding the actual impact on degraded soils is required.
Author Contributions
L.M.G.-G. and L.E.d.-B. participated in equal parts in conceptualizing, writing, and editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically Rethinking Agriculture for the 21st Century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, H.K.; Salmon, J.M. Mapping the world’s degraded lands. Appl. Geogr. 2015, 57, 12–21. [Google Scholar] [CrossRef]
- Shoshany, M.; Goldshleger, N.; Chudnovsky, A. Monitoring of agricultural soil degradation by remote-sensing methods: A review. Int. J. Remote Sens. 2013, 34, 6152–6181. [Google Scholar] [CrossRef]
- Ye, S.; Zeng, G.; Wu, H.; Zhang, C.; Liang, J.; Dai, J.; Liu, Z.; Xiong, W.; Wan, J.; Xu, P. Co-occurrence and interactions of pollutants, and their impacts on soil remediation—A review. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1528–1553. [Google Scholar] [CrossRef]
- Ossai, I.C.; Ahmed, A.; Hassan, A.; Hamid, F.S. Remediation of soil and water contaminated with petroleum hydrocarbon: A review. Environ. Technol. Innov. 2020, 17, 100526. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; Machado de Lima, N.M.; Chamizo, S.; Bowker, M.A. Restoring post-fire ecosystems with biocrusts: Living, photosynthetic soil surfaces. Curr. Opin. Environ. Sci. Health 2021, 23, 100273. [Google Scholar] [CrossRef]
- Abhilash, P.C. Restoring the Unrestored: Strategies for Restoring Global Land during the UN Decade on Ecosystem Restoration (UN-DER). Land 2021, 10, 201. [Google Scholar] [CrossRef]
- Bruckman, V.J.; Pumpanen, J. Biochar use in global forests: Opportunities and challenges. Dev. Soil Sci. 2019, 36, 427–453. [Google Scholar]
- Abinandan, S.; Subashchandrabose, S.R.; Venkateswarlu, K.; Megharaj, M. Soil microalgae and cyanobacteria: The biotechnological potential in the maintenance of soil fertility and health. Crit. Rev. Biotechnol. 2019, 39, 981–998. [Google Scholar] [CrossRef]
- Mutum, L.; Janda, T.; Ördög, V.; Molnár, Z. Biologia Futura: Potential of different forms of microalgae for soil improvement. Biol. Futur. 2022, 73, 1–8. [Google Scholar] [CrossRef]
- Ramakrishnan, B.; Maddela, N.R.; Venkateswarlu, K.; Megharaj, M. Potential of microalgae and cyanobacteria to improve soil health and agricultural productivity: A critical view. Environ.Sci. Adv. 2023, 2, 586–611. [Google Scholar] [CrossRef]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro- and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Pereg, L.; de-Bashan, L.E.; Bashan, Y. Assessment of affinity and specificity of Azospirillum for plants. Plant Soil 2016, 399, 389–414. [Google Scholar] [CrossRef]
- Galaviz, C.; Lopez, B.R.; de-Bashan, L.E.; Hirsch, A.M.; Maymon, M.; Bashan, Y. Root growth improvement of mesquite seedlings and bacterial rhizosphere and soil community changes are induced by inoculation with plant growth-promoting bacteria and promote restoration of eroded desert soil. Land Degrad. Dev. 2018, 29, 1453–1466. [Google Scholar] [CrossRef]
- Moreno, M.; de-Bashan, L.E.; Hernandez, J.-P.; Lopez, B.R.; Bashan, Y. Success of long-term restoration of degraded arid land using native trees planted 11 years earlier. Plant Soil 2017, 421, 83–92. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L. Plant growth-promoting. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elsevier: Oxford, UK, 2005; Volume 1, pp. 103–115. [Google Scholar]
- Bashan, Y.; Holguin, G. Azospirillum–plant relationships: Environmental and physiological advances (1990–1996). Can. J. Microbiol. 1997, 43, 103–121. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E. How the Plant Growth-Promoting Bacterium Azospirillum Promotes Plant Growth—A Critical Assessment. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2010; pp. 77–136. [Google Scholar] [CrossRef]
- Cassán, F.; Coniglio, A.; López, G.; Molina, R.; Nievas, S.; Carlan, C.L.N.D.; Donadio, F.; Torres, D.; Rosas, S.; Pedrosa, F.O.; et al. Everything you must know about Azospirillum and its impact on agriculture and beyond. Biol. Fertil. Soils 2020, 56, 461–479. [Google Scholar] [CrossRef]
- Tarfeen, N.; Nisa, K.U.; Hamid, B.; Bashir, Z.; Yatoo, A.M.; Dar, M.A.; Mohiddin, F.A.; Amin, Z.; Ahmad, R.A.; Sayyed, R.Z. Microbial Remediation: A Promising Tool for Reclamation of Contaminated Sites with Special Emphasis on Heavy Metal and Pesticide Pollution: A Review. Processes 2022, 10, 1358. [Google Scholar] [CrossRef]
- Naeem, U.; Qazi, M.A. Leading edges in bioremediation technologies for removal of petroleum hydrocarbons. Environ. Sci. Pollut. Res. 2020, 27, 27370–27382. [Google Scholar] [CrossRef]
- Zhang, B.; Li, W.; Guo, Y.; Zhang, Z.; Shi, W.; Cui, F.; Lens, P.N.; Tay, J.H. Microalgal-bacterial consortia: From interspecies interactions to biotechnological applications. Renew. Sustain. Energy Rev. 2020, 118, 109563. [Google Scholar] [CrossRef]
- González-González, L.M.; de-Bashan, L.E. Toward the Enhancement of Microalgal Metabolite Production through Microalgae–Bacteria Consortia. Biology 2021, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Smiraglia, D.; Ceccarelli, T.; Bajocco, S.; Salvati, L.; Perini, L. Linking trajectories of land change, land degradation processes and ecosystem services. Environ. Res. 2016, 147, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Prince, S.; Von Maltitz, G.; Zhang, F.; Byrne, K.; Driscoll, C.; Eshel, G.; Kust, G.; Martínez-Garza, C.; Metzger, J.; Midgley, G. Status and Trends of Land Degradation and Restoration and Associated Changes in Biodiversity and Ecosystem Functions; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES): Bonn, Germany, 2018; pp. 221–338.
- Jónsson, J.G.; Davíðsdóttir, B. Classification and valuation of soil ecosystem services. Agric. Syst. 2016, 145, 24–38. [Google Scholar] [CrossRef]
- Pereira, P.; Bogunovic, I.; Muñoz-Rojas, M.; Brevik, E.C. Soil ecosystem services, sustainability, valuation and management. Curr. Opin. Environ. Sci. Health 2018, 5, 7–13. [Google Scholar] [CrossRef]
- Lehmann, J.; Bossio, D.A.; Kögel-Knabner, I.; Rillig, M.C. The concept and future prospects of soil health. Nat. Rev. Earth Environ. 2020, 1, 544–553. [Google Scholar] [CrossRef]
- Keesstra, S.; Mol, G.; de Leeuw, J.; Okx, J.; Molenaar, C.; de Cleen, M.; Visser, S. Soil-Related Sustainable Development Goals: Four Concepts to Make Land Degradation Neutrality and Restoration Work. Land 2018, 7, 133. [Google Scholar] [CrossRef]
- Sutton, P.C.; Anderson, S.J.; Costanza, R.; Kubiszewski, I. The ecological economics of land degradation: Impacts on ecosystem service values. Ecol. Econ. 2016, 129, 182–192. [Google Scholar] [CrossRef]
- Nkonya, E.; Anderson, W.; Kato, E.; Koo, J.; Mirzabaev, A.; von Braun, J.; Meyer, S. Global cost of land degradation. In Economics of Land Degradation and Improvement–A Global Assessment for Sustainable Development; Springer: Bonn, Germany, 2016; pp. 117–165. [Google Scholar]
- Qiu, L.; Zhang, Q.; Zhu, H.; Reich, P.B.; Banerjee, S.; van der Heijden, M.G.; Sadowsky, M.J.; Ishii, S.; Jia, X.; Shao, M. Erosion reduces soil microbial diversity, network complexity and multifunctionality. ISME J. 2021, 15, 2474–2489. [Google Scholar] [CrossRef]
- Mabuhay, J.; Nakagoshi, N.; Isagi, Y. Influence of erosion on soil microbial biomass, abundance and community diversity. Land Degrad. Dev. 2004, 15, 183–195. [Google Scholar] [CrossRef]
- Dasgupta, D.; Brahmaprakash, G.P. Soil Microbes are Shaped by Soil Physico-chemical Properties: A Brief Review of Existing Literature. Int. J. Plant Soil Sci. 2021, 33, 59–71. [Google Scholar] [CrossRef]
- Wagg, C.; Hautier, Y.; Pellkofer, S.; Banerjee, S.; Schmid, B.; van der Heijden, M.G. Diversity and asynchrony in soil microbial communities stabilizes ecosystem functioning. eLife 2021, 10, e62813. [Google Scholar] [CrossRef]
- Maron, P.-A.; Sarr, A.; Kaisermann, A.; Lévêque, J.; Mathieu, O.; Guigue, J.; Karimi, B.; Bernard, L.; Dequiedt, S.; Terrat, S.; et al. High Microbial Diversity Promotes Soil Ecosystem Functioning. Appl. Environ. Microbiol. 2018, 84, e02738-17. [Google Scholar] [CrossRef]
- Hartmann, M.; Six, J. Soil structure and microbiome functions in agroecosystems. Nat. Rev. Earth Environ. 2022, 4, 4–18. [Google Scholar] [CrossRef]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Talukder, B.; Ganguli, N.; Matthew, R.; vanLoon, G.W.; Hipel, K.W.; Orbinski, J. Climate change-triggered land degradation and planetary health: A review. Land Degrad. Dev. 2021, 32, 4509–4522. [Google Scholar] [CrossRef]
- Webb, N.P.; Marshall, N.A.; Stringer, L.C.; Reed, M.S.; Chappell, A.; Herrick, J.E. Land degradation and climate change: Building climate resilience in agriculture. Front. Ecol. Environ. 2017, 15, 450–459. [Google Scholar] [CrossRef]
- Schiedek, D.; Sundelin, B.; Readman, J.W.; Macdonald, R.W. Interactions between climate change and contaminants. Mar. Pollut. Bull. 2007, 54, 1845–1856. [Google Scholar] [CrossRef]
- Noyes, P.D.; McElwee, M.; Miller, H.D.; Clark, B.W.; Van Tiem, L.A.; Walcott, K.C.; Erwin, K.N.; Levin, E.D. The toxicology of climate change: Environmental contaminants in a warming world. Environ. Int. 2009, 35, 971–986. [Google Scholar] [CrossRef]
- Coban, O.; De Deyn, G.B.; van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, abe0725. [Google Scholar] [CrossRef]
- Prăvălie, R. Exploring the multiple land degradation pathways across the planet. Earth-Sci. Rev. 2021, 220, 103689. [Google Scholar] [CrossRef]
- Rossi, F.; Olguin, E.J.; Diels, L.; De Philippis, R. Microbial fixation of CO2 in water bodies and in drylands to combat climate change, soil loss and desertification. Nat. Biotechnol. 2015, 32, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Prăvălie, R.; Patriche, C.; Borrelli, P.; Panagos, P.; Rosca, B.; Dumitrascu, M.; Nita, I.A.; Savulescu, I.; Birsan, M.V.; Bandoc, G. Arable lands under the pressure of multiple land degradation processes. A global perspective. Environ. Res. 2021, 194, 110697. [Google Scholar] [CrossRef] [PubMed]
- FAO. The Future of Food and Agriculture: Alternative Pathways to 2050; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018.
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Bashan, Y.; Salazar, B.G.; Moreno, M.; Lopez, B.R.; Linderman, R.G. Restoration of eroded soil in the Sonoran Desert with native leguminous trees using plant growth-promoting microorganisms and limited amounts of compost and water. J. Environ. Manag. 2012, 102, 26–36. [Google Scholar] [CrossRef]
- Gonzalez, E.J.; Hernandez, J.-P.; De-Bashan, L.E.; Bashan, Y. Dry micro-polymeric inoculant of Azospirillum brasilense is useful for producing mesquite transplants for reforestation of degraded arid zones. Appl. Soil Ecol. 2018, 129, 84–93. [Google Scholar] [CrossRef]
- Ramachandran, A.; Radhapriya, P. Restoration of Degraded Soil in the Nanmangalam Reserve Forest with Native Tree Species: Effect of Indigenous Plant Growth-Promoting Bacteria. Sci. World J. 2016, 2016, 5465841. [Google Scholar] [CrossRef]
- Schoebitz, M.; Mengual, C.; Roldán, A. Combined effects of clay immobilized Azospirillum brasilense and Pantoea dispersa and organic olive residue on plant performance and soil properties in the revegetation of a semiarid area. Sci. Total Environ. 2014, 466–467, 67–73. [Google Scholar] [CrossRef]
- Nouri, H.; Borujeni, S.C.; Nirola, R.; Hassanli, A.; Beecham, S.; Alaghmand, S.; Saint, C.; Mulcahy, D. Application of green remediation on soil salinity treatment: A review on halophytoremediation. Process. Saf. Environ. Prot. 2017, 107, 94–107. [Google Scholar] [CrossRef]
- Shilev, S. Plant-Growth-Promoting Bacteria Mitigating Soil Salinity Stress in Plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef]
- Mishra, P.; Mishra, J.; Arora, N.K. Plant growth promoting bacteria for combating salinity stress in plants—Recent developments and prospects: A review. Microbiol. Res. 2021, 252, 126861. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-tolerant plant growth-promoting Bacillus pumilus strain JPVS11 to enhance plant growth attributes of rice and improve soil health under salinity stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef]
- Hafez, M.; El-Ezz, S.F.A.; Popov, A.I.; Rashad, M. Organic Amendments Combined with Plant Growth-Promoting Rhizobacteria (Azospirillum brasilense) as an Eco-Friendly By-Product to Remediate and Enhance the Fertility of Saline Sodic-Soils in Egypt. Commun. Soil Sci. Plant Anal. 2021, 52, 1416–1433. [Google Scholar] [CrossRef]
- Lababpour, A. Potentials of the microalgae inoculant in restoration of biological soil crusts to combat desertification. Int. J. Environ. Sci. Technol. 2016, 13, 2521–2532. [Google Scholar] [CrossRef]
- Perera, I.; Subashchandrabose, S.R.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Consortia of cyanobacteria/microalgae and bacteria in desert soils: An underexplored microbiota. Appl. Microbiol. Biotechnol. 2019, 102, 7351–7363. [Google Scholar] [CrossRef]
- Barone, V.; Puglisi, I.; Fragalà, F.; Stevanato, P.; Baglieri, A. Effect of living cells of microalgae or their extracts on soil enzyme activities. Arch. Agron. Soil Sci. 2018, 65, 712–726. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Li, D.; Hu, C.; Rao, B. Feasibility of cyanobacterial inoculation for biological soil crusts formation in desert area. Soil Biol. Biochem. 2009, 41, 926–929. [Google Scholar] [CrossRef]
- Lan, S.; Zhang, Q.; Wu, L.; Liu, Y.; Zhang, D.; Hu, C. Artificially Accelerating the Reversal of Desertification: Cyanobacterial Inoculation Facilitates the Succession of Vegetation Communities. Environ. Sci. Technol. 2014, 48, 307–315. [Google Scholar] [CrossRef]
- Nisha, R.; Kaushik, A.; Kaushik, C. Effect of indigenous cyanobacterial application on structural stability and productivity of an organically poor semi-arid soil. Geoderma 2007, 138, 49–56. [Google Scholar] [CrossRef]
- Hu, C.; Liu, Y.; Song, L.; Zhang, D. Effect of desert soil algae on the stabilization of fine sands. J. Appl. Phycol. 2002, 14, 281–292. [Google Scholar] [CrossRef]
- Malam Issa, O.; Défarge, C.; Le Bissonnais, Y.; Marin, B.; Duval, O.; Bruand, A.; D’Acqui, L.P.; Nordenberg, S.; Annerman, M. Effects of the inoculation of cyanobacteria on the microstructure and the structural stability of a tropical soil. Plant Soil 2007, 290, 209–219. [Google Scholar] [CrossRef]
- Shanthakumar, S.; Abinandan, S.; Venkateswarlu, K.; Subashchandrabose, S.R.; Megharaj, M. Algalization of acid soils with acid-tolerant strains: Improvement in pH, carbon content, exopolysaccharides, indole acetic acid and dehydrogenase activity. Land Degrad. Dev. 2021, 32, 3157–3166. [Google Scholar] [CrossRef]
- Abinandan, S.; Shanthakumar, S.; Panneerselvan, L.; Venkateswarlu, K.; Megharaj, M. Algalization of acid soils with Desmodesmus sp. MAS1 and Heterochlorella sp. MAS3 enriches bacteria of ecological importance. ACS Agric. Sci. Technol. 2022, 2, 512–520. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; Román, J.; Roncero-Ramos, B.; Erickson, T.; Merritt, D.; Aguila-Carricondo, P.; Cantón, Y. Cyanobacteria inoculation enhances carbon sequestration in soil substrates used in dryland restoration. Sci. Total Environ. 2018, 636, 1149–1154. [Google Scholar] [CrossRef]
- Subashchandrabose, S.R.; Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Consortia of cyanobacteria/microalgae and bacteria: Biotechnological potential. Biotechnol. Adv. 2011, 29, 896–907. [Google Scholar] [CrossRef]
- Palacios, O.A.; López, B.R.; de-Bashan, L.E. Microalga Growth-Promoting Bacteria (MGPB): A formal term proposed for beneficial bacteria involved in microalgal–bacterial interactions. Algal Res. 2022, 61, 102585. [Google Scholar] [CrossRef]
- Palacios, O.A.; Lopez, B.R.; Bashan, Y.; de-Bashan, L.E. Early Changes in Nutritional Conditions Affect Formation of Synthetic Mutualism Between Chlorella sorokiniana and the Bacterium Azospirillum brasilense. Microb. Ecol. 2019, 77, 980–992. [Google Scholar] [CrossRef]
- Palacios, O.A.; Espinoza-Hicks, J.C.; Camacho-Davila, A.A.; Lopez, B.R.; de-Bashan, L.E. Differences in exudates between strains of Chlorella sorokiniana affect the interaction with the microalga growth-promoting bacteria Azospirillum brasilense. Microbiol. Ecol. 2022, 85, 1412–1422. [Google Scholar] [CrossRef]
- Palacios, O.A.; López, B.R.; Palacios-Espinosa, A.; Hernández-Sandoval, F.E.; de-Bashan, L.E. The immediate effect of riboflavin and lumichrome on the mitigation of saline stress in the microalga Chlorella sorokiniana by the plant-growth-promoting bacterium Azospirillum brasilense. Algal Res. 2021, 58, 102424. [Google Scholar] [CrossRef]
- Dao, G.; Wang, S.; Wang, X.; Chen, Z.; Wu, Y.; Wu, G.; Lu, Y.; Liu, S.; Hu, H. Enhanced Scenedesmus sp. growth in response to gibberellin secretion by symbiotic bacteria. Sci. Total Environ. 2020, 740, 140099. [Google Scholar] [CrossRef]
- Pagnussat, L.; Maroniche, G.; Curatti, L.; Creus, C. Auxin-dependent alleviation of oxidative stress and growth promotion of Scenedesmus obliquus C1S by Azospirillum brasilense. Algal Res. 2020, 47, 101839. [Google Scholar] [CrossRef]
- Peng, H.; de-Bashan, L.E.; Bashan, Y.; Higgins, B.T. Indole-3-acetic acid from Azosprillum brasilense promotes growth in green algae at the expense of energy storage products. Algal Res. 2020, 47, 101845. [Google Scholar] [CrossRef]
- Zayadan, B.K.; Matorin, D.N.; Baimakhanova, G.B.; Bolathan, K.; Oraz, G.D.; Sadanov, A.K. Promising microbial consortia for producing biofertilizers for rice fields. Microbiology 2014, 83, 391–397. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Prasanna, R.; Kumar, A.; Pattnaik, S.; Chakravarty, K.; Shivay, Y.S.; Singh, R.; Saxena, A.K. Evaluating the influence of novel cyanobacterial biofilmed biofertilizers on soil fertility and plant nutrition in wheat. Eur. J. Soil Biol. 2013, 55, 107–116. [Google Scholar] [CrossRef]
- Prasanna, R.; Triveni, S.; Bidyarani, N.; Babu, S.; Yadav, K.; Adak, A.; Khetarpal, S.; Pal, M.; Shivay, Y.S.; Saxena, A.K. Evaluating the efficacy of cyanobacterial formulations and biofilmed inoculants for leguminous crops. Arch. Agron. Soil Sci. 2014, 60, 349–366. [Google Scholar] [CrossRef]
- Manjunath, M.; Prasanna, R.; Sharma, P.; Nain, L.; Singh, R. Developing PGPR consortia using novel genera Providencia and Alcaligenes along with cyanobacteria for wheat. Arch. Agron. Soil Sci. 2011, 57, 873–887. [Google Scholar] [CrossRef]
- Bashan, Y.; Trejo, A.; de-Bashan, L.E. Development of two culture media for mass cultivation of Azospirillum spp. and for production of inoculants to enhance plant growth. Biol. Fertil. Soils 2011, 47, 963–969. [Google Scholar] [CrossRef]
- Lopez, B.R.; Bashan, Y.; Trejo, A.; de-Bashan, L.E. Amendment of degraded desert soil with wastewater debris containing immobilized Chlorella sorokiniana and Azospirillum brasilense significantly modifies soil bacterial community structure, diversity, and richness. Biol. Fertil. Soils 2013, 49, 1053–1063. [Google Scholar] [CrossRef]
- Kheirfam, H.; Sadeghi, S.H.; Darki, B.Z.; Homaee, M. Controlling rainfall-induced soil loss from small experimental plots through inoculation of bacteria and cyanobacteria. Catena 2017, 152, 40–46. [Google Scholar] [CrossRef]
- Rana, A.; Joshi, M.; Prasanna, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Babu, S.; Prasanna, R.; Bidyarani, N.; Nain, L.; Shivay, Y.S. Synergistic action of PGP agents and Rhizobium spp. for improved plant growth, nutrient mobilization and yields in different leguminous crops. Biocatal. Agric. Biotechnol. 2015, 4, 456–464. [Google Scholar] [CrossRef]
- Bidyarani, N.; Prasanna, R.; Babu, S.; Hossain, F.; Saxena, A.K. Enhancement of plant growth and yields in Chickpea (Cicer arietinum L.) through novel cyanobacterial and biofilmed inoculants. Microbiol. Res. 2016, 188–189, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Trejo, A.; de-Bashan, L.E.; Hartmann, A.; Hernandez, J.-P.; Rothballer, M.; Schmid, M.; Bashan, Y. Recycling waste debris of immobilized microalgae and plant growth-promoting bacteria from wastewater treatment as a resource to improve fertility of eroded desert soil. Environ. Exp. Bot. 2012, 75, 65–73. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental chemistry and ecotoxicology of hazardous heavy metals: Environmental persistence, toxicity, and bioaccumulation. J. Chem. 2019, 2019, 6730305. [Google Scholar] [CrossRef]
- Ahmed, F.; Fakhruddin, A. A review on environmental contamination of petroleum hydrocarbons and its biodegradation. Int. J. Environ. Sci. Nat. Resour. 2018, 11, 1–7. [Google Scholar]
- Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Ball, A.S.; Shahsavari, E. Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems—Fate and Microbial Responses. Molecules 2019, 24, 3400. [Google Scholar] [CrossRef]
- Wołejko, E.; Jabłońska-Trypuć, A.; Wydro, U.; Butarewicz, A.; Łozowicka, B. Soil biological activity as an indicator of soil pollution with pesticides—A review. Appl. Soil Ecol. 2020, 147, 103356. [Google Scholar] [CrossRef]
- de-Bashan, L.E.; Hernandez, J.-P.; Bashan, Y. The potential contribution of plant growth-promoting bacteria to reduce environmental degradation—A comprehensive evaluation. Appl. Soil Ecol. 2012, 61, 171–189. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Valentine, A.; Cornejo, P.; Kannan, V.R. Pseudomonas citronellolis strain SLP6 enhances the phytoremediation efficiency of Helianthus annuus in copper contaminated soils under salinity stress. Plant Soil 2020, 457, 241–253. [Google Scholar] [CrossRef]
- Rajkumar, M.; Freitas, H. Influence of metal resistant-plant growth-promoting bacteria on the growth of Ricinus communis in soil contaminated with heavy metals. Chemosphere 2008, 71, 834–842. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.; Mamirova, A.; Pranaw, K.; Shapoval, P.Y.; Trögl, J.; Nurzhanova, A. Potential role of plant growth-promoting bacteria in Miscanthus x giganteus phytotechnology applied to the trace elements contaminated soils. Int. Biodeterior. Biodegrad. 2020, 155, 105103. [Google Scholar] [CrossRef]
- Franchi, E.; Rolli, E.; Marasco, R.; Agazzi, G.; Borin, S.; Cosmina, P.; Pedron, F.; Rosellini, I.; Barbafieri, M.; Petruzzelli, G. Phytoremediation of a multi contaminated soil: Mercury and arsenic phytoextraction assisted by mobilizing agent and plant growth promoting bacteria. J. Soils Sediments 2016, 17, 1224–1236. [Google Scholar] [CrossRef]
- Rani, R.; Kumar, V.; Gupta, P.; Chandra, A. Application of plant growth promoting rhizobacteria in remediation of pesticides contaminated stressed soil. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 341–353. [Google Scholar] [CrossRef]
- Inthama, P.; Pumas, P.; Pekkoh, J.; Pathom-Aree, W.; Pumas, C. Plant Growth and Drought Tolerance-Promoting Bacterium for Bioremediation of Paraquat Pesticide Residues in Agriculture Soils. Front. Microbiol. 2021, 12, 604662. [Google Scholar] [CrossRef]
- Myresiotis, C.K.; Vryzas, Z.; Papadopoulou-Mourkidou, E. Biodegradation of soil-applied pesticides by selected strains of plant growth-promoting rhizobacteria (PGPR) and their effects on bacterial growth. Biodegradation 2012, 23, 297–310. [Google Scholar] [CrossRef]
- Hassen, W.; Neifar, M.; Cherif, H.; Najjari, A.; Chouchane, H.; Driouich, R.C.; Salah, A.; Naili, F.; Mosbah, A.; Souissi, Y. Pseudomonas rhizophila S211, a new plant growth-promoting rhizobacterium with potential in pesticide-bioremediation. Front. Microbiol. 2018, 9, 34. [Google Scholar] [CrossRef]
- Akbar, S.; Sultan, S.; Kertesz, M. Determination of Cypermethrin Degradation Potential of Soil Bacteria Along with Plant Growth-Promoting Characteristics. Curr. Microbiol. 2015, 70, 75–84. [Google Scholar] [CrossRef]
- Chen, X.; Liu, X.; Zhang, X.; Cao, L.; Hu, X. Phytoremediation effect of Scirpus triqueter inoculated plant-growth-promoting bacteria (PGPB) on different fractions of pyrene and Ni in co-contaminated soils. J. Hazard. Mater. 2017, 325, 319–326. [Google Scholar] [CrossRef]
- Sampaio, C.J.S.; de Souza, J.R.B.; Damião, A.O.; Bahiense, T.C.; Roque, M.R.A. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) in a diesel oil-contaminated mangrove by plant growth-promoting rhizobacteria. 3 Biotech 2019, 9, 155. [Google Scholar] [CrossRef]
- Iliev, I.; Petkov, G.; Lukavsky, J. An approach to bioremediation of mineral oil polluted soil. Genet. Plant Physiol. 2015, 5, 162–169. [Google Scholar]
- Wang, Y.; Li, Y.Q.; Lv, K.; Cheng, J.J.; Chen, X.L.; Ge, Y.; Yu, X.Y. Soil microalgae modulate grain arsenic accumulation by reducing dimethylarsinic acid and enhancing nutrient uptake in rice (Oryza sativa L.). Plant Soil 2018, 430, 99–111. [Google Scholar] [CrossRef]
- Lei, A.-P.; Hu, Z.-L.; Wong, Y.-S.; Tam, N.F.-Y. Removal of fluoranthene and pyrene by different microalgal species. Bioresour. Technol. 2007, 98, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Decesaro, A.; Rampel, A.; Machado, T.S.; Thomé, A.; Reddy, K.; Margarites, A.C.; Colla, L.M. Bioremediation of Soil Contaminated with Diesel and Biodiesel Fuel Using Biostimulation with Microalgae Biomass. J. Environ. Eng. 2017, 143, 04016091. [Google Scholar] [CrossRef]
- Cáceres, T.P.; Megharaj, M.; Naidu, R. Biodegradation of the Pesticide Fenamiphos by Ten Different Species of Green Algae and Cyanobacteria. Curr. Microbiol. 2008, 57, 643–646. [Google Scholar] [CrossRef]
- Sethunathan, N.; Megharaj, M.; Chen, Z.; Williams, B.D.; Lewis, G.; Naidu, R. Algal degradation of a known endocrine disrupting insecticide, α-endosulfan, and its metabolite, endosulfan sulfate, in liquid medium and soil. J. Agric. Food Chem. 2004, 52, 3030–3035. [Google Scholar] [CrossRef]
- Kumar, N.; Mukherjee, I.; Sarkar, B.; Paul, R.K. Degradation of tricyclazole: Effect of moisture, soil type, elevated carbon dioxide and Blue Green Algae (BGA). J. Hazard. Mater. 2017, 321, 517–527. [Google Scholar] [CrossRef]
- Subashchandrabose, S.R.; Venkateswarlu, K.; Venkidusamy, K.; Palanisami, T.; Naidu, R.; Megharaj, M. Bioremediation of soil long-term contaminated with PAHs by algal-bacterial synergy of Chlorella sp. MM3 and Rhodococcus wratislaviensis strain 9 in slurry phase. Sci. Total Environ. 2019, 659, 724–731. [Google Scholar] [CrossRef]
- Luo, S.; Chen, B.; Lin, L.; Wang, X.; Tam, N.F.-Y.; Luan, T. Pyrene Degradation Accelerated by Constructed Consortium of Bacterium and Microalga: Effects of Degradation Products on the Microalgal Growth. Environ. Sci. Technol. 2014, 48, 13917–13924. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).