

Green Tea Extract (Theaceae; Camellia sinensis L.): A Promising Antimicrobial, Anti-Quorum Sensing and Antibiofilm Candidate Against Multidrug-Resistant Campylobacter Species
 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Prevalence of Campylobacter Species in Animal, Environmental and Human Sources
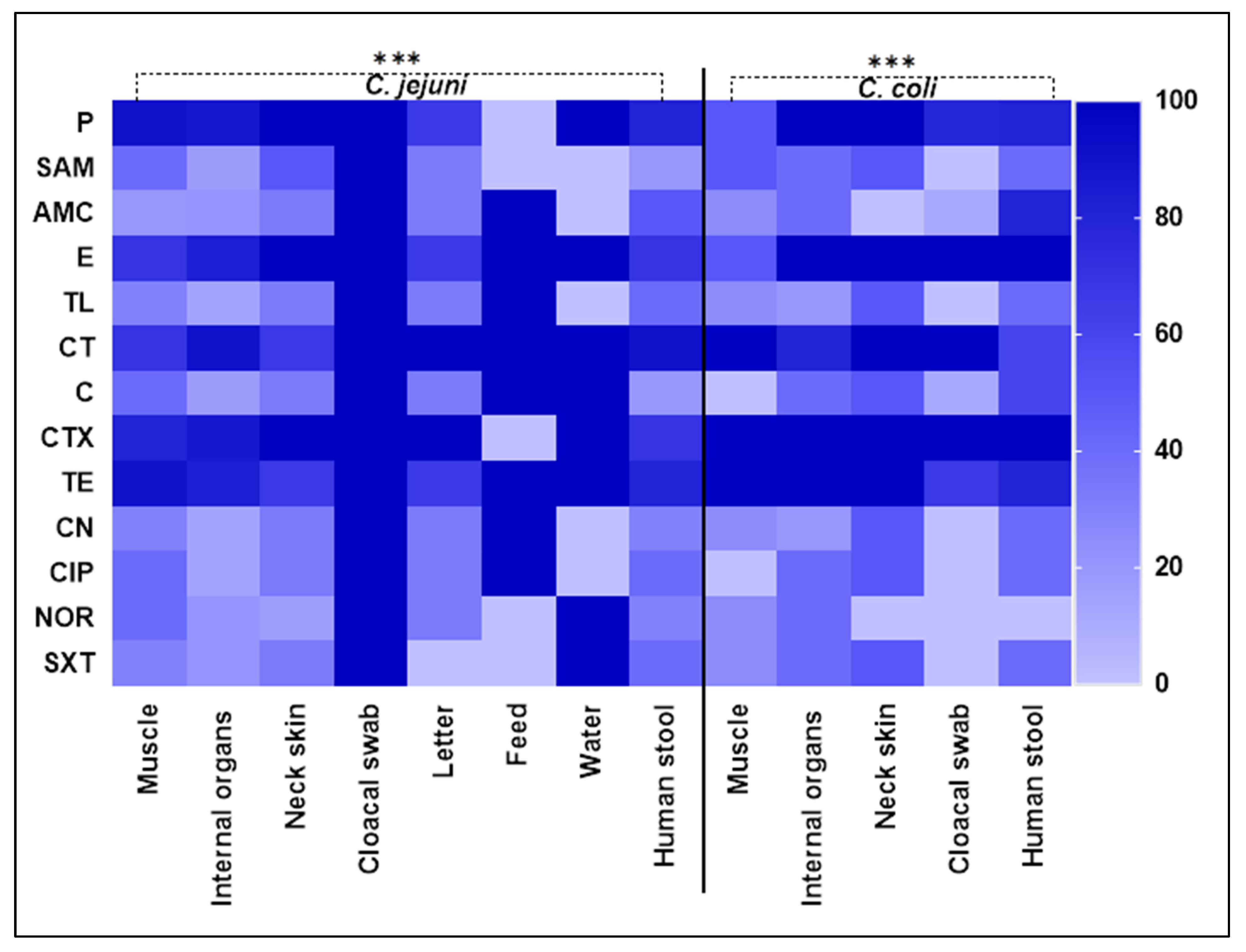
2.2. Antimicrobial Susceptibility Testing of Campylobacter Species
2.3. Biofilm Formation by Multidrug-Resistant Campylobacter Isolates
2.4. Antimicrobial Activities of Green Tea, Rosemary and Ginger Extracts Against MDR Campylobacter Species
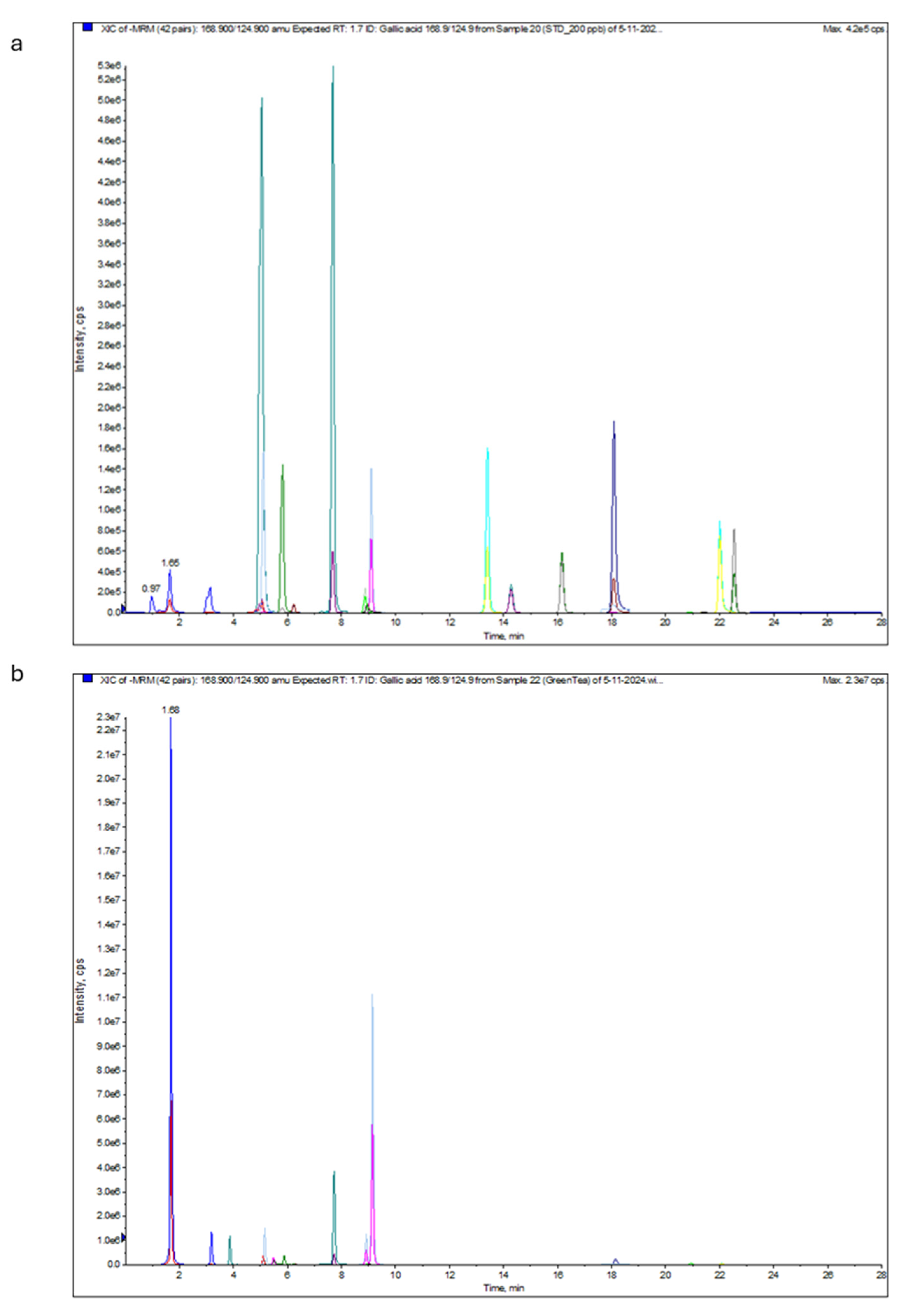
2.5. Characterization of Green Tea Extract Using HPLC
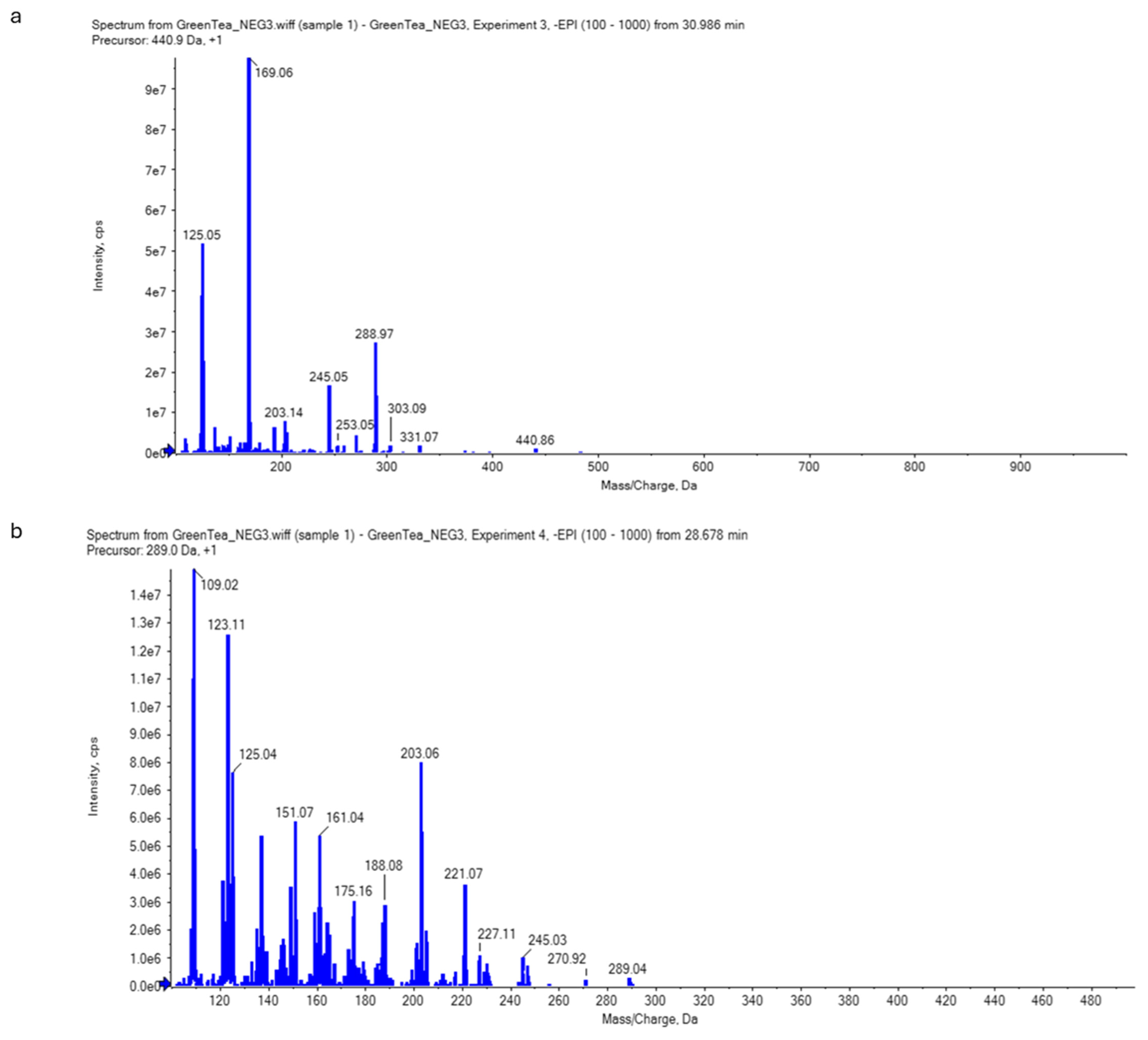
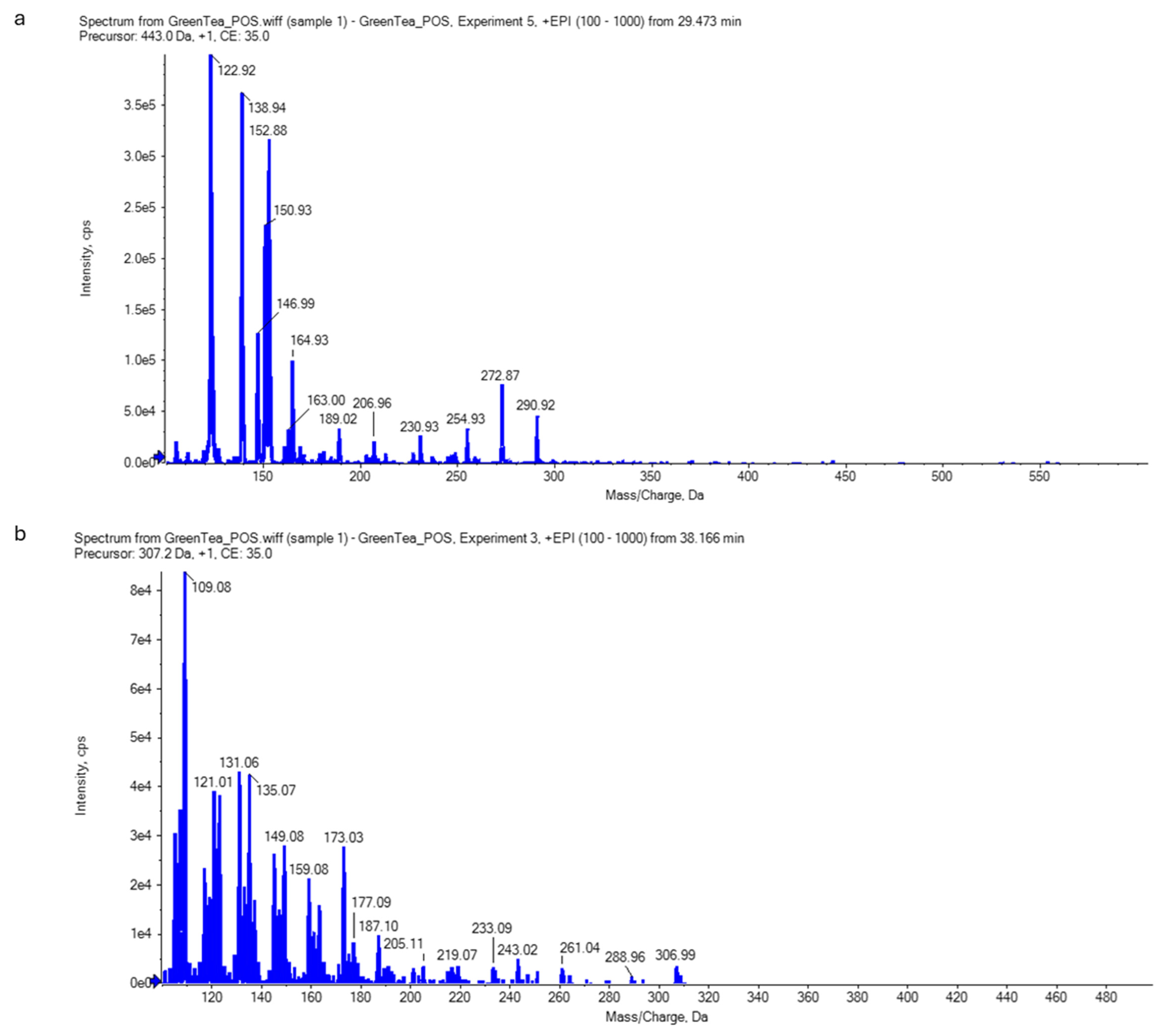
2.6. Analysis of Green Tea Extract Using Liquid Chromatography–Electrospray Ionization–Tandem Mass Spectrometry (LC-ESI-MS/MS)
2.6.1. Total Phenolic and Flavonoid Contents of Green Tea
2.6.2. Catechins of Green Tea Extract
2.7. Antibiofilm Activity of Green Tea Extract Against Campylobacter Species
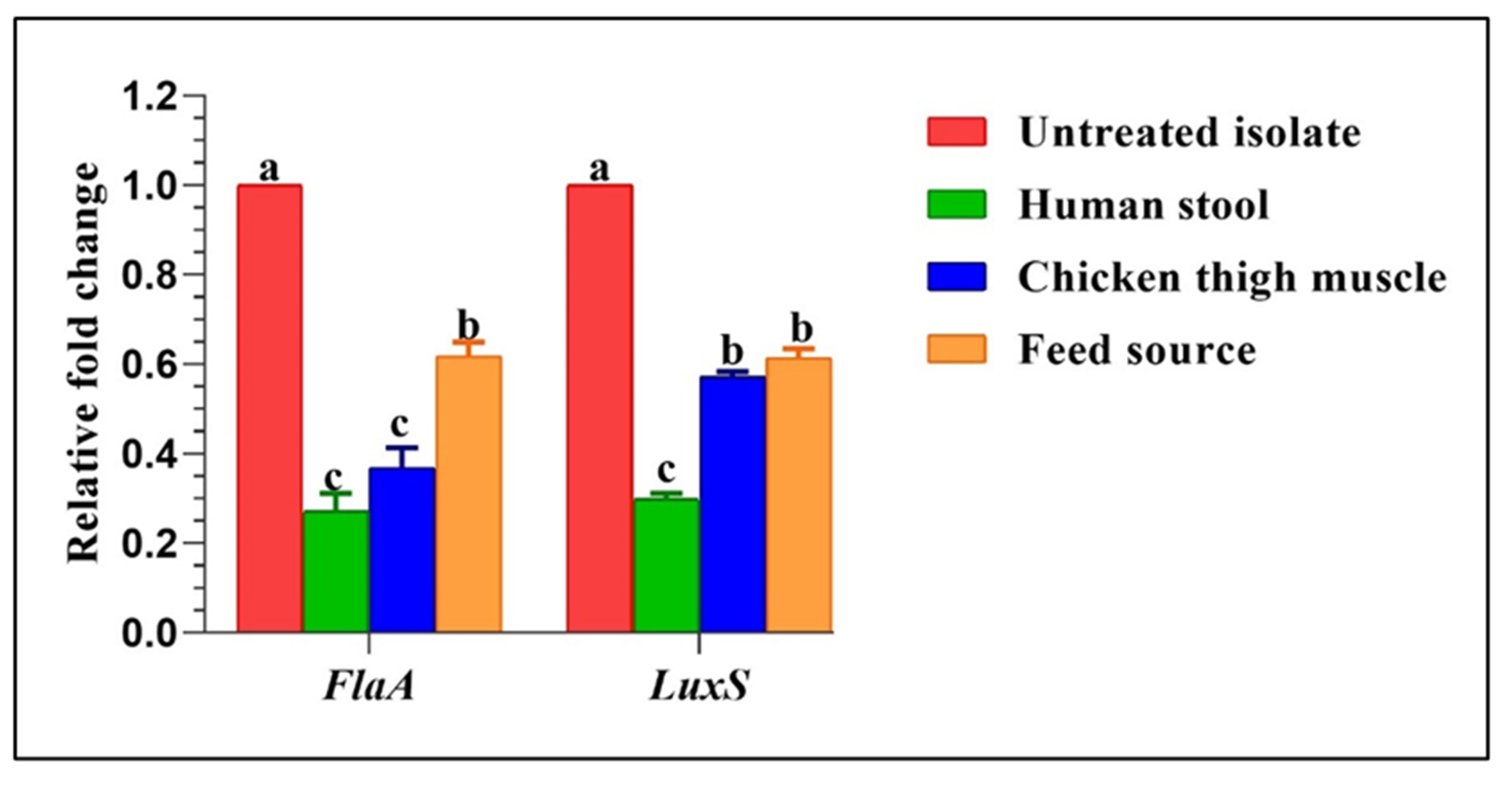
2.8. Transcriptional Changes of Biofilm Genes Post Treatment by Green Tea Extract
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Isolation and Identification of Thermophilic Campylobacter Species
4.3. Molecular Confirmation of Campylobacter Species
4.4. Antimicrobial Susceptibility Testing
4.5. Quantitative Assessment of Biofilm Formation by Campylobacter Species
4.6. Plant Extracts and Their Preparation
4.7. Antimicrobial Activities of the Plant Extracts
4.8. Antibiofilm Assay
4.9. High-Performance Liquid Chromatography (HPLC) Analysis of Green Tea Extract
4.10. Green Tea Analysis Using Liquid Chromatography–Electrospray Ionization–Tandem Mass Spectrometry (LC-ESI-MS/MS) [77]
4.10.1. Quantitative Analysis of the Total Phenolic and Flavonoid Contents of Green Tea Extract
4.10.2. Non-Targeted Screening for Catechins (Qualitative Analysis)
4.11. Reverse Transcriptase Quantitative Polymerase Chain Reaction (RT-qPCR)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization (WHO). Campylobacter. World Health Organisation Web Page. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/campylobacter (accessed on 22 May 2021).
- Salehi, M.; Shafaei, E.; Bameri, Z.; Zahedani, S.S.; Bokaeian, M.; Mirzaee, B.; Mirfakhraee, S.; Rigi, T.B.; Akbari, M. Prevalence and antimicrobial resistance of Campylobacter jejuni. Int. J. Infect. 2014, 1, 2. [Google Scholar] [CrossRef]
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Asakura, H.; Yamasaki, M.; Yamamoto, S.; Igimi, S. Deletion of peb4 gene impairs cell adhesion and biofilm formation in Campylobacter jejuni. FEMS Microbiol. Lett. 2007, 275, 278–285. [Google Scholar] [CrossRef]
- Elvers, K.T.; Park, S.F. Quorum sensing in Campylobacter jejuni: Detection of a luxS encoded signalling molecule. Microbiology 2002, 148 Pt 5, 1475–1481. [Google Scholar] [CrossRef]
- Kalmokoff, M.; Lanthier, P.; Tremblay, T.L.; Foss, M.; Lau, P.C.; Sanders, G.; Austin, J.; Kelly, J.; Szymanski, C.M. Proteomic analysis of Campylobacter jejuni 11,168 biofilms reveal a role for the motility complex in biofilm formation. J. Bacteriol. 2006, 188, 4312–4320. [Google Scholar] [CrossRef]
- Kim, J.S.; Park, C.; Kim, Y.J. Role of flgA for flagellar biosynthesis and biofilm formation of Campylobacter jejuni NCTC11168. J. Microbiol. Biotechnol. 2015, 25, 1871–1879. [Google Scholar] [CrossRef]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef]
- Silva, W.C.; Targino, B.N.; Gonçalves, A.G.; Silva, M.R.; Hungaro, H.M. Campylobacter: An important food safety issue. In Food Safety and Preservation; Elsevier: Amsterdam, The Netherlands, 2018; pp. 391–430. [Google Scholar]
- Ammar, A.M.; El-Hamid, A.; Marwa, I.; El-Malt, R.M.S.; Azab, D.S.; Albogami, S.; Al-Sanea, M.M.; Soliman, W.E.; Ghoneim, M.M.; Bendary, M.M. Molecular detection of fluoroquinolone resistance among multidrug-, extensively drug-, and pan-drug-resistant Campylobacter species in Egypt. Antibiotics 2021, 10, 1342. [Google Scholar] [CrossRef]
- Roua, M.A.; Altuwijri, L.A.; Aldosari, N.S.; Alsakabi, N.; Dawoud, T.M. Antimicrobial and synergistic properties of green tea catechins against microbial pathogens. J. King Saud Univ. Sci. 2024, 36, 103277. [Google Scholar]
- Alimohammadi, Z.; Shirzadi, H.; Taherpour, K.; Rahmatnejad, E.; Khatibjoo, A. Effects of cinnamon, rosemary and oregano on growth performance, blood biochemistry, liver enzyme activities, excreta microbiota and ileal morphology of Campylobacter jejuni-challenged broiler chickens. Vet. Med. Sci. 2024, 10, e70034. [Google Scholar] [CrossRef] [PubMed]
- Mohamedin, A.; Elsayed, A.; Shakurfow, F.A. Molecular effects and antibacterial activities of ginger extracts against some drug resistant pathogenic bacteria Egypt. J. Bot. 2018, 58, 133–143. [Google Scholar]
- Song, J.M.; Seong, B.L. Tea catechins as a potential alternative anti-infectious agent. Expert Rev. Anti-infect. Ther. 2007, 5, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Akela, M.A.; El-Atrash, A.; El-Kilany, M.I.; Tousson, E. Qualitative and quantitative characterization of biologically active compounds of Rosemary (Rosmarinus officinalis) Leaf Extract. J. ATBAS 2018, 2, 59–64. [Google Scholar]
- Tousson, E.; Masoud, A.; Hafez, E.; Almakhatreh, M. Protective role of rosemary extract against etoposide induced liver toxicity, injury and KI67 alterations in rats. J. Biosci. Appl. Res. 2019, 5, 1–7. [Google Scholar] [CrossRef]
- Alsherbiny, M.A.; Abd-Elsalam, W.H.; El Badawy, S.A.; Taher, E.; Fares, M.; Torres, A.; Chang, D.; Li, C.G. Ameliorative and protective effects of ginger and its main constituents against natural, chemical and radiation-induced toxicities: A comprehensive review. Food Chem. Toxicol. 2019, 123, 72–97. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Ammar, A.M.; Hamdy, M.M.; Gobouri, A.A.; Azab, E.; Sewid, A.H. First report of aacC5-aadA7Δ4 gene cassette array and phage tail tape measure protein on class 1 integrons of Campylobacter species isolated from animal and human sources in Egypt. Animals 2020, 10, 2067. [Google Scholar] [CrossRef]
- Poudel, S.; Li, T.; Chen, S.; Zhang, X.; Cheng, W.; Sukumaran, A.T.; Kiess, A.S.; Zhang, L. Prevalence, antimicrobial resistance, and molecular characterization of Campylobacter isolated from broilers and broiler meat raised without antibiotics. Microbiol. Spectr. 2022, 1, e00251-22. [Google Scholar] [CrossRef]
- Bryan, F.L.; Doyle, M.P. Health risks and consequences of Salmonella and Campylobacter jejuni in raw poultry. J. Food Protec. 1995, 58, 326–344. [Google Scholar] [CrossRef]
- Mohamed, K. Prevalence of Campylobacter spp. and its pathogenic genes in poultry meat, human and environment in Aswan, Upper Egypt. Assiut Vet. Med. J. 2019, 65, 151–158. [Google Scholar]
- Khalifa, N.O.; Afify, J.S.A.; Rabie, N.S. Zoonotic and molecular characterizations of Campylobacter jejuni and Campylobacter coli isolated from beef cattle and children. Glob. Vet. 2013, 11, 585–591. [Google Scholar]
- El-Naenaeey, E.Y.; Abd El-Aziz, N.K.; Sewid, A.H.; Hashem, A.; Hefny, A.A. Antimicrobial resistance, virulence-associated genes, and flagellin typing of thermophilic Campylobacter species isolated from diarrheic humans, raw milk, and broiler niches. Slov. Vet. Res. 2021, 58 (Suppl. S24), 155–164. [Google Scholar] [CrossRef]
- Awadallah, M.; Ahmed, H.; El-Gedawy, A.; Saad, A. Molecular identification of C. jejuni and C. coli in chicken and humans, at Zagazig, Egypt, with reference to the survival of C. jejuni in chicken meat at refrigeration and freezing temperatures. Int. Food Res. J. 2013, 21, 1801–1812. [Google Scholar]
- Chatur, Y.A.; Brahmbhatt, M.N.; Modi, S.; Nayak, J.B. Fluoroquinolone resistance and detection of topoisomerase gene mutation in Campylobacter jejuni isolated from animal and human sources. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 773–783. [Google Scholar]
- Rivera-Mendoza, D.; Martínez-Flores, I.; Santamaría, R.I.; Lozano, L.; Bustamante, V.H.; Pérez-Morales, D. Genomic analysis reveals the genetic determinants associated with antibiotic resistance in the zoonotic pathogen Campylobacter spp. distributed Globally. Front. Microbiol. 2020, 11, 513070. [Google Scholar] [CrossRef]
- Dasti, J.I.; Gross, U.; Pohl, S.; Lugert, R.; Weig, M.; Schmidt-Ott, R. Role of the plasmid-encoded tet(O) gene in tetracycline-resistant clinical isolates of Campylobacter jejuni and Campylobacter coli. J. Med. Microbiol. 2007, 56, 833–837. [Google Scholar] [CrossRef]
- El Baaboua, A.; El Maadoudi, M.; Bouyahya, A.; Kounnoun, A.; Bougtaib, H.; Belmehdi, O. Prevalence and antimicrobial profiling of Campylobacter spp. isolated from meats, animal, and human faeces in Northern of Morocco. Int. J. Food Microbiol. 2021, 349, 109202. [Google Scholar] [CrossRef]
- Debelo, M.; Mohammed, N.; Tiruneh, A.; Tolosa, T. Isolation, identification and antibiotic resistance profile of thermophilic Campylobacter species from bovine, knives and personnel at Jimma Town Abattoir, Ethiopia. PLoS ONE 2022, 17, 10. [Google Scholar] [CrossRef]
- Zhang, T.; Dong, J.; Cheng, Y.; Lu, Q.; Luo, Q.; Wen, G.; Liu, G.; Shao, H. Genotypic diversity, antimicrobial resistance and biofilm-forming abilities of Campylobacter isolated from chicken in Central China. Gut Pathog. 2017, 9, 62. [Google Scholar] [CrossRef]
- Ammar, A.M.; Abd El-Hamid, M.I.; Eid, S.E.A.; El Oksh, A.S. Insights into antimicrobial resistance and virulence genes of emergent multidrug resistant avian pathogenic Escherichia coli in Egypt: How closely related are they? Rev. Med. Vet. 2015, 166, 304–314. [Google Scholar]
- Samad, A.; Abbas, F.; Ahmed, Z.; Akbar, A.; Naeem, M.; Sadiq, M.B.; Ali, I.; Saima; Roomeela; Bugti, F.S.; et al. Prevalence, antimicrobial susceptibility, and virulence of Campylobacter jejuni isolated from chicken meat. J. Food Saf. 2019, 39, 12600. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Abd El-Hamid, M.I.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M.; Abd El-Aziz, N.K.; Abd El-Hamid, M.I.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M. Existence of vancomycin resistance among methicillin resistant S. aureus recovered from animal and human sources in Egypt. Slov. Vet. Res. 2018, 55, 221–230. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Ammar, A.M.; El-Naenaeey, E.Y.M.; El Damaty, H.M.; Elazazy, A.A.; Hefny, A.A.; Shaker, A.; Eldesoukey, I.E. Antimicrobial and antibiofilm potentials of cinnamon oil and silver nanoparticles against Streptococcus agalactiae isolated from bovine mastitis: New avenues for countering resistance. BMC Vet. Res. 2021, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, A.E.; Abd El-Aziz, N.K.; Elariny, E.Y.T.; Shindia, A.; Osman, A.; Hozzein, W.N.; Alkhalifah, D.H.M.; El-Hossary, D. Antibacterial activity of bioactive compounds extracted from red kidney bean (Phaseolus vulgaris L.) seeds against multidrug-resistant Enterobacterales. Front. Microbiol. 2022, 13, 1035586. [Google Scholar] [CrossRef] [PubMed]
- Abdelatti, M.A.I.; Abd El-Aziz, N.K.; El-Naenaeey, E.-s.Y.M.; Ammar, A.M.; Alharbi, N.K.; Alharthi, A.; Zakai, S.A.; Abdelkhalek, A. Antibacterial and anti-efflux activities of cinnamon essential oil against pan and extensive drug-resistant Pseudomonas aeruginosa isolated from human and animal sources. Antibiotics 2023, 12, 1514. [Google Scholar] [CrossRef]
- Taha, S.M.; Abd El-Aziz, N.K.; Abdelkhalek, A.; Pet, I.; Ahmadi, M.; El-Nabtity, S.M. Chitosan-loaded Lagenaria siceraria and Thymus vulgaris potentiate antibacterial, antioxidant, and immunomodulatory activities against extensive drug-resistant Pseudomonas aeruginosa and vancomycin-resistant Staphylococcus aureus: In Vitro and in vivo approaches. Antioxidants 2024, 13, 428. [Google Scholar] [CrossRef]
- Horne, M. The world is running out of antibiotics. Aust. Med. 2017, 29, 31. [Google Scholar]
- Reygaert, W.C. Green tea catechins: Their use in treating and preventing infectious diseases. Biomed. Res. Int. 2018, 2018, 9105261. [Google Scholar] [CrossRef]
- Butucel, E.; Balta, I.; Ahmadi, M.; Dumitrescu, G.; Morariu, F.; Pet, I.; Stef, L.; Corcionivoschi, N. Biocides as biomedicines against foodborne pathogenic bacteria. Biomedicines 2022, 10, 379. [Google Scholar] [CrossRef]
- Maftei, N.-M.; Raileanu, C.R.; Balta, A.A.; Ambrose, L.; Boev, M.; Marin, D.B.; Lisa, E.L. The potential impact of probiotics on human health: An update on their health-promoting properties. Microorganisms 2024, 12, 234. [Google Scholar] [CrossRef]
- Corcionivoschi, N.; Balta, I.; Butucel, E. Natural antimicrobial mixtures disrupt attachment and survival of E. coli and C. jejuni to non-organic and organic surfaces. Foods 2023, 12, 3863. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.; Heredia, N.; Garcia, S. 2(5H)-Furanone, epigallocatechin gallate, and a citricbased disinfectant disturb quorum-sensing activity and reduce motility and biofilm formation of Campylobacter jejuni. Folia Microbiol. 2015, 60, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Henika, P.R.; Mandrell, R.E. Bactericidal activities of plant essential oils and some of their isolated constituents against Campylobacter jejuni, Escherichia coli, Listeria monocytogenes, and Salmonella enterica. J. Food Prot. 2002, 65, 1545–1560. [Google Scholar] [CrossRef] [PubMed]
- Mutlu-Ingok, A.; Catalkaya, G.; Capanoglu, E.; Karbancioglu-Guler, F. Antioxidant and antimicrobial activities of fennel, ginger, oregano and thyme essential oils. Food Front. 2021, 2, 508–518. [Google Scholar] [CrossRef]
- Giriraju, A.; Yunus, G.Y. Assessment of antimicrobial potential of 10% ginger extract against Streptococcus mutans, Candida albicans, and Enterococcus faecalis: An in vitro study. Indian J. Dent. Res. 2013, 24, 397–400. [Google Scholar] [CrossRef]
- Thanissery, R.; Smith, D.P. Marinade with thyme and orange oils reduces Salmonella Enteritidis and Campylobacter coli on inoculated broiler breast fillets and whole wings. Poult. Sci. 2014, 93, 1258–1262. [Google Scholar] [CrossRef]
- Yeddes, W.; Majdi, H.; Gadhoumi, H.; Affes, T.G.; Mohamed, S.N.; Wannes, W.A. Optimizing ethanol extraction of rosemary leaves and their biological evaluations. J. Explor. Res. Pharmacol. 2022, 7, 85–94. [Google Scholar] [CrossRef]
- Vegara, S.; Funes, L.; Martí, N.; Saura, D.; Micol, V.; Valero, M. Bactericidal activities against pathogenic bacteria by selected constituents of plant extracts in carrot broth. Food Chem. 2011, 128, 872–877. [Google Scholar] [CrossRef]
- Tian, Q.; Wei, S.; Su, H.; Zheng, S.; Xu, S.; Liu, M.; Bo, R.; Li, J. Bactericidal activity of gallic acid against multi-drug resistance Escherichia coli. Microb. Pathog. 2022, 173 Pt A, 105824. [Google Scholar] [CrossRef]
- Keyvani-Ghamsari, S.; Rahimi, M.; Khorsand, K. An update on the potential mechanism of gallic acid as an antibacterial and anticancer agent. Food Sci. Nutr. 2023, 11, 5856–5872. [Google Scholar] [CrossRef]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial biofilms in the food industry-a comprehensive review. Int. J. Environ. Res. Public Health 2021, 18, 4. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Yang, D.; Zhao, L.; Shi, F.; Ye, G.; Fu, H.; Lin, J.; Guo, H.; He, R.; Li, J.; et al. EGCG-mediated potential inhibition of biofilm development and quorum sensing in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 4946. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Escaping the biofilm in more than one way: Desorption, detachment or dispersion. Curr. Opin. Microbiol. 2016, 30, 67–78. [Google Scholar] [CrossRef]
- Bae, J.; Oh, E.; Jeon, B. Enhanced transmission of antibiotic resistance in Campylobacter jejuni biofilms by natural transformation. Antimicrob. Agents Chemother. J. 2014, 58, 7573–7575. [Google Scholar] [CrossRef]
- Yuan, R.; Lin, Y. Traditional Chinese medicine: An approach to scientific proof and clinical validation. Pharmacol. Ther. 2000, 86, 191–198. [Google Scholar] [CrossRef]
- Davila, J.C.; Rodriguez, R.J.; Melchert, R.B.; Acosta, D., Jr. Predictive value of in vitro model systems in toxicology. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 63–96. [Google Scholar] [CrossRef]
- Bolton, F.J.; Hutchinson, D.N.; Coates, D. Blood-free, selective medium for isolation of C. jejuni from feces. J. Clin. Microbiol. 1984, 19, 169–171. [Google Scholar] [CrossRef]
- Quinn, P.J.; Carter, M.E.; Markey, B.; Carter, G.R. Clinical Veterinary Microbiology, 1st ed.; Wolf publishing Mosby: London, UK, 1994. [Google Scholar]
- Shin, E.; Lee, Y.H. Comparison of three different methods for Campylobacter isolation from porcine intestines. J. Microbiol. Biotechnol. 2009, 19, 647–650. [Google Scholar]
- Wang, G.; Clark, C.G.; Taylor, T.M.; Pucknell, C.; Barton, C.; Price, L.; Woodward, D.L.; Rodgers, F.G. Colony multiplex PCR assay for identification and differentiation of Campylobacter jejuni, C. coli, C. lari, C. upsaliensis, and C. fetus subsp. fetus. J. Clin. Microbiol. 2002, 40, 4744–4747. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 149–158. [Google Scholar] [CrossRef]
- CLSI. Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Supplement M100, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 7.1, 2017. Available online: https://www.megumed.de/wp-content/uploads/2024/02/v_14.0_Breakpoint_Tables.pdf (accessed on 27 May 2024).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G. Multidrug-resistant, extensively drug-resistant and pan-drug resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2011, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, R.; Dahiya, S.; Sayal, P. Evaluation of multiple antibiotic resistance (MAR) index and doxycycline susceptibility of Acinetobacter species among inpatients. Indian J. Microbiol. Res. 2016, 3, 299. [Google Scholar] [CrossRef]
- Tendolkar, P.M.; Baghdayan, A.S.; Gilmore, M.S.; Shankar, N. Enterococcal surface protein, Esp, enhances biofilm formation by Enterococcus faecalis. Infect. Immun. 2004, 72, 6032–6039. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by Staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Yam, T.S.; Shah, S.; Hamilton-Miller, J.T.M. Microbiological activity of whole and fractionated crude extract of tea (Camellia sinensis) and of tea components. FEMS. Microbiol. Lett. 1997, 152, 169–174. [Google Scholar] [CrossRef]
- Fani, M.M.; Kohanteb, J. Inhibitory activity of Aloe vera gel on some clinically isolated cariogenic and periodontopathic bacteria. J. Oral Sci. 2012, 54, 15–21. [Google Scholar] [CrossRef]
- Choi, O.; Choi, S.K.; Kim, J.; Park, C.G.; Kim, J. In vitro antibacterial activity and major bioactive components of Cinnamomum verum essential oils against cariogenic bacteria, Streptococcus mutans and Streptococcus sobrinus. Asian Pac. J. Trop. Biomed. 2016, 4, 308–314. [Google Scholar] [CrossRef]
- Raja, A.F.; Ali, F.; Khan, I.A.; Shwal, A.S.; Arora, D.S.; Shah, B.A.; Taneja, S.C. Antistaphylococcal and biofilm inhibitory activities of acetyl-11-keto-β-boswellic acid from Boswellia serrata. BMC Microbiol. 2011, 11, 54–62. [Google Scholar] [CrossRef]
- He, Q.; Yao, K.; Jia, D.; Fan, H.; Liao, X.; Shi, B. Determination of total catechins in tea extracts by HPLC and spectrophotometry. Nat. Prod. Res. 2009, 23, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Olech, M.; Nowak, R.; Los, R.; Rzymowska, J.; Malm, A.; Chruściel, K. Biological activity and composition of teas and tinctures prepared from Rosa rugosa Thunb. Cent. Eur. J. Biol. 2012, 7, 172–182. [Google Scholar] [CrossRef]
- Zheng, J.; Meng, J.; Zhao, S.; Singh, R.; Song, W. Adherence to and invasion of human intestinal epithelial cells by Campylobacter jejuni and Campylobacter coli isolates from retail meat products. J. Food Prot. 2006, 69, 768–774. [Google Scholar] [CrossRef]
- Wagle, B.R.; Upadhyay, A.; Upadhyaya, I.; Shrestha, S.; Arsi, K.; Liyanage, R.; Venkitanarayanan, K.; Donoghue, D.J.; Donoghue, A.M. Trans-cinnamaldehyde, eugenol and carvacrol reduce Campylobacter jejuni biofilms and modulate expression of select genes and proteins. Front. Microbiol. 2019, 10, 1837. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source (No.) | Sample Type (No.) | Overall Occurrence of Campylobacter Isolates No. (%) | Campylobacter Species No. (%) | p-Value | |
|---|---|---|---|---|---|
| C. jejuni | C. coli | ||||
| Chickens and their products (290) | Cloacal swabs (50) | 10 (20.00) | 1 (2.00) | 9 (18.00) | 0.0003 |
| Breast muscles (50) | 5 (10.00) | 4 (8.00) | 1 (2.00) | 0.0507 | |
| Thigh muscles (50) | 9 (18.00) | 6 (12.00) | 3 (6.00) | 0.1572 | |
| Liver (20) | 12 (60.00) | 11 (55.00) | 1 (5.00) | 0.0001 | |
| Gizzard (20) | 7 (35.00) | 5 (25.00) | 2 (10.00) | 0.1088 | |
| Neck skin (50) | 8 (16.00) | 6 (12.00) | 2 (4.00) | 0.0455 | |
| Cecal parts (50) | 15 (30.00) | 13 (26.00) | 2 (4.33) | 0.0001 | |
| Total positives | 66 (22.76) | 45 (15.52) | 21 (7.24) | 0.0001 | |
| Environmental samples (60) | Litter (20) | 3 (15.00) | 3 (15.00) | 0 (0.00) | 0.0326 |
| Feed (20) | 1 (5.00) | 1 (5.00) | 0 (0.00) | 0.0862 | |
| Water (20) | 1 (5.00) | 1 (5.00) | 0 (0.00) | 0.0862 | |
| Total positives | 5 (8.33) | 5 (8.33) | 0 (0.00) | 0.0862 | |
| Human (50) | Stool (50) | 15 (30.00) | 10 (20.00) | 5 (10.00) | 0.0678 |
| Total | 400 | 86 (21.5) | 61 (15.25) | 25 (6.25) | 0.0001 |
| AMA | C. jejuni Isolates (n = 61) | Total | p-Value | C. coli Isolates (n = 25) | Total | p-Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chicken Source (n = 46) | Environmental Source (n = 5) | Human Stool (n = 10) | Chicken Source (n = 20) | Human Stool (n = 5) | |||||||||||||
| Muscle (10) | Internal Organs (29) | Neck Skin (6) | Cloacal Swab (1) | Litter (3) | Feed (1) | Water (1) | Muscle (4) | Internal Organ (5) | Nick Skin (2) | Cloacal Swab (9) | |||||||
| P | 9 (90.00) | 25 (86.21) | 6 (100.00) | 1 (100.00) | 2 (66.66) | 0 (0.00) | 1 (100.00) | 8 (80.00) | 52 (85.25) | 0.0001 | 2 (50.00) | 5 (100.00) | 2 (100.00) | 7 (77.78) | 4 (80.00) | 20 (80.00) | 0.0001 |
| SAM | 4 (40.00) | 5 (17.24) | 3 (50.00) | 1 (100.00) | 1 (33.33) | 0 (0.00) | 0 (0.00) | 2 (20.00) | 16 (26.23) | 0.0001 | 2 (50.00) | 2 (40.00) | 1 (50.00) | 0 (0.00) | 2 (40.00) | 7 (28.00) | 0.0001 |
| AMC | 2 (20.00) | 6 (20.69) | 2 (33.33) | 1 (100.00) | 1 (33.33) | 1 (100.00) | 0 (0.00) | 5 (50.00) | 18 (29.51) | 0.0001 | 1 (25.00) | 2 (40.00) | 0 (0.00) | 1 (11.11) | 4 (80.00) | 8 (32.00) | 0.0001 |
| E | 7 (70.00) | 24 (82.76) | 6 (100.00) | 1 (100.00) | 2 (66.66) | 1 (100.00) | 1 (100.00) | 7 (70.00) | 49 (80.33) | 0.5416 | 2 (50.00) | 5 (100.00) | 2 (100.00) | 9 (100.00) | 5 (100.00) | 23 (92.00) | 0.5153 |
| TL | 3 (30.00) | 4 (13.79) | 2 (33.33) | 1 (100.00) | 1 (33.33) | 1 (100.00) | 0 (0.00) | 4 (40.00) | 16 (26.23) | 0.0001 | 1 (25.00) | 1 (20.00) | 1 (50.00) | 0 (0.00) | 2 (40.00) | 5 (20.00) | 0.0001 |
| CT | 7 (70.00) | 26 (89.66) | 4 (66.67) | 1 (100.00) | 3 (100.00) | 1 (100.00) | 1 (100.00) | 9 (90.00) | 52 (85.25) | 0.6129 | 4 (100.00) | 4 (80.00) | 2 (100.00) | 9 (100.00) | 3 (60.00) | 22 (88.00) | 0.7641 |
| C | 4 (40.00) | 5 (17.24) | 2 (33.33) | 1 (100.00) | 1 (33.33) | 1 (100.00) | 1 (100.00) | 2 (20.00) | 17 (27.87) | 0.0001 | 0 (0.00) | 2 (40.00) | 1 (50.00) | 1 (11.11) | 3 (60.00) | 7 (28.00) | 0.0001 |
| CTX | 8 (80.00) | 25 (86.21) | 6 (100.00) | 1 (100.00) | 3 (100.00) | 0 (0.00) | 1 (100.00) | 7 (70.00) | 51 (83.61) | 0.0001 | 4 (100.00) | 5 (100.00) | 2 (100.00) | 9 (100.00) | 5 (100.00) | 25 (100.00) | 1.000 |
| TE | 9 (90.00) | 24 (82.76) | 4 (66.67) | 1 (100.00) | 2 (66.66) | 1 (100.00) | 1 (100.00) | 8 (80.00) | 50 (81.97) | 0.4534 | 4 (100.00) | 5 (100.00) | 2 (100.00) | 6 (66.67) | 4 (80.00) | 21 (84.00) | 0.5686 |
| CN | 3 (30.00) | 4 (13.79) | 2 (33.33) | 0 (0.00) | 1 (33.33) | 1 (100.00) | 0 (0.00) | 3 (30.00) | 14 (22.95) | 0.0001 | 1 (25.00) | 1 (20.00) | 1 (50.00) | 0 (0.00) | 2 (40.00) | 5 (20.00) | 0.0001 |
| CIP | 4 (40.00) | 4 (13.79) | 2 (33.33) | 1 (100.00) | 1 (33.33) | 1 (100.00) | 0 (0.00) | 4 (40.00) | 17 (27.87) | 0.0001 | 0 (0.00) | 2 (40.00) | 1 (50.00) | 0 (0.00) | 2 (40.00) | 5 (20.00) | 0.0001 |
| NOR | 4 (40.00) | 6 (20.69) | 1 (16.61) | 1 (100.00) | 1 (33.33) | 0 (0.00) | 1 (100.00) | 3 (30.00) | 17 (27.87) | 0.0001 | 1 (25.00) | 2 (40.00) | 0 (0.00) | 0 (0.00) | 0 (0.00) | 3 (12.00) | 0.0001 |
| SXT | 3 (30.00) | 6 (20.69) | 2 (33.33) | 1 (100.00) | 0 (0.00) | 0 (0.00) | 1 (100.00) | 4 (40.00) | 17 (27.87) | 0.0001 | 1 (25.00) | 2 (40.00) | 1 (50.00) | 0 (0.00) | 2 (40.00) | 6 (24.00) | 0.0001 |
| Isolate No. | Source | Campylobacter Species | Antimicrobial Resistance Pattern | MAR Index | Biofilm Formation | |
|---|---|---|---|---|---|---|
| OD 570 * | Degree | |||||
| 1 | Human stool | C. jejuni | P, SAM, AMC, E, CIP, NOR, SXT, CN, TE, C, CT, CTX, TL | 1.00 | 0.52 | Strong |
| 2 | Chicken gizzard | C. jejuni | P, SAM, AMC, E, CIP, NOR, SXT, CN, TE, C, CT, CTX, TL | 1.00 | 0.55 | Strong |
| 3 | Chicken neck skin | C. jejuni | P, SAM, AMC, E, CIP, NOR, SXT, CN, TE, C, CT, CTX, TL | 1.00 | 0.63 | Strong |
| 4 | Chicken liver | C. jejuni | P, SAM, AMC, E, CIP, NOR, SXT, CN, C, CT, CTX, TL | 0.92 | 0.13 | Weak |
| 5 | Human stool | C. jejuni | P, SAM, E, CIP, NOR, SXT, CN, TE, CT, CTX, TL | 0.85 | 0.6 | Strong |
| 6 | Chicken litter | C. jejuni | P, SAM, E, CIP, NOR, CN, TE, C, CT, CTX, TL | 0.85 | 0.55 | Strong |
| 7 | Human stool | C. coli | P, SAM, AMC, CIP, NOR, SXT, CN, TE, CT, CTX, TL | 0.85 | 0.48 | Strong |
| 8 | Chicken liver | C. jejuni | P, SAM, CIP, NOR, SXT, CN, TE, C, CT, CTX, TL | 0.85 | 0.49 | Strong |
| 9 | Chicken breast muscle | C. coli | P, SAM, AMC, E, NOR, SXT, CN, TE, CT, CTX, TL | 0.85 | 0.29 | Moderate |
| 10 | Chicken neck skin | C. jejuni | P, SAM, AMC, E, CIP, SXT, CN, C, CT, CTX, TE | 0.85 | 0.50 | Strong |
| 11 | Chicken neck skin | C. coli | P, SAM, E, CIP, SXT, CN, TE, C, CT, CTX, TL | 0.85 | 0.21 | Moderate |
| 12 | Chicken thigh muscle | C. jejuni | P, SAM, E, CIP, NOR, SXT, TE, C, CT, CTX, TL | 0.85 | 0.59 | Strong |
| 13 | Chicken thigh muscle | C. jejuni | P, SAM, E, CIP, NOR, CN, TE, C, CT, CTX, TL | 0.85 | 0.20 | Moderate |
| 14 | Chicken cecal swab | C. jejuni | P, SAM, AMC, E, CIP, NOR, TE, C, CT, CTX, TL | 0.85 | 0.49 | Strong |
| 15 | Chicken cecal part | C. jejuni | P, SAM, AMC, E, CIP, NOR, SXT, CN, TE, CT, TL | 0.85 | 0.24 | Moderate |
| 16 | Chicken cecal part | C. coli | P, SAM, AMC, E, CIP, NOR, SXT, TE, C, CTX, TL | 0.85 | 0.44 | Strong |
| 17 | Chicken breast muscle | C. jejuni | P, E, CIP, NOR, SXT, CN, TE, C, CTX, TL | 0.77 | 0.20 | Moderate |
| 18 | Chicken breast muscle | C. jejuni | P, SAM, AMC, E, CIP, SXT, TE, C, CT, CTX | 0.77 | 0.60 | Strong |
| 19 | Feed | C. jejuni | P, E, CIP, CN, TE, C, CT, TL | 0.62 | 0.61 | Strong |
| 20 | Water | C. jejuni | P, NOR, SXT, TE, C, CT, CTX | 0.54 | 0.62 | Strong |
| Isolate No. | Campylobacter Species | Plant Extract | p-Value | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Green Tea | Rosemary | Ginger | |||||||||||||||||||||
| Disc Diffusion (mm) | MIC (mg/mL) | Disc Diffusion (mm) | MIC (mg/mL) | Disc Diffusion (mm) | MIC (mg/mL) | ||||||||||||||||||
| 50 | 25 | 12.5 | 6.25 | 3.12 | 1.56 | 50 | 25 | 12.5 | 6.25 | 3.12 | 1.56 | 50 | 25 | 12.5 | 6.25 | 3.12 | 1.56 | ||||||
| 1 | C. jejuni | 37 | 32 | 22 | 19 | 10 | 5 | 3.12 a | 15 | 11 | 5 | 2 | 0 | 0 | 25 a | 20 | 16 | 12 | 6 | 4 | 0 | 12.5 b | 0.0281 |
| 2 | C. jejuni | 36 | 31 | 25 | 20 | 11 | 8 | 1.56 a | 16 | 11 | 5 | 3 | 0 | 0 | 25 a | 19 | 13 | 10 | 5 | 0 | 0 | 12.5 b | 0.0108 |
| 3 | C. jejuni | 37 | 30 | 25 | 19 | 10 | 5 | 3.12 a | 14 | 7 | 5 | 0 | 0 | 0 | 50 a | 20 | 15 | 10 | 7 | 3 | 0 | 12.5 b | 0.0148 |
| 4 | C. jejuni | 37 | 30 | 24 | 20 | 10 | 5 | 3.12 a | 15 | 10 | 3 | 0 | 0 | 0 | 25 a | 14 | 11 | 8 | 4 | 0 | 0 | 12.5 b | 0.0084 |
| 5 | C. jejuni | 38 | 33 | 25 | 20 | 10 | 5 | 3.12 a | 15 | 10 | 4 | 2 | 0 | 0 | 25 a | 19 | 12 | 7 | 3 | 0 | 0 | 25 a | 0.0130 |
| 6 | C. jejuni | 37 | 30 | 23 | 18 | 8 | 4 | 3.12 a | 15 | 11 | 6 | 3 | 0 | 0 | 25 a | 20 | 17 | 9 | 5 | 0 | 0 | 12.5 b | 0.0469 |
| 7 | C. coli | 38 | 30 | 25 | 20 | 9 | 4 | 3.12 a | 12 | 9 | 5 | 0 | 0 | 0 | 25 a | 18. | 15 | 11 | 7 | 4 | 0 | 12.5 b | 0.0155 |
| 8 | C. jejuni | 35 | 30 | 25 | 20 | 9 | 3 | 3.12 a | 15 | 11 | 4 | 1 | 0 | 0 | 25 a | 15 | 10 | 7 | 4 | 0 | 0 | 25 a | 0.0139 |
| 9 | C. coli | 26 | 29 | 24 | 19 | 11 | 8 | 1.56 a | 18 | 10 | 4 | 0 | 0 | 0 | 25 a | 19 | 15 | 10 | 6 | 3 | 0 | 12.5 b | 0.0161 |
| 10 | C. jejuni | 36 | 31 | 24 | 20 | 12 | 8 | 1.56 a | 15 | 11 | 4 | 0 | 0 | 0 | 25 a | 17 | 12 | 6 | 4 | 0 | 0 | 25 a | 0.0052 |
| 11 | C. coli | 36 | 31 | 23 | 16 | 9 | 4 | 3.12 a | 16 | 10 | 6 | 3 | 0 | 0 | 25 a | 19. | 15 | 9 | 6 | 0 | 0 | 12.5 b | 0.0416 |
| 12 | C. jejuni | 37 | 30 | 22 | 17 | 8 | 4 | 3.12 a | 16 | 10 | 3 | 0 | 0 | 0 | 25 a | 19 | 13 | 6 | 3 | 0 | 0 | 25 a | 0.0311 |
| 13 | C. jejuni | 38 | 32 | 23 | 18 | 11 | 8 | 1.56 a | 16 | 11 | 5 | 2 | 0 | 0 | 25 a | 19 | 16 | 10 | 7 | 4 | 0 | 12.5 b | 0.0167 |
| 14 | C. jejuni | 37 | 32 | 23 | 18 | 9 | 4 | 3.12 a | 15 | 10 | 4 | 0 | 0 | 0 | 25 a | 18 | 13 | 9 | 5 | 0 | 0 | 12.5 b | 0.0227 |
| 15 | C. jejuni | 35 | 30 | 23 | 19 | 8 | 3 | 3.12 a | 15 | 10 | 4 | 0 | 0 | 0 | 25 a | 18 | 14 | 9 | 5 | 0 | 0 | 12.5 b | 0.0295 |
| 16 | C. coli | 37 | 30 | 23 | 19 | 9 | 5 | 3.12 a | 16 | 10 | 5 | 0 | 0 | 0 | 25 a | 19 | 15 | 10 | 5 | 0 | 0 | 12.5 b | 0.0267 |
| 17 | C. jejuni | 35 | 32 | 23 | 18 | 10. | 4 | 3.12 a | 16 | 10 | 3 | 0 | 0 | 0 | 25 a | 19 | 15 | 10 | 6 | 2 | 0 | 12.5 b | 0.0254 |
| 18 | C. jejuni | 37 | 32 | 24 | 19 | 9. | 3 | 3.12 a | 15 | 11 | 5 | 3 | 0 | 0 | 25 a | 19 | 16 | 9 | 5 | 0 | 0 | 12.5 b | 0.0345 |
| 19 | C. jejuni | 35 | 32 | 22 | 18 | 9 | 4 | 3.12 a | 15 | 10 | 3 | 0 | 0 | 0 | 25 a | 20 | 15 | 10 | 6 | 3 | 0 | 12.5 b | 0.0289 |
| 20 | C. jejuni | 37 | 29 | 24 | 21 | 9 | 5 | 3.12 a | 16 | 11 | 5 | 2 | 0 | 0 | 25 a | 19 | 15 | 11 | 6 | 3 | 0 | 12.5 b | 0.0255 |
| Overall average | 36.0 ± 0.57 | 30.8 ± 0.25 | 23.6 ± 0.23 | 18.9 ± 0.27 | 9.55 ± 0.24 | 4.95 ± 0.38 | p = 0.0175 b-reg = 0.576 | 15.3 ± 0.25 | 10.2 ± 0.21 | 4.4 ± 0.21 | 1.05 ± 0.28 | 0 ± 0.00 | 0 ± 0.00 | p = 0.0005 b-reg = 0.334 | 18.5 ± 1.25 | 14.1 ± 0.35 | 9.15 ± 0.41 | 5.25 ± 0.36 | 1.3 ± 0.27 | 0 ± 0.00 | p = 0.0048 b-reg = 0.370 | ||
| Constituents | Area | Concentration (µg/mL) |
|---|---|---|
| Gallic acid | 641.83 | 424.69 |
| Chlorogenic acid | 47.59 | 54.64 |
| Catechin | 77.77 | 150.77 |
| Methyl gallate | 0.00 | 0.00 |
| Coffeic acid | 78.38 | 52.68 |
| Syringic acid | 25.79 | 14.67 |
| Rutin | 129.69 | 176.50 |
| Ellagic acid | 82.75 | 56.96 |
| Coumaric acid | 0.00 | 0.00 |
| Vanillin | 0.00 | 0.00 |
| Ferulic acid | 0.00 | 0.00 |
| Naringenin | 0.00 | 0.00 |
| Rosmarinic acid | 0.00 | 0.00 |
| Daidzein | 0.00 | 0.00 |
| Querectin | 0.00 | 0.00 |
| Cinnamic acid | 40.74 | 6.24 |
| Kaempferol | 0.00 | 0.00 |
| Hesperetin | 0.00 | 0.00 |
| Component Name | Expected RT | Area * | RT | Conc. (µg/mL) |
|---|---|---|---|---|
| Gallic acid 168.9/124.9 | 1.67 | 8.34 × 107 | 1.69 | 86.51 |
| Caffeic acid 178/135 | 5.83 | 1.57× 106 | 5.88 | 0.5761 |
| Rutin 609/299.9 | 9.13 | 5.36 × 10 7 | 9.15 | 19.42 |
| Coumaric acid 162.9/119 | 7.7 | 1.97 × 10 7 | 7.73 | 1.747 |
| Naringenin 271/151 | 20.98 | 6.17 × 10 5 | 20.94 | 47.01 |
| Querectin 301/151 | 18.16 | 2.24 × 10 6 | 18.14 | 0.4713 |
| Ellagic acid 301/145 | 8.97 | 2.30 × 10 5 | 9.01 | 0.7675 |
| 3.4-Dihydroxybenzoic acid 152.9/109 | 3.13 | 5.98 × 10 6 | 3.19 | 5.9 |
| Hesperetin 301/136 | 22.62 | N/A | N/A | ND |
| Cinnamic acid 146.9/77 | 18.16 | N/A | N/A | ND |
| Methyl gallate 183/124 | 5.04 | 1.57 × 10 6 | 5.11 | ND |
| Kaempferol 284.7/93 | 22.08 | 5.46 × 10 5 | 22.06 | 0.1809 |
| Ferulic acid 192.8/133.9 | 8.89 | 1.84 × 10 5 | 8.94 | 0.324 |
| Syringic acid 196.8/181.9 | 6.23 | 6.13 × 10 4 | 6.3 | 0.3735 |
| Apigenin 269/151 | 21.47 | N/A | N/A | ND |
| Catechin 288.8/109 | 5.05 | 1.68 × 10 6 | 5.11 | 6.688 |
| Daidzein 253/132 | 16.22 | N/A | N/A | ND |
| Chlorogenic acid 353/191 | 5.1 | 6.63 × 10 6 | 5.16 | 1.628 |
| Resveratrol 227/185 | 14.27 | 4.00 × 10 4 | 14.36 | 0.1487 |
| Rosmarinic acid 359.1/161 | 13.42 | N/A | N/A | ND |
| Isolate No. | Campylobacter Species | OD Pre-Treatment | OD Post Treatment (Reduction %) * | p-Value |
|---|---|---|---|---|
| 1 | C.jejuni | 0.52 | 0.14 (73.1) | 0.0001 |
| 2 | C.jejuni | 0.55 | 0.25 (54.6) | 0.0001 |
| 3 | C.jejuni | 0.63 | 0.21 (66.7) | 0.0001 |
| 5 | C.jejuni | 0.6 | 0.21 (65.0) | 0.0001 |
| 6 | C.jejuni | 0.55 | 0.14 (74.6) | 0.0001 |
| 7 | C. coli | 0.48 | 0.10 (79.2) | 0.0001 |
| 8 | C.jejuni | 0.49 | 0.13 (73.5) | 0.0001 |
| 10 | C.jejuni | 0.50 | 0.15 (70.0) | 0.0001 |
| 12 | C.jejuni | 0.59 | 0.13 (79.0) | 0.0001 |
| 14 | C.jejuni | 0.49 | 0.12 (75.5) | 0.0001 |
| 16 | C. coli | 0.44 | 0.22 (50.0) | 0.0001 |
| 18 | C.jejuni | 0.60 | 0.22 (63.3) | 0.0001 |
| 19 | C.jejuni | 0.61 | 0.14 (77.1) | 0.0001 |
| 20 | C.jejuni | 0.62 | 0.24 (61.29) | 0.0001 |
| Overall average | 0.547 ± 0.01 a | 0.171 ± 0.01 b | 0.0001 | |
| ID | Q1 (m/z) | Q3 (m/z) | RT (min) | CE (V) | CXP (V) | DP (V) |
|---|---|---|---|---|---|---|
| Gallic acid 168.9/124.9 | 168.9 | 124.9 | 1.67 | −30 | −11 | −110 |
| Gallic acid 168.9/79 | 168.9 | 79 | 1.67 | −30 | −11 | −110 |
| Caffeic acid 178/135 | 178.9 | 135 | 5.83 | −22 | −9 | −115 |
| Caffeic acid 178/107 | 178.9 | 107 | 5.83 | −30 | −7 | −115 |
| Rutin 609/299.9 | 609 | 299.9 | 9.13 | −48 | −15 | −230 |
| Rutin 609/270.9 | 609 | 270.9 | 9.13 | −70 | −9 | −230 |
| Coumaric acid 162.9/119 | 162.9 | 119 | 7.7 | −20 | −7 | −90 |
| Coumaric acid 162.9/93 | 162.9 | 93 | 7.7 | −40 | −5 | −90 |
| Vanillin 151/136 | 151 | 136 | 7.48 | −12 | −9 | −140 |
| Vanillin 151/92 | 151 | 92 | 7.48 | −16 | −7 | −140 |
| Naringenin 271/151 | 271 | 151 | 20.98 | −24 | −25 | −130 |
| Naringenin 271/119 | 271 | 119 | 20.98 | −34 | −11 | −130 |
| Querectin 301/151 | 301 | 151 | 18.16 | −28 | −9 | −50 |
| Querectin 301/178.8 | 301 | 178.8 | 18.16 | −20 | −7 | −50 |
| Ellagic acid 301/145 | 301 | 145 | 8.97 | −40 | −14 | −120 |
| Ellagic acid 301/245 | 301 | 245 | 8.97 | −38 | −14 | −120 |
| 3.4-Dihydroxybenzoic acid 152.9/109 | 152.9 | 109 | 3.13 | −40 | −5 | −75 |
| 3,4-Dihydroxybenzoic acid 152.9/90.9 | 152.9 | 90.9 | 3.13 | −20 | −7 | −75 |
| Hesperetin 301/164 | 301 | 164 | 22.62 | −23 | −10 | −125 |
| Hesperetin 301/136 | 301 | 136 | 22.62 | −38 | −10 | −125 |
| Cinnamic acid 146.9/102.6 | 146.9 | 102.6 | 18.16 | −17 | −6 | −60 |
| Cinnamic acid 146.9/77 | 146.9 | 77 | 18.16 | −33 | −6 | −60 |
| Methyl gallate 183/124 | 183 | 124 | 5.04 | −30 | −10 | −110 |
| Methyl gallate 183/140 | 183 | 140 | 5.04 | −30 | −10 | −110 |
| Kaempferol 284.7/93 | 284.7 | 93 | 22.08 | −46 | −10 | −120 |
| Kaempferol 284.7/116.8 | 284.7 | 116.8 | 22.08 | −52 | −10 | −120 |
| Ferulic acid 192.8/133.9 | 192.8 | 133.9 | 8.89 | −16 | −5 | −25 |
| Ferulic acid 192.8/177.9 | 192.8 | 177.9 | 8.89 | −12 | −5 | −25 |
| Syringic acid 196.9/122.8 | 196.9 | 122.8 | 6.23 | −24 | −5 | −30 |
| Syringic acid 196.8/181.9 | 196.9 | 181.9 | 6.23 | −12 | −5 | −30 |
| Apigenin 269/151 | 269 | 151 | 21.47 | −15 | −7 | −35 |
| Apigenin 269/117 | 269 | 117 | 21.47 | −15 | −7 | −35 |
| Catechin 288.8/244.9 | 288.8 | 244.9 | 5.05 | −16 | −8 | −40 |
| Catechin 288.8/109 | 288.8 | 109 | 5.05 | −32 | −8 | −40 |
| Luteolin 284.7/132.9 | 284.7 | 132.9 | 18.14 | −38 | −10 | −50 |
| Luteolin 284.7/150.9 | 284.7 | 150.9 | 18.14 | −26 | −10 | −50 |
| Daidzein 253/132 | 253 | 132 | 16.22 | −55 | −10 | −65 |
| Daidzein 253/91 | 253 | 91 | 16.22 | −50 | −13 | −65 |
| Chlorogenic acid 353/191 | 353 | 191 | 5.1 | −23 | −10 | −60 |
| Chlorogenic acid 353/179 | 353 | 179 | 5.1 | −35 | −10 | −60 |
| Resveratrol227/185 | 277 | 187 | 14.27 | −53 | −26 | −10 |
| Resvertrol 277/143 | 277 | 143 | 14.27 | −53 | −40 | −10 |
| Romarinic acid 359.1/161 | 359.1 | 161.030 | 13.42 | −60 | −20 | −10 |
| Rosmarinic acid 359.1/197 | 359.1 | 197.050 | 13.42 | −60 | −20 | −10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emara, M.S.; Ammar, A.M.; Abdelwahab, A.M.O.; Elgdawy, A.A.; Abdelkhalek, A.; Pet, E.; Dumitrescu, G.; Ahmadi, M.; Abd El-Aziz, N.K. Green Tea Extract (Theaceae; Camellia sinensis L.): A Promising Antimicrobial, Anti-Quorum Sensing and Antibiofilm Candidate Against Multidrug-Resistant Campylobacter Species. Antibiotics 2025, 14, 61. https://doi.org/10.3390/antibiotics14010061
Emara MS, Ammar AM, Abdelwahab AMO, Elgdawy AA, Abdelkhalek A, Pet E, Dumitrescu G, Ahmadi M, Abd El-Aziz NK. Green Tea Extract (Theaceae; Camellia sinensis L.): A Promising Antimicrobial, Anti-Quorum Sensing and Antibiofilm Candidate Against Multidrug-Resistant Campylobacter Species. Antibiotics. 2025; 14(1):61. https://doi.org/10.3390/antibiotics14010061
Chicago/Turabian StyleEmara, Mona S., Ahmed M. Ammar, Ashraf M.O. Abdelwahab, Attia A. Elgdawy, Adel Abdelkhalek, Elena Pet, Gabi Dumitrescu, Mirela Ahmadi, and Norhan K. Abd El-Aziz. 2025. "Green Tea Extract (Theaceae; Camellia sinensis L.): A Promising Antimicrobial, Anti-Quorum Sensing and Antibiofilm Candidate Against Multidrug-Resistant Campylobacter Species" Antibiotics 14, no. 1: 61. https://doi.org/10.3390/antibiotics14010061
APA StyleEmara, M. S., Ammar, A. M., Abdelwahab, A. M. O., Elgdawy, A. A., Abdelkhalek, A., Pet, E., Dumitrescu, G., Ahmadi, M., & Abd El-Aziz, N. K. (2025). Green Tea Extract (Theaceae; Camellia sinensis L.): A Promising Antimicrobial, Anti-Quorum Sensing and Antibiofilm Candidate Against Multidrug-Resistant Campylobacter Species. Antibiotics, 14(1), 61. https://doi.org/10.3390/antibiotics14010061