Simple Summary
Begomovirus has a wide host range and threatens a significant amount of economic damage to many important crops such as tomatoes, beans, cassava, squash and cotton. There are many efforts directed at controlling this disease including the use of insecticides to control the insect vector as well as screening the resistant varieties. The use of synthetic virus or infectious clones approaches has allowed plant virologists to characterize and exploit the genome virus at the molecular and biological levels. By exploiting the DNA of the virus using the infectious clones strategy, the viral genome can be manipulated at specific regions to study functional genes for host–virus interactions. Thus, this review will provide an overview of the strategy to construct infectious clones of Begomovirus. The significance of established infectious clones in Begomovirus study will also be discussed.
Abstract
Begomovirus has become a potential threat to the agriculture sector. It causes significant losses to several economically important crops. Given this considerable loss, the development of tools to study viral genomes and function is needed. Infectious clones approaches and applications have allowed the direct exploitation of virus genomes. Infectious clones of DNA viruses are the critical instrument for functional characterization of the notable and newly discovered virus. Understanding of structure and composition of viruses has contributed to the evolution of molecular plant pathology. Therefore, this review provides extensive guidelines on the strategy to construct infectious clones of Begomovirus. Also, this technique’s impacts and benefits in controlling and understanding the Begomovirus infection will be discussed.
1. Introduction
The first infectious clones of bacteriophage QB was reported in 1978 and this was achieved by inserting the cDNA genome into a plasmid vector [1]. In 1980, the infectious clone of retroviral DNA was constructed [2] and was followed by cDNA of poliovirus genome that was transfected into the mammalian cell in the following year [3]. With these notable discoveries, members of infectious DNAs of almost all virus families have been reported. The construction of infectious clones of plant viruses has been established by plant virologists. To characterise a virus at the molecular level, constructing an infectious clone is necessary to study viral replication, range of host species, movement, pathogenicity, and genomic regions’ roles [4]. However, the construction of an infectious clone that involves the cloning of many subgenomic cDNA fragments is frequently laborious [5]. The construction of an infectious clone is still gruelling and many obstacles remain including unpredictability and toxicity in bacterial host–virus sequences [6,7].
The family Geminiviridae consists of nine genera that mainly affect a wide range of dicotyledonous plants. Of all the nine genera, Begomovirus is the largest member of the group, which has about 322 species. Begomoviruses have either monopartite or bipartite genome. It can only be infecting dicots plants through whitefly (Bemisia tabaci) as the vector. This virus is present in both Old World (both genomes) and the New World (mainly bipartite genome, with one describes as monopartite genomes recently) [8,9]. Bipartite begomoviruses encode seven or eight proteins, while monopartite viruses encode only five or six proteins. Alphasatellites, betasatellites and deltasatellites are three types of circular DNA satellites related to the begomoviruses [10,11]. The International Committee on Taxonomy of Viruses (ICTV) recently revised the International Code of Virus Classification and Nomenclature to allow satellite nucleic acid classification. This alteration creates Tolecusatellitidae, a brood of single-stranded DNA satellites linked with begomoviruses [12].
Bipartite Begomovirus contains two components in their genomes: the DNA-A and DNA-B with a size of 2.5–2.6 kb. DNA-A is the part that makes proteins responsible for functions like replicating the virus, encapsidation of both genomic molecules and viral gene expression regulation. However, DNA-B creates the protein responsible for the mechanism of viral DNA molecules in and among the host cells [13]. DNA-A components in the genomes can replicate autonomously. However, it needs DNA-B components for systemic infection to occur. The intergenic region of DNA-A and DNA-B consists of about 200 bp of conserved sequence (Common Region (CR)), including the TAATATTAC sequence v-ori in the conserved stem-loop. The monopartite Begomovirus genome organization resembles the DNA-A component in the bipartite Begomovirus genome [14]. Figure 1 shows the structure genomes of Begomovirus (adapted from the 10th online report of the International Committee on Taxonomy of Viruses (ICTV)).
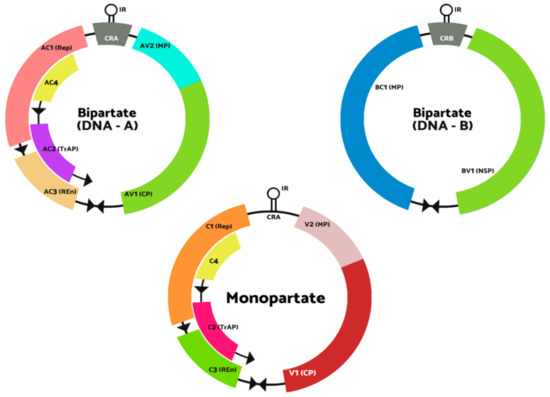
Figure 1.
Genomic structure of begomoviruses. The ORFs are labelled as V if it encodes the virion-sense genes and labelled C if it encodes in complementary-sense genes. According to the genome components, the bipartite genome (top) is tagged with A or B, DNA-A or DNA-B. The two shared “common regions” CRA and CRB genomic components of bipartite viruses are shown in grey inside the intergenic region (IR). The stem-loop position in the IR that contains the conserved TAATATTAC sequence is shown. CP, coat protein; MP, movement protein; NSP, nuclear shuttle protein; REn, replication enhancer protein; Rep, replication-associated protein; TrAP, transcriptional activator protein.
There are two components in the genomes of bipartite Begomovirus: DNA-A and DNA-B, whereas monopartite Begomovirus only contains one genome that resembles DNA-A. DNA-A and monopartite Begomovirus genome are made up of 6 open reading frames encoded for AV1 and AV2 in the virion strand. In contrast, in the complementary strand, there are four encoded proteins, which are AC1, AC2, AC3 and AC4 [14]. In general, AC1 protein is called the replication-associated protein (Rep), which functions in initiating viral DNA replication through the binding of the protein to the iterated motifs (iterons) and introduces a nick to the conserved region (CR) in the intergenic region (IR) [15,16]. According to Arguello-Astroga et al. [17], the Rep protein also coheres to the plant homolog of retinoblastoma protein (Rb) to support the viral DNA replication host factor. Sunter and Bisaro [18] and Bisaro [19] have reported that AC2 protein, which is known as transactivactional activation protein (TraP) that transactivates the expression of viral senses gene expression in DNA-A and B. It also helps in the regulation of transcriptional and post-transcriptional gene silencing (PTGS). AC4 protein plays a vital role as a symptom determinant, which affects cell-cycle control. AC4 protein also counters the host response towards the Rep expression [20]. The DNA-B compartment has two encoded proteins, which are translated from one of the virus strands and the other is from the complementary strand. The BV1 protein is a nuclear-shuttle protein, while BC1 is a movement protein [21,22].
Although the etiological agents causing Begomovirus were whiteflies under natural conditions, it is necessary to create artificial infections with cloned Begomovirus DNAs to study the various aspects of the virus, including the function of gene and virus-plant interactions [23]. Therefore, this review describes methods that are widely used to prepare and test infectious clones for different begomoviruses. The strategy to construct the full-length infectious clones of Begomovirus will be summarised. Also, the importance of infectious clones in understanding the Begomovirus infections and functional genomics will be highlighted.
2. Full-Length Infectious Clones Construction Strategy
An infectious clone is necessary for researchers to study viruses at the molecular and biological levels. An infectious clone has the capability to exploit the viral genome. Mutagenesis of specific regions can be introduced to study the coding part of the genome for studies on host–virus interactions. Figure 2 shows a summary of the methodology used to construct an infectious clone of begomoviruses. Starting from the total plant DNA isolation to obtain the viral genome, this is followed by the viral genome’s amplification. Next is cloning a full-length virus genome and multimer to produce a complete construct of an infectious clone. Finally, transformation into Agrobacterium tumefaciens and transfection to host plants for constructing an infectious clone for Begomovirus.
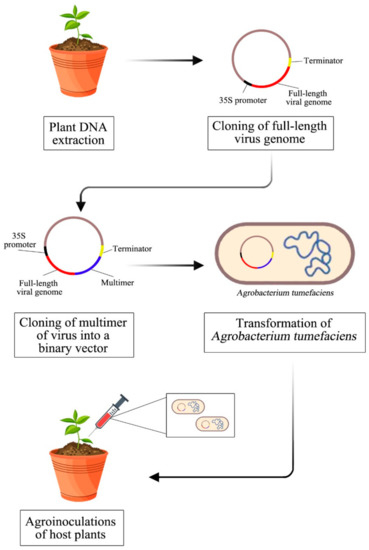
Figure 2.
Summary of the methodology of constructing an infectious clone for Begomovirus.
2.1. Isolation of Total Plant DNA
The first stage of constructing an infectious clone is the isolation of the genome of the virus. Through the extraction of host symptomatic plant total DNA, the viral genome can be obtained. Several protocols have been used widely to produce high-quality and sufficient amounts of DNA. The established protocol by Dellaporta et al. [24], Doyle and Doyle [25], and Palmer et al. [26] are examples of protocols that are well-known. In recent years, the advancement in technology has led to the development of rapid and straightforward kits to extract total plant DNA such as DNeasy Plant Mini Kit (Qiagen, Hilden, Germany), AuPrepTM DNA extraction kit (Life Technologies, Carlsbad, CA, USA), Nucleospin® Plant II total DNA purification kit (Macherey-Nagel, Düren, Germany) and Gentra Puregene DNA Reagents (QIAGEN, Valenia, CA, USA).
2.2. Amplification of Viral Genomes
The next step to produce a full-length infectious clone is the amplification of the complete viral genome from the total plant DNA extracts. There are two commonly used amplification practices for begomoviruses, which are the polymerase chain reaction (PCR) and rolling circle amplification (RCA) [27]. In a standard PCR method, an overlapping primer set needs to be constructed [28]. RCA is an isothermal enzymatic method to amplify a short DNA or RNA primer, forming a lengthy single-stranded DNA or RNA using a circular DNA template and unique phi29 polymerase (Templiphi™, GE, Healthcare). The amplification product would be a long single-stranded DNA with repeating bases that will complement the template. RCA is an advantageous technique to construct infectious clones for Begomovirus [29,30]. RCA is more advantageous than PCR as cloning sequence information is unnecessary, cost-effective and less laborious [31].
2.3. Cloning of Multimer and Full-Length Genome
In general, due to the size of the small genomes of Begomovirus, the cloning of the full genome is relatively easy. To produce a tandem repeat that comprises two origins of replication (V-ori), the construction of infectious clones of Begomovirus require more than one mer (unit) of the genome to boost their ability to infect the host cells [32]. From the amplification through the RCA methods that produce a circular ss-DNA (1.0 mer), a specific restriction enzyme (RE) will be used to linearize the circular DNA. The linearized product will be subsequently being inserted and ligated into a plasmid such as p-GEM-T Easy (Promega), pBlueScript (Stratagene), PTZ57R/T (Fermentas), Pjet1.2/blunt (ThermoFisher) and others that have been treated with the same RE. The plasmid + 1.0 mer were then treated with another RE to produce a multimer (0.01 mer–0.9 mer) that includes the origin of replication (V-ori). The multimer is then transformed into a destination vector and subsequently, a full length (1.0 mer) of the viral genome is ligated together in the destination vector to produce (destination vector + multimer + 1.0 mer).
2.4. Transformation into Agrobacterium Tumafaciens
There are three methods commonly used to transform genes of interest into Agrobacterium tumefaciens: electroporation, the freeze/thaw method and triparental mating [33]. The most efficient way of inserting a gene of interest into Agrobacterium tumefaciens is electroporation. The electroporation method uses electricity to create aqueous pores in the lipid membrane of bacteria. The pores formed are sufficient enough to allow the pass-through of DNA molecules to enter the cells. The regulation of electric field strength and duration of the pulse and the plasmid size determine this method’s efficiency [34,35]. Previously, research by Nair et al. [36] has reported successfully introduce a 200 kb plasmid into Agrobacterium tumefaciens.
As an alternative form of transforming Agrobacterium tumefaciens with foreign DNA, a freeze/thaw technique is considered a fast and straightforward method [37,38]. Based on this technique, the uptake of foreign DNA relies on the damage of the cell wall caused by the exposure to the rapid changes of temperature that alter the membrane fluidity of the bacterial cells [39]. This freeze/thaw technique is widely used as it does not require specialized equipment [33].
Triparental mating is an effective method for cloning a mobilized plasmid into Agrobacterium tumefaciens. This technique uses two Escherichia coli strains to transfer the plasmid carrying the gene of interest into Agrobacterium tumefaciens. The first strain of E. coli has a self-transmissible plasmid, also known as the helper plasmid, as it encodes for the protein that helps in the formation of the mating bridge and the transfer process of itself or another mobilized plasmid to the recipient cells. Once mating is secure, the helper plasmid will transfer itself into the second E. coli strain called the donor strain, which carries the plasmid of interest. Trans-acting functions of the helper plasmid act to transform the plasmid of interest by the donor strain into the Agrobacterium tumefaciens cells [40]. Table 1 below shows the strategy used by geminivirologists in producing infectious clones of different Begomovirus.

Table 1.
Compilation of strategy on cloning an infectious clone for Begomovirus.
2.5. Agroinoculation of Host Plant
Agroinoculation is a technique that allows the transmission of Begomovirus infectious clones into various host plants with the aid of Agrobacterium tumefaciens [95,96]. This method is cost-effective and more straightforward to employ than biolistic inoculation, which required sophisticated equipment such as the particle gun [97]. According to this method, the culture of Agrobacterium tumefaciens, which carries the plasmid of interest, is plated on agar with antibiotic selection specific to the plasmid of interest. The positive growing colonies are selected and grown in a liquid culture containing the antibiotic for further selection. The growing bacteria are then harvested and resuspended in sterile distilled water before subject to the absorbance reading at 600 nm (OD600). A syringe is used to infiltrate the abaxial side of selected plant leaves. A needle is used to incise a small hole beforehand to create a wound. A needled syringe containing Agrobacterium tumefaciens culture bearing the plasmid of interest is used to inoculate the bacteria culture to the side of the plant’s incision. The infiltrated plant is incubated for a short period (1–4 days) before conducting the microscopy analysis. In general, it is best to use the immature plants for agroinoculation as they are more prone to infection with Agrobacterium. Alternative to agroinoculation, the biolistic method by which host plants are bombarded with the DNA containing-gold particles has been used in several studies of Begomovirus infectivity [43,72,89,91].
3. The Significance of Established Infectious Clones in the Begomovirus Study
The construction of an infectious clone allows functional analysis of the viral at the molecular level and their biological roles in replication, pathogenesis and transmission [32,98]. Creating infectious clones that contain wild-type viral genomes enables the establishment of an inoculum that can be used for resistance screening, this is necessary to avoid a genetically inconsistent viral population. Also, this eliminates the need for the maintenance and passage of the virus within plants. Infectious clones also provide an alternative to ineffective inoculation procedures such as grafting and infections transmitted via insect vector [99]. Infectious clones can be altered to achieve virus-induced gene silencing (VIGS) without the need for stable plant transgenesis as well as expression vectors for the plant where it can be applied in plant pathology and used in plant gene function study [100,101]. Despite this, the ability to reconstitute the virus that is fully functional and transmissible will lead to the uncontrolled use of infectious clones that may pose a risk to the environment. These risks could arise due to their inherent pathogenicity and the effect of any introduced genetic modifications [99].
In Begomovirus, the construction of infectious clones has enabled the acquisition of a huge amount of knowledge. Artificial manipulation of Begomovirus genomes such as site-directed mutagenesis, deletion or insertion and rearrangement have reported gaining new insights related to virus-host cell interactions. Infectious clones in the genus Begomovirus also essential for screening resistant varieties.
3.1. Functional Study on the Begomovirus Genomes Using Infectious Clones
Infectious clones for plant viruses are necessary tools in molecular virology as the cloned genome is easily manipulated. The characterization of a virus is possible by generating infectious clones [102].
3.1.1. V2 Protein Functional Analysis
In monopartite Begomovirus, V2 plays its role as movement protein, whereas in bipartite Begomovirus, its function is being replaced by the BC1 protein of DNA-B. Infectivity analysis of Tomato leaf curl Palampur virus (ToLCPalV) and its AV2 mutants of ToLCPalV have revealed that AV2 protein is necessary for symptoms development such as leaves curling down, epinasty and chlorotic leaves. AV2 protein of Okra enation leaf virus (OELV) also reported to function as symptomatic induction, the OELV AV2 mutants cannot infect Nicotiana benthamiana, no necrosis or leaf-curling symptom observed [103,104].
3.1.2. Functional Studies of C1 Protein
Replication-initiator protein (Rep) encoded by the AC1 gene is vital for virus replication. In the African Cassava mosaic virus (ACMV), Rep protein has been reported to contribute to viral infections [105]. Throughout Rolling Circle Replication (RCR), Rep will cleave the strand of virion-sense in a common region inside the mononucleotide sequence and secure it after one round of replication, similar actions as an endonuclease and nucleotidyltransferase [15,106]. A potential cyclin interaction motif (RXL) in the ACMV Rep sequence can be an alternative cycle controls to recognise interaction with retinoblastoma protein (pRBR) within plant homologs. The RXL motif also is necessary for Nicotiana benthamiana plants’ virus infection in producing the symptoms {105].
3.1.3. Functional Studies of C2 Protein
Elucidation of the role of C2 protein in symptom determination and replication in the Bhendi yellow mosaic virus (BYMV) has been conducted through the construction of C2 mutants in BYMV. Two stop codons were inserted into the ORF that encodes the C2 protein. The host plant inoculated with BYMV bearing a mutated C2 protein displays the suppression of symptoms and a decrease in viral load. This suggests that BYMV C2 protein is vital in infection as it involves symptom determination and the viral replication process [107]. However, in different Begomovirus, C2 protein may play a different role. Stanley et al. [108] reported the C2 protein in Beet curly top virus (BCTV) does not play any role in infectivity. In contrast, a study conducted by Baliji et al. [109] in Spinach curly top virus (SCTV) has suggested that C2 mutant of SCTV produces a milder symptom of infection, which in turn indicates that C2 protein is required for infectivity.
3.1.4. C4 Protein Functional Analysis
In general, ORF encodes for AC4 protein in Begomovirus function in the development of symptoms. The loss of C4 function leads to the attenuation of the symptom and reduced infectivity [110,111,112,113]. In bipartite Begomovirus, the function of AC4 protein is less conserved as compared to monopartite Begomovirus. This is shown by the alignment of the AC4 region sequence of bipartite Begomovirus, indicating that the sequence is less conserved. Bipartite African cassava mosaic virus with AC4 mutants (ACMV) expressed symptoms and infection [114,115]. While in Tomato golden mosaic virus (TGMV), mutants of AC4 show no effect on the virus’s symptom development and infectivity [116]. On the other hand, the AC4 protein in some bipartite Old World Begomovirus may function as a suppressor of PTGS by binding to miRNAs and impeding plant growth [117,118]. This shows that the function of AC4 protein is diverse and depends on the type of virus.
3.1.5. Functional Studies on Betasatellite DNAβ of Monopartite Begomovirus
Monopartite Begomovirus genome mimics the DNA-A from bipartite Begomovirus [116]. In recent years, DNAβ, a symptom modulating betasatellites, has been associated with monopartite Begomovirus [119,120]. DNAβ is about 1.3–1.4 kb in size. Approximately half the virus genome’s size DNAβ carries a βC1 gene that is conserved, can suppress RNA silencing and hold together to ssDNA dsDNA in vitro in a sequence non-specific manner [121]. A study on the Tomato leaf curl China virus (ToLCCNV) by Yang et al. [122] found that this virus is related to a betasatellite molecule (ToLCCNB). TolCCNV is inoculated on Nicotiana benthamiana and no symptoms were observed. However, the plant begins to show a downward leaf curling symptom when transfected with ToLCCNV and ToLCCNB. This infectivity test showed that ToLCCNB is needed for the symptoms to appear. Also, the DNAβ with a mutated βC1 retains its capability for silencing suppressor, no typical symptom in Nicotiana benthamiana is observed when it is coinfected with ToLCCNV [122]. Table 2 shows the summary of functional studies conducted on Begomovirus genome.
Table 2.
Summary of functional studies conducted on the genome of Begomovirus.
3.2. Infectious Clones for Screening Resistant Varieties
One of the approaches to managing and controlling Begomovirus-causative diseases is based on the viral vector’s control, which is the whitefly (Bemisia tabaci) even this approach is known to be laborious [123]. Without infectious clones, researchers need to maintain a virus-free whitefly colony and propagate a certain whitefly biotype that transmits the plant’s disease and about 1500 whiteflies per plant are needed to successfully transmit African cassava mosaic virus (ACMV) clones into the cassava plant [124]. This indicates that a vast number of whiteflies are required to transmit the clone into the plant successfully. A previous study has reported that the whitefly transmission is controlled by selecting preferences of the vector and biotype compatibility with tested plants [125]. Additionally, wide-ranging screening genotypes of cassava for resistance to whitefly transmission has also shown favourable whitefly manifestation among cassava genotypes [126]. To deploy a resistant variety, early screening for resistance in the breeding line is desirable. The availability of infectious virus clones without depending on the natural infestation of viruliferous whiteflies could help early screening for the resistant varieties. Hence, resistant cultivar has become the best alternative for disease control [98,127].
As for early screening for resistant variety, biolistic inoculation was used to deliver cloned virus into cassava plant infection, speeding up evaluating cassava cultivars’ tolerance in the breeding program. The usage of biolistic has been reported to help determine the resistance level to cassava mosaic disease in cassava breeding lines [128]. The biolistic inoculation of DNA Cassava mosaic virus (Begomovirus) is more rapid in drawing out the infection symptom, which is as early as 10 days post-inoculation. Moreover, successfully infected plants produced are more significant in number. Hence, this indicates that using biolistic mediated transmission is more efficient than the whitefly-transmission technique [123,128].
4. Future Perspectives
For decades, the assembly of DNA constructs and viral clones has been conducted using the use of restriction endonucleases to create compatible ends. The presence or absence of restriction sites in the viral genome and vector sequences prevented this approach from being successful. Plant virologists have adopted several cloning methods to overcome these limitations such as overlap-based methods, Gibson assembly, Golden Gate cloning and yeast homologous recombination. Overlap-based methods are versatile, flexible and overcome the major constraints associated with the use of restriction enzymes. Overlap-based cloning allows for the rapid and efficient assembly of infectious clones. A simple approach to obtain infectious clones of the Bean golden mosaic virus was demonstrated using the PCR–Gibson Assembly technique [129]. The dimeric constructs that contain two replication origins are needed for each genomic component of Begomovirus infectious clones, and the single-step GA was successfully used to assemble the genome of the virus in an isothermal reaction. Golden Gate cloning and yeast homologous recombination has not yet been reported in the development of infectious clones in begomoviruses, and this might be due to their small genomes and cloning of the genome being relatively easy.
The potential of synthetic biology has been demonstrated through the development of functional and artificial genomes. In particular, the assembly of synthetic replicons from virus genomes has been shown to be an effective method for de novo synthesis. Several infectious clones from plant virus’s genomes have been reported to have been synthesized entirely using de novo synthesis and assembly approach [130,131,132]. In Begomovirus, the presence of conserved sequences allows the Begomovirus to utilize its degenerate “universal” primers to amplify the entire genome, even from total plant DNA. Genotypic sequences of begomoviruses were generated through a viral metagenomic approach using RCA–NGS (rolling circle amplification–next generation sequencing). The method was described to enable the analysis of the diversity of begomoviruses in North America, and 19 complete genome begomoviruses and one alphasatellite were assembled [133]. With the use of next-generation sequencing technology, the authenticity and reproducibility of begomoviral sequences will be verified by comparing them to those obtained using conventional methods such as PCR and RCA.
Cell-free cloning techniques have been used to obtain infectious clones from plant viruses [134,135]. Uncloned genome copies can be obtained by in vitro amplification and the products will be used to inoculate plants by rubbing or biolistic approach. This method is typically not suitable for reverse genomic studies [136]. The discovery that Agrobacterium can successfully infect plants with infectious clones opens new avenues of study in plant virology. As for delivering plants DNA or RNA viruses, agro-inoculation provides the most efficient and universal method. Its potential is greatly increased by the use of synthetic biology strategies [137,138]. Full-length infectious clones can be engineered to be optimized for industrial applications, sources of biomaterial and nanotechnology tools and viral vectors [132].
5. Conclusions
The availability of infectious clones for begomoviruses has proven to be a powerful molecular tool. This technique allows the functional study of a viral gene and its biological properties, and facilitates the genetic screening of germplasm for resistance to viruses. The agroinoculation of viral clones using Agrobacterium tumefaciens and its binary vectors is a cost-effective and highly efficient method for analyzing the Begomovirus’s infectivity. Thus, it is widely used by virologists. To date, the processes for producing a full-length infectious clone construct have been simplified and continuous developments of kits and methodologies have been made available to allow a better understanding of viral infectivity.
Author Contributions
M.F.M.S., A.R.S. and H.B. prepared the original draft of the manuscript. M.A.A., S.N.B., A.B.R., N.T. and H.B. have reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Higher Education Malaysia, grant number FRGS/1/2019/STG05/UKM/02/2 and Universiti Kebangsaan Malaysia, grant number DIP-2020-019.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing does not apply to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Taniguchi, T.; Palmieri, M.; Weissmann, C. QB DNA-containing hybrid plasmids giving rise to QB phage formation in the bacterial host. Nature 1978, 274, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Lowy, D.R.; Rands, E.; Chattopadhyay, S.K.; Garon, C.F.; Hager, G.L. Molecular cloning of infectious integrated murine leukemia virus DNA from infected mouse cells. Proc. Natl. Acad. Sci. USA 1980, 77, 614–618. [Google Scholar] [CrossRef]
- Racaniello, V.R.; Baltimore, D. Cloned poliovirus complementary DNA is infectious in mammalian cells. Science 1981, 214, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Fang, X.-D.; Qiao, J.-H.; Gao, Q.; Wang, X.-B. Reverse genetics system of plant negative-strand RNA viruses are difficult to developed but powerful for virus-host interaction studies and virus-based vector applications. Phytopathol. Res. 2020, 2. [Google Scholar] [CrossRef]
- Kannan, M.; Zainal, Z.; Ismail, I.; Baharum, S.N.; Bunawan, H. Application of reverse genetics in functional genomics of Potyvirus. Viruses 2020, 12, 803. [Google Scholar] [CrossRef]
- Aubry, F.; Nougairede, A.; Gould, E.A.; Lamballerie, X. Flavivirus reverse genetic systems, construction techniques and applications: A historical perspective. Antivir. Res. 2015, 114, 67–85. [Google Scholar] [CrossRef]
- Kannan, M.; Ismail, I.; Bunawan, H. Maize Dwarf Mosaic Virus: From Genome to Disease Management. Viruses 2018, 10, 492. [Google Scholar] [CrossRef]
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Sau, A.R.; Nazmie, N.M.F.; Yusop, M.S.M.; Akbar, M.A.; Saad, M.F.M.; Baharum, S.N.; Talip, N.; Goh, H.H.; Kassim, H.; Bunawan, H. First report of pepper vein yellows virus (PeVYV) and pepper yellow leaf curl virus (PepYLCV) infecting Chili Pepper (Capsicum annuum L.) in Malaysia. Plant Dis. 2020, 104. [Google Scholar] [CrossRef]
- Zhou, X. Advances in understanding begomovirus satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef]
- Lozano, G.; Trenado, H.P.; Fiallo-Olivé, E.; Chirinos, D.; Geraud-Pouey, F.; Briddon, R.W.; Navas-Castillo, J. Characterization of Non-coding DNA Satellites Associated with Sweepoviruses (Genus Begomovirus, Geminiviridae)—Definition of a Distinct Class of Begomovirus-Associated Satellites. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Molecular and Biological Characterization of a New World Mono-/Bipartite Begomovirus/Deltasatellite Complex Infecting Corchorus siliquosus. Front. Microbiol. 2020, 11, 1755. [Google Scholar] [CrossRef]
- Pandey, V.; Srivastava, A.; Gaur, R.K. Begomovirus: A curse for the agricultural crops. Arch. Phytopathol. Plant Prot. 2021. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 105–140. [Google Scholar] [CrossRef]
- Laufs, J.; Traut, W.; Heyraud, F.; Matzeit, V.; Rogers, S.G.; Schell, J.; Gronenborn, B. In vitro cleavage and joining at the viral origin of replication by the replication initiator protein of tomato yellow leaf curl virus. Proc. Natl. Acad. Sci. USA 1995, 92, 3879–3883. [Google Scholar] [CrossRef]
- Fontes, E.P.; Eagle, P.A.; Sipe, P.S.; Luckow, V.A.; Hanley-Bowdoin, L. Interaction between a geminivirus replication protein and origin DNA is essential for viral replication. J. Biol. Chem. 1994, 269, 8459–8465. [Google Scholar] [CrossRef]
- Arguello-Astorga, G.; Lopez-Ochoa, L.; Kong, L.J.; Orozco, B.M.; Settlage, S.B.; Hanley-Bowdoin, L. A novel motif in geminivirus replication proteins interacts with the plant retinoblastoma-related protein. J. Virol. 2004, 78, 4817–4826. [Google Scholar] [CrossRef]
- Sunter, G.; Bisaro, D.M. Transactivation of geminivirus AR1 and BR1 gene expression by the viral AL2 gene product occurs at the level of transcription. Plant Cell 1992, 4, 1321–1331. [Google Scholar] [CrossRef]
- Bisaro, D.M. Silencing suppression by geminivirus proteins. Virology 2006, 344, 158–168. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Bejarano, E.R.; Robertson, D.; Mansoor, S. Geminiviruses: Masters at redirecting and reprogramming plant processes. Nat. Reviews. Microbiol. 2013, 11, 777–788. [Google Scholar] [CrossRef]
- Noueiry, A.O.; Lucas, W.J.; Gilbertson, R.L. Two proteins of a plant DNA virus coordinate nuclear and plasmodesmal transport. Cell 1994, 76, 925–932. [Google Scholar] [CrossRef]
- Sanderfoot, A.A.; Lazarowitz, S.G. Getting it together in plant virus movement: Cooperative interactions between bipartite geminivirus movement proteins. Trends Cell Biol. 1996, 6, 353–358. [Google Scholar] [CrossRef]
- Kushawaha, A.K.; Dasgupta, I. Infectivity of cloned begomoviral DNAs: An appraisal. Virus Dis. 2018, 30, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Molacular Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation from small amount of fresh leaf tissue. Phytochemistry 1987, 19, 11–15. [Google Scholar]
- Palmer, K.E.; Schnippenkoetter, W.H.; Rybicki, E.P. Geminivirus isolation and DNA extraction. Methods Mol. Biol. 1998, 81, 41–52. [Google Scholar] [CrossRef]
- Inoue-Nagata, A.K.; Albuquerque, L.C.; Rocha, W.B.; Nagata, T. A simple method for cloning the complete begomovirus genome using the bacteriophage phi29 DNA polymerase. J. Virol. Methods 2004, 116, 209–211. [Google Scholar] [CrossRef]
- Kiss, M.M.; Ortoleva-Donnelly, L.; Beer, N.R.; Warner, J.; Bailey, C.G.; Colston, B.W.; Rothberg, J.M.; Link, D.R.; Leamon, J.H. High-throughput quantitative polymerase chain reaction in picoliter droplets. Anal. Chem. 2008, 80, 8975–8981. [Google Scholar] [CrossRef]
- Nagata, T.; Inoue-Nagata, A.K. Simplified methods for the construction of RNA and DNA virus infectious clones. Methods Mol. Biol. (Cliftonn. J.) 2015, 1236, 241–254. [Google Scholar] [CrossRef]
- Wu, C.Y.; Lai, Y.C.; Lin, N.S.; Hsu, Y.H.; Tsai, H.T.; Liao, J.Y.; Hu, C.C. A simplified method of constructing infectious clones of begomovirus employing limited restriction enzyme digestion of products of rolling circle amplification. J. Virol. Methods 2008, 147, 355–359. [Google Scholar] [CrossRef]
- Bang, B.; Lee, J.; Kim, S.; Park, J.; Nguyen, T.T.; Seo, Y.S. A Rapid and Efficient Method for Construction of an Infectious Clone of Tomato yellow leaf curl virus. Plant Pathol. J. 2014, 30, 310–315. [Google Scholar] [CrossRef]
- Trenado, H.P.; Orílio, A.F.; Márquez-Martín, B.; Moriones, E.; Navas-Castillo, J. Sweepoviruses cause disease in sweet potato and related Ipomoea spp.: Fulfilling Koch’s postulates for a divergent group in the genus begomovirus. PLoS ONE 2011, 6, e27329. [Google Scholar] [CrossRef]
- Wise, A.A.; Liu, Z.; Binns, A.N. Three methods for the introduction of foreign DNA into Agrobacterium. Methods Mol. Biol. (Cliftonn. J.) 2006, 343, 43–53. [Google Scholar] [CrossRef]
- Dower, W.J.; Miller, J.F.; Ragsdale, C.W. High efficiency transformation of E. coli by high voltage electroporation. Nucleic Acids Res. 1988, 16, 6127–6145. [Google Scholar] [CrossRef]
- Mersereau, M.; Pazour, G.J.; Das, A. Efficient transformation of Agrobacterium tumefaciens by electroporation. Gene 1990, 90, 149–151. [Google Scholar] [CrossRef]
- Nair, G.R.; Liu, Z.; Binns, A.N. Reexamining the role of the accessory plasmid pAtC58 in the virulence of Agrobacterium tumefaciens strain C58. Plant Physiol. 2003, 133, 989–999. [Google Scholar] [CrossRef][Green Version]
- An, G. Binary TI vectors for plant transformation and promoter analysis. Methods Enzymol. 1987, 153, 292–293. [Google Scholar] [CrossRef]
- Chen, H.; Nelson, R.S.; Sherwood, J.L. Enhanced recovery of transformants of Agrobacterium tumefaciens after freeze-thaw transformation and drug selection. Biotechniques 1994, 16, 664–670. [Google Scholar]
- Yusop, M.; Saad, M.; Talip, N.; Baharum, S.N.; Bunawan, H. A Review on Viruses Infecting Taro (Colocasia esculenta (L.) Schott). Pathogens 2019, 8, 56. [Google Scholar] [CrossRef]
- Waters, V.L. Conjugative transfer in the dissemination of beta-lactam and aminoglycoside resistance. Front. Biosci. 1999, 4, D433–D456. [Google Scholar] [CrossRef]
- Srivastava, A.; Raj, S.K.; Kumar, S.; Snehi, S.K.; Kulshreshtha, A.; Hallan, V.; Pande, S.S. Molecular identification of Ageratum enation virus, betasatellite and alphasatellite molecules isolated from yellow vein diseased Amaranthus cruentus in India. Virus Genes 2013, 47, 584–590. [Google Scholar] [CrossRef]
- Xiong, Q.; Fan, S.; Wu, J.; Zhou, X. Ageratum yellow vein China virus Is a Distinct Begomovirus Species Associated with a DNAbeta Molecule. Phytopathology 2007, 97, 405–411. [Google Scholar] [CrossRef]
- Briddon, R.W.; Mansoor, S.; Bedford, I.D.; Pinner, M.S.; Markham, P.G. Clones of cotton leaf curl geminivirus induce symptoms atypical of cotton leaf curl disease. Virus Genes 2000, 20, 19–26. [Google Scholar] [CrossRef]
- Wu, J.; Zulfiqar, A.; Huang, C. Infectivity of Euphorbia leaf curl virus and interaction with Tomato yellow leaf curl China betasatellite. Arch. Virol. 2011, 156, 517–521. [Google Scholar] [CrossRef]
- Marwal, A.; Kumar, R.; Paul Khurana, S.M.; Gaur, R.K. Complete nucleotide sequence of a new geminivirus isolated from Vitis vinifera in India: A symptomless host of Grapevine red blotch virus. Virus Dis. 2019, 30, 106–111. [Google Scholar] [CrossRef]
- Wu, J.; Mugiira, R.B.; Zhou, X. Malvastrum leaf curl Guangdong virus is a distinct monopartite begomovirus. Plant Pathol. 2007, 56, 771–776. [Google Scholar] [CrossRef]
- Guo, W.; Jiang, T.; Zhang, X.; Li, G.; Zhou, X. Molecular variation of satellite DNA beta molecules associated with Malvastrum yellow vein virus and their role in pathogenicity. Appl. Environ. Microbiol. 2008, 74, 1909–1913. [Google Scholar] [CrossRef][Green Version]
- Jiang, T.; Liu, P.; Liao, B.L.; Wu, J.X.; Huang, C.J. Malvastrum yellow vein Yunnan virus is amonopartite begomovirus. Acta Virol. 2010, 54, 21–26. [Google Scholar] [CrossRef]
- Zhang, H.; Ma, X.Y.; Qian, Y.J.; Zhou, X.P. Molecular characterization and infectivity of Papaya leaf curl China virus infecting tomato in China. J. Zhejiang Univ. Sci. B 2010, 11, 109–114. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fontenele, R.S.; Abreu, R.A.; Lamas, N.S.; Alves-Freitas, D.; Vidal, A.H.; Poppiel, R.R.; Melo, F.L.; Lacorte, C.; Martin, D.P.; Campos, M.A.; et al. Passion Fruit Chlorotic Mottle Virus: Molecular Characterization of a New Divergent Geminivirus in Brazil. Viruses 2018, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Fan, S.; Guo, X.; Zhou, X. Stachytarpheta leaf curl virus is a novel monopartite begomovirus species. Arch. Virol. 2005, 150, 2257–2270. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Lee, G.; Park, J.; Lee, T.K.; Choi, H.S.; Lee, S. Molecular characterization and an infectious clone construction of sweet potato leaf curl virus (SPLCV) isolated from Korea. Acta Virol. 2012, 56, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Zhang, P. Molecular characterization of two sweepoviruses from China and evaluation of the infectivity of cloned SPLCV-JS in Nicotiana benthamiana. Arch. Virol. 2012, 157, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xie, Y.; Zhou, X. Tobacco curly shoot virus DNAbeta Is Not Necessary for Infection but Intensifies Symptoms in a Host-Dependent Manner. Phytopathology 2005, 95, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Jiang, T.; Zhou, X. Agroinoculation Shows Tobacco leaf curl Yunnan virus is a Monopartite Begomovirus. Eur. J. Plant Pathol. 2006, 115, 369–375. [Google Scholar] [CrossRef]
- Khan, A.J.; Akhtar, S.; Singh, A.K.; Al-Shehi, A.A.; Al-Matrushi, A.M.; Ammara, U.; Briddon, R.W. Recent evolution of a novel begomovirus causing tomato leaf curl disease in the Al-Batinah region of Oman. Arch. Virol. 2014, 159, 445–455. [Google Scholar] [CrossRef]
- Khan, A.J.; Akhtar, S.; Singh, A.K.; Briddon, R.W. A Distinct Strain of Tomato leaf curl Sudan virus Causes Tomato Leaf Curl Disease in Oman. Plant Dis. 2013, 97, 1396–1402. [Google Scholar] [CrossRef]
- Al-Saleh, M.A.; Al-Shahwan, I.M.; Brown, J.K.; Idris, A.M. Molecular characterization of a naturally occurring intraspecific recombinant begomovirus with close relatives widespread in southern Arabia. Virol. J. 2014, 11, 103. [Google Scholar] [CrossRef]
- Kon, T.; Dolores, L.M.; Murayama, A.; Bajet, N.B.; Hase, S.; Takahashi, H.; Ikegami, M. Genome organization of an infectious clone of Tomato leaf curl virus (Philippines), a new monopartite begomovirus. J. Phytopathol. 2002, 150, 587–591. [Google Scholar] [CrossRef]
- Ueda, S.; Kimura, T.; Onuki, M.; Hanada, K.; Iwanami, T. Three distinct groups of isolates of Tomato yellow leaf curl virus in Japan and construction of an infectious clone. J. Gen. Plant Pathol. 2004, 70, 232–238. [Google Scholar] [CrossRef]
- Romay, G.; Geraud-Pouey, F.; Chirinos, D.T.; Mahillon, M.; Gillis, A.; Mahillon, J.; Bragard, C. Tomato Twisted Leaf Virus: A Novel Indigenous New World Monopartite Begomovirus Infecting Tomato in Venezuela. Viruses 2019, 11, 327. [Google Scholar] [CrossRef]
- Davino, S.; Napoli, C.; Dellacroce, C.; Miozzi, L.; Noris, E.; Davino, M.; Accotto, G.P. Two new natural begomovirus recombinants associated with the tomato yellow leaf curl disease co-exist with parental viruses in tomato epidemics in Italy. Virus Res. 2009, 143, 15–23. [Google Scholar] [CrossRef]
- Heydarnejad, J.; Kamali, M.; Hassanvand, V.; Massumi, H.; Shamshiri, M.; Varsani, A. Turnip leaf curl disease associated with two begomoviruses in south-eastern Iran. Trop. Plant Pathol. 2018, 43, 165–169. [Google Scholar] [CrossRef]
- Just, K.; Leke, W.N.; Sattar, M.N.; Luik, A.; Kvarnheden, A. Detection of tomato yellow leaf curl virus in imported tomato fruit in northern europe. Plant Pathol. 2014, 63, 1454–1460. [Google Scholar] [CrossRef]
- Salati, R.; Nahkla, M.K.; Rojas, M.R.; Guzman, P.; Jaquez, J.; Maxwell, D.P.; Gilbertson, R.L. Tomato yellow leaf curl virus in the Dominican Republic: Characterization of an Infectious Clone, Virus Monitoring in Whiteflies, and Identification of Reservoir Hosts. Phytopathology 2002, 92, 487–496. [Google Scholar] [CrossRef]
- Urbino, C.; Thébaud, G.; Granier, M.; Blanc, S.; Peterschmitt, M. A novel cloning strategy for isolating, genotyping and phenotyping genetic variants of geminiviruses. Virol. J. 2008, 5, 135. [Google Scholar] [CrossRef]
- García-Andrés, S.; Monci, F.; Navas-Castillo, J.; Moriones, E. Begomovirus genetic diversity in the native plant reservoir Solanum nigrum: Evidence for the presence of a new virus species of recombinant nature. Virology 2006, 350, 433–442. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, H.; Zhou, X. Molecular characterization and pathogenicity of tomato yellow leaf curl virus in China. Virus Genes 2009, 39, 249–255. [Google Scholar] [CrossRef]
- Packialakshmi, R.M.; Usha, R. A simple and efficient method for agroinfection of Vernonia cinerea with infectious clones of Vernonia yellow vein virus. Virus Genes 2011, 43, 465–470. [Google Scholar] [CrossRef]
- Garrido-Ramirez, E.R.; Sudarshana, M.R.; Gilbertson, R.L. Bean golden yellow mosaic virus from Chiapas, Mexico: Characterization, Pseudorecombination with Other Bean-Infecting Geminiviruses and Germ Plasm Screening. Phytopathology 2000, 90, 1224–1232. [Google Scholar] [CrossRef]
- Li, J.; Zhou, X. Molecular characterization and experimental host-range of two begomoviruses infecting Clerodendrum cyrtophyllum in China. Virus Genes 2010, 41, 250–259. [Google Scholar] [CrossRef]
- Blawid, R.; Fontenele, R.S.; Lacorte, C.; Ribeiro, S.G. Molecular and biological characterization of corchorus mottle virus, a new begomovirus from Brazil. Arch. Virol. 2013, 158, 2603–2609. [Google Scholar] [CrossRef][Green Version]
- Sharma, D.; Kulshreshtha, A.; Roshan, P.; Hallan, V. Molecular characterization and infectivity analysis of a bipartite begomovirus associated with cotton leaf curl Multan betasatellite naturally infecting Rumex nepalensis in northern India. J. Plant Pathol. 2019, 101, 935–941. [Google Scholar] [CrossRef]
- Jailani, A.; Kumar, A.; Mandal, B.; Sivasudha, T.; Roy, A. Agroinfection of tobacco by croton yellow vein mosaic virus and designing of a replicon vector for expression of foreign gene in plant. Virus Dis. 2016, 27, 277–286. [Google Scholar] [CrossRef]
- Hagen, C.; Rojas, M.R.; Sudarshana, M.R.; Xoconostle-Cazares, B.; Natwick, E.T.; Turini, T.A.; Gilbertson, R.L. Biology and Molecular Characterization of Cucurbit leaf crumple virus, an Emergent Cucurbit-Infecting Begomovirus in the Imperial Valley of California. Plant Dis. 2008, 92, 781–793. [Google Scholar] [CrossRef]
- Ndunguru, J.; Legg, J.; Fofana, B.; Aveling, T.; Thompson, G.; Fauquet, C. Identification of a defective molecule derived from DNA-A of the bipartite begomovirus of East African Cassava Mosaic Virus. Plant Pathol. 2006, 55, 2–10. [Google Scholar] [CrossRef]
- Gao, S.; Qu, J.; Chua, N.H.; Ye, J. A new strain of Indian cassava mosaic virus causes a mosaic disease in the biodiesel crop Jatropha curcas. Arch. Virol. 2010, 155, 607–612. [Google Scholar] [CrossRef]
- Polston, J.E.; Londoño, M.A.; Capobianco, H. The complete genome sequence of New World jatropha mosaic virus. Arch. Virol. 2014, 159, 3131–3136. [Google Scholar] [CrossRef]
- Ilyas, M.; Qazi, J.; Mansoor, S.; Briddon, R.W. Molecular characterisation and infectivity of a "Legumovirus" (genus Begomovirus: Family Geminiviridae) infecting the leguminous weed Rhynchosia minima in Pakistan. Virus Res. 2009, 145, 279–284. [Google Scholar] [CrossRef]
- Kumar, S.; Tanti, B.; Patil, B.L.; Mukherjee, S.K.; Sahoo, L. RNAi-derived transgenic resistance to Mungbean yellow mosaic India virus in cowpea. PLoS ONE 2017, 12, e0186786. [Google Scholar] [CrossRef]
- Haq, Q.M.; Rouhibakhsh, A.; Ali, A.; Malathi, V.G. Infectivity analysis of a blackgram isolate of Mungbean yellow mosaic virus and genetic assortment with MYMIV in selective hosts. Virus Genes 2011, 42, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Shakir, S.; Nawaz-ul-Rehman, M.S.; Mubin, M.; Ali, Z. Characterization, phylogeny and recombination analysis of Pedilanthus leaf curl virus-Petunia isolate and its associated betasatellite. Virol. J. 2018, 15. [Google Scholar] [CrossRef] [PubMed]
- Koeda, S.; Homma, K.; Tanaka, Y.; Onizaki, D.; Kesumawati, E.; Zakaria, S. Inoculation of capsicums with pepper yellow leaf curl indonesia virus by combining agroinoculation and grafting. Hortic. J. 2018, 87, 364–371. [Google Scholar] [CrossRef]
- Urbino, C.; Polston, J.E.; Patte, C.P.; Caruana, M.L. Characterization and genetic diversity of potato yellow mosaic virus from the Caribbean. Arch. Virol. 2004, 149, 417–424. [Google Scholar] [CrossRef]
- Al-Aqeel, H.A.; Iqbal, Z.; Polston, J.E. Characterization of sida golden mottle virus isolated from Sida santaremensis Monteiro in Florida. Arch. Virol. 2018, 163, 2907–2911. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Tovar, R.; Navas-Castillo, J. Deciphering the biology of deltasatellites from the New World: Maintenance by New World begomoviruses and whitefly transmission. New Phytol. 2016, 212, 680–692. [Google Scholar] [CrossRef]
- Kon, T.; Dolores, L.M.; Bajet, N.B.; Hase, S.; Takahashi, H.; Ikegami, M. Molecular characterisation of a strain of Squash leaf curl China virus from the Philippines. J. Phytopathol. 2003, 151, 535–539. [Google Scholar] [CrossRef]
- Ferreira, P.T.O.; Lemos, T.O.; Nagata, T.; Inoue-Nagata, A.K. One-step cloning approach for construction of agroinfectious begomovirus clones. J. Virol. Methods 2008, 147, 351–354. [Google Scholar] [CrossRef]
- Malik, A.H.; Briddon, R.W.; Mansoor, S. Infectious clones of Tomato leaf curl Palampur virus with a defective DNA B and their pseudo-recombination with Tomato leaf curl New Delhi virus. Virol. J. 2011, 8. [Google Scholar] [CrossRef]
- Guo, W.; Yang, X.; Xie, Y.; Cui, X.; Zhou, X. Tomato yellow leaf curl Thailand virus-[Y72] from Yunnan is a monopartite begomovirus associated with DNAbeta. Virus Genes 2009, 38, 328–333. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Hernández-Zepeda, C.; Trejo-Saavedra, D.; Carillo-Tripp, J.; Rivera-Bustamante, R.F.; Martinez-Zubiaur, Y. Complete genome and pathogenicity of Tomato yellow leaf distortion virus, a bipartite begomovirus infecting tomato in Cuba. Eur. J. Plant Pathol. 2012, 134, 13–21. [Google Scholar] [CrossRef]
- Zaim, M.; Kumar, Y.; Hallan, V.; Zaidi, A.A. Velvet bean severe mosaic virus: A distinct begomovirus species causing severe mosaic in Mucuna pruriens (L.) DC. Virus Genes 2011, 43, 138–146. [Google Scholar] [CrossRef]
- Al-Musa, A.; Anfoka, G.; Al-Abdulat, A.; Misbeh, S.; Haj Ahmed, F.; Otri, I. Watermelon chlorotic stunt virus (WmCSV): A serious disease threatening watermelon production in Jordan. Virus Genes 2011, 43, 79–89. [Google Scholar] [CrossRef]
- Collins, A.M.; Mujaddad-ur-Rehman, M.; Brown, J.K.; Reddy, C.; Wang, A.; Fondong, V.; Roye, M.E. Molecular characterization and experimental host range of an isolate of Wissadula golden mosaic St. Thomas virus. Virus Genes 2009, 39, 387–395. [Google Scholar] [CrossRef]
- Boulton, M.I. Construction of infectious clones for DNA viruses: Mastreviruses. Methods Mol. Biol. 2008, 451, 503–523. [Google Scholar] [CrossRef]
- Bunawan, H.; Dusik, L.; Bunawan, S.N.; Amin, N.M. Rice Tungro Disease: From Identification to Disease Control. World Appl. Sci. J. 2014, 31, 1221–1226. [Google Scholar] [CrossRef]
- Biswas, K.K.; Varma, A. Agroinoculation: A method of screening germplasm resistance to mungbean yellow mosaic Geminivirus. Indian Phytopathol. 2001, 54, 240–245. [Google Scholar]
- Picó, B.; Díez, M.J.; Nuez, F. Viral diseases causing the greatest economic losses to the tomato crop. II. The Tomato yellow leaf curl virus—A review. Sci. Hortic. 1996, 67, 151–196. [Google Scholar] [CrossRef]
- Brewer, H.C.; Hird, D.L.; Bailey, A.M.; Seal, S.E.; Foster, G.D. A guide to the contained use of plant virus infectious clones. Plant Biotechnol. J. 2018, 16, 832–843. [Google Scholar] [CrossRef]
- Lange, M.; Yellina, A.L.; Orashakova, S.; Becker, A. Virus-induced gene silencing (VIGS) in plants: An overview of target species and the virus-derived vector systems. Methods Mol. Biol. 2013, 975, 1–14. [Google Scholar] [CrossRef]
- Lee, W.S.; Hammond-Kosack, K.E.; Kanyuka, K. Barley stripe mosaic virus-mediated tools for investigating gene function in cereal plants and their pathogens: Virus-induced gene silencing, host-mediated gene silencing, and virus-mediated overexpression of heterologous protein. Plant Physiol. 2012, 160, 582–590. [Google Scholar] [CrossRef]
- Moury, B.; Morel, C.; Johansen, E.; Guilbaud, L.; Souche, S.; Ayme, V.; Caranta, C.; Palloix, A.; Jacquemond, M. Mutations in potato virus Y genome-linked protein determine virulence toward recessive resistances in Capsicum annuum and Lycopersicon hirsutum. Mol. Plant-Microbe Interact. Mpmi 2004, 17, 322–329. [Google Scholar] [CrossRef]
- Roshan, P.; Kulshreshtha, A.; Kumar, S.; Purohit, R.; Hallan, V. AV2 protein of tomato leaf curl Palampur virus promotes systemic necrosis in Nicotiana benthamiana and interacts with host Catalase2. Sci. Rep. 2018, 8, 1273. [Google Scholar] [CrossRef]
- Saeed, F.; Naeem, M.; Hameed, U.; Ilyas, M.; Saleem, M.; Hamza, M.; Mansoor, S.; Amin, I. Infectivity of okra enation leaf curl virus and the role of its V2 protein in pathogenicity. Virus Res. 2018, 255, 90–94. [Google Scholar] [CrossRef]
- Hipp, K.; Rau, P.; Schäfer, B.; Gronenborn, B.; Jeske, H. The RXL motif of the African cassava mosaic virus Rep protein is necessary for rereplication of yeast DNA and viral infection in plants. Virology 2014, 462–463, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J. Analysis of African cassava mosaic virus recombinants suggests strand nicking occurs within the conserved nonanucleotide motif during the initiation of rolling circle DNA replication. Virology 1995, 206, 707–712. [Google Scholar] [CrossRef]
- Chandran, S.A.; Jeyabharathy, C.; Usha, R. The C2 protein of Bhendi yellow vein mosaic virus plays an important role in symptom determination and virus replication. Virus Genes 2014, 48, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.; Latham, J.R.; Pinner, M.S.; Bedford, I.; Markham, P.G. Mutational analysis of the monopartite geminivirus beet curly top virus. Virology 1992, 191, 396–405. [Google Scholar] [CrossRef]
- Baliji, S.; Sunter, J.; Sunter, G. Transcriptional analysis of complementary sense genes in Spinach curly top virus and functional role of C2 in pathogenesis. Mol. Plant-Microbe Interact. Mpmi 2007, 20, 194–206. [Google Scholar] [CrossRef]
- Iqbal, Z.; Sattar, M.N.; Kvarnheden, A.; Mansoor, S.; Briddon, R.W. Effects of the mutation of selected genes of Cotton leaf curl Kokhran virus on infectivity, symptoms and the maintenance of Cotton leaf curl Multan betasatellite. Virus Res. 2012, 169, 107–116. [Google Scholar] [CrossRef]
- Jupin, I.; De Kouchkovsky, F.; Jouanneau, F.; Gronenborn, B. Movement of tomato yellow leaf curl geminivirus (TYLCV): Involvement of the protein encoded by ORF C4. Virology 1994, 204, 82–90. [Google Scholar] [CrossRef]
- Piroux, N.; Saunders, K.; Page, A.; Stanley, J. Geminivirus pathogenicity protein C4 interacts with Arabidopsis thaliana shaggy-related protein kinase AtSKeta, a component of the brassinosteroid signalling pathway. Virology 2007, 362, 428–440. [Google Scholar] [CrossRef]
- Rojas, M.R.; Jiang, H.; Salati, R.; Xoconostle-Cázares, B.; Sudarshana, M.R.; Lucas, W.J.; Gilbertson, R.L. Functional analysis of proteins involved in movement of the monopartite begomovirus, Tomato yellow leaf curl virus. Virology 2001, 291, 110–125. [Google Scholar] [CrossRef]
- Etessami, P.; Saunders, K.; Watts, J.; Stanley, J. Mutational analysis of complementary-sense genes of African cassava mosaic virus DNA A. J. Gen. Virol. 1991, 72 Pt 5, 1005–1012. [Google Scholar] [CrossRef]
- Hong, Y.; Stanley, J. Regulation of African cassava mosaic virus complementary-sense gene expression by N-terminal sequences of the replication-associated protein AC1. J. Gen. Virol. 1995, 76 Pt 10, 2415–2422. [Google Scholar] [CrossRef]
- Pooma, W.; Petty, I.T. Tomato golden mosaic virus open reading frame AL4 is genetically distinct from its C4 analogue in monopartite geminiviruses. J. Gen. Virol. 1996, 77 Pt 8, 1947–1951. [Google Scholar] [CrossRef]
- Chellappan, P.; Vanitharani, R.; Fauquet, C.M. MicroRNA-binding viral protein interferes with Arabidopsis development. Proc. Natl. Acad. Sci. USA 2005, 102, 10381–10386. [Google Scholar] [CrossRef]
- Vanitharani, R.; Chellappan, P.; Pita, J.S.; Fauquet, C.M. Differential roles of AC2 and AC4 of cassava geminiviruses in mediating synergism and suppression of posttranscriptional gene silencing. J. Virol. 2004, 78, 9487–9498. [Google Scholar] [CrossRef]
- Mansoor, S.; Zafar, Y.; Briddon, R.W. Geminivirus disease complexes: The threat is spreading. Trends Plant Sci. 2006, 11, 209–212. [Google Scholar] [CrossRef]
- Jose, J.; Usha, R. Bhendi yellow vein mosaic disease in India is caused by association of a DNA Beta satellite with a begomovirus. Virology 2003, 305, 310–317. [Google Scholar] [CrossRef]
- Cui, X.; Li, G.; Wang, D.; Hu, D.; Zhou, X. A Begomovirus DNAbeta-encoded protein binds DNA, functions as a suppressor of RNA silencing, and targets the cell nucleus. J. Virol. 2005, 79, 10764–10775. [Google Scholar] [CrossRef]
- Yang, X.; Guo, W.; Ma, X.; An, Q.; Zhou, X. Molecular characterization of Tomato leaf curl China virus, infecting tomato plants in China, and functional analyses of its associated betasatellite. Appl. Environ. Microbiol. 2011, 77, 3092–3101. [Google Scholar] [CrossRef]
- Ariyo, O.A.; Atiri, G.I.; Dixon, A.G.; Winter, S. The use of biolistic inoculation of cassava mosaic begomoviruses in screening cassava for resistance to cassava mosaic disease. J. Virol. Methods 2006, 137, 43–50. [Google Scholar] [CrossRef]
- Briddon, R.W.; Liu, S.; Pinner, M.S.; Markham, P.G. Infectivity of African cassava mosaic virus clones to cassava by biolistic inoculation. Arch. Virol. 1998, 143, 2487–2492. [Google Scholar] [CrossRef]
- Berrie, L.C.; Rybicki, E.P.; Rey, M. Complete nucleotide sequence and host range of South African cassava mosaic virus: Further evidence for recombination amongst begomoviruses. J. Gen. Virol. 2001, 82 Pt 1, 53–58. [Google Scholar] [CrossRef]
- Ariyo, O.A.; Dixon, A.G.; Atiri, G.I. Whitefly Bemisia tabaci (Homoptera: Aleyrodidae) infestation on cassava genotypes grown at different ecozones in Nigeria. J. Econ. Entomol. 2005, 98, 611–617. [Google Scholar] [CrossRef]
- Lapidot, M.; Friedmann, M. Breeding for resistance to whitefly-transmitted geminviruses. Ann. Appl. Biol. 2005, 140, 109–127. [Google Scholar] [CrossRef]
- Ayeh, K.O.; Ramsell, J.N.E. The use of biolistic inoculation of cassava mosaic begomoviruses in screening cassava cultivars from Ghana for resistance/susceptibility to cassava mosaic disease. Afr. J. Biotechnol. 2008, 7, 2353–2358. [Google Scholar]
- Ferro, M.M.M.; Ramos-Sobrinho, R.; Xavier, C.A.D.; Zerbini, F.M.; Lima, G.S.A.; Nagata, T.; Assunção, I.P. New approach for the construction of infectious clones of a circular DNA plant virus using Gibson Assembly. J. Virol. Methods 2019, 263, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B. Proof by synthesis of Tobacco mosaic virus. Genome Biol. 2014, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Lovato, A.; Faoro, F.; Gambino, G.; Maffi, D.; Bracale, M.; Polverari, A.; Santi, L. Construction of a synthetic infectious cDNA clone of Grapevine Algerian latent virus (GALV-Nf) and its biological activity in Nicotiana benthamiana and grapevine plants. Virol. J. 2014, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Pasin, F.; Menzel, W.; Daròs, J.-A. Harnessed viruses in the age of metagenomics and synthetic biology: An update on infectious clone assembly and biotechnologies of plant viruses. Plant Biotechnol. J. 2019, 17, 1010–1026. [Google Scholar] [CrossRef] [PubMed]
- Bornancini, V.A.; Irazoqui, J.M.; Flores, C.R.; Vaghi Medina, C.G.; Amadio, A.F.; López Lambertini, P.M. Reconstruction and characterization of full-length begomovirus and alphasatellite genomes infecting pepper through metagenomics. Viruses 2020, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Fakhfakh, H.; Vilaine, F.; Makni, M.; Robaglia, C. Cell-free cloning and biolistic inoculation of an infectious cDNA of potato virus Y. J. Gen. Virol. 1996, 77, 519–523. [Google Scholar] [CrossRef]
- Jailani, A.A.K.; Shilpi, S.; Mandal, B. Rapid demonstration of infectivity of a hybrid strain of potato virus Y occurring in India through overlapping extension PCR. Physiol. Mol. Plant Pathol. 2017, 98, 62–68. [Google Scholar] [CrossRef]
- Youssef, F.; Marais, A.; Faure, C.; Gentit, P.; Candresse, T. Strategies to facilitate the development of uncloned or cloned infectious full-length viral cDNAs: Apple chlorotic leaf spot virus as a case study. Virol. J. 2011, 8. [Google Scholar] [CrossRef]
- Roy, A.; Zhai, Y.; Ortiz, J.; Neff, M.; Mandal, B.; Mukherjee, S.K.; Pappu, H.R. Multiplexed editing of a begomovirus genome restricts escape mutant formation and disease development. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Khan, S.; Mahmood, M.S.; Ur Rahman, S.; Rizvi, F.; Ahmad, A. Evaluation of the CRISPR/Cas9 system for the development of resistance against Cotton leaf curl virus in model plants. Plant Protect. Sci. 2020, 56, 154–162. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).