
Luteogenesis and Embryo Implantation Are Enhanced by Exogenous hCG in Goats Subjected to an Out-of-Season Fixed-Time Artificial Insemination Protocol


 ,
,  ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
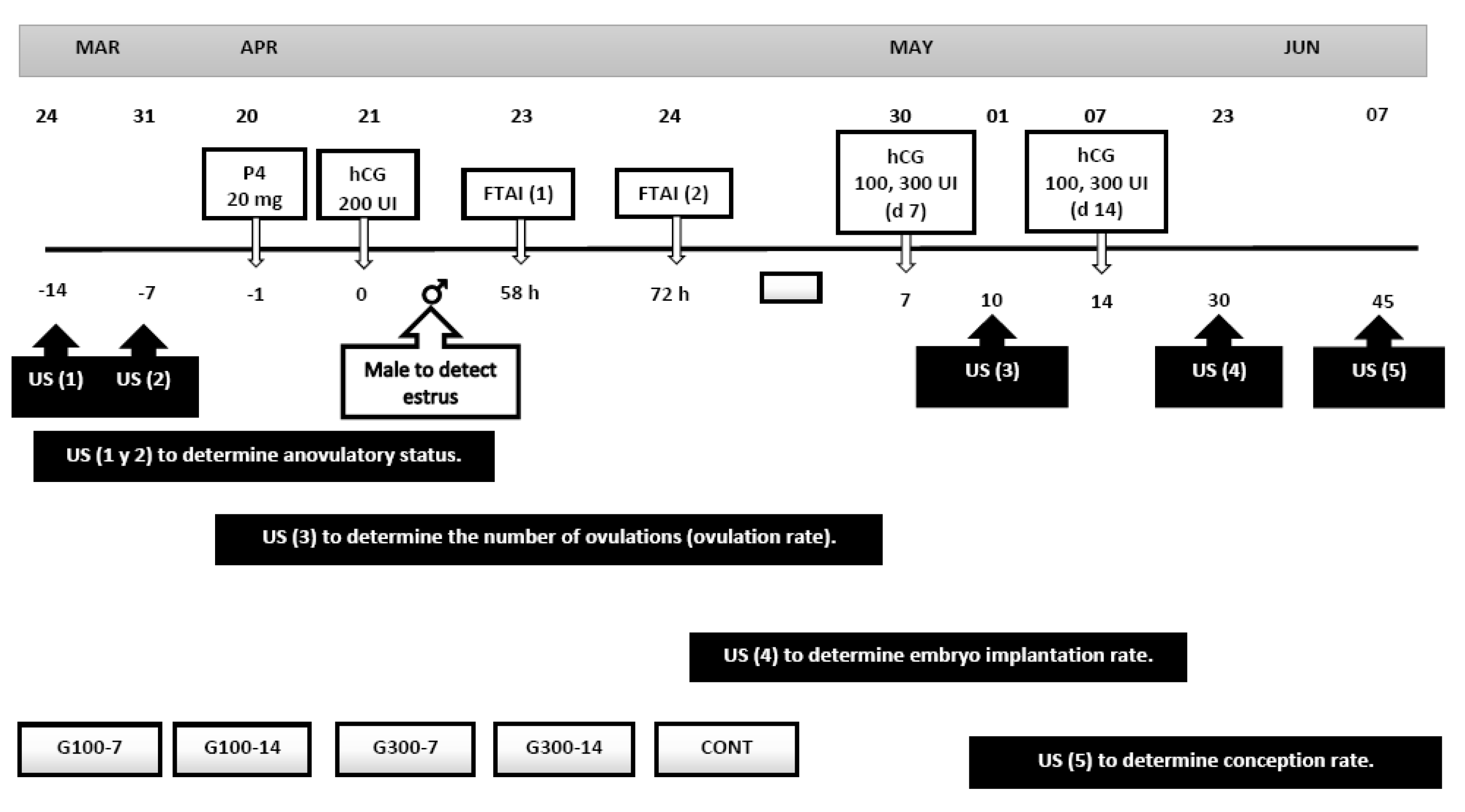
2. Materials and Methods
2.1. General
2.2. Location, and Environmental Conditions
2.3. Animals and Their Management
2.3.1. Female Goats: Confirmation of the Anovulatory Status, Estrus Induction Protocol, and Estrus Detection with Aproned Males
2.3.2. Male Goats: Setting Bucks for Semen Extraction and Semen Quality Assessment
2.4. Fixed-Time Artificial Insemination (FTAI)
2.5. Conformation of the Experimental Groups: The Use of hCG Post-FTAI to Enhance the Embryo Implantation Rate
2.6. Response Variables
2.6.1. Body Weight, Body Condition Score, and Estrus Induction
2.6.2. Ovulation Percentage, Ovulatory Rate, and Luteogenesis
2.6.3. Embryo Implantation Rate, and Other Reproductive Variables
2.7. Statistical Analyses
3. Results
3.1. Body Weight, Body Condition Score, and Estrus Induction
3.2. Conception Rate, Fertility Rate, Prolificacy Rate, and Fecundity Rate
3.3. Ovulation Rate, Corpus Luteum Area and Diameter, Embryo Number, Embryo Implantation Rate, and Embryo Efficiency Indexes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castel, J.M.; Ruiz, F.A.; Mena, Y.; Sánchez-Rodríguez, M. Present situation and future perspectives for goat production systems in Spain. Small Rumin. Res. 2010, 89, 207–210. [Google Scholar] [CrossRef]
- Escareño, L.; Wurzinger, M.; Iñiguez, L.; Soelkner, J.; Salinas, H.; Meza-Herrera, C.A. Dairy goat production systems in dry areas: Status-quo, perspectives and challenges. Trop. Anim. Health Prod. 2013, 45, 17–34. [Google Scholar] [CrossRef]
- SIAP (Sistema de Información Agroalimentaria y Pesquera). Resumen Nacional. Población Ganadera, Avícola Y Apícola. SAGARPA. 2019. Available online: www.siap.gob.mx/ganaderia (accessed on 15 February 2020).
- Isidro-Requejo, L.M.; Meza-Herrera, C.; Pastor-López, F.J.; Maldonado, J.A.; Salinas-Gonzalez, H. Physicochemical characterization of goat milk produced in the Comarca Lagunera, Mexico. Anim. Sci. J. 2019, 90, 563–573. [Google Scholar] [CrossRef]
- Navarrete-Molina, C.; Meza-Herrera, C.; Herrera-Machuca, M.; Macias-Cruz, U.; Veliz-Deras, F. Not all ruminants were created equal: Environmental and socio-economic sustainability of goats under a marginalextensive production system. J. Clean. Prod. 2020, 255, 120237. [Google Scholar] [CrossRef]
- Álvarez, L.; Zarco, L. Los fenómenos de bioestimulación sexual en ovejas y cabras. Vet Méx. 2001, 32, 117–129. [Google Scholar]
- Meza-Herrera, C.A.; Serradilla, J.M.; Muñoz-Mejias, M.E.; Baena-Manzano, F.; Menendez-Buxadera, A. Effect of breed and some environmental factors on body weights till weaning and litter size in five goat breeds in Mexico. Small Rum. Res. 2014, 121, 215–219. [Google Scholar] [CrossRef]
- Meza-Herrera, C.A.; Santamaría-Estrada, C.E.; Flores-Hernández, A.; Cano-Villegas, O.; De la Peña, C.G.; Macias-Cruz, U.; Calderón-Leyva, G.; Angel-García, O.; Mellado, M.; Carrillo-Moreno, D. The Opuntia Effect upon the out-of-season embryo implantation rate in goats: Corpus luteal number, corpus luteal diameter and serum progesterone concentrations. Livest. Sci. 2019, 228, 201–206. [Google Scholar] [CrossRef]
- Carrillo, E.; Meza-Herrera, C.A.; Véliz, F.G. Estacionalidad reproductiva de los machos cabríos de la raza Alpino-Francés adaptados al subtrópico Mexicano. Rev. Mex. de Cien. Pec. 2010, 1, 169–178. [Google Scholar]
- Contreras-Villarreal, V.; Meza-Herrera, C.A.; Rivas-Muñoz, R.; Angel-Garcia, O.; Luna-Orozco, J.R.; Carrillo, E.; Mellado, M.; Veliz-Deras, F.G. Reproductive performance of seasonally anovular mixed-bred dairy goats induced to ovulate with a combination of progesterone and eCG or estradiol. Anim. Sci. J. 2015, 87, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Espino, A.S.; Meza-Herrera, C.A.; Carrillo, E.; González-Álvarez, V.H.; Guillen-Muñoz, J.M.; Ángel-García, O.; Mellado, M.; Véliz-Deras, F.G. Reproductive outcomes of Alpine goats primed with progesterone and treated with human chorionic gonadotropin during the anestrus-to-estrus transition season. Anim. Reprod. Sci. 2016, 167, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, R.; Meza-Herrera, C.A.; Tapia-Robles, K.I.; Alvarado-Espino, A.S.; Luna-Orozco, J.R.; Leyva, C. Effect of two routes of administration of human chorionic gonadotropin upon oestrus induction and reproductive outcomes in adult acyclic mix-breed goats. J. Appl. Anim. Res. 2017, 46, 190–194. [Google Scholar] [CrossRef]
- Alvarado-Espino, A.S.; Menchaca, A.; Meza-Herrera, C.A.; Mellado, M.; Arellano, F.; Véliz, F. Use of injectable progesterone and hCG for fixed-time artificial insemination during the non-breeding season in goats. Theriogenology 2019, 127, 21–25. [Google Scholar] [CrossRef]
- Alvarado-Espino, A.S.; Menchaca, A.; Meza-Herrera, C.A.; Carrillo-Moreno, D.I.; Zúñiga-García, S.; Arellano-Rodríguez, F.; Mellado, M.; Véliz, F.G. Ovarian response is not affected by the stage of seasonal anestrus or breed of goats when using a progesterone injection plus human chorionic gonadotropin-based protocol. Anim. Reprod. Sci. 2019, 204, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Kawate, N.; Yamazaki, M.; Tamada, H.; Inaba, T.; Sawada, T. Effect of low dose of hCG on induction of fertile estrus in shiba goats pretreated intravaginally with progesterone during the early postpartum nursing period. J. Reprod. Dev. 2002, 48, 497–504. [Google Scholar] [CrossRef][Green Version]
- Catalano, R.; Teruel, M.; Gonzalez, C.; Williams, S.; Videla Dorna, I.; Callejas, S. The effect of hCG treatment on follicular characteristics and luteal function in seasonally anestrous ewes lambs. J. Anim. Vet. Adv. 2012, 11, 2091–2096. [Google Scholar] [CrossRef]
- Bazer, F.W.; Wu, G.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C.; Bayless, K. Novel pathways for implantation and establishment and maintenance of pregnancy in mammals. Mol. Hum. Reprod. 2010, 16, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Thatcher, W.W. Chronicling the discovery of interferon tau. Reproduction 2017, 154, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Burghardt, R.C.; Johnson, G.A.; Bazer, F.W. Conceptus signals for establishment and maintenance of pregnancy. Anim. Reprod. Sci. 2004, 82, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C. Implantation mechanisms: Insights from the sheep. Reproduction 2004, 128, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Vera, A.H.R.; Urrutia, M.J.; Espinosa, M.M.A.; Estrada, C.E.; Jiménez, S.H. Nutrición, estacionalidad reproductiva y mantenimiento de la gestación en caprinos. In Libro técnico Núm; INIFAP-CENID Fisiología y Mejoramiento Animal: Ajuchitlán, Querétaro, Mexico, 2013. [Google Scholar]
- Khan, T.; Beck, N.; Khalid, M. The effect of hCG treatment on day 12 post-mating on ovarian function and reproductive performance of ewes and ewe lambs. Anim. Reprod. Sci. 2009, 116, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Brooks, K.; Burns, G.; Spencer, T.E. Conceptus elongation in ruminants: Roles of progesterone, prostaglandin, interferon tau and cortisol. J. Anim. Sci. Biotechnol. 2014, 16, 50–53. [Google Scholar] [CrossRef]
- Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Spencer, T.E.; Wu, G. Mechanisms for the establishment and maintenance of pregnancy: Synergies from scientific collaborations. Biol. Reprod. 2018, 99, 225–241. [Google Scholar] [CrossRef]
- Mellado, M.; Meza-Herrera, C. Influence of season and environment on fertility of goats in a hot-arid environment. J. Agric. Sci. 2002, 138, 97–102. [Google Scholar] [CrossRef]
- Mellado, M.; Pastor, F. Aborto no infeccioso en caprinos. Ciênc. Anim. Bras. 2006, 7, 167–175. [Google Scholar]
- FASS. Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching, 3rd ed.; Federation Animal Science Society: Champaing, IL, USA, 2010; p. 177. [Google Scholar]
- NAM-National Academy of Medicine. Guide for the Care and Use of Laboratory Animals, 1st ed.; National Academy of Medicine-Mexico and the Association for Assessment and Accreditation of Laboratory Animal Care International: Harlan, Mexico, 2010. [Google Scholar]
- INEGI (Instituto Nacional de Estadística y Geografía). Información Nacional por Entidad Federativa y Municipios. Available online: http://www.inegi.org.mx/sistemas/mexicocifras/default.aspx (accessed on 19 February 2015).
- INIFAP (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias). Boletín técnico. Coeficientes de agostadero de la República Mexicana: Estados de Baja California, Sonora, Chihuahua, Zacatecas, Coahuila, Tamaulipas, Nuevo León, Durango y San Luis Potosí. 2: 13-78 SAG. México. Available online: http://www.gob.mx/inifap/archivo/documentos/coeficientesdeagostader (accessed on 1 March 2010).
- National Research Council (NRC). Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Martínez, A.; Vega-Pla, J.; León, J.; Camacho, M.; Delgado, J.; Ribeiro, M. Is the Murciano-Granadina a single goat breed? A molecular genetics approach. Arq. Bras. Med. Vet. Zootec. 2010, 62, 1191–1198. [Google Scholar] [CrossRef]
- Montaldo, H.H.; Meza-Herrera, C.A. Genetic goat resources in Mexico: Bio-economical efficiency of local and specialized genotypes. Wool Technol. Sheep Breed. 1999, 47, 184–198. [Google Scholar]
- León, J.; Quiroz, J.; Pleguezuelos, J.; Camacho, E.; Delgado, J. Esquema de selección para preservar la biodiversidad intraracial de la cabra Murciano-Granadina. Arch. Zootec. 2007, 56, 661–662. [Google Scholar]
- Luna-Orozco, J.R.; Guillen-Muñoz, J.M.; De Santiago-Miramontes, M.d.l.A.; García, J.E.; Rodríguez-Martínez, R.; Meza-Herrera, C.A.; Mellado, M.; Véliz, F.G. Influence of sexually inactive bucks subjected to long photoperiod or testosterone on the induction of estrus in anovulatory goats. Trop. Anim. Health Prod. 2012, 44, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Membrillo-Ortega, A.; Córdova-Izquierdo, A.; Hicks-Gómez, J.J.; Valencia-Méndez, J.J.; Castillo-Juárez, H. Efecto de la adición de antioxidantes en el diluyente de semen de macho cabrío antes de congelar y después de descongelar. Rev. Vet. 2011, 22, 85–90. [Google Scholar] [CrossRef]
- Walkden-Brown, S.; Restall, B.; Scaramuzzi, R.; Martin, G.; Blackberry, M. Seasonality in male Australian cashmere goats: Long term effects of castration and testosterone or oestradiol treatment on changes in LH, FSH and prolactin concentrations, and body growth. Small Rumin. Res. 1997, 26, 239–252. [Google Scholar] [CrossRef]
- Giordano, J.O.; Fricke, P.M.; Guenther, J.N.; Lopes, G.; Herlihy, M.M.; Nascimento, A.B. Effect of progesterone on magnitude of the luteinizing hormone surge induced by two different doses of gonadotropin-releasing hormone in lactating dairy cows. J. Dairy Sci. 2012, 95, 3781–3793. [Google Scholar] [CrossRef] [PubMed]
- Mellado, M.; Valdez, R.; Lara, L.M.; García, J.E. Risk factors affecting conception, abortion and kidding rates of goats under extensive conditions. Small Rum. Res. 2004, 55, 191–198. [Google Scholar] [CrossRef]
- Arrebola, F.A.; Pardo, B.; Sanchez, M.; Lopez, M.D.; Perez-Marin, C.C. Factors influencing the success of an artificial insemination program in Florida goats. Span. J. Agric. Res. 2012, 10, 338. [Google Scholar] [CrossRef]
- Fernandez, J.; Galarraga, M.M.B.; Soto, A.T.; Sota, R.L.; Cueto, M.I.; Lacau-Mengido, I.M. Effect of GnRH or hCG administration on Day 4 post insemination on reproductive performance in Merino sheep of North Patagonia. Theriogenology 2019, 126, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Raheem, K.A. An insight into maternal recognition of pregnancy in mammalian species. J. Saudi Soc. Agric. Sci. 2015, 16, 1–6. [Google Scholar] [CrossRef]
- Lonergan, P.; Forde, N.; Spencer, T. Role of progesterone in embryo development in cattle. Reprod. Fertil. Dev. 2016, 28, 66–74. [Google Scholar] [CrossRef]
- Lonergan, P.; Forde, N. Maternal-embryo interaction leading up to the initiation of implantation of pregnancy in cattle. Animal 2014, 8, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Meza-Herrera, C.A.; Tena-Sempere, M. Interface between Nutrition and Reproduction. In Animal Reproduction in Livestock—Encyclopedia of Life Support Systems; Astiz, S., Gonzalez, A., Eds.; EOLSS Publishers: Oxford, UK, 2012. [Google Scholar]
- Rostami, B.; Hajizadeh, R.; Shahir, M.; Aliyari, D. The effect of post-mating hCG or progesterone administration on reproductive performance of Afshariv-Booroola-Merino crossbred ewes. Trop. Anim. Health. Prod. 2017, 49, 245–250. [Google Scholar]
- Khan, T.H.; Beck, N.F.G.; Khalid, M. The effects of GnRH analogue (buserelin) or hCG (Chorulon) on Day 12 of pregnancy on ovarian function, plasma hormone concentrations, conceptus growth and placentation in ewes and Ewe lambs. Anim. Reprod. Sci. 2007, 102, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Côrtes, L.R.; Souza-Fabjan, J.M.G.; Dias, D.S.; Martins, B.B.; Maia, A.L.R.S.; Veiga, M.O.; Arashiro, E.K.N.; Brandão, F.Z.; Oliveira, M.E.F.; Bartlewski, P.M.; et al. Administration of a single dose of 300 IU of human chorionic gonadotropin seven days after the onset of estrus improves pregnancy rate in dairy goats by an unknown mechanism. Dom. Anim. Endo. 2020, 74, 106579. [Google Scholar] [CrossRef]
- Catalano, R.; Teruel, M.; Callejas, S. Human chorionic gonadotrophin and its relationship to luteal function and reproductive performance in ovine. Revision Bibliografica. Vet 2016, 18, 49–58. [Google Scholar]
- Figueira, L.M.; Fonseca, J.F.; Arashiro, E.; Souza-Fabjan, J.; Ribeiro, A.; Oba, E.; Viana, J.; Brandão, F.Z. Colour Doppler ultrasonography as a tool to assess luteal function in Santa Inês ewes. Reprod. Domest. Anim. 2015, 50, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.R.; Sinedino, L.D.P.; Spencer, T.E. Paracrine and endocrine actions of interferon tau (ifnt). Reproduction 2017, 154, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Makrigiannakis, A.; Vrekoussis, T.; Zoumakis, E.; Kalantaridou, S.; Jeschke, U. The Role of hCG in Implantation: A mini-review of molecular and clinical evidence. Int. J. Mol. Sci. 2017, 18, 1305. [Google Scholar] [CrossRef] [PubMed]
- Al-Samawi, K.; Al-Hassan, M.; Migdadi, H.; Ammar, M.; Alghamdi, S. Interferon-stimulated gene 15 and interferon-1 stimulated gene 17 messenger rna-based detection of early pregnancy in Aardi goats in Saudi Arabia. J. Zool. 2021, 53, 1005. [Google Scholar] [CrossRef]
- Roberts, R.M.; Ezashi, T.; Rosenfeld, C.S.; Ealy, A.D.; Kubisch, H.M. Evolution of the interferon tau genes and their promoters, and maternal-trophoblast interactions in control of their expression. Reprod. Suppl. 2003, 61, 239–251. [Google Scholar]
- Ealy, A.D.; Wooldridge, L.K. The evolution of interferon-tau. Reproduction 2017, 154, 1–10. [Google Scholar] [CrossRef] [PubMed][Green Version]


{kind=link}
| Variables | G100-7 (n = 13) | G100-14 (n = 12) | G300-7 (n = 12) | G300-14 (n = 12) | CONT (n = 12) | p Value |
|---|---|---|---|---|---|---|
| BW, (kg) | 45.5 ± 0.80 | 45.8 ± 1.2 | 45.9 ± 0.82 | 45.2 ± 1.50 | 45.6 ± 1.71 | 0.99 |
| BCS, (units) | 1.9 ± 0.09 | 2.0 0.07 | 1.8 ± 0.07 | 1.9 ± 0.11 | 1.8 ± 0.07 | 0.62 |
| EI, (n, %) | 13/14 (92.8) | 12/15 (80.0) | 12/13 (92.3) | 12/13 (92.3) | (12/14) 85.71 | 0.64 |
| Variables | G100-7 (n = 13) | G100-14 (n = 12) | G300-7 (n = 12) | G300-14 (n = 12) | CONT (n = 12) | p Value |
|---|---|---|---|---|---|---|
| CR (n, %) | 5/13 (38.5 ± 0.15) | 5/12 (41.7 ± 0.28) | 3/12 (25 ± 0.22) | 7/12 (58.3 ± 0.20) | 4/12 (33.3± 0.30) | 0.54 |
| FR (n, %) | 3/13 (23.1) | 3/12 (25) | 3/12 (25) | 6/12 (50) | 2/12 (16.7) | 0.71 |
| PR (n) | 1.7 ± 0.29 | 0.6 ± 0.33 | 1.3 ± 0.33 | 1.4 ± 0.21 | 0.5 ± 0.29 | 0.53 |
| FC (n, %) | 5/13 (38) b | 2/12 (17) b | 4/12 (33) b | 8/12 (67) a | 2/12 (17) b | 0.06 |
| Variables | G100-7 (n = 13) | G100-14 (n = 12) | G300-7 (n = 12) | G300-14 (n = 12) | CONT (n = 12) | p Value |
|---|---|---|---|---|---|---|
| OVR n | 0.61 ± 0.17 | 0.75 ± 0.22 | 0.58 ± 0.19 | 1.27 ± 0.17 | 0.58 ± 0.23 | 0.3 |
| CLD (mm) | 9.54 ± 0.48 | 9.91 ± 0.77 | 11.88 ± 0.71 | 11.22 ± 0.47 | 8.21 ± 0.51 | 0.10 |
| CLA (mm2) | 72.73 ± 0.18 b | 79.37 ± 0.22 b | 113.30 ± 0.19 a | 103.04 ± 0.17 a | 45.96 ± 0.19 b | 0.02 |
| EN n | 1.75 ± 0.13 a | 1.40 ± 0.24 | 1.67 ± 0.21 | 1.57 ± 0.20 | 1.50 ± 0.24 | 0.78 |
| EIR n (%) | 8/13 (61.5) | 5/12 (41.7) | 6/12 (50) | 7/12 (58.3) | 4/12 (33.3) | 0.61 |
| EEI1 (%) 1 | 23.68 ± 0.12 b | 17.39 ± 0.24 b | 12.5 ± 0.21 b | 33.99 ± 0.20 a | 11.09 ± 0.24 b | 0.02 |
| EEI2 (%) 2 | 19.00 ± 0.37 b | 9.91 ± 0.20 b | 20.30 ± 0.25 a | 27.94 ± 0.30 a | 5.66 ± 0.18 b | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustamante-Andrade, J.A.; Meza-Herrera, C.A.; Rodríguez-Martínez, R.; Santos-Jimenez, Z.; Ángel-García, O.; Gaytán-Alemán, L.R.; Gutierrez-Guzman, U.N.; Esquivel-Romo, A.; Véliz-Deras, F.G. Luteogenesis and Embryo Implantation Are Enhanced by Exogenous hCG in Goats Subjected to an Out-of-Season Fixed-Time Artificial Insemination Protocol. Biology 2021, 10, 429. https://doi.org/10.3390/biology10050429
Bustamante-Andrade JA, Meza-Herrera CA, Rodríguez-Martínez R, Santos-Jimenez Z, Ángel-García O, Gaytán-Alemán LR, Gutierrez-Guzman UN, Esquivel-Romo A, Véliz-Deras FG. Luteogenesis and Embryo Implantation Are Enhanced by Exogenous hCG in Goats Subjected to an Out-of-Season Fixed-Time Artificial Insemination Protocol. Biology. 2021; 10(5):429. https://doi.org/10.3390/biology10050429
Chicago/Turabian StyleBustamante-Andrade, Jorge A., César A. Meza-Herrera, Rafael Rodríguez-Martínez, Zurisaday Santos-Jimenez, Oscar Ángel-García, Leticia R. Gaytán-Alemán, Ulises N. Gutierrez-Guzman, Amaury Esquivel-Romo, and Francisco G. Véliz-Deras. 2021. "Luteogenesis and Embryo Implantation Are Enhanced by Exogenous hCG in Goats Subjected to an Out-of-Season Fixed-Time Artificial Insemination Protocol" Biology 10, no. 5: 429. https://doi.org/10.3390/biology10050429
APA StyleBustamante-Andrade, J. A., Meza-Herrera, C. A., Rodríguez-Martínez, R., Santos-Jimenez, Z., Ángel-García, O., Gaytán-Alemán, L. R., Gutierrez-Guzman, U. N., Esquivel-Romo, A., & Véliz-Deras, F. G. (2021). Luteogenesis and Embryo Implantation Are Enhanced by Exogenous hCG in Goats Subjected to an Out-of-Season Fixed-Time Artificial Insemination Protocol. Biology, 10(5), 429. https://doi.org/10.3390/biology10050429