Abstract
Campylobacteriosis is the leading cause of human bacterial gastroenteritis, very often associated with poultry consumption. Thermophilic Campylobacter (Campylobacter jejuni and Campylobacter coli) isolates (n = 158) recovered from broiler neck skin and caecal contents in Ireland over a one-year period, resistant to at least one of three clinically relevant antimicrobial classes, were screened for resistance determinants. All ciprofloxacin-resistant isolates (n = 99) harboured the C257T nucleotide mutation (conferring the Thr-86-Ile substitution) in conjunction with other synonymous and nonsynonymous mutations, which may have epidemiological value. The A2075G nucleotide mutation and amino acid substitutions in L4 and L22 were detected in all erythromycin-resistant isolates (n = 5). The tetO gene was detected in 100% (n = 119) of tetracycline-resistant isolates and three of which were found to harbour the mosaic tetracycline resistance gene tetO/32/O. Two streptomycin-resistant C. jejuni isolates (isolated from the same flock) harboured ant(6)-Ib, located in a multidrug resistance genomic island, containing aminoglycoside, streptothricin (satA) and tetracycline resistance genes (truncated tetO and mosaic tetO/32/O). The ant(6)-Ie gene was identified in two streptomycin-resistant C. coli isolates. This study highlights the widespread acquisition of antimicrobial resistance determinants among chicken-associated Campylobacter isolates, through horizontal gene transfer or clonal expansion of resistant lineages. The stability of such resistance determinants is compounded by the fluidity of mobile genetic element.
1. Introduction
Campylobacter is the most commonly reported foodborne bacterial pathogen causing human gastroenteritis in the European Union (EU) and Ireland, most often associated with the broiler reservoir [1]. Ireland was found to have a 98% prevalence of campylobacter-contaminated broiler carcasses in 2008 [2]. Frequent isolation of antimicrobial-resistant Campylobacter spp. of food animal origin continues to limit the spectrum of clinically useful antimicrobials and is internationally recognised as a major societal challenge. Veterinary antimicrobials used therapeutically and prophylactically are often the same as, or belong to the same class as those used clinically [3].
Macrolides, fluoroquinolones and aminoglycosides are classified as critically important antimicrobials, while tetracycline is considered a highly important antimicrobial [3]. Resistance to (fluoro)quinolones and tetracyclines is highly prevalent in clinical and broiler-associated Campylobacter spp. isolates, while resistance to erythromycin is typically low to moderate across Europe [4,5,6]. Macrolides are the first line antibiotic for the treatment of enteric gastroenteritis, while fluoroquinolones and tetracyclines remain as alternatives [7,8,9]. Systemic infections are routinely treated with aminoglycosides [9,10] and resistance to aminoglycosides is low in clinical and broiler isolates across Europe [5]. Cross-resistance between aminoglycoside antimicrobials is incomplete and although streptomycin is not used clinically to treat campylobacteriosis, resistance can be used as an indicator for acquired aminoglycoside resistance genes.
In Gram negative bacteria, DNA gyrase is the primary target of fluoroquinolones [11]. DNA gyrase is a heterotetrameric type IIA topoisomerase, consisting of two polypeptide subunits (GyrA and GyrB, encoded by gyrA and gyrB, respectively), catalysing ATP-dependent negative supercoiling of DNA to regulate replication, repair and gene expression [12,13,14]. Resistance to (fluoro)quinolones among Campylobacter spp. is largely mediated by chromosomal mutations in the quinolone resistance-determining region (QRDR) of gyrA, typically conferred by the C257T nucleotide mutation (Thr-86-Ile) [15]. The QRDR is located near the Tyr-125 active site, involved in DNA-protein bridge formation during DNA strand passage [15].
Macrolides act by binding to the 50S bacterial ribosomal subunit and inhibit translational elongation, and interfere with protein synthesis and subsequent ribosomal subunit assembly [16,17]. Polymorphisms in the 23S ribosomal RNA (rRNA), mutations in 50S ribosomal proteins L4 and L22 (encoded by rplD and rplV, respectively) or the presence of the emerging ermB gene contribute to macrolide resistance [8,18]. Βeta-hairpin extensions from 50S ribosomal proteins L4 and L22 are involved in the regulation of nascent peptide exit from the large ribosomal subunit [19,20]. Mutations in L4 and L22, combined with the overexpression of antimicrobial efflux genes have been reported to contribute to high-level macrolide resistance [21]. The ribosomal methylase encoded by ermB was reported recently for the first time in thermophilic Campylobacter spp., located on a chromosomal multidrug resistance genomic island (MDRGI) (likely originating from a Gram positive species) in a high-level erythromycin-resistant Campylobacter coli (C. coli) isolate (ZTC113) of swine origin in China [8,22]. ErmB dimethylates adenine at position 2074 of the 23S rRNA gene, reducing the binding affinity of macrolides [23].
The intrinsic, chromosomally encoded resistance-nodulation-division (RND) CmeABC (Campylobacter multidrug efflux) efflux pump in Campylobacter spp. contributes to baseline resistance against structurally diverse antimicrobials [6,24,25,26]. The cmeABC operon encodes a tripartite multidrug efflux pump that consists of an outer membrane channel protein (cmeC), an inner membrane efflux transporter (cmeB) and a periplasmic fusion protein (cmeA) [27]. Repressor (cmeR) binding to an inverted repeat (IR) (TGTAATAAATATTACA) in the intergenic region between cmeR and cmeA transcriptionally represses the cmeABC operon [28,29]. Consequently, polymorphisms in the repressing site induce efflux pump overexpression and enhanced resistance to antimicrobials, most notably, erythromycin [8,30].
Tetracycline resistance in Campylobacter spp. is largely conferred by a ribosomal protection protein (RPP), TetO, capable of displacing tetracycline from its primary binding site on the 30S ribosomal subunit [17,31]. Bacterial resistance to tetracycline is also associated with ATP-dependent efflux or enzymatic inactivation of tetracycline [32,33,34]. Campylobacter tetO can be located chromosomally but is often plasmid-mediated [31,33,35,36,37,38,39]. Tetracycline RPPs are widely distributed among bacterial genera and it has been reported that the tetO gene exists in at least eleven bacterial genera, including four Gram negative and seven Gram positive genera [33]. The conserved acquisition of tetO between members of different bacterial genera indicates that conjugative plasmids, transposons, or recombination events contribute to the dissemination and maintenance of the tetO gene [37].
Although tetO acquisition is the most prevalent genetic event conferring tetracycline resistance among Campylobacter spp., mosaic tet genes (specifically tetO/32/O) have also been reported within the genus [34]. Mosaic tetracycline resistance genes are derived from the interclass (double-crossover) recombination of two or more RPP-encoding gene (predominantly tetO, W, 32 [34,40,41,42] and tetM, S [43]) to form functional chimera [34]. Mosaic tet genes are widespread among Gram positive and Gram negative genera in human and animal isolates [41].
Aminoglycosides are broad spectrum antimicrobials and inhibit protein synthesis by binding to 16S rRNA of the 30S ribosome [44,45]. Campylobacter spp. resistance to aminoglycosides is mediated by reduced antimicrobial binding affinity for target sites due to enzymatic modification, via acetylation, phosphorylation or adenylation of amino or hydroxyl groups of the aminocyclitol nucleus or sugar moieties [44,45,46]. Although there are two main nomenclature systems used to identify aminoglycoside modifying enzymes [47,48,49], we followed the system proposed by Shaw et al. (1993), later extended to include an expanded panel of aminoglycoside 6-nucleotidyltransferases (also known as adenyltransferases) [50,51]. The designation proposed by Shaw et al. (1993) is as follows: the type of modification (nucleotidyltransferase/adenyltransferases (ANT)); the modification site (6’); a roman numeral to denote unique resistance profiles (I), and a letter to represent unique protein sequences (b) [48]. Genes for ANT enzymes are found on transposons, plasmids or chromosome, often in associated with other resistance genes and very often as part of the transposon-associated aminoglycoside-streptothricin resistance gene cluster (ant(6)-I-sat4-aphA3), first isolated from Staphylococci [52,53]. ANT(6)-I encoding genes are widely distributed among clinical and animal streptomycin-resistant thermophilic Campylobacter spp. isolates [51].
Despite the high prevalence of Campylobacter in broilers on the island of Ireland, in the last twenty years, only a few reports of the molecular mechanisms contributing to resistance exist for broiler [54,55,56,57,58,59], clinical [54,56,59,60,61], domestic animal [62] or ruminant [56,63] isolates. We report the antimicrobial resistance determinants circulating among 158 resistant Campylobacter jejuni (C. jejuni) and C. coli isolates recovered from Irish broiler neck skin and caecal samples over a one-year period (2017–2018).
2. Results
2.1. Fluoroquinolone Resistance
Isolates resistant to ciprofloxacin/nalidixic acid were screened for mutations in the gyrA gene and 100% of isolates (n = 99) harboured a C257T point mutation, which is the dominant mutation conferring resistance among campylobacters. Resistant isolates were grouped into C. jejuni gyrA and C. coli gyrA (arbitrarily named GTJs and GTCs, respectively) sequence types based on the presence of synonymous and nonsynonymous mutations present in the portion of the gyrA gene sequenced (Table 1). Ciprofloxacin-resistant C. jejuni isolates (n = 85) were grouped into three GTJs (GTJ-I, -II, and -III). A large proportion (47.1%) carried the Thr-86-Ile substitution exclusively (GTJ1). Synonymous mutations T72C, C243T, T357C, C360T, C471T, T483C, and C622T were exclusively associated with C. jejuni and were present in both GTJ-II and GTJ-III. Nonsynonymous Ser-22-Gly (A64G) and Ala-206-Thr (G616A) mutations were present in 35.3% and 17.7% of isolates of ciprofloxacin-resistant C. jejuni isolates, respectively and were the basis of defining GTJ-II and GTJ-III, respectively. Both GTJ-II and GTJ-III were associated with the Asn-203-Ser (A608G) substitution. All CIP-resistant C. coli isolates tested (n = 14) harboured one nonsynonymous mutation only (Thr-86-Ile), but were grouped into seven GTCs based on the presence of various synonymous mutations (Table 1).

Table 1.
GyrA sequence types (GTs) and associated polymorphisms distributed among 85 ciprofloxacin-resistant C. jejuni isolates (GTJs) and 14 ciprofloxacin-resistant C. coli isolates (GTCs). Polymorphisms causing an amino acid substitution (nonsynonymous mutations) are highlighted in black.
No high-level moxifloxacin resistance was detected among the 99 (fluoro)quinolone-resistant isolates tested and minimum inhibitory concentrations (MICs) ranged from 0.5–8 mg/L.
2.2. Erythromycin Resistance
Five erythromycin-resistant isolates were screened for mutations contributing to erythromycin resistance. All five erythromycin-resistant isolates harboured the A2075G mutation in the 23S rRNA gene. A T82C mutation (Ser-28-Pro substitution) in the rplD gene was detected in the five isolates, and the partial sequences shared 100% homology with the rplD gene of erythromycin-sensitive C. coli isolates (GenBank accession numbers: MH084640.1 and MH084639.1) [64]. Identical mutations were also observed in the rplV sequence in all erythromycin-resistant isolates: double point mutation at positions 308 and 309 (Ala-103-Val), A325G (Thr-109-Ala), a double point mutation at positions 332 and 333 (Ala-111-Glu), G340A (Ala-114-Thr) and C358A (Pro-120-Thr). Nonsynonymous mutations (T282A, C294T, A306G, T321G) in rplV were also identified in all erythromycin-resistant isolates. Partial rplV sequences in this study were homologous to a high-level erythromycin-resistant clinical C. coli isolate rplV gene (GenBank accession number: GU384982.1) [28]. Similarly, all five erythromycin-resistant isolates harboured a 9 base pair deletion (positions 45–54) and an insertion at position 45 (G) in cmeR-cmeA intergenic region, upstream of the IR (positions 66-80), homologous to sequences derived from erythromycin-resistant (GenBank accession number: FJ797673.1) and erythromycin-sensitive (GenBank accession number: FJ797671.1) strains [24]. The ermB gene was not detected among the erythromycin-resistant isolates tested.
2.3. Tetracycline Resistance
A portion of the tetO gene was detected in 100% of tetracycline-resistant isolates (n = 119). Three isolates (CITCj625-18, CITCj727-18, and CITCc3448-18), accounting for 2.5% of the tetracycline-resistant isolates, harboured the mosaic tetO/32/O type II gene, confirmed by PCR/partial sequencing and genomic sequencing (Figure 1B). Isolates CITCj625-18 and CITCj727-18 carried identical mosaic tetracycline genes, but differed from CITCCc3448-18 by Thr-118-Ile and Glu-176-Asp substitutions and A684G and A789G point mutations. The mosaic tetracycline gene detected among the Irish broilers isolates in this study was very similar to a Streptococcus suis (S. suis) (GenBank accession number: KY994102.1) tetO/32/O gene (Table 2). Equally, the TetO/32/O sequences were >99.5% identical to Campylobacter spp. (GenBank accession numbers: WP_052855148.1) and Clostridiales (WP_117823345.1) TetO/32/O sequences.
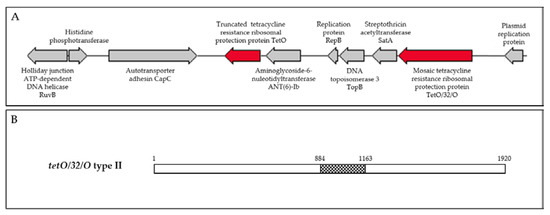
Figure 1.
(A) Schematic of Campylobacter jejuni isolate CITCj625-18 multidrug resistance genomic island. (B) Schematic of the mosaic tetO/32/O type II gene detected in CITCj625-18 and CITCj727-18. Figure adapted from Warburton et al. (2016). White bars are tetO and central, checked bar is tet32 (297 bp).
Table 2.
Percentage identity (percentage cover in brackets) of mosaic tetracycline resistance gene (tetO/32/O) type II in three tetracycline-resistant Campylobacter spp. isolates detected in this study and Streptococcus suis tetO/32/O gene (GenBank accession number: KY994102.1).
2.4. Streptomycin Resistance
C. jejuni isolates CITCj625-18 and CITCj727-18 were found to harbour multiple resistance genes, including a truncated tetO (873 bp, truncated at the 3’ end), mosaic tetO/32/O type II (1920 bp), aminoglycoside-6-nucleotidyltransferase (ant(6)-Ib)), and streptothricin acetyltransferase (satA). These antimicrobial resistance genes were located circumjacent to proteins involved in replication and recombination (Table 3, Figure 1A). The MDRGI contained 10 open reading frames in a region of 35.9% GC content, compared to the average genomic GC content of 30.5%. Genomic sequences of C. jejuni isolates CITCj625-18 (first thin) (1,630,363 bp) and CITCj727-18 (final thin, isolated a week later) (1,636,524 bp) were nearly identical with an average nucleotide identity (ANI) of 99.99% and 30.5% GC content, indicating that the isolates had been circulating within and had been maintained by the flock. The pair belonged to sequence type ST-45 clonal complex (ST-137) and harboured LOS locus class C.
Table 3.
Description of CITCj625-18 (representative of CITCj727-18) multidrug resistance genomic island.
CITCc1631-18 and CITCc3448-18 belonged to ST-828 clonal complex (ST-6543 and ST-1096, respectively) and harboured the ant(6)-Ie gene (900 bp), with almost identical sequences (99.89% identity). ANT(6)-Ie in this study shared 99.66% amino acid identity with each other, where CITCc3448-18 harboured a nonsynonymous G820A mutation (Val-274-Ile) and was identical to a C. coli ANT(6)-Ie protein (GenBank: WP_052786298.1). CITCc1631-18 and CITCc3448-18 ANT(6)-Ie shared between 29.7-36.1% identity to ANT(6)-Ia, ANT(6)-Ib, ANT(6)-Ic, and ANT(6)-Id amino acid sequences (GenBank: AFJ97257.1, AFJ97264.1, AAR10415.1, and WP_001258597.1, respectively) [50,51,65].
Aminoglycoside 3-N-acetyltransferase (AAC(3)) (261 amino acids) was also detected in both CITCj625-18 and CITCj727-18, 44.3% identical to a aminoglycoside 3-N-acetyltransferase (AAC(3)) variant (239 amino acids) in both CITCc1631-18 and CITCc3448-18. However, these isolates were gentamicin-susceptible, although this protein may confer resistance to other aminoglycoside antibiotics [66]. The C. coli variant was identical to AAC(3) genes reported in Campylobacter spp., 37.5% identical to AAC(3)-Ia described in Serratia marcescens (GenBank accession number: Q7B9H0), and 19–32.25% similar to AAC(3) orthologous, including types AAC(3)-Ib/Ic/Id/Ie/IIa/IIb/IIc/IId/IIe/IIf/IIIa/IIIb/IIIc/IVa/VIa/VIIIa/Ixa/Xa/XIa. The AAC(3) variant in CITCj625-18 and CITCj727-18 was identical to Campylobacter AAC(3) variants and was 19.3–25.84% similar to orthologues reported in other bacterial genera.
2.5. Overall Distribution of Antimicrobial Resistance
The antimicrobial resistance rates of this pool of broiler-associated thermophilic Campylobacter spp. isolates (resistant to at least one antimicrobial; n = 158) have been detailed previously [67], and are summarised below (Figure 2).
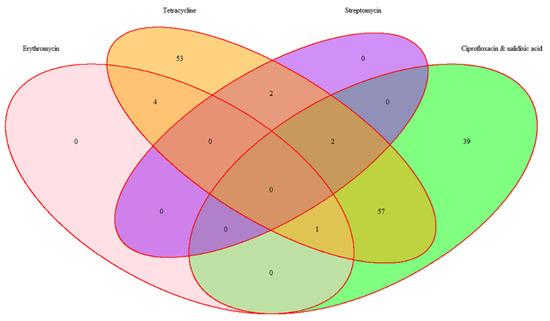
Figure 2.
Distribution of antimicrobial resistance among 158 thermophilic Campylobacter spp.
3. Discussion
This study reports the antimicrobial resistance determinants circulating among Irish broiler-associated Campylobacter isolates, collected throughout the Republic of Ireland, from the three largest poultry processors, over a one-year period (2017–2018).
The Thr-86-Ile mutation is the predominant genetic alteration conferring (fluoro)quinolone resistance among Campylobacter spp. [68], and was detected in all (fluoro)quinolone-resistant isolates (n = 99) tested in the current study, similar to reports published worldwide [6,69,70,71]. Some studies have not detected the Thr-86-Ile mutation universally in the QRDR of (fluoro)quinolone-resistant Campylobacter spp. isolates [58,62,72], indicating that other factors are responsible for, or contribute to, (fluoro)quinolone resistance.
Fluoroquinolone-resistant Campylobacter spp. are ecologically competitive and persistent even in the absence of antimicrobial selective pressure [73,74,75]. Despite a distinct rise in ciprofloxacin resistance among broiler Campylobacter isolates (3.1% to 28.9%) in Ireland between 1998 and 2000 [54,76], in 2017–8 resistance to ciprofloxacin remained stable (28.3%) [67]. Clonal expansion of resistant lineages has likely contributed to the persistence of ciprofloxacin resistance in Ireland, considering that fluoroquinolones typically account for less than 1% of all veterinary antimicrobials sold in Ireland [77,78,79].
C. jejuni isolates harbouring multiple amino acid substitutions in the gyrA QRDR (GTJ-II and GTJ-III) (Table 1) had ciprofloxacin MICs ranging from 8–16 mg/L, except one GTJ-II isolate had an MIC of 4 mg/L, while isolates harbouring the Thr-86-Ile mutation exclusively (GTJ-I) had MICs ranging from 4–16 mg/L. Similarly, Ekkapobyotin et al. (2008) observed varying ciprofloxacin/nalidixic acid MICs in isolates harbouring identical GyrA amino acid substitutions [80]. Moreover, the Ser-22-Gly, Asn-203-Ser and Ala-206-Thr mutations have been reported in fluoroquinolone-sensitive strains [81,82,83]. To confirm the apparent lack of involvement of these accessory gyrA mutations in the development of fluoroquinolone resistance, the introduction of these mutations in fluoroquinolone-susceptible strains could be investigated. Authors have previously reported that double mutations in gyrA (at amino acid positions 86 and 90) were necessary to produce high level moxifloxacin resistance [84,85]. Moxifloxacin is a potent fluoroquinolone with activity against fluoroquinolone-resistant campylobacters that harbour a single mutation in gyrA [86]. In this study, no high-level resistance to moxifloxacin was observed among the (fluoro)quinolone-resistant isolates. These data indicate that mutations outside the gyrA QRDR have a negligible effect on (fluoro)quinolone resistance.
Variation in gyrA alleles within a population of Campylobacter isolates have been identified as epidemiological markers and may serve as a supplementary approach to classical epidemiological typing methods [81,83,87,88]. In our study, the GTs detected were species specific, although Ragimbeau et al. (2014) reported the presence of a typical C. coli gyrA type in 0.23% (n = 1) of 430 C. jejuni isolates tested, and 1.4% (n = 4) C. coli isolates harboured a typical C. jejuni gyrA type. The amino acid substitutions present in each of the three GTJ (Table 1) lineages have been associated with poultry Campylobacter isolates previously [89,90,91]. Only one nonsynonymous mutation (Thr-86-Ile) was detected among the seven GTCs detected (Table 1), similar to previous studies reporting a single nonsynonymous mutation (Thr-86-Ile substitution) present in (fluoro)quinolone-resistant C. coli QRDR sequences [83,89,90]. It is likely that additional variants of Campylobacter spp. gyrA alleles exist, and may reflect ecological evolution [83].
Identical mutations in the 23S rRNA, rplD, and rplV genes were detected in erythromycin-resistant isolates (n = 5), while MICs ranged from 128 mg/L to ≥ 128 mg/L. Three of these isolates were collected from the same flock in north-central Ireland while one isolate was collected from a farm approximately 10 km away, the following week. The fifth erythromycin-resistant isolate was recovered from a farm in the mid-south-west of Ireland, two months previously, but all birds from these farms were processed in the same processing plant.
The A2075G point mutation in the 23S rRNA gene remains the most prevalent genetic event conferring macrolide resistance [8,92] and was detected in all erythromycin-resistant isolates in this study. The 23S rRNA A2074G and A2074C mutations were not detected. Mutations in the rplV gene, encoding the 50S ribosomal protein L22 were detected in the C-terminal region (amino acids 109–142) [18], including Thr-109-Ala, Ala-111-Glu, Ala-114-Thr, and Pro-120-Thr. Nonsynonymous mutations (T282A and C294T) were also observed in the region encoding the highly conserved β-hairpin loop at amino acids 78–98 [18]. Mutations in the RplD β-hairpin (spanning amino acid positions 55–77 [18,93]) are often associated with bacterial macrolide resistance, and such mutations were not observed among the Irish erythromycin-resistant isolates tested in this study. Moreover, polymorphisms were detected outside the cmeR regulatory IR. The effects of mutations detected in this study in rplD, rplV, and the intergenic region of cmeR-cmeA are unknown, but may contribute to erythromycin resistance.
The ermB gene was not detected among the erythromycin-resistant isolates tested. Resistance mediated by ermB in Campylobacter spp. is largely confined to China, which may reflect the extensive use of antimicrobials in food producing animals in China [8]. Three reports of genetically distinct ermB-positive C. coli isolates recovered from poultry exist in Europe [94,95,96] and an ermB-positive isolate was detected for the first time in the United States in a C. jejuni isolate of clinical origin [97], while the ermB gene was recently detected in 18.3% of 240 thermophilic Campylobacter spp. retail meat associated-isolates in South Africa [98]. Mutations in C. jejuni 23S rRNA has been associated with a fitness cost and reduced doubling times [99,100,101], although tolerance to low temperatures may facilitate persistence in the environment and transmission of resistant strains through the food supply [100].
All tetracycline-resistant isolates harboured a portion of the tetO gene, while three isolates harboured the mosaic tetO/32/O type II gene were detected. Mosaic tetracycline resistance genes in Campylobacter spp. are typically derived from tetO and tet32 sequences in the type II conformation, with a shorter central tet32 segment [34], although there are limited reports of these resistance genes circulating among Campylobacter spp. The first mosaic tetracycline RPP gene was detected in Megasphera elsdenii, composed of a central tetW region flanked by two tetO regions [102]. However, the progenitors of these mosaic genes are based only on the order in which they were discovered, and the current classification system does not adequately reflect the variable nature of tetracycline RPPs [34,43]. The tetO primers [103] used in this study amplify a region at the beginning of the tetO gene and enable the detection of mosaic tet genes with a central portion flanked by an initial tetO region until position 228. The tetO/32/O type II gene reported here was associated exclusively with an MIC of 64 µg/L. However, 27 of 119 (22.7%) tetracycline-resistant Campylobacter spp. isolates tested had MICs of 64 µg/L or ≥ 64 µg/L, indicating that other factors contribute to enhanced tetracycline resistance. It should be noted that in this study, all three isolates harbouring mosaic tetracycline genes were also co-resistant to streptomycin, enabling co-selection and persistence of these antimicrobials. The burden of mosaic tetracycline resistance genes within the genus should be considered as part of the approach to elucidate developing and newly acquired antimicrobial resistance determinants within the genus.
All streptomycin resistant (n = 4) isolates had MICs of ≥ 16 mg/L. Streptomycin-resistant C. jejuni isolates CITCj625-18 and CITCj727-18 (ST-137) harboured sialylated LOS locus class C, which has been identified as a risk factor for post-infectious Guillain-Barré syndrome and increased severity of enteric disease [104]. CITCj625 and CITCj727-18 belonged to ST-137 and members of the diverse C. jejuni ST-45 clonal complex [105,106]. ST-137 is frequently isolated from cases of enteric campylobacteriosis [105] and broilers/avian [107,108,109], porcine [105], and bovine [110] hosts. The ST-137 genotype is widely dispersed and represents an ecologically successful clone [106]. A study by Dearlove et al. (2016) reported that ST-45 clonal complex was a generalist lineage capable of frequent transmission between hosts.
Genomic sequencing of tetracycline-/streptomycin-resistant C. jejuni isolates (CITCj625-18 and CITCj727-18, isolated from the same flock, first and final thin) revealed identical genes in a multidrug resistance genomic island (Table 3, Figure 1A). Both isolates harboured a truncated tetO gene and a mosaic tetO/32/O type II gene, homologous that of Gram positive (GenBank accession number: KY994102.1) and Campylobacter spp. (GenBank accession number: WP_002823161.1). The presence of multiple tet genes (coding for similar or different mechanisms) in Gram negative isolates has also been documented [33]. However, the truncated form detected in this study (CITCj625-18 and CITCj727-18) is likely a remnant of a region of insertion or recombination. Truncated tetO genes have also been reported in C. coli MDGRI containing aadE (ant(6)-Ib) and ermB [94,111]. Aminoglycoside (ant(6)-Ib (867 bp)) and streptothricin resistance (satA) genes were also located within the multidrug resistance island (Figure 1A). Streptothricin acetyltransferase A (satA) is frequently reported in Gram positive bacilli [112] and shares less than 40% identity with the streptothricin acetyltransferase A (sat4) reported in Campylobacter [52,111,113]. Plasmid replication proteins within the MDRGI suggest a plasmid as the insertion vehicle of the resistance genes.
CITCc3448-18 belonged to ST-828 clonal complex (ST-1096). ST-1096 has been isolated from a case of gastroenteritis in the United Kingdom (UK) in 2016 [114] and has also been previously reported in C. coli of swine origin in Spain, America, and Grenada [51,115,116]. CITCc1631 was ST-6543 (ST-828 clonal complex), which has been associated with clinical and chicken-associated isolates [117]. There are a total of fifteen depositions (all are UK associated) of C. coli ST-6543 on the PubMLST Campylobacter database at the time of writing [114], including eleven clinical isolates (faeces), two chicken-associated isolates, and two isolates with no source allocation.
Both streptomycin-resistant C. coli isolates (CITCc1631-18 and CITCc3448-18) harboured the emerging ant(6)-Ie gene (900 bp), found widely disseminated among clinical and animal C. coli isolates [51,65]. Unlike other Campylobacter ant(6) genes, ant(6)-Ie appears to be inherent to C. coli and does not have a Gram positive ancestor [65]. The ant(6)-Ie gene was originally detected in a hypervariable genomic region, unaccompanied by other resistance genes [65]. Similarly, the CITCc1631-18 and CITCc3448-18 ant(6)-Ie genes were located chromosomally, and were not located near other resistance determinants. Both streptomycin-resistant C. coli isolates were co-resistant to ciprofloxacin/nalidixic acid (GTC-V) and tetracycline. C. coli isolate 1631-18 harboured tetO (1920 bp), while C. coli isolate CITCc3448-18 also harboured the mosaic tetracycline resistance gene, tetO/32/O (1290 bp), highly homologous with that detected in CITCj625-18 and CITCj727-18.
4. Materials and Methods
4.1. Bacterial Isolate Culture Conditions and Susceptibility Testing
A total of 350 thermophilic Campylobacter isolates (296 C. jejuni and 54 C. coli) were recovered from free range and intensively-reared broiler carcasses (neck skin and caecal contents) using ISO 10272-2:2017 [2]. Isolates were collected between September 2017 and September 2018, from the three largest poultry processing plants in the Republic of Ireland. The collection of isolates was speciated using matrix-assisted laser desorption ionisation-time of flight (MALDI-TOF) mass spectrometry (MS) (Bruker, Billerica, MA, United States). Isolates were previously tested [67] for their MIC to six clinically relevant antimicrobials, namely ciprofloxacin, nalidixic acid, erythromycin, tetracycline, gentamicin, and streptomycin according to ISO 20776:2006 and EC Decision 2013/652/EU [118,119]. Overall, 158 (140 C. jejuni and 18 C. coli) isolates tested were resistant to at least one antimicrobial and were subsequently tested for resistance determinants.
4.2. DNA Extraction
Briefly, isolates were recovered from −80 °C on Columbia blood agar (Fannin Ltd, Dublin, Ireland) and incubated for 24 h at 42 °C, under microaerobic conditions (5% O2, 10% CO2, 85% N2) and subcultured. DNA was extracted using the PureLink Genomic DNA Mini Kit (Invitrogen, Carlsbad, CA, USA), according to manufacturer’s instructions and DNA was standardised to 50–100 ng/μL.
4.3. Genotypic Characterisation of Antimicrobial Resistance—PCR Amplification and Sequencing
Primer sets, target genes and annealing temperatures are listed in Table 4. Primers to detect mosaic tetracycline resistance genes (tetO/32/O and tetO/W/O) were designed on SnapGene2.3.2 software (from Insightful Science; available at snapgene.com) and regions of primer complementarity were assessed on Primer-BLAST [120].
Table 4.
Primer sequences and target genes used for the detection of antimicrobial resistance determinants in resistant thermophilic Campylobacter spp. isolates and sialylated lipooligosaccharide (LOS) locus classes A, B and C in streptomycin-resistant C. jejuni isolates.
PCRs were performed with 2.5 U of Amplitaq polymerase (Applied Biosystems, Foster City, CA, USA), 1 × PCR buffer I and 2.5 mM magnesium chloride (Applied Biosystems), 0.2 mM of each deoxyribonucleotide (Sigma Aldrich, St Louis, MO, USA), 0.2 pmol/μL of each primer, and 1 µL (1–2 ng/μL) of DNA. Reaction conditions were denaturation at 94 °C for 2 min, 35 cycles of denaturation at 94 °C for 30 s, annealing (Table 4) for 30 s and extension at 72 °C for 1 min followed by a final extension at 72 °C for 5 min. PCR products were purified using the High Pure PCR Purification Kit (Sigma Aldrich). Purified PCR products were Sanger sequenced (forward and reverse reads) (Table 4) by Eurofins Genomics (Eurofins Genomics, Ebersberg, Germany). Consensus sequences were aligned and assembled on SeqMan Pro (Lasergene) (DNAStar, Madison, WI, USA).
Ciprofloxacin-resistant isolates (n = 99) were screened for mutations in the QRDR of the gyrA gene [15]. Products were purified and sequenced, as described above. Consensus sequences were aligned to the gyrA of the reference C. jejuni (GenBank accession number: L04566.1 and AL111168.1) and C. coli sequences (GenBank accession number: AF092101.1 and NZ_UIGM01000003.1) on SnapGene 2.3.2.
The primers used for the amplification and sequencing of domain V of 23S rRNA, rplD, and rplV (encoding L4 and L22 ribosomal proteins, respectively) and the regulatory site of the cmeABC operon in five erythromycin-resistant C. coli isolates are listed in Table 4. Partial multiple alignment to reference C. coli type strain NCTC 11366 (ATCC 33559) 23S rRNA (GenBank accession number: GQ167698.1), rplD (GenBank accession number: DQ639752.1 and UIGM01000003.1), rplV (GenBank accession number: UIGM01000003.1) and cmeABC operon (GenBank accession number: FJ797670.1) sequences was performed on SnapGene 2.3.2. Isolates were also screened for the presence of ermB, according to Wang et al. (2014).
Tetracycline-resistant isolates (n = 119) were screened for the presence of tetO, according to Aminov et al. (2001) and products were visualised on 2% agarose gel electrophoresis. The tetO amplicon of a tetracycline-resistant C. jejuni isolate (CITCj382-18) from this study was purified and sequenced (as described above) as a positive control. Consensus sequences were aligned to the C. jejuni tetO gene (GenBank accession number: M18896.2).
Primers were designed to target tetO/W/O (Table 4) based on alignments between tetO (GenBank accession number: M18896.2) and mosaic tetO/WO genes (GenBank accession numbers: EF065524.1 and AY196921.1). Two tetO/32/O-targeting primers (Table 4) were designed based on regions of homology between tetO (GenBank accession number: M18896.2) and tetO/32/O type I genes with a longer central tet32 region (GenBank accession numbers: AJ295238.3 and JQ740052.1) and tetO/32/O type II genes with a shorter central tet32 segment (GenBank accession numbers: KY994102.1, FP929050.1, AIOQ01000025.1, NZ_AUJS01000017.1 and AABYPB010000033.1) [34].
Streptomycin-resistant isolates (n = 4) were screened for sialylated LOS locus class A/B and C (Table 4) by PCR, using C. jejuni 81-176 (ATCC BAA2151) and C. jejuni NCTC 11168 (DSM 27585), respectively, as positive controls.
4.4. Moxifloxacin Minimum Inhibitory Concentration Testing
All (fluoro)quinolone-resistant isolates (n = 99) were tested for moxifloxacin susceptibility. Briefly, isolates were recovered from −80 °C on CBA (Fannin Ltd) for 24 h at 42 °C under microaerobic conditions, and subcultured to CBA for 20 ± 2 h at 42 °C under microaerobic conditions. In microtiter plates, 100 µL serial dilutions of moxifloxacin (Sigma Aldrich) were prepared in of Mueller Hinton broth with lysed horse blood (Thermo Fisher Scientific, Waltham, MA, USA) ranging from 0.125–16 mg/L. A 0.5 McFarland inoculum was prepared in 5 mL demineralised water (Thermo Fisher Scientific) and 100 µL was transferred to 11 mL of Mueller Hinton broth with lysed horse blood (Thermo Fisher Scientific). Moxifloxacin serial dilutions were inoculated with 100 µL of cell suspension and incubated for 20 ± 2 hat 42 °C under microaerobic conditions.
4.5. Genome Sequencing and Genomic Analysis
The genomes of four streptomycin-resistant isolates (C. jejuni isolates CITCj625-18 and CITCj727-18 and C. coli isolates CITCc1631-18 and CITCc3448-18) were sequenced. DNA was quantified in triplicate with the Quant-iT dsDNA Assay Kit (Thermo Fisher Scientific). Genomic DNA libraries were prepared using the Nextera-XT protocol (Illumina, San Diego, CA, USA), with changes including 2 ng of input DNA and a minute PCR elongation time. DNA quantification and library preparation were performed on a Hamilton Microlab STAR system. Pooled libraries were quantified using the Kapa Biosystems library quantification kit on a Roche light cycler 96 qPCR machine. Libraries were sequenced on the Illumina HiSeq using a 250 bp paired-end protocol. Reads were adapter trimmed using Trimmomatic 0.30 with a sliding window quality cut-off of Q15 [124]. De novo assembly was performed using SPAdes version 3.7 [125] and assembly quality was assessed using QUAST [126].
Genomes were annotated using Prokka 1.14.3 [127]. Similarity searches were performed using the BLAST suit of programs [128] and InterProScan [129]. Multi locus sequence typing (MLST) patterns was determined using PubMLST [114]. ANI was calculated using EzBioCloud [130].
4.6. Data Availability
This whole-genome project has been deposited at DDBJ/ENA/GenBank under the accession number PRJNA612628. CITCj625-18, CITCj727-18, CITCc1631-18, and CITCc3448-18 genomes can be accessed using SAMN14379027, SAMN14379028, SAMN14379029, and SAMN14379030. The partial and complete gene sequences deposited in GenBank are listed in Table 5. Erythromycin-resistant isolates harboured identical ribosomal mutations and CITCc1303-18 partial sequences were submitted. One representative gyrA GT was submitted for each type.
Table 5.
Accession numbers of partial sequences submitted to GenBank in this study.
5. Conclusions
Although non-poultry sources contribute to campylobacteriosis incidence, poultry are natural thermophilic Campylobacter spp. hosts. The broiler industry serves as a reservoir for the dissemination of resistant campylobacters. The enrichment and stability of Campylobacter spp. resistance determinants is noteworthy but the natural competence and potential of recombination or acquisition of mobile genetic elements contributes to the Campylobacter. Taken together, the data collected in this study point to the diversity of resistance determinants circulating among Irish broilers, contributing to the development of resistance to clinically relevant antimicrobials.
Author Contributions
Investigation, C.T.L., S.B., and K.H.; methodology, C.T.L., H.L., C.B. and C.M.C.; original draft preparation, C.T.L., B.L., and A.C.; writing—review and editing, C.T.L., C.B., B.L., and A.C.; supervision, B.L. and A.C.; funding acquisition, A.C., J.E., P.W., and D.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Department of Agriculture Food and the Marine, Food Institutional Research Measure, grant number 15F641. C.T.L. was funded by a Teagasc Walsh Fellowship, grant number 2017265.
Acknowledgments
Genome sequencing was provided by MicrobesNG (http://www.microbesng.uk) which is supported by the BBSRC (grant number BB/L024209/1).
Conflicts of Interest
The authors declare no conflict of interest.
References
- European Food Safety Authority. Scientific Opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 94, 2105. [Google Scholar]
- European Food Safety Authority. Analysis of the baseline survey on the prevalence of Campylobacter in broiler batches and of Campylobacter and Salmonella on broiler carcasses in the EU. EFSA J. 2010, 8, 1522. [Google Scholar]
- World Health Organisation Critically Important Antimicrobials for Human Medicine 6th Revision. 2018. Available online: https://www.who.int/foodsafety/publications/antimicrobials-sixth/en/ (accessed on 1 April 2020).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, aimals and food in 2017. EFSA J. 2019, 17, e04694. [Google Scholar]
- European Food Safety Authority; European Centre for Disease Prevention and Control. European union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16, 4380. [Google Scholar]
- Duarte, A.; Santos, A.; Manageiro, V.; Martins, A.; Fraqueza, M.J.; Caniç, M.; Domingues, F.C.; Oleastro, M. Human, food and animal Campylobacter spp. isolated in Portugal: High genetic diversity and antibiotic resistance rates. Int. J. Antimicrob. Agents 2014, 44, 306–313. [Google Scholar] [CrossRef]
- Alfredson, D.A.; Korolik, V. Antibiotic resistance and resistance mechanisms in Campylobacter jejuni and Campylobacter coli. FEMS Microbiol. Lett. 2007, 227, 123–132. [Google Scholar] [CrossRef]
- Bolinger, H.; Kathariou, S. The current state of macrolide resistance in Campylobacter spp.: Trends and impacts of resistance mechanisms. Appl. Environ. Microbiol. 2017, 83, e00416–e00417. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Engberg, J. Antimicrobial resistance of thermophilic Campylobacter. Vet. Res. 2001, 32, 311–321. [Google Scholar] [CrossRef]
- Nielsen, H.; Hansen, K.K.; Gradel, K.O.; Kristensen, B.; Ejlertsen, T.; Østergaard, C.; Schønheyder, H.C. Bacteraemia as a result of Campylobacter species: A population-based study of epidemiology and clinical risk factors. Clin. Microbiol. Infect. 2010, 16, 57–61. [Google Scholar] [CrossRef]
- Payot, S.; Bolla, J.-M.; Corcoran, D.; Fanning, S.; Mégraud, F.; Zhang, Q. Mechanisms of fluoroquinolone and macrolide resistance in Campylobacter spp. Microbes Infect. 2006, 8, 1967–1971. [Google Scholar] [CrossRef]
- Drlica, K.; Zhao, X. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol. Mol. Biol. Rev. 1997, 61, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Klostermeier, D. Why two? On the role of (A-)symmetry in negative supercoiling of DNA by gyrase. Int. J. Mol. Sci. 2018, 19, 1489. [Google Scholar] [CrossRef] [PubMed]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Nasht, H.A. DNA gyrase: An enzyme that introduces superhelical turns into DNA (Escherichia coli/ATP-dependent reaction/superhelix density). Proc. Natl. Acad. Sci. USA 1976, 73, 3872–3876. [Google Scholar] [CrossRef] [PubMed]
- Zirnstein, G.; Li, Y.; Swaminathan, B.; Angulo, F. Ciprofloxacin resistance in Campylobacter jejuni isolates: Detection of gyrA resistance mutations by mismatch amplification mutation assay PCR and DNA sequence analysis. J. Clin. Microbiol. 1999, 37, 3276–3280. [Google Scholar] [CrossRef]
- Siibak, T.; Peil, L.; Xiong, L.; Mankin, A.; Remme, J.; Tenson, T. Erythromycin- and chloramphenicol-induced ribosomal assembly defects are secondary effects of protein synthesis inhibition. Antimicrob. Agents Chemother. 2009, 53, 563–571. [Google Scholar] [CrossRef]
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef]
- Cagliero, C.; Mouline, C.; Cloeckaert, A.; Payot, S. Synergy between efflux pump CmeABC and modifications in ribosomal proteins L4 and L22 in conferring macrolide resistance in Campylobacter jejuni and Campylobacter coli. Antimicrob. Agents Chemother. 2006, 50, 3893–3896. [Google Scholar] [CrossRef]
- Zengel, J.M.; Jerauld, A.; Walker, A.; Wahl, M.C.; Lindahl, L. The extended loops of ribosomal proteins L4 and L22 are not required for ribosome assembly or L4-mediated autogenous control. Rna 2003, 9, 1188–1197. [Google Scholar] [CrossRef]
- Gong, F.; Yanofsky, C. Instruction of translating ribosome by nascent peptide. Science 2002, 297, 1864–1867. [Google Scholar] [CrossRef]
- Hao, H.; Yuan, Z.; Shen, Z.; Han, J.; Sahin, O.; Liu, P.; Zhang, Q. Mutational and transcriptomic changes involved in the development of macrolide resistance in Campylobacter jejuni. Antimicrob. Agents Chemother. 2013, 57, 1369–1378. [Google Scholar] [CrossRef][Green Version]
- Qin, S.; Wang, Y.; Zhang, Q.; Zhang, M.; Deng, F.; Shen, Z.; Wu, C.; Wang, S.; Zhang, J.; Shen, J. Report of ribosomal RNA methylase gene erm(B) in multidrug-resistant Campylobacter coli. J. Antimicrob. Chemother. 2014, 69, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, R. Mechanisms of resistance to macrolides and lincosamides: Nature of the resistance elements and their clinical implications. Clin. Infect. Dis. 2002, 34, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Lin, J.; Reynolds, D.L.; Zhang, Q. Contribution of the multidrug efflux transporter CmeABC to antibiotic resistance in different Campylobacter species. Foodborne Pathog. Dis. 2010, 7, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Gibreel, A.; Wetsch, N.M.; Taylor, D.E. Contribution of the CmeABC efflux pump to macrolide and tetracycline resistance in Campylobacter jejuni. Antimicrob. Agents Chemother. 2007, 51, 3212–3216. [Google Scholar] [CrossRef]
- Yao, H.; Shen, Z.; Wang, Y.; Deng, F.; Liu, D.; Naren, G.; Dai, L.; Su, C.C.; Wang, B.; Wang, S.; et al. Emergence of a potent multidrug efflux pump variant that enhances Campylobacter resistance to multiple antibiotics. MBio 2016, 7, e01543-16. [Google Scholar] [CrossRef]
- Pumbwe, L.; Piddock, L.J.V. Identification and molecular characterisation of CmeB, a Campylobacter jejuni multidrug efflux pump. FEMS Microbiol. Lett. 2002, 206, 185–189. [Google Scholar] [CrossRef]
- Pérez-Boto, D.; López-Portolés, J.A.; Simón, C.; Valdezate, S.; Echeita, M.A. Study of the molecular mechanisms involved in high-level macrolide resistance of Spanish Campylobacter jejuni and Campylobacter coli strains. J. Antimicrob. Chemother. 2010, 65, 2083–2088. [Google Scholar] [CrossRef]
- Cagliero, C.; Maurel, M.-C.; Cloeckaert, A.; Payot, S. Regulation of the expression of the CmeABC efflux pump in Campylobacter jejuni: Identification of a point mutation abolishing the binding of the CmeR repressor in an in vitro -selected multidrug-resistant mutant. FEMS Microbiol. Lett. 2007, 267, 89–94. [Google Scholar] [CrossRef]
- Cagliero, C.; Mouline, C.; Payot, S.; Cloeckaert, A. Involvement of the CmeABC efflux pump in the macrolide resistance of Campylobacter coli. J. Antimicrob. Chemother. 2005, 56, 948–950. [Google Scholar] [CrossRef]
- Connell, S.R.; Trieber, C.A.; Dinos, G.P.; Einfeldt, E.; Taylor, D.E.; Nierhaus, K.H. Mechanism of Tet(O)-mediated tetracycline resistance. EMBO J. 2003, 22, 945–953. [Google Scholar] [CrossRef]
- Roberts, M.C. Tetracycline Therapy: Update. Clin. Infect. Dis. 2003, 36, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef]
- Warburton, P.J.; Amodeo, N.; Roberts, A.P. Mosaic tetracycline resistance genes encoding ribosomal protection proteins. J. Antimicrob. Chemother. 2016, 71, 3333–3339. [Google Scholar] [CrossRef] [PubMed]
- Gibreel, A.; Tracz, D.M.; Nonaka, L.; Ngo, T.M.; Connell, S.R.; Taylor, D.E. Incidence of antibiotic resistance in Campylobacter jejuni isolated in Alberta, Canada, from 1999 to 2002, with special reference to tet(O)-mediated tetracycline resistance. Antimicrob. Agents Chemother. 2004, 48, 3442–3450. [Google Scholar] [CrossRef] [PubMed]
- Pratt, A.; Korolik, V. Tetracycline resistance of Australian Campylobacter jejuni and Campylobacter coli isolates. J. Antimicrob. Chemother. 2005, 55, 452–460. [Google Scholar] [CrossRef]
- Friis, L.M.; Pin, C.; Taylor, D.E.; Pearson, B.M.; Wells, J.M. A role for the tet(O) plasmid in maintaining Campylobacter plasticity. Plasmid 2007, 57, 18–28. [Google Scholar] [CrossRef]
- Kemper, N. Veterinary antibiotics in the aquatic and terrestrial environment. Ecol. Indic. 2008, 8, 1–13. [Google Scholar] [CrossRef]
- Crespo, M.D.; Altermann, E.; Olson, J.; Miller, W.G.; Chandrashekhar, K.; Kathariou, S. Novel plasmid conferring kanamycin and tetracycline resistance in the turkey-derived Campylobacter jejuni strain 11601MD. Plasmid 2016, 86, 32–37. [Google Scholar] [CrossRef]
- Stanton, T.B.; McDowall, J.S.; Rasmussen, M.A. Diverse tetracycline resistance genotypes of Megasphaera elsdenii strains selectively cultured from swine feces. Appl. Environ. Microbiol. 2004, 70, 3754–3757. [Google Scholar] [CrossRef][Green Version]
- Patterson, A.J.; Rincon, M.T.; Flint, H.J.; Scott, K.P. Mosaic tetracycline resistance genes are widespread in human and animal fecal samples. Antimicrob. Agents Chemother. 2007, 51, 1115–1118. [Google Scholar] [CrossRef][Green Version]
- Melville, C.M.; Scott, K.P.; Mercer, D.K.; Flint, H.J. Novel tetracycline resistance gene, tet(32), in the Clostridium-related human colonic anaerobe K10 and its transmission in vitro to the rumen anaerobe Butyrivibrio fibrisolvens. Antimicrob. Agents Chemother. 2001, 45, 3246–3249. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Freitas, A.R.; Silveira, E.; Baquero, F.; Peixe, L.; Roberts, A.P.; Coque, T.M. A tet(S/M) hybrid from CTn6000 and CTn916 recombination. Microbiology. 2012, 158, 2710–2711. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed]
- Kotra, L.P.; Haddad, J.; Mobashery, S. Aminoglycosides: Perspectives on mechanisms of action and resistance and strategies to counter resistance. Antimicrob. Agents Chemother. 2000, 44, 3249–3256. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef]
- Novick, R.P.; Clowes, R.C.; Cohen, S.N.; Curtiss, R.I.; Datta, N.; Falkow, S. Uniform nomenclature for bacterial plasmids: A proposal. Bacteriol. Rev. 1976, 40, 168–189. [Google Scholar] [CrossRef]
- Shaw, K.J.; Rather, P.N.; Hare, R.S.; Miller, G.H. Molecular genetics of aminoglycoside resistance genes and familial relationships of the aminoglycoside-modifying enzymes. Microbiol. Rev. 1993, 57, 138–163. [Google Scholar] [CrossRef]
- Vanhoof, R.; Hannecart-Pokorni, E.; Content, J. Nomenclature of genes encoding aminoglycoside-modifying enzymes. Antimicrob. Agents Chemother. 1998, 42, 483. [Google Scholar] [CrossRef]
- Abril, C.; Brodard, I.; Perreten, V. Two novel antibiotic resistance genes, tet(44) and ant(6)-Ib, are located within a transferable pathogenicity island in Campylobacter fetus subsp. fetus. Antimicrob. Agents Chemother. 2010, 54, 3052–3055. [Google Scholar] [CrossRef]
- Hormeño, L.; Ugarte-Ruiz, M.; Palomo, G.; Borge, C.; Florez-Cuadrado, D.; Vadillo, S.; Píriz, S.; Domínguez, L.; Campos, M.J.; Quesada, A. ant(6)-I genes encoding aminoglycoside O-nucleotidyltransferases are widely spread among streptomycin resistant strains of Campylobacter jejuni and Campylobacter coli. Front. Microbiol. 2018, 9, 2515. [Google Scholar] [CrossRef]
- Werner, G.; Hildebrandt, B.; Witte, W. Linkage of erm(B) and aadE-sat4-aphA-3 in multiple-resistant Enterococcus faecium isolates of different ecological origins. Microb. Drug Resist. 2003, 9 (Suppl. 1), S9–S16. [Google Scholar] [CrossRef] [PubMed]
- Nirdnoy, W.; Mason, C.J.; Guerry, P. Mosaic structure of a multiple-drug-resistant, conjugative plasmid from Campylobacter jejuni. Antimicrob. Agents Chemother. 2005, 49, 2454–2459. [Google Scholar] [CrossRef]
- Lucey, B.; Cryan, B.; O’Halloran, F.; Wall, P.G.; Buckley, T.; Fanning, S. Trends in antimicrobial susceptibility among isolates of Campylobacter species in Ireland and the emergence of resistance to ciprofloxacin. Vet. Rec. 2002, 151, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Bolton, D.; Patriarchi, A.; Fox, Á.; Fanning, S. A study of the molecular basis of quinolone and macrolide resistance in a selection of Campylobacter isolates from intensive poultry flocks. Food Control. 2013, 30, 222–226. [Google Scholar] [CrossRef]
- Lucey, B.; Crowley, D.; Moloney, P.; Cryan, B.; Daly, M.; O’ Halloran, F.; Threlfall, E.J.; Fanning, S. Integronlike structures in Campylobacter spp. of human and animal origin. Emerg. Infect. Dis. 2000, 6, 50–55. [Google Scholar]
- Corcoran, D.; Quinn, T.; Cotter, L.; O’Halloran, F.; Fanning, S. Characterization of a cmeABC operon in a quinolone-resistant Campylobacter coli isolate of Irish origin. Microb. Drug Resist. 2005, 11, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Oza, A.N.; McKenna, J.P.; McDowell, S.W.J.; Menzies, F.D.; Neill, S.D. Antimicrobial susceptibility of Campylobacter spp. isolated from broiler chickens in Northern Ireland. J. Antimicrob. Chemother. 2003, 52, 220–223. [Google Scholar] [CrossRef][Green Version]
- Moore, J.E.; Mclernon, P.; Wareing, D.; Xu, J.; Murphy, P.G.; Mclernon, P.; Hospital, R.P.; Lane, S.G.; Pr, P. Characterisation of resistant Campylobacter species isolated from human beings and chickens. Vet. Rec. 2002, 150, 518–520. [Google Scholar] [CrossRef]
- Redondo, N.; Carroll, A.; Mcnamara, E. Molecular characterization of Campylobacter causing human clinical infection using whole-genome sequencing: Virulence, antimicrobial resistance and phylogeny in Ireland. PLoS ONE 2019, 14, e0219088. [Google Scholar] [CrossRef]
- Corcoran, D.; Quinn, T.; Cotter, L.; Fanning, S. An investigation of the molecular mechanisms contributing to high-level erythromycin resistance in Campylobacter. Int. J. Antimicrob. Agents 2006, 27, 40–45. [Google Scholar] [CrossRef]
- Acke, E.; McGill, K.; Quinn, T.; Jones, B.R.; Fanning, S.; Whyte, P. Antimicrobial resistance profiles and mechanisms of resistance in Campylobacter jejuni isolates from pets. Foodborne Pathog. Dis. 2009, 6, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Minihan, D.; Whyte, P.; O’Mahony, M.; Cowley, D.; O’Halloran, F.; Corcoran, D.; Fanning, S.; Collins, J.D. Phenotypic and genotypic anti-microbial resistance profiles of Campylobacters from untreated feedlot cattle and their environment. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2006, 53, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Kang, M. Molecular basis of macrolide resistance in Campylobacter strains isolated from poultry in South Korea. Biomed. Res. Int. 2018, 2018, 4526576. [Google Scholar] [CrossRef] [PubMed]
- Olkkola, S.; Culebro, A.; Juntunen, P.; Hänninen, M.L.; Rossi, M.M. Functional genomics in Campylobacter coli identified a novel streptomycin resistance gene located in a hypervariable genomic region. Microbiology 2016, 162, 1157–1166. [Google Scholar] [CrossRef]
- Fabre, A.; Oleastro, M.; Nunes, A.; Santos, A.; Ducournau, A.; Floch, P.; Lehours, P. Whole-genome sequence analysis of multidrug-resistant Campylobacter isolates: A focus on aminoglycoside resistance. J. Clin. Microbiol. 2018, 56, e00390-e18. [Google Scholar] [CrossRef]
- Lynch, C.T.; Lynch, H.; Egan, J.; Whyte, P.; Bolton, D.; Coffey, A.; Lucey, B. Antimicrobial resistance of Campylobacter isolates recovered from broilers in the Republic of Ireland in 2017 and 2018: An update. Br. Poult. Sci. 2020. [Google Scholar] [CrossRef]
- Moore, J.E.; Corcoran, D.; Dooley, J.S.G.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Megraud, F.; Millar, C.B.; O’ Mahony, R.; et al. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, M.; Zhou, J.; Pang, L.; Wang, G.; Hou, F. The molecular mechanisms of ciprofloxacin resistance in clinical Campylobacter jejuni and their genotyping characteristics in Beijing, China. Foodborne Pathog. Dis. 2017, 14, 386–392. [Google Scholar] [CrossRef]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Kielsznia, A.; Korzekwa, K.; Tobiasz, A.; Korzeniowska-Kowal, A.; Wieliczko, A. High prevalence of resistance to fluoroquinolones and tetracycline Campylobacter spp. isolated from poultry in Poland. Microb. Drug Resist. 2018, 24, 314–322. [Google Scholar] [CrossRef]
- Sierra-Arguello, Y.M.; Perdoncini, G.; Morgan, R.B.; Salle, C.T.P.; Moraes, H.L.S.; Gomes, M.J.P.; do Nascimento, V.P. Fluoroquinolone and macrolide resistance in Campylobacter jejuni isolated from broiler slaughterhouses in southern Brazil. Avian Pathol. 2016, 45, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Álvarez-Ordóñez, A.; Duarte, A.; Dierick, K.; Botteldoorn, N. Genetic basis and clonal population structure of antibiotic resistance in Campylobacter jejuni isolated from broiler carcasses in Belgium. Front. Microbiol. 2018, 9, 1014. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Pereira, S.; Sahin, O.; Lin, J.; Huang, S.; Michel, L.; Zhang, Q. Enhanced in vivo fitness of fluoroquinolone-resistant Campylobacter jejuni in the absence of antibiotic selection pressure. Proc. Natl. Acad. Sci. USA 2005, 102, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lin, J.; Pereira, S. Fluoroquinolone-resistant Campylobacter in animal reservoirs: Dynamics of development, resistance mechanisms and ecological fitness. Anim. Heal. Res. Rev. 2003, 4, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Whelan, M.X.; Ardill, L.; Koide, K.; Nakajima, C.; Suzuki, Y.; Simpson, J.C.; Ó Cróinín, T. Acquisition of fluoroquinolone resistance in Campylobacter jejuni leads to an increase in biofilm formation and virulence. Sci. Rep. 2019, 9, 18216. [Google Scholar] [CrossRef] [PubMed]
- Lucey, B.; Feurer, C.; Greer, P.; Moloney, P.; Cryan, B.; Fanning, S. Antimicrobial resistance profiling and DNA Amplification Fingerprinting (DAF) of thermophilic Campylobacter spp. in human, poultry and porcine samples from the Cork region of Ireland. J. Appl. Microbiol. 2000, 89, 727–734. [Google Scholar] [CrossRef]
- Health Products Regulatory Authority. Report on Consumption of Veterinary Antibiotics in Ireland during 2017. Available online: http://www.hpra.ie/docs/default-source/default-document-library/draft-report-on-consumption-of-veterinary-antibiotics-in-ireland-during-2017.pdf?sfvrsn=0 (accessed on 1 April 2020).
- Health Products Regulatory Authority. Report on consumption of veterinary antibiotics in Ireland during 2014. 2014. Available online: https://www.hpra.ie/docs/default-source/publications-forms/newsletters/report-on-consumption-of-veterinary-antibiotics-in-ireland-during-2014.pdf?sfvrsn=9 (accessed on 1 April 2020).
- Health Products Regulatory Authority. Report on Consumption of Veterinary Antibiotics in Ireland During 2018. Available online: https://www.hpra.ie/docs/default-source/default-document-library/report_consumption_vet_antimicrobials_2011.pdf?sfvrsn=0 (accessed on 1 April 2020).
- Ekkapobyotin, C.; Padungtod, P.; Chuanchuen, R. Antimicrobial resistance of Campylobacter coli isolates from swine. Int. J. Food Microbiol. 2008, 128, 525–528. [Google Scholar] [CrossRef]
- Kinana, A.D.; Cardinale, E.; Bahsoun, I.; Tall, F.; Sire, J.M.; Garin, B.; Boye, C.-B.; Dromigny, J.-A.; Perrier-Gros-Claude, J.-D. Analysis of topoisomerase mutations in fluoroquinolone-resistant and -susceptible Campylobacter jejuni strains isolated in Senegal. Int. J. Antimicrob. Agents 2007, 29, 397–401. [Google Scholar] [CrossRef]
- Dionisi, A.M.; Luzzi, I.; Carattoli, A. Identification of ciprofloxacin-resistant Campylobacter jejuni and analysis of the gyrA gene by the LightCycler mutation assay. Mol. Cell. Probes 2004, 18, 255–261. [Google Scholar] [CrossRef]
- Ragimbeau, C.; Colin, S.; Devaux, A.; Decruyenaere, F.; Cauchie, H.M.; Losch, S.; Penny, C.; Mossong, J. Investigating the host specificity of Campylobacter jejuni and Campylobacter coli by sequencing gyrase subunit A. BMC Microbiol. 2014, 14, 205. [Google Scholar] [CrossRef]
- Ruiz, J.; Moreno, A.; Jimenez de Anta, M.T.; Vila, J. A double mutation in the gyrA gene is necessary to produce high levels of resistance to moxifloxacin in Campylobacter spp. clinical isolates. Int. J. Antimicrob. Agents 2005, 25, 542–545. [Google Scholar] [CrossRef]
- Bachoual, R.; Ouabdesselam, S.; Mory, F.; Lascols, C.; Soussy, C.-J.; Tankovic, J. Single or double mutational alterations of GyrA associated with fluoroquinolone resistance in Campylobacter jejuni and Campylobacter coli. Microb. Drug Resist. 2001, 7, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Tankovic, J.; Bachoual, R.; Ouabdesselam, S.; Boudjadja, A.; Soussy, C.-J. In-vitro activity of moxifloxacin against fluoroquinolone-resistant strains of aerobic Gram-negative bacilli and Enterococcus faecalis. J. Antimicrob. Chemother. 1999, 43, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.V.; Ricci, V.; Pumbwe, L.; Everett, M.J.; Griggs, D.J. Fluoroquinolone resistance in Campylobacter species from man and animals: Detection of mutations in topoisomerase genes. J. Antimicrob. Chemother. 2003, 51, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hakanen, A.; Jalava, J.; Kotilainen, P.; Jousimies-Somer, H.; Siitonen, A.; Huovinen, P. gyrA polymorphism in Campylobacter jejuni: Detection of gyrA mutations in 162 C. jejuni isolates by single-strand conformation polymorphism and DNA sequencing. Antimicrob. Agents Chemother. 2002, 46, 2644–2647. [Google Scholar] [CrossRef]
- Ge, B.; White, D.G.; Mcdermott, P.F.; Girard, W.; Zhao, S.; Hubert, S.; Meng, J. Antimicrobial-Resistant Campylobacter Species from Retail Raw Meats. Appl. Environ. Microbiol. 2003, 69, 3005–3007. [Google Scholar] [CrossRef]
- Thomrongsuwannakij, T.; Blackall, P.J.; Chansiripornchai, N. A study on Campylobacter jejuni and Campylobacter coli through commercial broiler production chains in Thailand. Avian Dis. 2017, 61, 186–197. [Google Scholar] [CrossRef]
- Oishi, A.; Murakami, K.; Etoh, Y.; Sera, N.; Horikawa, K. Antimicrobial susceptibility and resistance mutations in Campylobacter jejuni and C. coli isolates from human and meat sources. Kansenshogaku Zasshi. 2015, 89, 244–253. [Google Scholar] [CrossRef]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef]
- Chittum, H.S.; Champney, W.S. Ribosomal protein gene sequence changes in erythromycin-resistant mutants of Escherichia coli. J. Bacteriol. 1994, 176, 6192–6198. [Google Scholar] [CrossRef]
- Florez-Cuadrado, D.; Ugarte-Ruiz, M.; Quesada, A.; Palomo, G.; Domínguez, L.; Porrero, M.C. Description of an erm(B)-carrying Campylobacter coli isolate in Europe. J. Antimicrob. Chemother. 2016, 71, 841–843. [Google Scholar] [CrossRef]
- Florez-Cuadrado, D.; Ugarte-Ruiz, M.; Meric, G.; Quesada, A.; Porrero, M.C.; Pascoe, B.; Sáez-Llorente, J.L.; Orozco, G.L.; Domínguez, L.; Sheppard, S.K. Genome comparison of erythromycin resistant campylobacter from Turkeys identifies hosts and pathways for horizontal spread of erm(B) genes. Front. Microbiol. 2017, 8, 2240. [Google Scholar] [CrossRef] [PubMed]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Álvarez-Ordóñez, A.; Dierick, K.; Botteldoorn, N. Molecular epidemiology and antimicrobial resistance mechanisms of Campylobacter coli from diarrhoeal patients and broiler carcasses in Belgium. Transbound. Emerg. Dis. 2019, 66, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Tagg, K.A.; Joung, Y.J.; Bennett, C.; Watkins, L.F.; Eikmeier, D.; Folster, J.P. Report of erm(B) Campylobacter jejuni in the United States. Antimicrob. Agents Chemother. 2018, 62, e02615-17. [Google Scholar] [CrossRef] [PubMed]
- Igwaran, A.; Okoh, A.L. Campylobacteriosis agents in meat carcasses collected from two district municipalities in the Eastern cape province, South Africa. Foods 2020, 9, 203. [Google Scholar] [CrossRef]
- Luangtongkum, T.; Shen, Z.; Seng, V.W.; Sahin, O.; Jeon, B.; Liu, P.; Zhang, Q. Impaired fitness and transmission of macrolide-resistant Campylobacter jejuni in its natural host. Antimicrob. Agents Chemother. 2012, 56, 1300–1308. [Google Scholar] [CrossRef]
- Han, F.; Pu, S.; Wang, F.; Meng, J.; Ge, B. Fitness cost of macrolide resistance in Campylobacter jejuni. Int. J. Antimicrob. Agents 2009, 34, 462–466. [Google Scholar] [CrossRef]
- Zeitouni, S.; Collin, O.; Andraud, M.; Ermel, G.; Kempf, I. Fitness of macrolide resistant Campylobacter coli and Campylobacter jejuni. Microb. Drug Resist. 2012, 18, 101–108. [Google Scholar] [CrossRef]
- Stanton, T.B.; Humphrey, S.B. Isolation of tetracycline-resistant Megasphaera elsdenii strains with novel mosaic gene combinations of tet(O) and tet(W) from swine. Appl. Environ. Microbiol. 2003, 69, 3874–3882. [Google Scholar] [CrossRef]
- Aminov, R.I.; Garrigues-Jeanjean, N.; Mackie, R.I. Molecular ecology of tetracycline resistance: Development and validation of primers for detection of tetracycline resistance genes encoding ribosomal protection proteins. Appl. Environ. Microbiol. 2001, 67, 22–32. [Google Scholar] [CrossRef]
- Mortensen, N.P.; Kuijf, M.L.; Ang, C.W.; Schiellerup, P.; Krogfelt, K.A.; Jacobs, B.C.; van Belkum, A.; Endtz, H.P.; Bergman, M.P. Sialylation of Campylobacter jejuni lipo-oligosaccharides is associated with severe gastro-enteritis and reactive arthritis. Microbes Infect. 2009, 11, 988–994. [Google Scholar] [CrossRef]
- Dearlove, B.L.; Cody, A.J.; Pascoe, B.; Méric, G.; Wilson, D.J.; Sheppard, S.K. Rapid host switching in generalist Campylobacter strains erodes the signal for tracing human infections. ISME J. 2016, 10, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Llarena, A.K.; Zhang, J.; Vehkala, M.; Välimäki, N.; Hakkinen, M.; Hänninen, M.L.; Roasto, M.; Mäesaar, M.; Taboada, E.; Barker, D.; et al. Monomorphic genotypes within a generalist lineage of Campylobacter jejuni show signs of global dispersion. Microb. Genom. 2016, 2, e000088. [Google Scholar] [CrossRef] [PubMed]
- Mohan, V.; Stevenson, M.; Marshall, J.; Fearnhead, P.; Holland, B.R.; Hotter, G.; French, N.P. Campylobacter jejuni colonization and population structure in urban populations of ducks and starlings in New Zealand. Microbiologyopen 2013, 2, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Kang, M.; Jang, H.K. Genetic characterization and epidemiological implications of Campylobacter isolates from wild birds in South Korea. Transbound. Emerg. Dis. 2019, 66, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Mäesaar, M.; Meremäe, K.; Ivanova, M.; Roasto, M. Antimicrobial resistance and multilocus sequence types of Campylobacter jejuni isolated from Baltic broiler chicken meat and Estonian human patients. Poult. Sci. 2018, 97, 3645–3651. [Google Scholar] [CrossRef] [PubMed]
- Aksomaitiene, J.; Ramonaite, S.; Tamuleviciene, E.; Novoslavskij, A.; Alter, T.; Malakauskas, M. Overlap of antibiotic resistant Campylobacter jejuni MLST genotypes isolated from humans, broiler products, dairy cattle and wild birds in Lithuania. Front. Microbiol. 2019, 10, 1377. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Deng, F.; Shen, Z.; Wu, C.; Zhang, J.; Zhang, Q.; Shen, J. Emergence of multidrug-resistant Campylobacter species isolates with a horizontally acquired rRNA methylase. Antimicrob. Agents Chemother. 2014, 58, 5405–5412. [Google Scholar] [CrossRef]
- Burckhardt, R.M.; Escalante-Semerena, J.C. Insights into the function of the N-acetyltransferase SatA that detoxifies streptothricin in Bacillus subtilis and Bacillus anthracis. Appl. Environ. Microbiol. 2019, 85, e03029-18. [Google Scholar] [CrossRef]
- Qin, S.; Wang, Y.; Zhang, Q.; Chen, X.; Shen, Z.; Deng, F.; Wu, C.; Shen, J. Identification of a novel genomic island conferring resistance to multiple aminoglycoside antibiotics in Campylobacter coli. Antimicrob. Agents Chemother. 2012, 56, 5332–5339. [Google Scholar] [CrossRef] [PubMed]
- PubMLST. Available online: https://pubmlst.org/ (accessed on 24 February 2020).
- Wright, S.; Wilson, S.; Miller, W.G.; Mandrell, R.E.; Siletzky, R.M.; Kathariou, S. Differences in methylation at GATC sites in genomic DNA of Campylobacter coli from turkeys and swine. Appl. Environ. Microbiol. 2010, 76, 7314–7317. [Google Scholar] [CrossRef] [PubMed]
- Amadi, V.A.; Matthew-Belmar, V.; Subbarao, C.; Kashoma, I.; Rajashekara, G.; Sharma, R.; Hariharan, H.; Stone, D. Campylobacter species isolated from pigs in Grenada exhibited novel clones: Genotypes and antimicrobial resistance profiles of sequence types. Foodborne Pathog. Dis. 2017, 14, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Cody, A.J.; McCarthy, N.D.; Van Rensburg, M.J.; Isinkaye, T.; Bentley, S.D.; Parkhill, J.; Dingle, K.E.; Bowler, I.C.J.W.; Jolley, K.A.; Maiden, M.C.J. Real-time genomic epidemiological evaluation of human Campylobacter isolates by use of whole-genome multilocus sequence typing. J. Clin. Microbiol. 2013, 51, 2526–2534. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. EU Protocol for Harmonised Monitoring of Antimicrobial Resistance in Human Salmonella and Campylobacter Isolates. Available online: https://www.ecdc.europa.eu/sites/default/files/media/en/publications/Publications/antimicrobial-resistance-Salmonella-Campylobacter-harmonised-monitoring.pdf (accessed on 7 June 2016).
- European Commission. Commission Implementing Decision of 12 November 2013 on the monitoring and reporting of antimicrobial resistance in zoonotic and commensal bacteria. Off. J. Eur. Union 2013, L 303, 26–39. [Google Scholar]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Vacher, S.; Ménard, A.; Bernard, E.; Mégraud, F. PCR-restriction fragment length polymorphism analysis for detection of point mutations associated with macrolide resistance in Campylobacter spp. Antimicrob. Agents Chemother. 2003, 47, 1125–1128. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Godschalk, P.C.R.; Kuijf, M.L.; Li, J.; Michael, F.S.; Ang, C.W.; Jacobs, B.C.; Karwaski, M.-F.; Brochu, D.; Moterassed, A.; Endtz, H.P.; et al. Structural characterization of Campylobacter jejuni lipooligosaccharide outer cores associated with Guillain-Barré and Miller Fisher Syndromes. Infect. Immun. 2007, 75, 1245–1254. [Google Scholar] [CrossRef]
- Guyard-Nicodème, M.; Rivoal, K.; Houard, E.; Rose, V.; Quesne, S.; Mourand, G.; Rouxel, S.; Kempf, I.; Guillier, L.; Gauchard, F.; et al. Prevalence and characterization of Campylobacter jejuni from chicken meat sold in French retail outlets. Int. J. Food Microbiol. 2015, 203, 8–14. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Genome analysis Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. Genome analysis QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).