



Phenotypic Plasticity Strategy of Aeluropus lagopoides Grass in Response to Heterogenous Saline Habitats

 ,
,  ,
,  ,
, 
 and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Surveyed Regions and Soil Analysis
2.2. Morphological Traits Measurements
2.3. Determination of Ecophysiological Traits
2.4. Statistical Analysis
3. Results
3.1. Soil Layer Variations among the Regions of A. lagopoides
3.2. Morphological Traits Variations among the Studied Regions of A. lagopoides
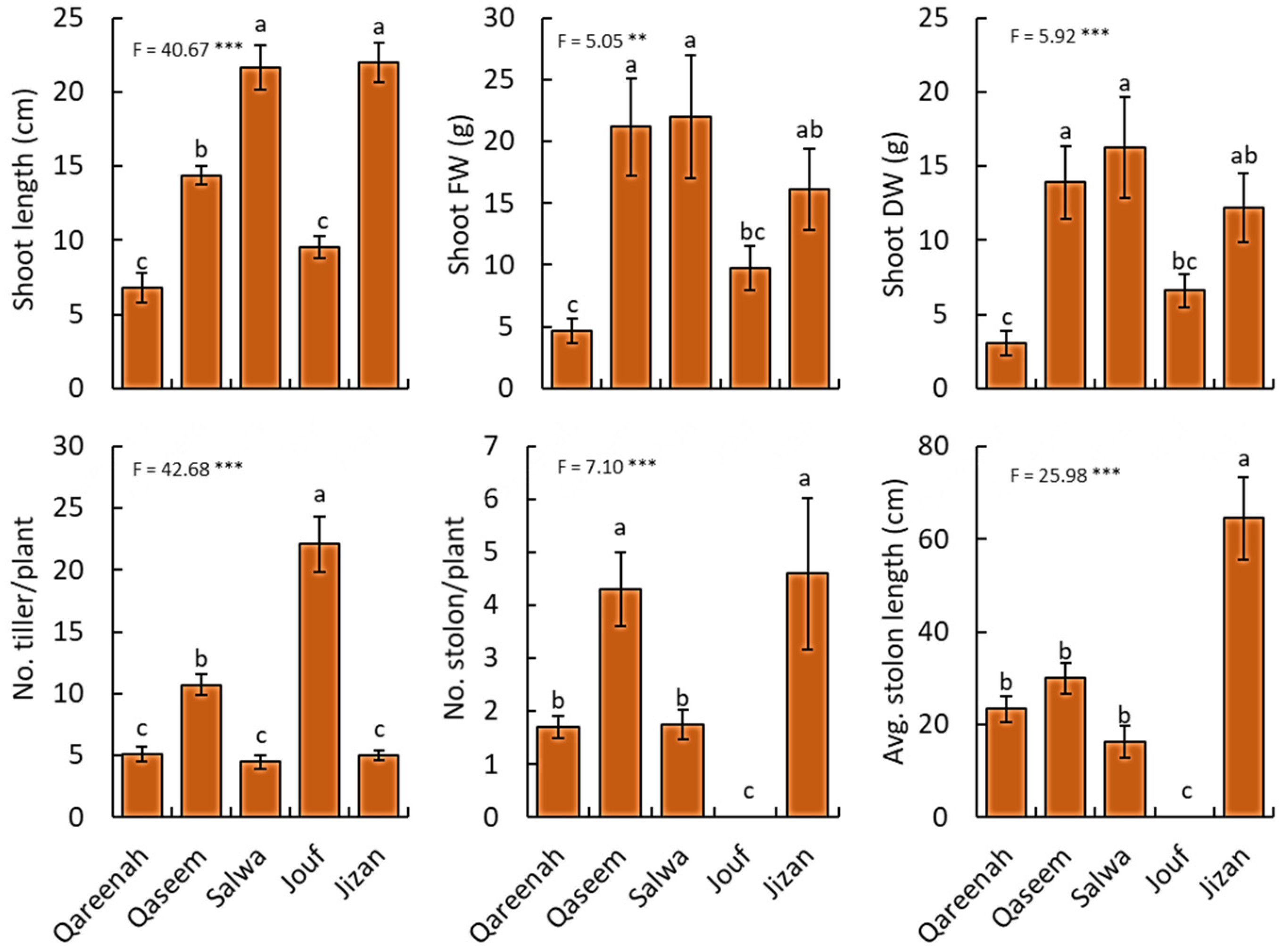
3.2.1. Shoot Traits
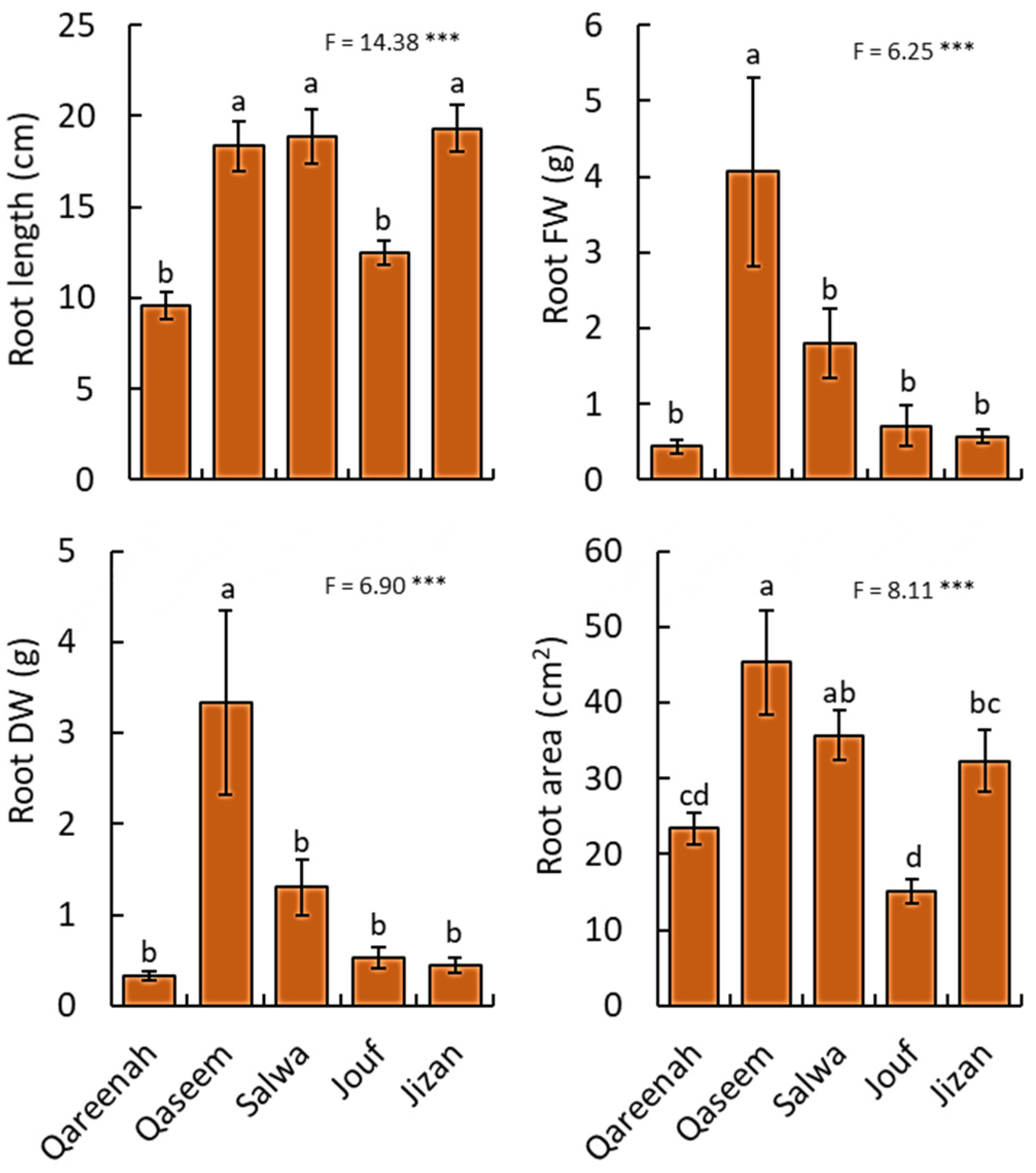
3.2.2. Root Traits
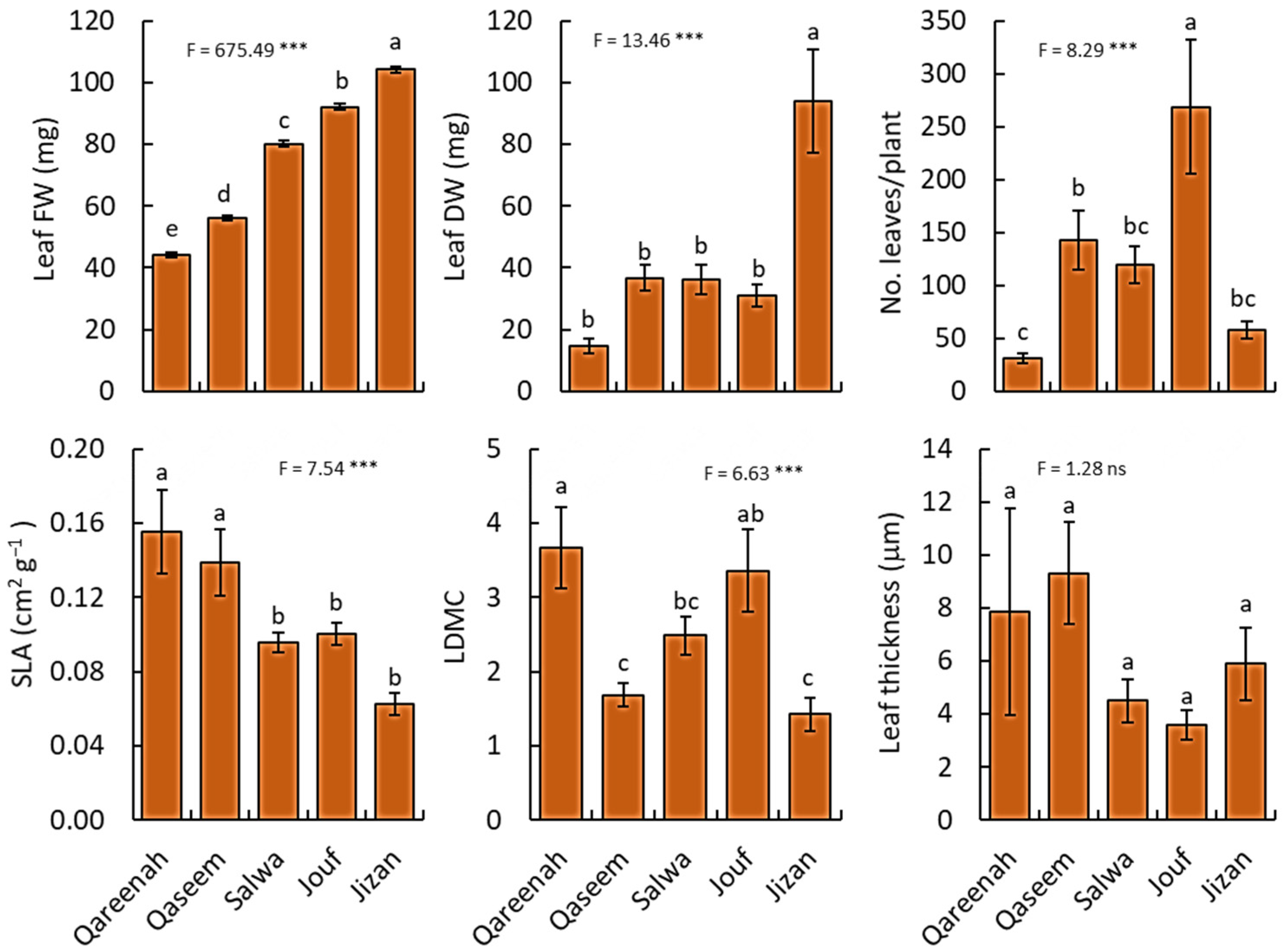
3.2.3. Leaf Traits
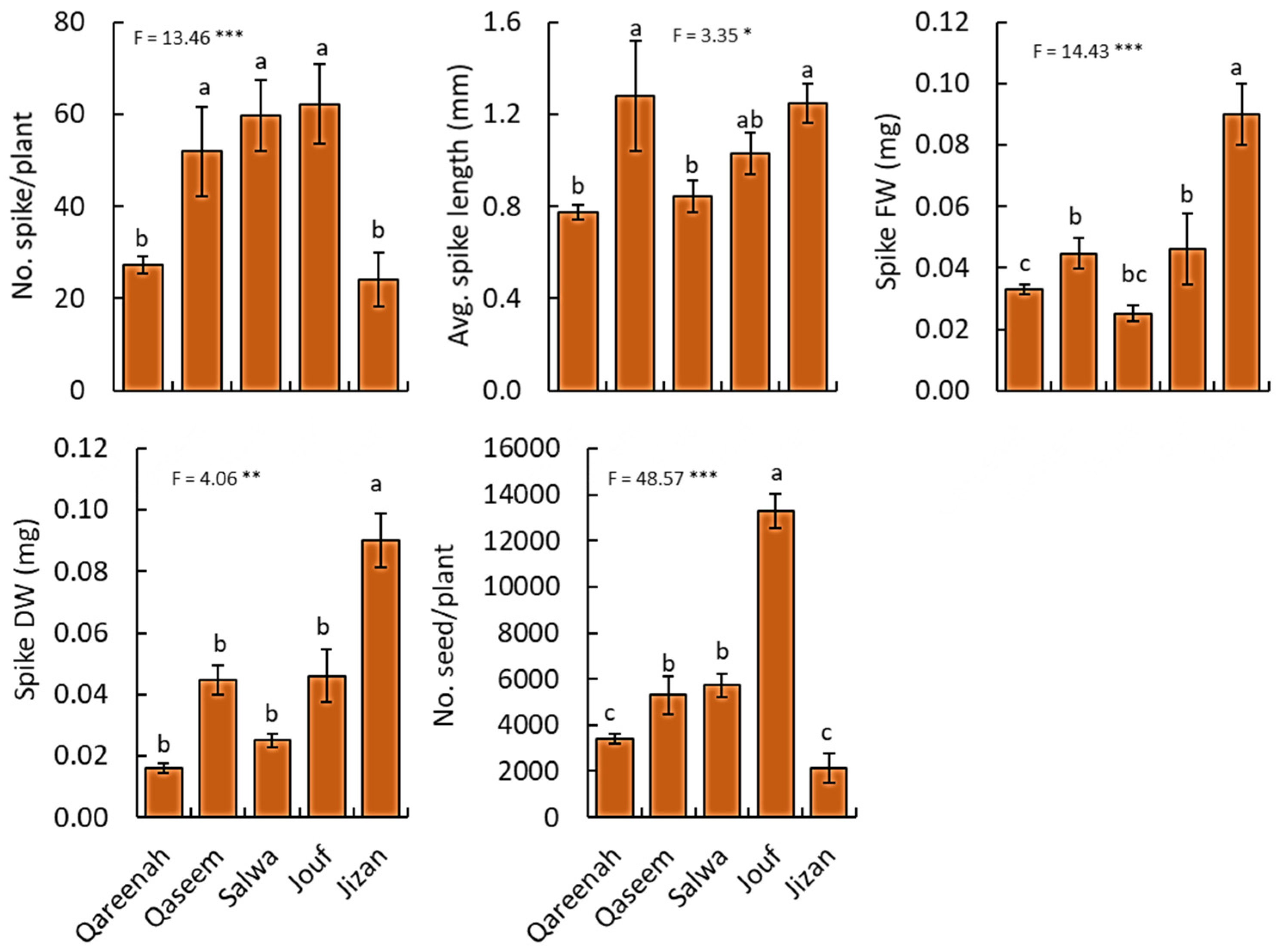
3.2.4. Reproductive Traits
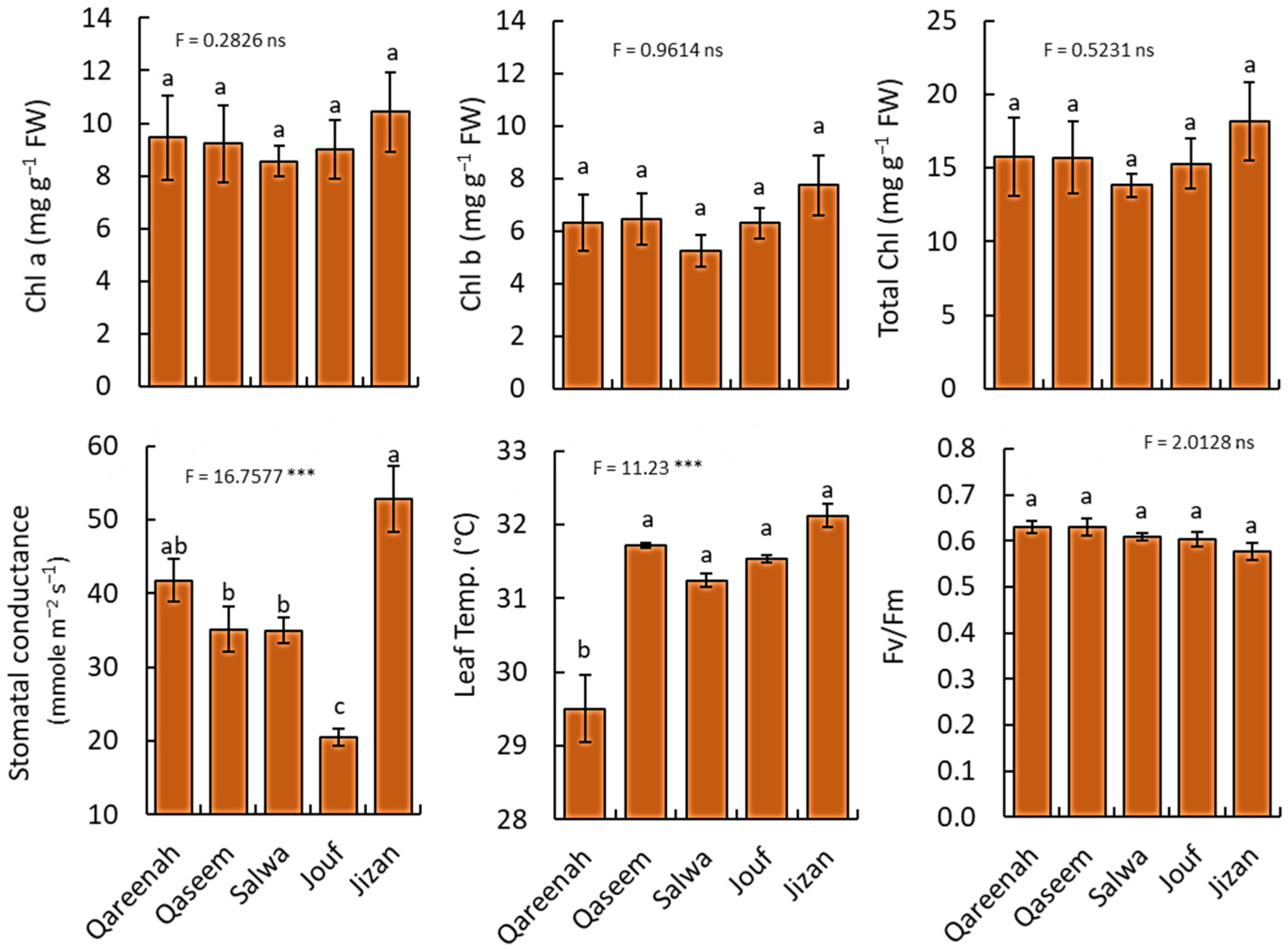
3.3. Variation in Ecophysiological Parameters of A. lagopoides among Different Regions
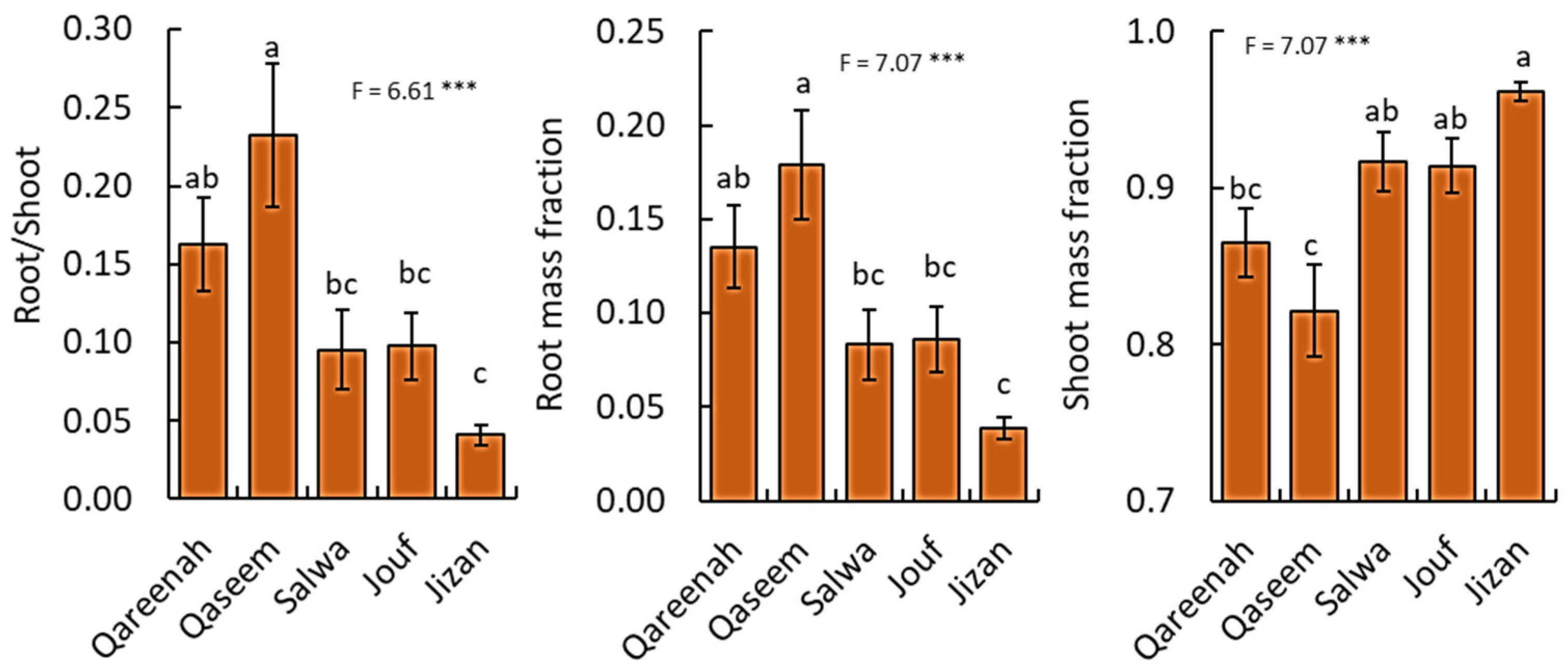
3.4. Biomass Allocation of A. lagopoides in Response to Different Habitats
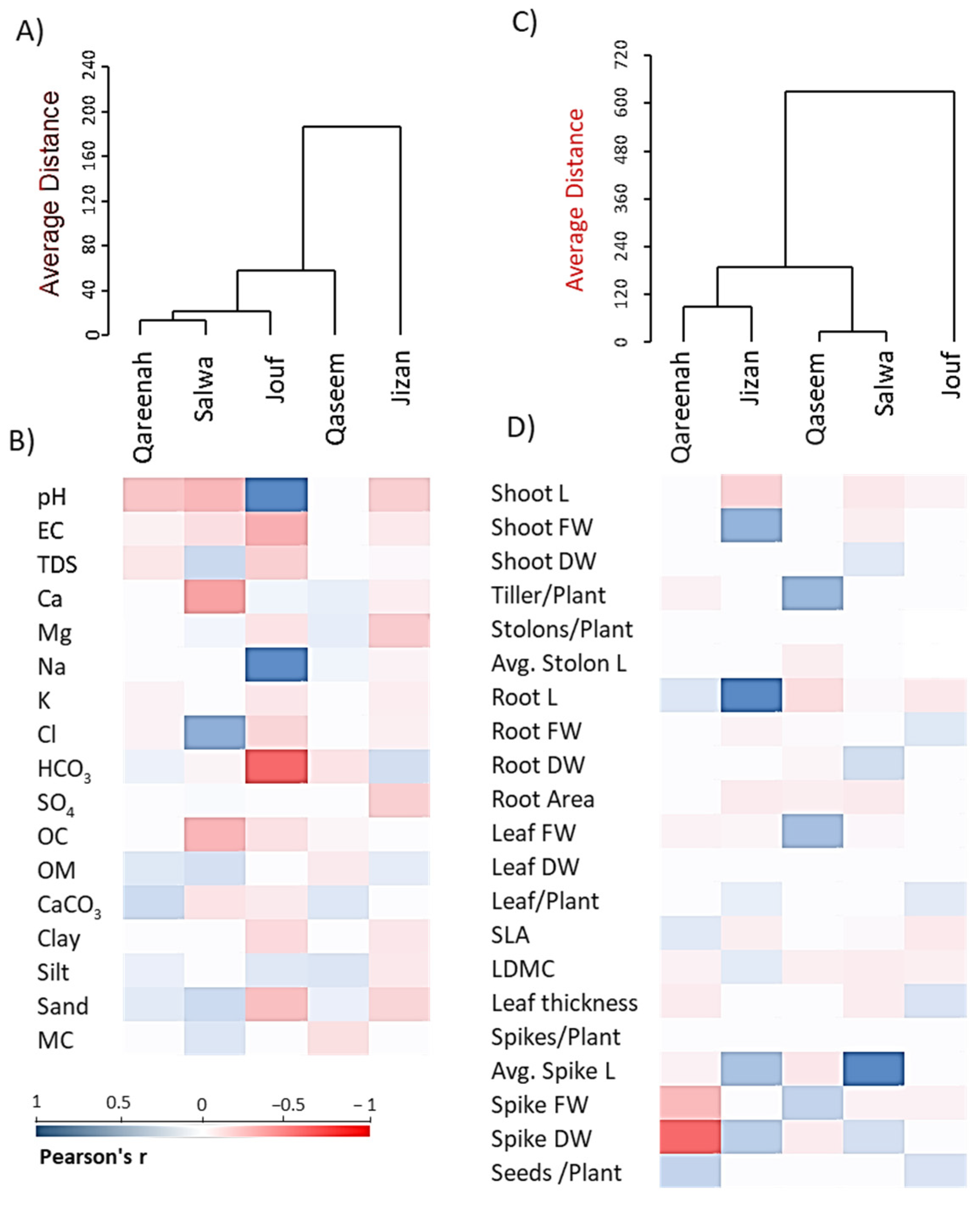
3.5. Correlation Analysis among Functional Plant Traits, Regions, and Soil Variables
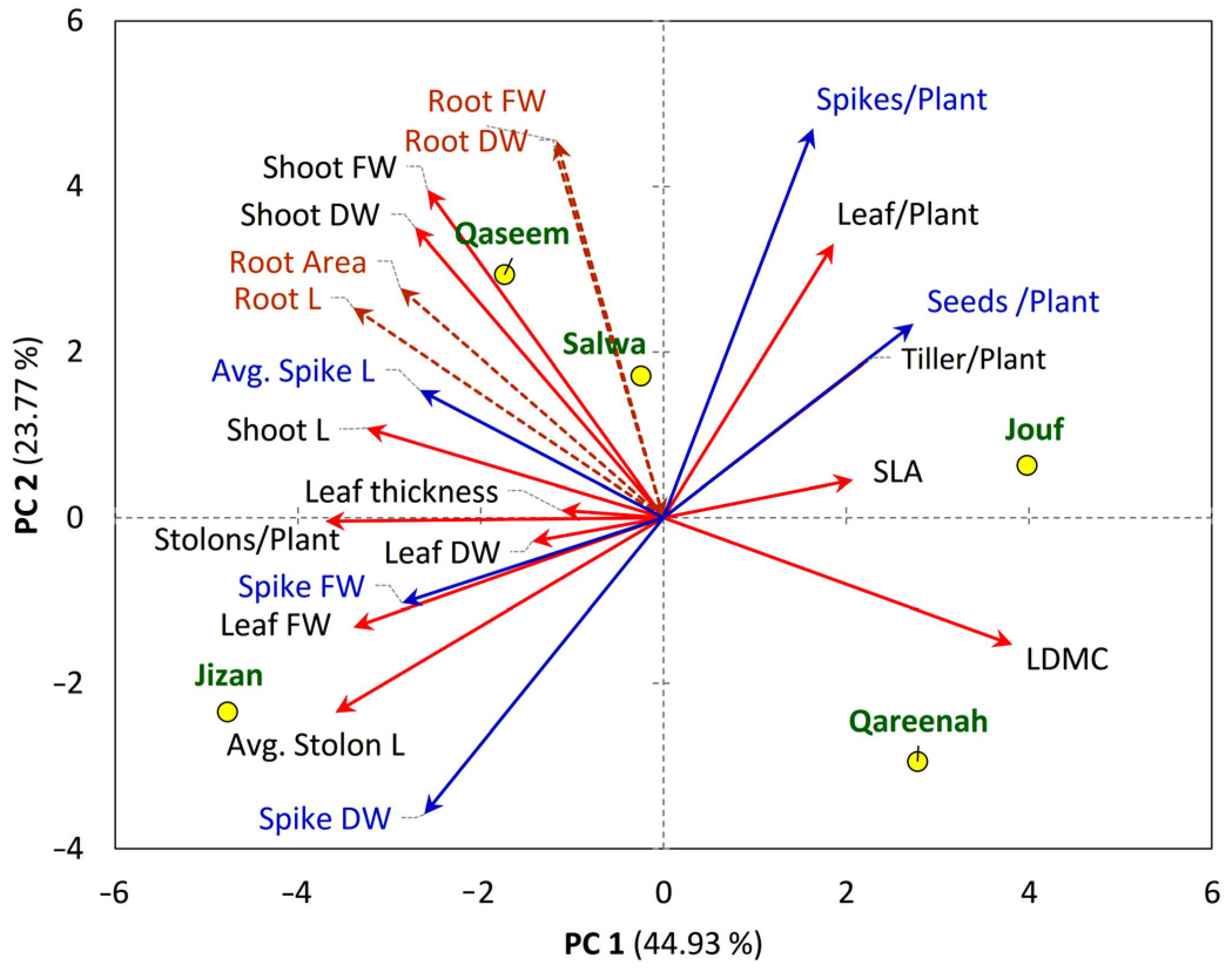
3.5.1. Plant Functional Traits-Regions Correlations
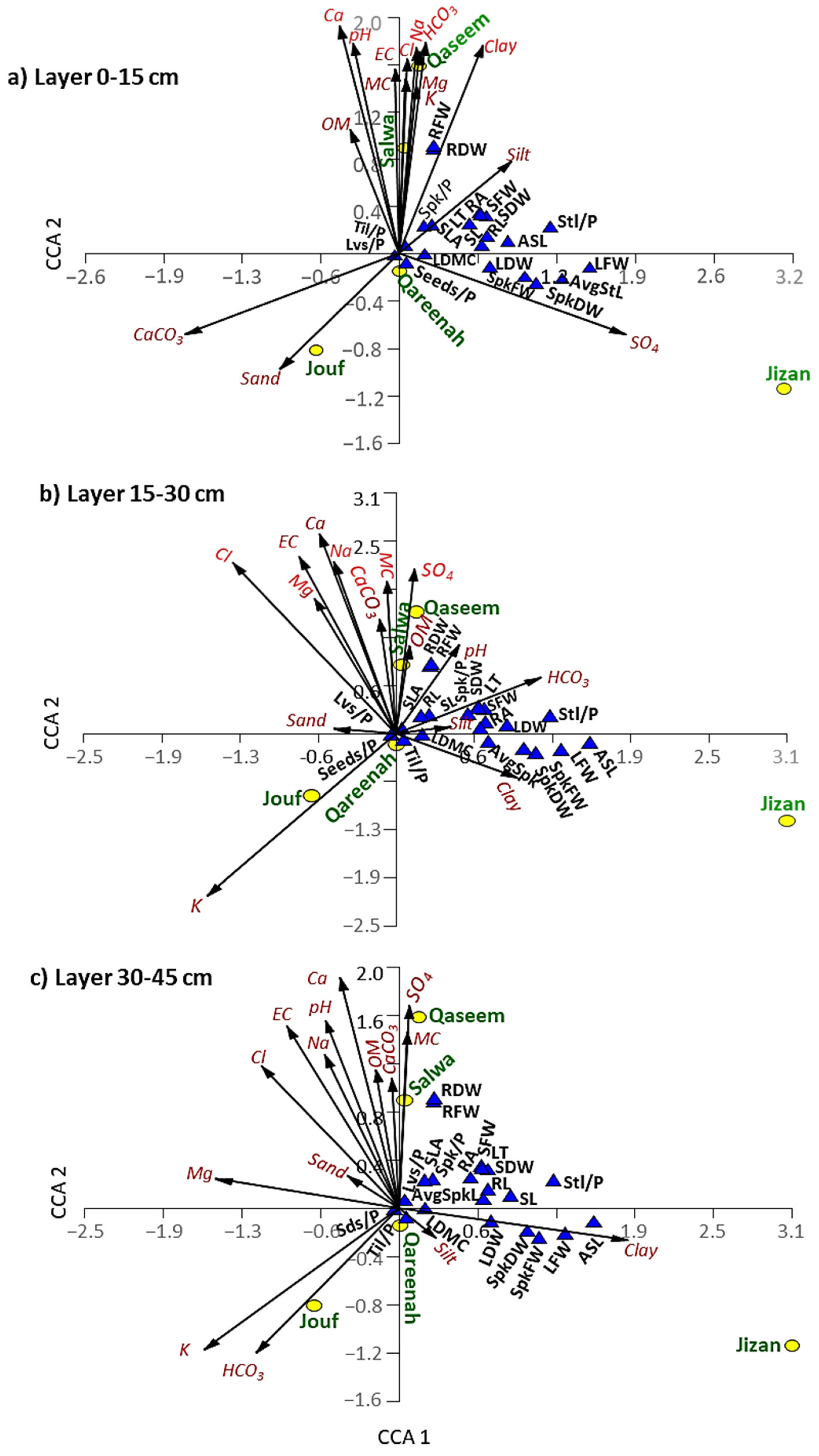
3.5.2. Correlations among Soil Variables, Plant Functional Traits, and Regions
3.6. Cluster Analysis of Regions Based on Soil and Plant Functional Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Hufford, K.M.; Mazer, S.J. Plant ecotypes: Genetic differentiation in the age of ecological restoration. Trends Ecol. Evol. 2003, 18, 147–155. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef]
- Ren, L.; Guo, X.; Liu, S.; Yu, T.; Guo, W.; Wang, R.; Ye, S.; Lambertini, C.; Brix, H.; Eller, F. Intraspecific variation in Phragmites australis: Clinal adaption of functional traits and phenotypic plasticity vary with latitude of origin. J. Ecol. 2020, 108, 2531–2543. [Google Scholar] [CrossRef]
- Scheiner, S.M.; Barfield, M.; Holt, R.D. The genetics of phenotypic plasticity XVII. Response to climate change. Evol. Appl. 2020, 13, 388–399. [Google Scholar] [CrossRef]
- Savolainen, O.; Pyhäjärvi, T.; Knürr, T. Gene flow and local adaptation in trees. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 595–619. [Google Scholar] [CrossRef]
- Schmitt, J. Reaction norms of morphological and life-history traits to light availability in Impatiens capensis. Evolution 1993, 47, 1654–1668. [Google Scholar]
- Dudley, S.A.; Schmitt, J. Testing the adaptive plasticity hypothesis: Density-dependent selection on manipulated stem length in Impatiens capensi. Am. Nat. 1996, 147, 445–465. [Google Scholar] [CrossRef]
- Sultan, S.E. Phenotypic plasticity in plants: A case study in ecological development. Evol. Dev. 2003, 5, 25–33. [Google Scholar] [CrossRef]
- Crick, J.; Grime, J. Morphological plasticity and mineral nutrient capture in two herbaceous species of contrasted ecology. New Phytol. 1987, 107, 403–414. [Google Scholar] [CrossRef]
- Hester, M.W.; Mendelssohn, I.A.; McKee, K.L. Intraspecific variation in salt tolerance and morphology in the coastal grass Spartina patens (Poaceae). Am. J. Bot. 1996, 83, 1521–1527. [Google Scholar] [CrossRef]
- Florin, A.-B.; Höglund, J. Absence of population structure of turbot (Psetta maxima) in the Baltic Sea. Mol. Ecol. 2007, 16, 115–126. [Google Scholar] [CrossRef]
- Hong-bo, S. Plant gene regulatory network system under abiotic stress. Acta Biol. Szeged. 2006, 50, 1–9. [Google Scholar]
- Gulzar, S.; Khan, M.A.; Ungar, I.A. Effects of salinity on growth, ionic content, and plant–water status of Aeluropus lagopoides. Commun. Soil Sci. Plant Anal. 2003, 34, 1657–1668. [Google Scholar] [CrossRef]
- Matesanz, S.; Gianoli, E. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 2014, 1206, 35–55. [Google Scholar] [CrossRef]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef]
- Chevin, L.-M.; Hoffmann, A.A. Evolution of phenotypic plasticity in extreme environments. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160138. [Google Scholar] [CrossRef]
- Jacquemyn, H.; De Meester, L.; Jongejans, E.; Honnay, O. Evolutionary changes in plant reproductive traits following habitat fragmentation and their consequences for population fitness. J. Ecol. 2012, 100, 76–87. [Google Scholar] [CrossRef]
- Gurkok, T.; Dolarslan, M.; Gul, E.; Turktas Erken, M. Adaptive Roles of Plant Development and Genome Structure against Soil Composition. Fresenius Environ. Bull. 2021, 30, 4031–4042. [Google Scholar]
- Petrik, P.; Petek, A.; Konopkova, A.; Bosela, M.; Fleischer, P.; Frydl, J.; Kurjak, D. Stomatal and leaf morphology response of European Beech (Fagus sylvatica L.) provenances transferred to contrasting climatic conditions. Forests 2020, 11, 1359. [Google Scholar] [CrossRef]
- Leimu, R.; Fischer, M. A meta-analysis of local adaptation in plants. PLoS ONE 2008, 3, e4010. [Google Scholar] [CrossRef] [PubMed]
- Youssef, A.M. Salt tolerance mechanisms in some halophytes from Saudi Arabia and Egypt. Res. J. Agric. Biol. Sci. 2009, 5, 191–206. [Google Scholar]
- Stanton-Geddes, J.; Shaw, R.G.; Tiffin, P. Interactions between soil habitat and geographic range location affect plant fitness. PLoS ONE 2012, 7, e36015. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, A.; Santo, A. Effects of photoperiod, thermoperiod, and salt stress on Gymnocarpos decandrus seeds: Potential implications in restoration ecology activities. Botany 2017, 95, 1093–1098. [Google Scholar] [CrossRef]
- Elnaggar, A.; El-Keblawy, A.; Mosa, K.A.; Navarro, T. Adaptive drought tolerance during germination of Salsola drummondii seeds from saline and nonsaline habitats of the arid Arabian deserts. Botany 2019, 97, 123–133. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Shamsi, N.; Mosa, K. Effect of maternal habitat, temperature and light on germination and salt tolerance of Suaeda vermiculata, a habitat-indifferent halophyte of arid Arabian deserts. Seed Sci. Res. 2018, 28, 140–147. [Google Scholar] [CrossRef]
- Miner, B.G.; Sultan, S.E.; Morgan, S.G.; Padilla, D.K.; Relyea, R.A. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol. 2005, 20, 685–692. [Google Scholar] [CrossRef]
- Rúa, M.A.; Antoninka, A.; Antunes, P.M.; Chaudhary, V.B.; Gehring, C.; Lamit, L.J.; Piculell, B.J.; Bever, J.D.; Zabinski, C.; Meadow, J.F. Home-field advantage? Evidence of local adaptation among plants, soil, and arbuscular mycorrhizal fungi through meta-analysis. BMC Evol. Biol. 2016, 16, 1–15. [Google Scholar] [CrossRef]
- Diekmann, M.; Müller, J.; Heinken, T.; Dupré, C. Wiederansiedlungen von Gefäßpflanzenarten in Deutschland—Eine Übersicht und statistische Auswertung. Tuexenia 2015, 35, 249–265. [Google Scholar]
- Bolnick, D.I.; Amarasekare, P.; Araújo, M.S.; Bürger, R.; Levine, J.M.; Novak, M.; Rudolf, V.H.W.; Schreiber, S.J.; Urban, M.C.; Vasseur, D.A. Why intraspecific trait variation matters in community ecology. Trends Ecol. Evol. 2011, 26, 183–192. [Google Scholar] [CrossRef]
- Chapin, F.S. Integrated responses of plants to stress. BioScience 1991, 41, 29–36. [Google Scholar] [CrossRef]
- Shipley, B.; De Bello, F.; Cornelissen, J.H.C.; Laliberté, E.; Laughlin, D.C.; Reich, P.B. Reinforcing loose foundation stones in trait-based plant ecology. Oecologia 2016, 180, 923–931. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Pallas, B.; Da Silva, D.; Valsesia, P.; Yang, W.; Guillaume, O.; Lauri, P.-E.; Vercambre, G.; Génard, M.; Costes, E. Simulation of carbon allocation and organ growth variability in apple tree by connecting architectural and source–sink models. Ann. Bot. 2016, 118, 317–330. [Google Scholar] [CrossRef]
- Mensah, S.; Kakaï, R.G.; Seifert, T. Patterns of biomass allocation between foliage and woody structure: The effects of tree size and specific functional traits. Ann. For. Res. 2016, 59, 49–60. [Google Scholar] [CrossRef]
- Yin, Q.; Tian, T.; Han, X.; Xu, J.; Chai, Y.; Mo, J.; Lei, M.; Wang, L.; Yue, M. The relationships between biomass allocation and plant functional trait. Ecol. Indic. 2019, 102, 302–308. [Google Scholar] [CrossRef]
- Ivanova, K.; Geneva, M.; Anev, S.; Georgieva, T.; Tzvetkova, N.; Stancheva, I.; Markovska, Y. Effect of soil salinity on morphology and gas exchange of two Paulownia hybrids. Agrofor. Syst. 2019, 93, 929–935. [Google Scholar] [CrossRef]
- Hobohm, C.; Schaminée, J.; van Rooijen, N. Coastal Habitats, Shallow Seas and Inland Saline Steppes: Ecology, Distribution, Threats and Challenges. In Perspectives for Biodiversity and Ecosystems; Hobohm, C., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 279–310. [Google Scholar]
- Goodall, T.M.; Al-Belushi, J.D. A Glossary of Arabic Desert Terminology Used in Southeastern Arabia. In Quaternary Deserts and Climatic Change; CRC Press: Boca Raton, FL, USA, 2020; pp. 611–619. [Google Scholar]
- Lõhmus, K.; Balke, T.; Kleyer, M. Spatial and temporal patterns of initial plant establishment in salt marsh communities. J. Veg. Sci. 2020, 31, 1122–1132. [Google Scholar] [CrossRef]
- Westerband, A.C.; Funk, J.L.; Barton, K.E. Intraspecific trait variation in plants: A renewed focus on its role in ecological processes. Ann. Bot. 2021, 127, 397–410. [Google Scholar] [CrossRef]
- Weigelt, A.; Mommer, L.; Andraczek, K.; Iversen, C.M.; Bergmann, J.; Bruelheide, H.; Fan, Y.; Freschet, G.T.; Guerrero-Ramírez, N.R.; Kattge, J. An integrated framework of plant form and function: The belowground perspective. New Phytol. 2021, 232, 42–59. [Google Scholar] [CrossRef]
- Pennings, S.C.; Richards, C.L. Effects of wrack burial in salt-stressed habitats: Batis maritima in a southwest Atlantic salt marsh. Ecography 1998, 21, 630–638. [Google Scholar] [CrossRef]
- Pennings, S.C.; Bertness, M.D. Salt marsh communities. Mar. Community Ecol. 2001, 11, 289–316. [Google Scholar]
- Richards, C.L.; Pennings, S.C.; Donovan, L.A. Habitat range and phenotypic variation in salt marsh plants. Plant Ecol. 2005, 176, 263–273. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Zahran, M.A.; Murphy, K.J.; Mashaly, I.A.; Khedr, A.A. On the ecology of some halophytes and psammophytes in the Mediterranean coast of Egypt. Verh.-Ges. Fur Okologie 1996, 25, 133–146. [Google Scholar]
- Podda, L.; Santo, A.; Leone, C.; Mayoral, O.; Bacchetta, G. Seed germination, salt stress tolerance and seedling growth of Opuntia ficus-indica (Cactaceae), invasive species in the Mediterranean Basin. Flora 2017, 229, 50–57. [Google Scholar] [CrossRef]
- Azad, M.S.; Mollick, A.S.; Ranon, R.J.K.; Khan, M.N.I.; Kamruzzaman, M. Plasticity of leaf morphology of Bruguiera sexangula to salinity zones in Bangladesh’s Sundarbans. J. For. Res. 2022, 33, 1857–1866. [Google Scholar] [CrossRef]
- Yao, B.; Zhao, C.; Deng, J.; Zhou, H.; Zhao, X.; Liu, J. Phenotypic plasticity of Thellungiella salsaginea in response to saline stress. Evol. Ecol. Res. 2013, 15, 829–846. [Google Scholar]
- Cope, T. Flora of Pakistan, No. 143: Poaceae; Pakistan Agricultural Research Council Islamabad and University of Karachi: Islamabad, Pakistan, 1982; p. 678. [Google Scholar]
- Mandaville, J.P. Flora of Eastern Saudi Arabia; Routledge: Abingdon, UK, 2013. [Google Scholar]
- Alfarhan, A.H.; Al-Turki, T.A.; Basahy, A.Y. Flora of Jizan region. Final Rep. Support King Abdulaziz City Sci. Technol. 2005, 1, 545. [Google Scholar]
- Dar, B.A.; Assaeed, A.M.; Al-Rowaily, S.L.; Al-Doss, A.A.; Abd-ElGawad, A.M. Vegetation composition of the halophytic grass Aeluropus lagopoides communities within coastal and inland sabkhas of Saudi Arabia. Plants 2022, 11, 666. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.M.; Assaeed, A.M.; Al-Rowaily, S.L.; Dar, B.M.; Malik, J.A. Moisture and Salinity Drive the Vegetation Composition of Wadi Hargan, Riyadh, Saudi Arabia. Diversity 2021, 13, 587. [Google Scholar] [CrossRef]
- Phondani, P.C.; Bhatt, A.; Elsarrag, E.; Alhorr, Y.M.; El-Keblawy, A. Criteria and indicator approach of global sustainability assessment system for sustainable landscaping using native plants in Qatar. Ecol. Indic. 2016, 69, 381–389. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; Lata, C.; Kumar, S. Eco-physiological responses of Aeluropus lagopoides (grass halophyte) and Suaeda nudiflora (non-grass halophyte) under individual and interactive sodic and salt stress. South Afr. J. Bot. 2016, 105, 36–44. [Google Scholar] [CrossRef]
- Bowles, J.E. Engineering Properties of Soils and Their Measurement; McGraw-Hill, Inc.: New York, NY, USA, 1992. [Google Scholar]
- Rowell, D. Soil Science: Methods and Applications; Longman Group: Harlow, UK, 1994. [Google Scholar]
- Rhoades, J. Methods of Soil Analysis. Part 2, Chemical and Microbiological Properties. In Soluble Salts; Amer Society of Agronomy: Madison, Wisconsin, 1982; pp. 167–179. [Google Scholar]
- Vile, D.; Garnier, E.; Shipley, B.; Laurent, G.; Navas, M.-L.; Roumet, C.; Lavorel, S.; Díaz, S.; Hodgson, J.G.; Lloret, F. Specific leaf area and dry matter content estimate thickness in laminar leaves. Ann. Bot. 2005, 96, 1129–1136. [Google Scholar] [CrossRef]
- Perez-Harguindeguy, N.; Díaz, S.; Garnier Eric, L.S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.; Cornwelf, W.; Craine, J.; Gurvich, D.; Urcelay, C. New handbook for standardized measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Kovach, W. MVSP-A Multivariate Statistical Package for Windows, ver. 3.1; Kovach Computing Services: Pentraeth, Anglesey, 1999; p. 137. [Google Scholar]
- Weber, D. Adaptive mechanisms of halophytes in desert regions. In Salinity and Water Stress; Springer: Dordrecht, Netherlands, 2009; pp. 179–185. [Google Scholar]
- Mansoor, U.; Fatima, S.; Hameed, M.; Naseer, M.; Ahmad, M.S.A.; Ashraf, M.; Ahmad, F.; Waseem, M. Structural modifications for drought tolerance in stem and leaves of Cenchrus ciliaris L. ecotypes from the Cholistan Desert. Flora 2019, 261, 151485. [Google Scholar] [CrossRef]
- Kang, S.; Zhang, J. Controlled alternate partial root-zone irrigation: Its physiological consequences and impact on water use efficiency. J. Exp. Bot. 2004, 55, 2437–2446. [Google Scholar] [CrossRef]
- Nalina, M.; Saroja, S.; Chakravarthi, M.; Rajkumar, R.; Radhakrishnan, B.; Chandrashekara, K. Water deficit-induced oxidative stress and differential response in antioxidant enzymes of tolerant and susceptible tea cultivars under field condition. Acta Physiol. Plant. 2021, 43, 1–17. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, G.; New, T. Abiotic factors influencing the distribution of vegetation in coastal estuary of the Liaohe Delta, Northeast China. Estuaries Coasts 2009, 32, 937–942. [Google Scholar] [CrossRef]
- Grunstra, M.; Van Auken, O. Using GIS to display complex soil salinity patterns in an inland salt marsh. Dev. Environ. Sci. 2007, 5, 407–431. [Google Scholar]
- Lu, S.G.; Tang, C.; Rengel, Z. Combined effects of waterlogging and salinity on electrochemistry, water-soluble cations and water dispersible clay in soils with various salinity levels. Plant Soil 2004, 264, 231–245. [Google Scholar] [CrossRef]
- Barzegargolchini, B.; Movafeghi, A.; Dehestani, A.; Mehrabanjoubani, P. Morphological and anatomical changes in stems of Aeluropus littoralis under salt stress. J. Plant Mol. Breed. 2017, 5, 40–48. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Khan, W.-U.-D.; Tanveer, M.; Shaukat, R.; Ali, M.; Pirdad, F. An Overview of Salinity Tolerance Mechanism in Plants. In Salt and Drought Stress Tolerance in Plants: Signaling Networks and Adaptive Mechanisms; Hasanuzzaman, M., Tanveer, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–16. [Google Scholar]
- Rubio, G.; Walk, T.; Ge, Z.; Yan, X.; Liao, H.; Lynch, J.P. Root gravitropism and below-ground competition among neighbouring plants: A modelling approach. Ann. Bot. 2001, 88, 929–940. [Google Scholar] [CrossRef]
- Wang, W.; Ding, G.-D.; White, P.J.; Wang, X.-H.; Jin, K.-M.; Xu, F.-S.; Shi, L. Mapping and cloning of quantitative trait loci for phosphorus efficiency in crops: Opportunities and challenges. Plant Soil 2019, 439, 91–112. [Google Scholar] [CrossRef]
- Khan, M.A.; Ungar, I.A.; Showalter, A.M. Effects of salinity on growth, ion content, and osmotic relations in Halopyrum mucronatum (L.) Stapf. J. Plant Nutr. 1999, 22, 191–204. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 1–18. [Google Scholar] [CrossRef]
- Pujol, J.A.; Calvo, J.F.; Ramírez-Díaz, L. Seed germination, growth, and osmotic adjustment in response to NaCl in a rare succulent halophyte from southeastern Spain. Wetlands 2001, 21, 256–264. [Google Scholar] [CrossRef]
- Shabala, S.; Mackay, A. Ion transport in halophytes. In Advances in Botanical Research; Elsevier: New York, NY, USA, 2011; Volume 57, pp. 151–199. [Google Scholar]
- Liu, H.-S.; Li, F.-M.; Xu, H. Deficiency of water can enhance root respiration rate of drought-sensitive but not drought-tolerant spring wheat. Agric. Water Manag. 2004, 64, 41–48. [Google Scholar] [CrossRef]
- Todea, I.M.; Gonzalez-Orenga, S.; Boscaiu, M.; Plazas, M.; Sestras, A.F.; Prohens, J.; Vicente, O.; Sestras, R.E. Responses to water deficit and salt stress in silver fir (Abies alba Mill.) seedlings. Forests 2020, 11, 395. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A. Screening methods for salinity tolerance: A case study with tetraploid wheat. Plant Soil 2003, 253, 201–218. [Google Scholar] [CrossRef]
- Naz, N.; Hameed, M.; Nawaz, T.; Batool, R.; Ashraf, M.; Ahmad, F.; Ruby, T. Structural adaptations in the desert halophyte Aeluropus lagopoides (Linn.) Trin. ex Thw. under high salinity. J. Biol. Res. 2013, 19, 150–164. [Google Scholar]
- Zhao, K.F.; Li, F.Z.; Zhang, F.S. Chinese Halophytes, 2nd ed.; China Science Press: Beijing, China, 2013; pp. 71–74. [Google Scholar]
- Hussain, S.; Shaukat, M.; Ashraf, M.; Zhu, C.; Jin, Q.; Zhang, J. Salinity stress in arid and semi-arid climates: Effects and management in field crops. In Climate Change and Agriculture; Hussain, S., Ed.; IntechOpen: London, UK, 2019; Volume 13. [Google Scholar]
- Xiao, Y.; Tang, J.; Qing, H.; Zhou, C.; An, S. Effects of salinity and clonal integration on growth and sexual reproduction of the invasive grass Spartina alterniflora. Flora 2011, 206, 736–741. [Google Scholar] [CrossRef]
- Volis, S. Plasticity, its cost, and phenotypic selection under water and nutrient stress in two annual grasses. Biol. J. Linn. Soc. 2009, 97, 581–593. [Google Scholar] [CrossRef]
- Ilyas, M.; Nisar, M.; Khan, N.; Hazrat, A.; Khan, A.H.; Hayat, K.; Fahad, S.; Khan, A.; Ullah, A. Drought tolerance strategies in plants: A mechanistic approach. J. Plant Growth Regul. 2021, 40, 926–944. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef]
- Kleyer, M.; Minden, V. Why functional ecology should consider all plant organs: An allocation-based perspective. Basic Appl. Ecol. 2015, 16, 1–9. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, M.; Shi, P.; Zong, N.; Zhang, X.; Zhang, X. Variation of leaf and root traits and ecological adaptive strategies along a precipitation gradient on Changtang Plateau. Acta Ecol. Sin. 2020, 40, 295–309. [Google Scholar]
- Tan, J.; Ben-Gal, A.; Shtein, I.; Bustan, A.; Dag, A.; Erel, R. Root structural plasticity enhances salt tolerance in mature olives. Environ. Exp. Bot. 2020, 179, 104224. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Layer | Region | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Qareenah | Qaseem | Salwa | Jouf | Jizan | |||
| pH | 0–15 cm | 8.38 ± 0.18 A,a | 8.39 ± 0.141 A,a | 8.39 ± 0.140 A,a | 8.19 ± 0.108 B,C,a | 8.09 ± 0.220 C,a | 0.0011 ** |
| 15–30 cm | 8.47 ± 0.04 A,a | 8.16 ± 0.027 B,b | 8.22 ± 0.022 AB,b | 8.02 ± 0.051 C,b | 8.10 ± 0.192 C,a | ||
| 30–45 cm | 8.19 ± 0.07 B,b | 8.15 ± 0.034 B,b | 8.27 ± 0.115 A,B,a,b | 8.02 ± 0.097 C,b | 7.88 ± 0.207 C,b | ||
| p-value | 0.0219 * | ||||||
| EC (dS·m−1) | 0–15 cm | 15.39 ± 0.92 B,a | 25.95 ± 3.87 A,a | 7.29 ± 0.17 C,a | 2.74 ± 0.13 C,b | 1.032 ± 0.07 D,a | <0.0001 *** |
| 15–30 cm | 5.17 ± 0.78 B,b | 10.28 ± 1.83 A,b | 5.25 ± 0.26 B,b | 4.52 ± 1.03 C,a | 1.10 ± 0.16 D,a | ||
| 30–45 cm | 3.87 ± 0.46 B,c | 6.32 ± 1.14 A,c | 3.53 ± 0.58 C,c | 3.59 ± 1.12 C,a | 0.94 ± 0.07 D,b | ||
| p-value | <0.0001 *** | ||||||
| Ca2+ (meq/L) | 0–15 cm | 19.90 ± 2.69 C,a | 39.86 ± 3.74 A,a | 31.87 ± 0.43 B,a | 16.45 ± 1.38 D,a | 2.76 ± 0.36 E,a | <0.0001 *** |
| 15–30 cm | 14.08 ± 1.45 C,b | 23.66 ± 3.51 A,b | 24.55 ± 0.84 B,b | 11.12 ± 3.16 D,b | 3.14 ± 0.65 E,a | ||
| 30–45 cm | 15.30 ± 1.54 B,b | 20.17 ± 3.46 A,b | 19.30 ± 2.74 A,c | 9.02 ± 3.06 C,c | 2.15 ± 0.45 E,b | ||
| p-value | <0.0001 *** | ||||||
| Mg2+ (meq/L) | 0–15 cm | 35.40 ± 2.73 B,a | 60.63 ± 8.30 A,a | 8.25 ± 0.43 C,a | 5.77 ± 0.39 C,b | 1.67 ± 0.25 D,b | <0.0001 *** |
| 15–30 cm | 16.10 ± 0.89 B,b | 33.74 ± 3.28 A,b | 8.56 ± 0.36 D,a | 13.51 ± 0.95 C,a | 2.80 ± 0.24 E,a,b | ||
| 30–45 cm | 6.96 ± 0.53 C,c | 11.49 ± 0.55 A,b | 5.83 ± 0.60 D,b | 10.34 ± 1.03 B,a | 2.55 ± 0.32 E,a | ||
| p-value | <0.0001 *** | ||||||
| Na+ (meq/L) | 0–15 cm | 45.05 ± 6.52 B,a | 134.81 ± 33.36 A,a | 30.57 ± 0.71 B,C,a | 5.12 ± 0.06 C,b | 5.38 ± 0.61 C,a | <0.0001 *** |
| 15–30 cm | 25.18 ± 4.89 B,b | 44.34 ± 13.19 A,b | 18.04 ± 1.09 B,C,b | 15.53 ± 4.70 C,a | 3.76 ± 0.60 D,b | ||
| 30–45 cm | 15.43 ± 2.41 B,b | 27.34 ± 6.36 A,b | 9.73 ± 2.83 C,c | 13.49 ± 5.22 B,C,a | 4.54 ± 0.18 c | ||
| p-value | <0.0001 *** | ||||||
| K+ (meq/L) | 0–15 cm | 14.63 ± 4.24 B,a | 20.22 ± 4.97 A,a | 2.16 ± 0.04 C,a | 0.61 ± 0.11 D,c | 0.51 ± 0.09 D,b | 0.0001 *** |
| 15–30 cm | 2.78 ± 0.47 B,b | 1.42 ± 0.34B C,b | 1.75 ± 0.08B C,b | 9.95 ± 2.98 A,a | 2.57 ± 0.08 B,a | ||
| 30–45 cm | 1.88 ± 0.49 B,c | 1.52 ± 0.36 B,C,b, | 1.25 ± 0.40 B,C,b | 3.34 ± 2.36 A,b | 0.52 ± 0.05 C,b | ||
| p-value | <0.0001 *** | ||||||
| Cl− (meq/L) | 0–15 cm | 96.90 ± 13.08 B,a | 237.60 ± 41.19 A,a | 66.36 ± 1.27 B,C,a | 13.04 ± 1.24 C,D,c | 8.65 ± 0.78 D,a | <0.0001 *** |
| 15–30 cm | 48.02 ± 4.62 B,b | 73.30 ± 18.32 A,b | 48.00 ± 1.76 B,C,b | 41.60 ± 10.16 C,a | 7.25 ± 1.42 D,b | ||
| 30–45 cm | 30.94 ± 3.40 B,c | 51.00 ± 11.90 A,c | 25.67 ± 8.35 B,C,c | 32.58 ± 9.77 B,a,b | 7.07 ± 0.78 C,b | ||
| p-value | <0.0001 *** | ||||||
| HCO3− (meq/L) | 0–15 cm | 3.00 ± 0.137 A,a | 3.58 ± 0.54 A,a | 2.43 ± 0.21 A,a | 1.13 ± 0.03 B,b | 1.22 ± 0.03 B,b | 0.2300 ns |
| 15–30 cm | 1.51 ± 0.130Bb | 2.36 ± 0.18Aa | 0.9 ± 0.12Cb | 1.38 + 0.15Bb | 2.09 ± 0.27Aa | ||
| 30–45 cm | 1.14 ± 0.091 B,b | 2.13 ± 0.19 A,a | 1.09 ± 0.05 B,a | 4.14 ± 2.55 A,a | 1.47 ± 0.19 B,b | ||
| p-value | 0.3849 | ||||||
| SO42− (meq/L) | 0–15 cm | 15.04 ± 4.32 B,a | 14.19 ± 3.03 B,C,b | 3.92 ± 0.61 D,b | 12.84 ± 0.16 C,a | 71.80 ± 0.05 A,a | <0.0001 *** |
| 15–30 cm | 6.36 ± 1.39 B,b | 27.13 ± 1.14 A,a | 3.05 ± 0.59 C,b | 1.86 ± 0.43 D,b | 0.96 ± 0.19 E,b | ||
| 30–45 cm | 10.97 ± 1.77 A,a,b | 9.42 ± 2.52 A,c | 8.14 ± 2.60 B,a | 1.13 ± 0.56 C,b | 0.60 ± 0.07 D,b | ||
| p-value | 0.0058 ** | ||||||
| OM (%) | 0–15 cm | 1.63 ± 0.19 A,a | 1.78 ± 0.37 A,a | 0.55 ± 0.02 B,b | 0.81 ± 0.12 B,a | 0.33 ± 0.04 B,b | <0.0001 *** |
| 15–30 cm | 0.98 ± 0.17 A,b | 0.87 ± 0.20 A,b | 0.35 ± 0.02 C,a | 0.45 ± 0.14 B,b | 0.44 ± 0.12 B,a | ||
| 30–45 cm | 0.91 ± 0.33 A,b | 0.54 ± 0.06 B,c | 0.98 ± 0.29 A,a | 0.39 ± 0.13 C,c | 0.26 ± 0.03 D,c | ||
| p-value | 0.0003 *** | ||||||
| CaCO3 (%) | 0–15 cm | 34.84 ± 2.10 A,b | 18.02 ± 2.01 D,a | 20.77 ± 0.77 C,b | 32.58 ± 0.14 B,a | 1.23 ± 0.09 E | <0.0001 *** |
| 15–30 cm | 36.62 ± 0.94 A,b | 16.76 ± 1.04 C,b | 18.31 ± 0.55 B,c | 4.08 ± 0.56 D,b | 0.00 ± 0.00 E | ||
| 30–45 cm | 45.35 ± 1.68 A,a | 17.01 ± 0.88 C,c | 29.36 ± 2.17 B,a | 4.03 ± 0.78 D,b | 0.00 ± 0.00 E | ||
| p-value | <0.0001 *** | ||||||
| Clay (%) | 0–15 cm | 12.48 ± 065 C,a | 16.80 ± 1.26 A,a | 15.14 ± 1.64 B,a | 12.67 ± 0.08 C,a | 14.33 ± 1.03B,a | 0.0023 ** |
| 15–30 cm | 10.84 ± 0.64 C,b | 12.72 ± 0.41 B,b | 7.79 ± 0.13 D,c | 11.20 ± 0.53 B,b | 13.58 ± 1.76 A,b | ||
| 30–45 cm | 10.60 ± 0.54 C,b | 10.84 ± 0.83 C,c | 11.20 ± 0.56 B,b | 10.80 ± 0.83 C,c | 12.32 ± 0.86 A,b | ||
| p-value | <0.0001 *** | ||||||
| Silt (%) | 0–15 cm | 17.28 ± 1.86 C,b | 46.40 ± 2.15 A,b | 6.88 ± 1.17 E,b | 15.25 ± 1.04 D,c | 36.70 ± 1.88 B,b | <0.0001 *** |
| 15–30 cm | 22.48 ± 1.49 D,a | 54.77 ± 1.46 A,a | 2.04 ± 0.27 E,c | 32.28 ± 1.37 C,a | 41.30 ± 2.58 B,a | ||
| 30–45 cm | 22.68 ± 1.51 D,a | 32.54 ± 1.75 A,b | 12.10 ± 1.41 E,a | 26.08 ± 2.56 C,b | 29.61 ± 2.26 B,c | ||
| p-value | <0.0001 *** | ||||||
| Sand (%) | 0–15 cm | 70.24 ± 2.50 C,a | 36.80 ± 1.634 E,b | 78.58 ± 1.46 A,b | 74.41 ± 2.52 B,a | 48.68 ± 2.11 D,b | <0.0001 *** |
| 15–30 cm | 66.68 ± 1.23 B,b | 34.51 ± 0.99 E,b | 90.17 ± 0.36 A,a | 56.53 ± 1.83 C,c | 45.12 ± 2.41 D,c | ||
| 30–45 cm | 66.72 ± 0.72 B,b | 56.62 ± 1.49 D,a | 76.70 ± 1.74 A,b | 63.12 ± 2.72 C,b | 58.07 ± 2.36 D,a | ||
| p-value | <0.0001 *** | ||||||
| MC (%) | 0–15 cm | 7.06 ± 0.55 B,a | 28.83 ± 0.75 A,a | 5.78 ± 0.44 B,a | 4.97 ± 1.01 B,a | 1.64 ± 0.25 C,c | <0.0001 *** |
| 15–30 cm | 5.14 ± 0.62 B,b | 19.40 ± 0.79 A,a | 3.80 ± 0.81 C,c | 4.79 ± 1.08 C,b | 2.55 ± 0.52 C,b | ||
| 30–45 cm | 3.34 ± 0.40 B,c | 23.88 ± 0.96 A,a | 3.75 ± 0.80 B,b | 4.60 ± 0.88 B,b | 3.45 ± 0.52 B,c | ||
| p-value | <0.0001 *** | ||||||
| Functional Traits | Unit | SS | MS | F Value | p Value |
|---|---|---|---|---|---|
| Shoot length | cm | 1901.38 | 475.34 | 40.56 | <0.0001 *** |
| Shoot fresh weight | g | 2221.49 | 555.37 | 5.04 | 0.0019 ** |
| Shoot dry weight | g | 1182.11 | 295.52 | 5.91 | 0.0006 *** |
| Number of tillers/plant | No. | 2251.37 | 562.84 | 42.67 | <0.0001 *** |
| Number of stolons/plant | No. | 151.02 | 37.75 | 7.10 | 0.0002 *** |
| Average stolon length | cm | 22701.03 | 5675.25 | 25.97 | <0.0001 *** |
| Root length | cm | 773.21 | 193.30 | 14.38 | <0.0001 *** |
| Root fresh weight | g | 92.62 | 23.15 | 6.25 | 0.0004 *** |
| Root dry weight | g | 63.22 | 15.80 | 6.89 | 0.0002 *** |
| Root area | cm2 | 5335.64 | 1333.91 | 8.10 | 0.0001 *** |
| Leaf fresh weight | g | 36182.34 | 9045.58 | 13.46 | <0.0001 *** |
| Leaf dry weight | g | 24768.00 | 6192.00 | 675.49 | <0.0001 *** |
| Number of leaves/plant | No. | 342601.02 | 85650.25 | 8.28 | <0.0001 *** |
| Specific leaf area | cm2/g | 0.05 | 0.01 | 7.53 | 0.0001 *** |
| Leaf dry matter content | g | 39.38 | 9.84 | 6.63 | 0.0003 *** |
| Leaf thickness | µm | 223.61 | 55.90 | 1.28 | 0.2902 NS |
| Number of spikes/plant | No. | 12999.52 | 3249.88 | 6.07 | 0.0005 *** |
| Average. spike length | cm | 2.10 | 0.52 | 3.35 | 0.0174 * |
| Spike fresh weight | g | 0.03 | 0.01 | 14.42 | <0.0001 *** |
| Spike dry weight | g | 0.01 | 0.01 | 4.05 | 0.0068 ** |
| Number of seeds/plant | No. | 75 × 107 | 1.87 | 48.56 | <0.0001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assaeed, A.M.; Dar, B.A.; Al-Doss, A.A.; Al-Rowaily, S.L.; Malik, J.A.; Abd-ElGawad, A.M. Phenotypic Plasticity Strategy of Aeluropus lagopoides Grass in Response to Heterogenous Saline Habitats. Biology 2023, 12, 553. https://doi.org/10.3390/biology12040553
Assaeed AM, Dar BA, Al-Doss AA, Al-Rowaily SL, Malik JA, Abd-ElGawad AM. Phenotypic Plasticity Strategy of Aeluropus lagopoides Grass in Response to Heterogenous Saline Habitats. Biology. 2023; 12(4):553. https://doi.org/10.3390/biology12040553
Chicago/Turabian StyleAssaeed, Abdulaziz M., Basharat A. Dar, Abdullah A. Al-Doss, Saud L. Al-Rowaily, Jahangir A. Malik, and Ahmed M. Abd-ElGawad. 2023. "Phenotypic Plasticity Strategy of Aeluropus lagopoides Grass in Response to Heterogenous Saline Habitats" Biology 12, no. 4: 553. https://doi.org/10.3390/biology12040553
APA StyleAssaeed, A. M., Dar, B. A., Al-Doss, A. A., Al-Rowaily, S. L., Malik, J. A., & Abd-ElGawad, A. M. (2023). Phenotypic Plasticity Strategy of Aeluropus lagopoides Grass in Response to Heterogenous Saline Habitats. Biology, 12(4), 553. https://doi.org/10.3390/biology12040553