Abstract
The emergence of multi-drug resistant pathogens is a major public health problem, leading us to rethink and innovate our bacterial control strategies. Here, we explore the antibiofilm and antivirulence activities of nineteen 6-polyaminosterol derivatives (squalamine-based), presenting a modulation of their polyamine side chain on four major pathogens, i.e., carbapenem-resistant A. baumannii (CRAB) and P. aeruginosa (CRPA), methicillin-resistant S. aureus (MRSA), and vancomycin-resistant E. faecium (VRE) strains. We screened the effect of these derivatives on biofilm formation and eradication. Derivatives 4e (for CRAB, VRE, and MRSA) and 4f (for all the strains) were the most potent ones and displayed activities as good as those of conventional antibiotics. We also identified 11 compounds able to decrease by more than 40% the production of pyocyanin, a major virulence factor of P. aeruginosa. We demonstrated that 4f treatment acts against bacterial infections in Galleria mellonella and significantly prolonged larvae survival (from 50% to 80%) after 24 h of CRAB, VRE, and MRSA infections. As shown by proteomic studies, 4f triggered distinct cellular responses depending on the bacterial species but essentially linked to cell envelope. Its interesting antibiofilm and antivirulence properties make it a promising a candidate for use in therapeutics.
1. Introduction
Gram-positive and Gram-negative pathogens exist as planktonic cells only at limited times during their life cycle. In response to environmental signals, pathogenic bacteria can adopt varied cellular fates, which involve the activation of virulence gene programs and/or the induction of a sessile lifestyle to form multicellular surface-attached communities called biofilms.
Acinetobacter baumannii and Pseudomonas aeruginosa are representatives of Gram-negative bacterial pathogens, both recognized as very problematic pathogens due to the spectacular increase of infections caused by multidrug-resistant (MDR) strains, in parti-cular, carbapenem-resistant strains [1]. The virulence of A. baumannii is a multifactorial process that does not rely on toxin production but rather on a “persist and resist” virulence strategy [2], where biofilm formation plays an important role. Several adhesins such as OmpA, Ata, Bap-like proteins, FhaC, tip pilin of fimbriae, the poly-N-acetylglucosamine polymer and capsular polysaccharides, and outer membrane (OM) vesicles, which all participate in biofilm formation, have been recognized as virulence factors [3]. Biofilms also participate in A. baumannii resistance against desiccation, disinfectants, or host immune systems. Reducing the formation of biofilms is one of the main ways of combating this pathogen. For its part, P. aeruginosa displays a large and complex arsenal of cell-associated (pili, secretion systems…) and extracellular virulence factors (toxins, enzymes…), which play an important role in colonization, bacterial survival, and tissue invasion. During acute infections, P. aeruginosa secretes virulence factors, including the phenazine pyocyanin that contributes to tissue damage and neutrophil defense inactivation [4,5]. Cell-associated factors are involved in biofilm formation, leading to chronic infections to avoid host defenses and compete with host microbiota [6].
Staphylococcus aureus and Enterococcus faecium are representatives of Gram-positive bacteria pathogens. S. aureus is a prominent human pathogen that causes systemic diseases, i.e., endocarditis, osteomyelitis, and bacteremia. These infections are often provoked by biofilms encountered on therapeutic devices. The pathogenicity of S. aureus mainly depends on the expression of multiple virulence factors, like hemolytic toxins, coagulase, and adhesion factors [7]. S. aureus shows an important ability to form biofilm on the surface of prosthetic devices. Thus, given the dissemination of methicillin-resistant phenotype (MRSA), the treatment of biofilm-associated infections is increasingly challenging. Unlike staphylococci, E. faecium does not produce pro-inflammatory toxins but synthetizes adhesion proteins that mediate adherence to host tissues [8] as well as biofilm formation [9]. Enterococci also have exceptional abilities to survive host or environmental stresses, as well as antimicrobial agents (like cephalosporin or aminoglycosides). They have the important ability to acquire and disseminate antibiotic-resistance determinants (like vancomycin-resistance genes for VRE strains), a malleable genome that can accumulate mutations, and exogenous genes that confer additional resistance [10]. E. faecium is intrinsically resistant to several antibiotics like ampicillin and aminoglycosides. Vancomycin is the usual alternative treatment for infections caused by ampicillin-resistant enterococci, but the emergence of high-level resistance has led to many therapeutic deadlocks [11].
For all these pathogens, the discovery of an effective antivirulence strategy could offer an interesting alternative to antibiotic treatments, whose effectiveness is diminishing.
In 1993, squalamine 1 and trodusquemine 2 (Figure 1) were identified as the first na-tural aminosterols from the dogfish shark, Squalus acanthias, to exhibit potent antimicrobial activities against both Gram-positive and Gram-negative MDR bacterial species [12,13].
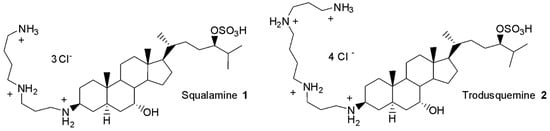
Figure 1.
Structure of squalamine 1 and trodusquemine 2.
Since then, polyaminosterol derivatives have been designed and evaluated for their antimicrobial activities, particularly due to their original mechanism of action [13,14]. Indeed, it was reported that in planktonic growth, the squalamine and claramine A1 derivatives have a bactericidal effect on Gram-positive bacteria via fast membrane depolarization and disruption [15]. On Gram-negative bacteria, a membrane permeabilization effect (similar to the one of antibacterial polymyxin [16]) associated with a disruption of the proton gradient makes these water-soluble compounds very promising therapeutics [15].
In a previous study, we easily prepared 6-polyaminosteroid compounds 4a–4r (Figure 2) derived from squalamine that presented interesting antibacterial activities against both Gram-positive and Gram-negative MDR strains and were able to potentialize carbapenems, vancomycin, or oxacillin [17].
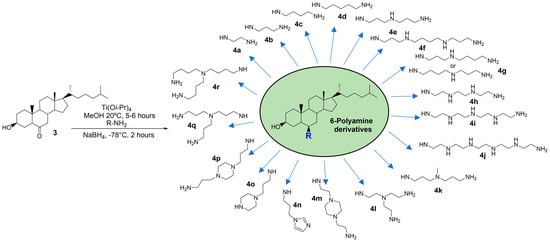
Figure 2.
Scheme presenting the easily prepared 6-polyaminosteroid derivatives tested in this study. They are numbered from 4a to 4r according to the modification of their polyamine “R” side chain.
Here, to explore new routes to combat resistant bacteria, we examined the antibiofilm and antivirulence properties of these derivatives. Since biofilm is a common trait of the four pathogens tested, we first assayed their effect on biofilm formation and eradication for each strain. Then, for P. aeruginosa, which is known to produce numerous secreted virulence factors involved in acute infections, the derivatives were also tested for their effect on pyocyanin production. The capacity to lower the bacterial virulence in vivo using a Galleria mellonella infection model was observed in the most efficient antibacterial 6-polyaminosteroid 4f derivative. Finally, to understand the mode of action of this derivative, we carried out proteomic experiments upon exposure to sub-inhibitory conditions and showed that the 4f compound interfered with different cellular pathways according to the bacterial species.
2. Results and Discussion
To assay the antibiofilm and antivirulence properties of the promising antibacterial 6-polyaminosteroid derivatives [17], we used resistant archetype strains, i.e., a carbapenem-resistant A. baumannii (CRAB), a carbapenem-resistant P. aeruginosa (CRPA), a vancomycin-resistant E. faecium (VRE), and a methicillin-resistant S. aureus (MRSA).
2.1. Antibiofilm Activity of 6-Polyaminosteroid Derivatives
As described above, one of the virulence features common to all the pathogens is their ability to form biofilms. In this context, we evaluated the impact of 6-polyaminosterols on biofilms (Table 1) by measuring the biofilm prevention concentration (BPC) and the minimal biofilm eradication concentration (MBEC) of these compounds on the four resistant pathogens.

Table 1.
Antibacterial and antibiofilm activities of 6-polyaminosteroid derivatives (mg/L).
As shown in Table 1, 6-polyaminosteroid derivatives showed noticeable antibiofilm activities. Indeed, of the 19 derivatives tested, 5, 2, 14, and 16 prevented the formation of biofilms in CRAB, CRPA, VRE, or MRSA, respectively, with a BPC < IC50. BPC values were very close to the MIC values for all the molecules tested. This may be explained by the bactericidal activity of these compounds, previously demonstrated in [17], which allowed for the prevention of the 24 h biofilm formation as efficiently as the planktonic growth.
Biofilm eradication activity was then determined for derivatives presenting MIC < IC50. Interestingly, some of them allowed for the biofilm eradication of both Gram-negative and Gram-positive resistant bacteria (Table 1). For Gram-negative bacteria, the MBEC values of conventional antibiotics are usually higher than 256 mg/L [18,19,20]. The most active antibiotics on pre-formed biofilms are colistin and aminoglycosides, with MBEC values that can be as low as 32 mg/L on susceptible A. baumannii strains and 160 mg/L on P. aeruginosa [17,19,20]. S. aureus is also highly resistant in biofilm form, with an MBEC higher than 256 mg/L for antibiotics like oxacillin or around 32 mg/L for vancomycin [21]. Some E. faecium isolates can have MBEC values that can reach 128 mg/L for daptomycin, linezolid, or vancomycin [22].
Here, three of the tested compounds presented interesting activities on biofilms of CRAB: 4e, 4r, and 4f with MBEC values of 32, 128, and 256 mg/L, respectively. 4e had biofilm eradication activity similar to that of squalamine and, indeed, they were both as active as the best conventional antibiotics to eradicate biofilms. 4r and 4f possessed similar structures, with either an OH or MeO group in position 3 as well as spermine as a polyamine lateral chain and, interestingly, they had similar biofilm eradication activities. However, against P. aeruginosa, 4f was the only derivative able to eradicate the biofilm at 256 mg/L. For E. faecium, 4e and 4f were also the most potent derivatives, with a biofilm eradication value of 64 mg/L, i.e., more effective than conventional antibiotics. Finally, against MRSA, four derivatives presented MBEC values below or equal to 128 mg/L, i.e., 4b, 4d, 4e, and 4f, and were even more active than squalamine for biofilm eradication. Altogether, 4e (for CRAB, VRE, and MRSA) and 4f (for all the strains) were the more potent 6-polyaminosterols to prevent or eradicate biofilms of these resistant species. It has to be noted that these compounds presented a norspermidine (4e) and a spermine (4f) lateral chain.
Since these 6-polyaminosteroid analogs of squalamine differed essentially by the flexible lateral chain, one can assume that the variations in MICs, BPC, or MBEC can be attributed at least partly to these moieties. Polyamines are small polycationic molecules involved in various bacterial physiological processes [23,24,25]. They can be involved in bacterial growth [26], surface motility, or siderophore synthesis; act as extracellular or intracellular signaling molecules; and regulate biofilm formation [24,27,28]. Numerous studies have reported that the addition of exogenous polyamines to biofilms results in various effects (inhibition or stimulation), depending on the bacterial species [24]. Here, a hypothesis of an effect of the polyamine side chain of derivatives on biofilms can be envisioned. Indeed, it was previously shown that S. aureus does not produce polyamines and presents sensibility to exogenous polyamines (inhibition of planktonic growth [27] at concentrations known to exist within human hosts [29]). A spermidine structural analog was also reported to inhibit biofilm formation and maintenance in S. aureus [30]. Polyamines production, like spermidine or norspermidine, is a species-level trait in enterococci and is encountered in E. faecalis but is rare in E. faecium (absent in our studied strain) [31]. This may explain the VRE hyper-susceptibility to the polyamines studied, like in S. aureus [29]. In P. aeruginosa, which is able to produce polyamines as putrescine or spermidine [32], the effect of exogenous polyamine addition is more mixed and depends on the polyamine tested. The putrescine was shown to promote biofilm formation by acting possibly as a host-associated signal that triggers bacteria lifestyle switching [33]; norspermidine was shown to inhibit biofilm formation and eradicate 24 h mature biofilm in P. aeruginosa PAO1 and clinical strains [34]. A similar effect of the polyamine lateral chain on A. baumannii biofilms is less likely. For this species, which only produces 1,3 diaminopropane but no polyamine with longer carbon chain, it was previously shown that the addition of diverse exogenous polyamines like spermine, spermidine, cadaverine, or putrescine did not impact biofilm development [35]. However, the amphiphilicity of the molecule associated with the positive charge of amines in precise spacing may act as a biofilm disruptor, as it was previously shown [30].
2.2. The Effect of 6-Polyaminosterols on CRPA Pyocyanin Production
To get further insights into the effects of 6-polyaminosterols on CRPA virulence, we assayed the production of one of its major secreted and soluble virulence factors, i.e., pyocyanin. This blue-green pigment is a redox-active phenazine that stimulates redox cycling in bacteria and enhances oxidative metabolism, which, in turn, increases the formation of intracellular reactive oxygen species (ROS) [36]. In the host, pyocyanin inhibits cell growth and ciliary movements and induces cell death and cytokine release [36]. The effect of 6-polyaminosterols on CRPA has been evaluated in a range of concentrations from 0.5 to 32 mg/L. As in the case of MBEC evaluation, we chose to focus on compounds, the MIC of which was below the IC50. Data represent the compound concentration leading to >40% and >70% reductions in terms of pyocyanin production (Table 2).
Table 2.
Anti-pyocyanin activity of 6-polyaminosteroid derivatives (mg/L).
As shown in Table 2, each tested compound led to reduced pyocyanin production by at least 40%. The most efficient 6-polyaminosterol derivatives were indicated as reducing the pyocyanin by more than 70% at a concentration of 32 mg/L, except for compound 4j, which was capable of reducing pyocyanin by 70% at 16 mg/L. Accordingly, the polyaminoisoprenyl compound NV716 was shown to reduce pyocyanin production [37]. Since NV716 was also shown to reduce biofilm formation, it was suggested that reducing pyocyanin production decreased the release of a major matrix component of P. aeruginosa that is extracellular DNA [37]. Noticeably, the compounds that were the most efficient in pyocyanin production also displayed antibiofilm activity, suggesting that a similar mechanism could occur.
2.3. G. mellonella Treatment Assays
The 4f compound was the most potent derivative against all the studied strains. Consequently, we studied the effect of treatment with this compound or with squalamine against bacterial infections of G. mellonella larvae. G. mellonella provides an alternative model to study Gram-positive and Gram-negative bacterial infections. Like all insects, it lacks an adaptive immune response but possesses an innate immune response that shows remarkable similarities with that of vertebrates, allowing for an initial evaluation of antivirulence properties. The use of the G. mellonella model can consequently give indications about the appropriateness of compounds for therapeutic use. Larvae were infected with bacteria and, after 2 h, the 4f or squalamine compounds were introduced separately at 14 mg/kg (or the equivalent of ¼ MIC). Larvae survival was then monitored for 24 h. As shown in Figure 3, the squalamine and 4f compounds significantly prolonged the survival of larvae infected by CRAB, VRE, and MRSA during the first 20 h compared to the control. Moreover, the 4f compound was more efficient than squalamine, with a survival rate ranging from 50% to 80% after 24 h against 0 to 20%, respectively. The equivalent of ¼ MIC of 4f or squalamine was injected into the larvae, so the treatments were not able to inhibit bacterial growth. Consequently, the enhanced survival of larvae was not due to antibiotic activity but rather likely due to the antivirulence properties of this molecule. This is in accordance with data that previously reported that in S. aureus and E. faecalis, the reduced concentration of polyamines due to the activity of acetyltransferase increased bacterial survival within the host [38,39]. This conclusion is also supported by the fact that polyamines attenuate virulence in Gram-positive pathogens like Streptococcus pneumoniae [40] and could be used as a treatment for infectious diseases [25]. It must be noted that no significant effect was observed for CRPA.
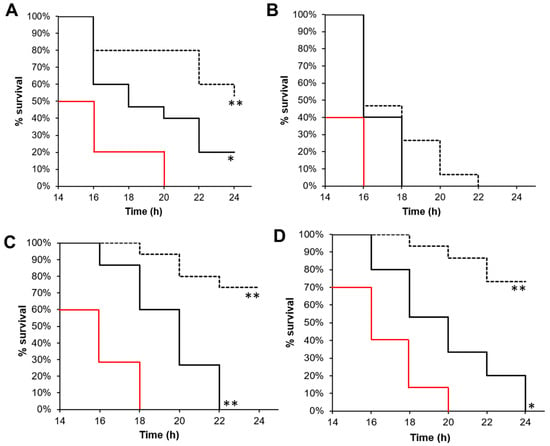
Figure 3.
4f and squalamine antivirulence activities. Survival curves of G. mellonella inoculated with (A) CRAB, (B) CRPA, (C) MRSA, or (D) VRE and treated 2 h post-infection with physiological solution (red line, control), ¼ MIC of squalamine (solid black line), or 4f compound (dotted black line), as described in the methods section. Log-rank test * p < 0.01, ** p < 0.001.
2.4. Contributions to the Mechanism of Action of the 4f Derivative
To gain a better understanding of how this particularly effective compound 4f works, a label-free semi-quantitative proteomic analysis was used to compare the protein expression of the different bacterial species in the presence and absence of exposure to this molecule at a concentration equivalent to ¼ of the MIC. Differentially expressed proteins were characterized by a fold change value greater than 1.4, a p-value < 0.05, and then were categorized according to the Clusters of Orthologous Groups (COG, Figure 4). Following 2 h exposure to a ¼ MIC concentration, we identified 69, 35, 40, and 34 differentially expressed proteins in CRAB, CRPA, MRSA, and VRE, respectively (Figure S1).
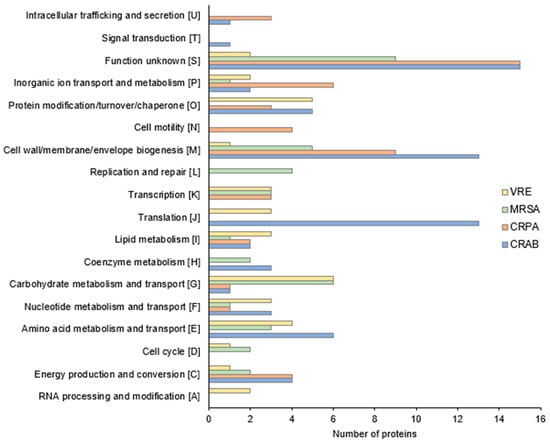
Figure 4.
Functional COG classification of CRAB, CRPA, MRSA, and VRE differentially expressed proteins (fold change > 1.4, p-value < 0.05).
Concerning Gram-negative bacteria, as shown in Figure 4 and Supplementary Tables S1 and S2, 4f exposure induced notable alterations, primarily within the proteins responsible for the biogenesis of the cell wall and membrane, translation, and amino acid metabolic processes.
Interestingly, for A baumannii, we observed that the abundance of proteins involved in lipooligosaccharide biosynthesis (ABYAL2345, +2.6-fold; ABYAL3113, +1.6-fold) and OM lipoprotein transport (ABYAL3248, +1.5-fold) significantly increased following treatment with 4f (Supplementary Table S1). These findings suggest that the principal mode of action of 4f likely involves the OM destabilization of this Gram-negative bacterium through a mechanism resembling detergents [41]. A similar observation was noticed in A. baumannii when exposed to the cationic surfactants chlorhexidine and cetyltrimethylammonium bromide [42]. In line with this mode of action, the exposure of CRAB to 4f also increased the abundance of ABYAL2090 (+4-fold) and ABYAL2091 (+4.5-fold) belonging to the multidrug efflux pump AdeABC (Table S1). A high level of transcription of this pump was indeed induced by benzalkonium and chlorhexidine [42]. These observations were therefore consistent with a mode of action via a detergent-like membrane permeabilization. In addition, proteins associated with oxidative stress defense (superoxide dismutase, +1.6-fold; NADH-quinone oxidoreductase, +1.5-fold; glutathione peroxidase, +1.5-fold; universal stress protein, +1.4-fold) and DNA repair (Excinuclease, +3.4-fold) were also increased in abundance. These proteins could be considered as the detoxification pathway of CRAB, limiting the entry of noxious compounds and the response to the oxidative damage of the cellular death pathway [43,44]. It is possible that 4f exerted its lethal effect by additionally promoting the formation of harmful hydroxyl radicals, a common mechanism of cellular death underlying all classes of bactericidal antibiotics [45]. A decrease in the abundance of a cluster of ribosomal subunit proteins 30S and 50S (ABYAL3632, ABYAL3642, ABYAL2700, ABYAL3638, ABYAL3621, ABYAL0637) was also observed after 4f treatment (Table S1 and Figure 4). These findings hint that 4f did not only impact the cell wall but also potentially hindered the translation machinery of A. baumannii. This translation inhibition and the ability to perturb lipooligosaccharides could elucidate the reduction of A. baumannii virulence in the presence of 4f, as evidenced by the results observed in the G. mellonella experiments.
Proteomic investigations of the effects of 4f on CRPA revealed 35 proteins whose abundance was affected (Supplementary Table S2). The most striking data for P. aeruginosa were that several proteins of the OM or that are involved in OM composition and/or modification were affected in response to 4f exposure. We indeed noticed a lower abundance of six porins, among which were the substrate-specific proteins OprB, OpdC, OpdH, OprQ, and OprG; PhoP/Q; and low Mg2+ inducible OprH. Since P. aeruginosa displays 26 porins, our data indicate that about ¼ of these OM β-barrel proteins were less abundant in the CRPA envelope in response to 4f. Such a deep reduction in OMP production and/or abundance has been previously reported in P. aeruginosa that was subjected to cell envelope stress, as in the case of a sigX mutant [46,47]. SigX is an extracytoplasmic function sigma factor that is activated in response to membrane stresses [48,49]. In addition, 4f exposure led to abundance alterations of proteins involved in the lipopolysaccharide (LPS) biosynthetic and/or modification pathways. Indeed, on one side, the LPS assembly protein LptD, lipid A biosynthetic acyltransferase HtrB1, and A-band LPS synthetic enzyme WbpW were reduced in response to 4f treatment. Interestingly, a lower abundance of WbpW was shown to reduce the uptake of tobramycin [50]. On the other hand, L-Ara4N transferase ArnT was increased upon 4f treatment. Noticeably, the arn cluster is involved in the aminoarabinose modification of LPS, thus decreasing the negative charge of the OM and limiting the uptake of cationic antimicrobials [51]. Altogether, these data suggest that 4f exposure led to reduced LPS biogenesis and the modification of the LPS structure at the level of lipid A, leading possibly to adaptive resistance in a way that could be similar to that observed for polymyxins and cationic peptides [52]. It is thus conceivable that 4f derivatives could disturb the OM by interacting with divalent cation binding sites on LPS, causing the disruption of these sites and promoting 4f uptake, as was suggested for squalamine or claramine A1 [13,15]. This mode of action, different from the detergent-like permeabilization of A. baumannii, may originate from a different composition of the outer membranes of these bacteria as the OM of P. aeruginosa has LPS in its outer leaflet when lipooligosaccharide constitutes the one of A. baumannii.
For VRE, proteomic profiling showed that deregulated proteins were mainly involved in post-translational modification. More precisely, proteins implicated in proteolysis (UPI0001B6F78E, UPI00003C508F, UPI000037F70A) were considerably induced (Figure 4, Supplementary Table S3). We can also observe that important expression changes occurred for the metabolism proteins of carbohydrates (six proteins) and amino-acids (four proteins). We can thus hypothesize that the degradation products triggered by these catabolic activities could act in favor of the toxic effect of compound 4f. Interestingly, in VRE incubated in the presence of 4f, proteins involved in the osmotic stress response appeared to be less abundant. The amounts of the Na+/H+ antiporter NhaC and sodium/dicarboxylate symporter family protein, as well as two enzymes linked to the accumulation of osmoprotectant glycine betaine, were significantly reduced [53]. These data indicate that 4f could impact the ability of Enterococcus to cope with osmotic challenge, which may contribute to its effects on virulence and/or antibacterial activity.
MRSA was the only strain showing the overproduction of proteins involved in DNA replication and repair (Figure 4). In addition, ftsK and hup gene products involved in chromosomal segregation and condensation, respectively, were also more abundant (Supplementary Table S4). Polyamines are positively charged molecules known to interact with DNA, but the correlation between the increased quantity of these polypeptides and the phenotypes observed when Staphylococcus is confronted with the 4f molecule remains to be elucidated. However, the most spectacular overproduction observed when the MRSA cells were challenged with the 4f compound was that of autolysin Atl. Peptidoglycan hydrolases (autolysins) are important enzymes in cell wall growth and turnover and cell separation, but they are also involved in antibiotic-induced lysis [54]. We can thus suspect that the deleterious effect of the 4f molecule in S. aureus was linked to the induction of this lytic enzyme, as observed for penicillin. Of note, Szweda and collaborators proposed that autolysin can be used as a weapon against Staphylococci in combined therapy due to the enhanced antimicrobial activity of some antibiotics [55]. In this context, the 4f molecule, in addition to its interesting antivirulent, antibacterial, and antibiofilm activities associated with the induction of autolysin production, could be a candidate as an “adjuvant” for treatment against MRSA.
In summary, our proteomic data revealed that the 4f molecule triggered distinct cellular responses depending on the bacterial species. The higher number of differentially expressed proteins in Gram-negative bacteria, particularly A. baumannii, highlighted the flexibility of these pathogens to rapidly adapt to changing environments. Proteomic changes in the presence of 4f within the Gram-negative group suggested a significant stress response in these bacteria, particularly affecting their cell envelope. Additionally, the observed alterations in translation, carbohydrate metabolism, and the autolysin process do not exclude them as additive action mechanisms of this compound on both Gram-negative and Gram-positive bacteria. These varied cellular response mechanisms may contribute to explaining the potential antivirulence effects observed in G. mellonella experiments. However, although our proteomic analysis has enabled us to gain a better understanding of the impact of compound 4f on the overall physiology of the bacterial cell, the targets envisaged require confirmation by additional experiments at the molecular level.
3. Materials and Methods
3.1. The Synthesis of 6-Polaminosteroid Derivatives
All the compounds were prepared and fully characterized according to the procedure previously reported [17].
3.2. Bacterial Strains and Culture Conditions
The bacterial strains used in this study are listed in Table 3. Cultures of S. aureus and E. faecium were performed in brain–heart infusion (BHI) broth (Biokar diagnostics, Allonne, France) for preculture, and Mueller–Hinton MHB cation-adjusted (MHBII) (Biokar diagnostics) was used for biofilm formation and incubated at 37 °C without shaking. For A. baumannii and P. aeruginosa, biofilms were grown in MHBII in the dark at 37 °C without shaking.
Table 3.
Strains used in this study.
3.3. Biofilm Prevention and Eradication Assays
For the determination of the BPC, biofilm formation was assessed in MHBII by the microtiter plate test in 96-well polystyrene plates, as previously described, with the following modifications [59]. For S. aureus and E. faecium, the inocula were adjusted to an OD600 nm of 0.2 in MHBII. The 6-polyaminosteroid derivatives (diluted in sterile ultra-pure water) were added at the same time as the inoculum. After 24 h at 37 °C for CRAB, CRPA, and MRSA and 48 h for VRE, biofilms were quantified by crystal violet assay [60]. BPC was defined as the first concentration of compound that prevented the detection of crystal violet staining (OD580 nm ≤ 0.1).
The MBEC for MRSA and VRE was assessed as previously described [60], with the following changes. Biofilms were exposed to 6-polyaminosteroid derivatives in MHBII for 24 h at 37 °C and then washed twice in saline solution (NaCl 0.9%) before the sonication of cells in fresh sterile MHB. The recovery plate was incubated at 37 °C for 24 h. OD600 nm was measured for each well to determine the MBEC value, defined as the lowest concentration of a compound that prevented measurable growth in the recovery medium [61]. For CRAB and CRPA, the MBEC was determined as previously described [20].
All assays were performed at least in duplicate in a minimum of three independent experiments.
3.4. Pyocyanin Quantification Assays
CRPA was grown in MHBII from an initial inoculum adjusted to an OD580 nm of 0.08 in a 96-well microtiter plate at 37 °C for 24 h with shaking (180 rpm). After incubation for 24 h, cell growth was determined by measuring OD580 nm. The pyocyanin quantification assay was carried out as described previously [62,63]. Briefly, one volume of chloroform was used to extract free-cell supernatant samples. Then, ½ volume of 0.5 M HCl was added to the chloroform layer (blue layer). The absorbance of the HCl layer (red-pink layer) was recorded at 520 nm using the Spark 20 M multimode microplate reader controlled by SparkControlTM software Version 2.1 (Tecan Group Ltd., Männedorf, Switzerland) and the data were normalized for bacterial cell density (OD580 nm).
3.5. Antivirulence Assays
The protocol used for these assays was inspired by [64]. Briefly, bacterial strains were cultured in BHI, harvested at OD600 = 0.6, and washed twice in 0.9% NaCl. Cultures were adjusted to reach a density of 2.108, 2.109, 1.108, and 5.106 CFU/mL for MRSA, VRE, CRAB, and CRPA, respectively. Fifteen randomly selected G. mellonella larvae around 280 mg were used for each test group. A 10 µL, an inoculum was injected into the last proleg using a 26 G needle (Terumo, Shibuya City, Japan) and a syringe pump (Thermofisher, Waltham, MA, USA). The inoculum size was checked by plate counting on BHI agar. Saline solution or the 4f compound (based on ¼ MIC or the equivalent of 14 mg/kg for squalamine and 3.5, 14, 1.75, and 7 for 4f in CRAB, CRPA, MRSA, and VRE, respectively) was administered 2 h post-infection. Larvae were incubated at 37 °C and their survival was monitored for 48 h every 2 h, from 16 to 48 h post-infection. Assays were performed in triplicate.
3.6. The Preparation of Protein Extracts and Proteomic Analyses
CRAB and CRPA strains were grown in MHBII broth to an OD600 nm of approximately 0.4. On the other hand, MRSA and VRE strains were grown in MH broth to an OD600 nm of about 0.6. Subsequently, the cells were harvested using centrifugation at 8000× g for 10 min and resuspended in fresh media containing ¼ MIC of the 4f compound (n = 4) or without treatment (control group, n = 4). After a 2 h incubation at 37 °C, cells were harvested by centrifugation (8000× g, 15 min, 4 °C) and washed once in the extraction buffer (50 mM Tris-HCl pH 7.4, 50 mM Na2SO4, 15% glycerol). Cells were then resuspended in the same buffer and lysed by using the FastPrep device (MP Biomedicals, Illkirch Graffenstaden, France). Then, the samples were centrifuged at 4 °C and 12,000× g for 20 min to collect the supernatant. The protein concentration was measured using the Pierce BCA Protein Assay Kit (Thermoscientific, Rockford, IL, USA). The proteomic analysis of four strains under 4f treatment was then conducted in two distinct proteomic laboratories.
Concerning the CRAB and CRPA samples, the enzymatic digestion of protein extracts and quantitative analysis by mass spectrometry analyses were performed as previously described by Sauvage et al. [65] and Robin et al. [20]. Briefly, 25 μg of proteins were loaded onto a 7% polyacrylamide gel (Acrylamide/Bis-Acrylamide 30% [29:1], Sigma-Aldrich, Steinheim, Germany). Migration was performed for a short period of 90 min at 10–20 mA/gel. After Coomassie blue staining, the revealed protein bands were excised and first immersed in a reductive buffer (5 mM DTT, Sigma-Aldrich) and then an alkylated buffer (20 mM iodoacetamide, Sigma-Aldrich). After washing, the gel bands were digested with 1 μg of trypsin (Promega, Madison, WI, USA) overnight at 37 °C. Then, several steps of peptide extraction were performed using acetonitrile (Fisher, Hampton, NH, USA), and the peptides were dried and stored at −20 °C. The CRAB peptide samples were then analyzed using an LTQ Orbitrap Elite (Thermo Fisher Scientific, Bremen, Germany) coupled to an Easy nLC II system (Thermo Fisher Scientific, Bremen, Germany). Peptide samples from CRPA were analyzed on an Orbitrap Eclipse™ Tribrid™ (Thermo Fisher Scientific, San Jose, CA, USA) coupled to an UltiMate™ 3000 RSLCnano (Thermo Fisher Scientific, San Jose, CA, USA). The detailed chromatography and mass spectrometry analysis protocols are provided in the Supplementary Information part. Raw data after MS analysis were processed using Progenesis LC-MS software (Nonlinear Dynamics, version 4.1.6675.48614, Newcastle, UK). The retention times of all samples were aligned on one reference sample within the experiment. One-way analysis of variance (ANOVA) calculations were performed on aligned and normalized data. The MS/MS spectra from retained peptides were identified using Mascot (Matrix Science, Columbus, OH, USA) against the databases restricted to A. baumannii ATCC17978 (http://www.genoscope.cns.fr/, accessed on 12 September 2022) and P. aeruginosa PAO1 (https://www.pseudomonas.com/, accessed on 22 April 2021).
For MRSA and VRE samples, 5 µg of each protein extract was prepared using a modified gel-aided sample preparation protocol [66]. Samples were digested with trypsin/Lys-C overnight at 37 °C. For nano-LC fragmentation, protein or peptide samples were first desalted and concentrated onto a µC18 Omix (Agilent, Santa Clara, CA, USA) before analysis. The analysis of MRSA and VRE peptide samples was carried out on a TIMS-TOF Pro mass spectrometer (Bruker Daltonics, Billerica, MA, USA) with a modified nano-electrospray ion source (CaptiveSpray, Bruker Daltonics) coupled with a NanoElute (Bruker Daltonics) ultra-high-pressure nanoflow chromatography system. The detailed chromatography and mass spectrometry analysis protocols are provided in the Supplemental Information. Before post-processing, the samples were analyzed using Preview software (version 3.7.4, ProteinMetrics, Cupertino, CA, USA) to estimate the quality of the tryptic digestion and predict the post-translational modifications present. The result was used for the “bank research/identification” part. The fragmentation pattern was used to determine the sequence of the peptides. Database searching was performed using Peaks XPro software. Updated UniProt E. faecium and S. aureus databases were used.
To quantify the relative levels of protein abundance between different groups, samples were analyzed using the label-free quantification feature of Progenesis LC-MS (for CRAB and CRPA) and PEAKS XPro software (for MRSA and VRE). A 1.4-fold increase in relative abundance and a p-value < 0.05 using a Student’s t-test from PERSEUS were used to determine the enriched proteins. Volcano plots were performed with Genoppi [67]. Enrichments in molecular processes, cellular processes, and pathways (KEGG/COG) were performed using the ClueGo app (Version 2.5.9) from Cytoscape software (Version 3.8.2). Enrichments were performed using a Bonferroni step-down method. The mass spectrometry proteomics data were deposited in the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD047333 (for CRAB et CRPA) and the iProX partner repository (for VRE and MRSA) with the dataset identifier IPX0007659001.
4. Conclusions
In the current context, where the emergence of antibiotic resistance is extremely worrying, the discovery and development of molecules capable of reducing bacterial virulence appears to be an attractive and alternative strategy against MDR pathogens [68,69]. The 6-polyaminosterol derivatives we tested in this study were previously reported for their interesting activities as bactericidal molecules against major resistant pathogens. They have also presented synergistic or additive activities when associated with imipenem, vancomycin, or oxacillin [17]. Here, we highlighted their capacities to prevent and eradicate the biofilms of these pathogens, with two of them (4e and 4f) being as efficient as classical antibiotics. The determination of 4f anti-virulence properties in the invertebrate infection model of G. mellonella is very promising but requires additional evidence regarding its effectiveness on rodent models or human organoids. Future assessments of 4f anti-virulence activities on a range of clinical strains, including both drug-resistant and susceptible isolates, would also be necessary to confirm its efficacy in the current clinical situation. However, thanks to their potential antivirulence properties, these 6-polyaminosterols could be used as adjuvants in therapeutics to reduce antibiotic dosages and treatment durations, which may contribute to antibiotic resistance mitigation.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/antibiotics13010008/s1, Table S1. Proteins differentially expressed in CRAB after 4f treatment. Table S2. Proteins differentially expressed in CRPA after 4f treatment. Table S3. Proteins differentially expressed in VRE after 4f treatment. Table S4. Proteins differentially expressed in MRSA after 4f treatment. Supplementary Information. Chromatography and mass spectrometry analysis methods. Figure S1. Volcano plots of differentially expressed proteins in CRAB, CRPA, VRE, and MRSA groups after 2 h of 4f treatment.
Author Contributions
Conceptualization, E.D., C.M., S.C., J.M.B. and J.-C.G.; methodology, E.D., C.M., J.M.B., S.C., O.L., J.-C.G., A.T., F.N., M.B., F.D., D.V., A.S., H.L. and B.B.; formal analysis, E.D., C.M., J.M.B., S.C., O.L., J.-C.G., A.T., F.N., M.B., F.D., D.V., A.S., H.L. and B.B.; investigation, E.D., C.M., J.M.B., S.C., J.-C.G., F.N., M.B., D.V., A.S., H.L. and B.B.; writing—original draft preparation, E.D., C.M., J.M.B., S.C., D.V., J.-C.G., H.L., B.B. and A.T.; writing—review and editing, E.D., C.M., S.C., J.M.B., J.-C.G. and F.D.; supervision, E.D., C.M., J.M.B., S.C. and J.-C.G.; project administration, E.D., C.M., S.C. and J.-C.G.; funding acquisition, E.D., C.M., S.C. and J.-C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Normandie Region (SéSAD research network, France) and the European Union in the framework of the ERDF-ESF program 2014–2020. Europe is involved in Normandie through the European Regional Development Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD047333 (for CRAB et CRPA) and the iProX partner repository (for VRE and MRSA) with the dataset identifier IPX0007659001.
Acknowledgments
We thank Nicolas Nalpas of Univ Rouen, Ophélie Grard, Sabrina Gueulle, Isabelle Rincé, and Julien Pontin of Univ Caen for their expert technical expertise and Julie Hardouin for the helpful discussions. The authors thank the Proteomic Platform PISSARO University of Rouen, Mont-Saint-Aignan, France, for the proteome analysis facility.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Maure, A.; Robino, E.; Van der Henst, C. The intracellular life of Acinetobacter baumannii. Trends Microbiol. 2023, 31, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, C.C.; Chen, Y.; Goetzmann, H.S.; Hao, Y.; Borchers, M.T.; Hassett, D.J.; Young, L.R.; Mavrodi, D.; Thomashow, L.; Lau, G.W. Pseudomonas aeruginosa exotoxin pyocyanin causes cystic fibrosis airway pathogenesis. Am. J. Pathol. 2009, 175, 2473–2488. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.; Dockrell, D.H.; Pattery, T.; Lee, D.G.; Cornelis, P.; Hellewell, P.G.; Whyte, M.K.B. Pyocyanin Production by Pseudomonas aeruginosa Induces Neutrophil Apoptosis and Impairs Neutrophil-Mediated Host Defenses In Vivo. J. Immunol. 2005, 174, 3643–3649. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Shang, W.; Peng, H.; Rao, Y.; Zhao, X.; Hu, Z.; Yang, Y.; Hu, Q.; Tan, L.; Xiong, K.; et al. Virulence determinants are required for brain abscess formation through staphylococcus aureus infection and are potential targets of antivirulence factor therapy. Front. Microbiol. 2019, 10, 447050. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef]
- Tan, C.A.Z.; Antypas, H.; Kline, K.A. Overcoming the challenge of establishing biofilms in vivo: A roadmap for Enterococci. Curr. Opin. Microbiol. 2020, 53, 9–18. [Google Scholar] [CrossRef]
- Lebreton, F.; Manson, A.L.; Saavedra, J.T.; Straub, T.J.; Earl, A.M.; Gilmore, M.S. Tracing the Enterococci from Paleozoic Origins to the Hospital. Cell 2017, 169, 849–861.e13. [Google Scholar] [CrossRef]
- Werner, G.; Coque, T.M.; Hammerum, A.M.; Hope, R.; Hryniewicz, W.; Johnson, A.; Klare, I.; Kristinsson, K.G.; Leclercq, R.; Lester, C.H.; et al. Emergence and spread of vancomycin resistance among enterococci in Europe. Eurosurveillance 2008, 13, 19046. [Google Scholar] [CrossRef] [PubMed]
- Bourguet-Kondracki, M.L.; Brunel, J.M. Promises of the Unprecedented Aminosterol Squalamine. Outst. Mar. Mol. Chem. Biol. Anal. 2014, 12, 265–284. [Google Scholar] [CrossRef]
- Salmi, C.; Loncle, C.; Vidal, N.; Letourneux, Y.; Fantini, J.; Maresca, M.; Taïeb, N.; Pagès, J.-M.; Brunel, J.M. Squalamine: An appropriate strategy against the emergence of multidrug resistant gram-negative bacteria? PLoS ONE 2008, 3, e2765. [Google Scholar] [CrossRef]
- Alhanout, K.; Malesinki, S.; Vidal, N.; Peyrot, V.; Rolain, J.M.; Brunel, J.M. New insights into the antibacterial mechanism of action of squalamine. J. Antimicrob. Chemother. 2010, 65, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, M.; Borselli, D.; Rodallec, A.; Peiretti, F.; Vidal, N.; Bolla, J.; Digiorgio, C.; Morrison, K.R.; Wuest, W.M.; Brunel, J.M. Claramines: A New Class of Broad-Spectrum Antimicrobial Agents with Bimodal Activity. ChemMedChem 2018, 13, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, E.; Salmi-Smail, C.; Brunel, J.-M.; Sanchez, P.; Fantini, J.; Maresca, M. Biophysical studies of the interaction of squalamine and other cationic amphiphilic molecules with bacterial and eukaryotic membranes: Importance of the distribution coefficient in membrane selectivity. Chem. Phys. Lipids 2010, 163, 131–140. [Google Scholar] [CrossRef]
- Vergoz, D.; Nilly, F.; Desriac, F.; Barreau, M.; Géry, A.; Lepetit, C.; Sichel, F.; Jeannot, K.; Giard, J.-C.; Garon, D.; et al. 6-Polyaminosteroid Squalamine Analogues Display Antibacterial Activity against Resistant Pathogens. Int. J. Mol. Sci. 2023, 24, 8568. [Google Scholar] [CrossRef]
- Dosler, S.; Karaaslan, E. Inhibition and destruction of Pseudomonas aeruginosa biofilms by antibiotics and antimicrobial peptides. Peptides 2014, 62, 32–37. [Google Scholar] [CrossRef]
- Eze, E.C.; El Zowalaty, M.E.; Pillay, M. Antibiotic resistance and biofilm formation of Acinetobacter baumannii isolated from high-risk effluent water in tertiary hospitals in South Africa. J. Glob. Antimicrob. Resist. 2021, 27, 82–90. [Google Scholar] [CrossRef]
- Robin, B.; Nicol, M.; Le, H.; Tahrioui, A.; Schaumann, A.; Vuillemenot, J.-B.; Vergoz, D.; Lesouhaitier, O.; Jouenne, T.; Hardouin, J.; et al. MacAB-TolC Contributes to the Development of Acinetobacter baumannii Biofilm at the Solid-Liquid Interface. Front. Microbiol. 2022, 12, 785161. [Google Scholar] [CrossRef]
- Brady, A.J.; Laverty, G.; Gilpin, D.F.; Kearney, P.; Tunney, M. Antibiotic susceptibility of planktonic-and biofilm-grown staphylococci isolated from implant-associated infections: Should MBEC and nature of biofilm formation replace MIC? J. Med. Microbiol. 2017, 66, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, A.; Rasmussen, M. Antibiotic regimens with rifampicin for treatment of Enterococcus faecium in biofilms. Int. J. Antimicrob. Agents 2014, 44, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, M.L.; Campilongo, R.; Casalino, M.; Micheli, G.; Colonna, B.; Prosseda, G. Polyamines: Emerging players in bacteria–host interactions. Int. J. Med. Microbiol. 2013, 303, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Michael, A.J. Polyamine function in archaea and bacteria. J. Biol. Chem. 2018, 293, 18693–18701. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Nakada, Y.; Itoh, Y. Identification of the putrescine biosynthetic genes in Pseudomonas aeruginosa and characterization of agmatine deiminase and N-carbamoylputrescine amidohydrolase of the arginine decarboxylase pathway. Microbiology 2003, 149 Pt 3, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Maezato, Y.; Kim, S.H.; Kurihara, S.; Liang, J.; Michael, A.J. Polyamine-independent growth and biofilm formation, and functional spermidine/spermine N-acetyltransferases in Staphylococcus aureus and Enterococcus faecalis. Mol. Microbiol. 2019, 111, 159–175. [Google Scholar] [CrossRef]
- Thongbhubate, K.; Irie, K.; Sakai, Y.; Itoh, A.; Suzuki, H. Improvement of putrescine production through the arginine decarboxylase pathway in Escherichia coli K-12. AMB Express 2021, 11, 168. [Google Scholar] [CrossRef]
- Joshi, G.S.; Spontak, J.S.; Klapper, D.G.; Richardson, A.R. Arginine catabolic mobile element encoded speG abrogates the unique hypersensitivity of Staphylococcus aureus to exogenous polyamines. Mol. Microbiol. 2011, 82, 9–20. [Google Scholar] [CrossRef]
- Böttcher, T.; Kolodkin-Gal, I.; Kolter, R.; Losick, R.; Clardy, J. Synthesis and activity of biomimetic biofilm disruptors. J. Am. Chem. Soc. 2013, 135, 2927–2930. [Google Scholar] [CrossRef]
- Ladero, V.; Fernández, M.; Calles-Enríquez, M.; Sánchez-Llana, E.; Cañedo, E.; Martín, M.C.; Alvarez, M.A. Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Wang, H.; Huang, J.; Liu, Z.; Chen, Q.; Yu, G.; Xu, Z.; Cheng, P.; Liang, Z.; Zhang, L.-H. Spermidine Is an Intercellular Signal Modulating T3SS Expression in Pseudomonas aeruginosa. Microbiol. Spectr. 2022, 10, e00644-22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Hossain, S.S.; Moreira, Z.M.; Haney, C.H. Putrescine and Its Metabolic Precursor Arginine Promote Biofilm and c-di-GMP Synthesis in Pseudomonas aeruginosa. J. Bacteriol. 2022, 204, e00297-21. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; She, P.; Wang, Y.; Liu, F.; Zhang, D.; Chen, L.; Luo, Z.; Xu, H.; Qi, Y.; Wu, Y. Effects of norspermidine on Pseudomonas aeruginosa biofilm formation and eradication. Microbiologyopen 2016, 5, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Armalytė, J.; Čepauskas, A.; Šakalytė, G.; Martinkus, J.; Skerniškytė, J.; Martens, C.; Sužiedėlienė, E.; Garcia-Pino, A.; Jurėnas, D. A polyamine acetyltransferase regulates the motility and biofilm formation of Acinetobacter baumannii. Nat. Commun. 2023, 14, 3531. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.V.; Zychlinsky, A. The bacterial pigment pyocyanin inhibits the NLRP3 inflammasome through intracellular reactive oxygen and nitrogen species. J. Biol. Chem. 2018, 293, 4893–4900. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Brunel, J.-M.; Preusse, M.; Mozaheb, N.; Willger, S.D.; Larrouy-Maumus, G.; Baatsen, P.; Häussler, S.; Bolla, J.-M.; Van Bambeke, F. The membrane-active polyaminoisoprenyl compound NV716 re-sensitizes Pseudomonas aeruginosa to antibiotics and reduces bacterial virulence. Commun. Biol. 2022, 5, 871. [Google Scholar] [CrossRef] [PubMed]
- Martini, C.; Michaux, C.; Bugli, F.; Arcovito, A.; Iavarone, F.; Cacaci, M.; Sterbini, F.P.; Hartke, A.; Sauvageot, N.; Sanguinetti, M.; et al. The polyamine N-acetyltransferase-like enzyme PmvE plays a role in the virulence of Enterococcus faecalis. Infect. Immun. 2015, 83, 364–371. [Google Scholar] [CrossRef]
- Thurlow, L.R.; Joshi, G.S.; Clark, J.R.; Spontak, J.S.; Neely, C.J.; Maile, R.; Richardson, A.R. Functional modularity of the arginine catabolic mobile element contributes to the success of USA300 methicillin-resistant Staphylococcus aureus. Cell Host Microbe 2013, 13, 100–107. [Google Scholar] [CrossRef]
- Ware, D.; Watt, J.; Swiatlo, E. Utilization of Putrescine by Streptococcus pneumoniae During Growth in Choline-limited Medium. J. Microbiol. 2005, 43, 398–405. [Google Scholar]
- Boll, J.M.; Tucker, A.T.; Klein, D.R.; Beltran, A.M.; Brodbelt, J.S.; Davies, B.W.; Trent, M.S. Reinforcing lipid a acylation on the cell surface of Acinetobacter baumannii promotes cationic antimicrobial peptide resistance and desiccation survival. mBio 2015, 6, e00478-15. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Short, F.L.; Hassan, K.A.; Naidu, V.; Pokhrel, A.; Nagy, S.S.; Prity, F.T.; Shah, B.S.; Afrin, N.; Baker, S.; et al. Systematic analyses identify modes of action of ten clinically relevant biocides and antibiotic antagonism in Acinetobacter baumannii. Nat. Microbiol. 2023, 8, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Baharoglu, Z.; Mazel, D. SOS, the formidable strategy of bacteria against aggressions. FEMS Microbiol. Rev. 2014, 38, 1126–1145. [Google Scholar] [CrossRef] [PubMed]
- Esterházy, D.; King, M.S.; Yakovlev, G.; Hirst, J. Production of reactive oxygen species by complex I (NADH:ubiquinone oxidoreductase) from Escherichia coli and comparison to the enzyme from mitochondria. Biochemistry 2008, 47, 3964–3971. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A Common Mechanism of Cellular Death Induced by Bactericidal Antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Gicquel, G.; Bouffartigues, E.; Bains, M.; Oxaran, V.; Rosay, T.; Lesouhaitier, O.; Connil, N.; Bazire, A.; Maillot, O.; Bénard, M.; et al. The Extra-Cytoplasmic Function Sigma Factor SigX Modulates Biofilm and Virulence-Related Properties in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e80407. [Google Scholar] [CrossRef]
- Fléchard, M.; Duchesne, R.; Tahrioui, A.; Bouffartigues, E.; Depayras, S.; Hardouin, J.; Lagy, C.; Maillot, O.; Tortuel, D.; Azuama, C.O.; et al. The absence of SigX results in impaired carbon metabolism and membrane fluidity in Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 17212. [Google Scholar] [CrossRef]
- Chevalier, S.; Bouffartigues, E.; Bazire, A.; Tahrioui, A.; Duchesne, R.; Tortuel, D.; Maillot, O.; Clamens, T.; Orange, N.; Feuilloley, M.G.; et al. Extracytoplasmic function sigma factors in Pseudomonas aeruginosa. Biochim. Biophys. Acta BBA—Gene Regul. Mech. 2019, 1862, 706–721. [Google Scholar] [CrossRef]
- Chevalier, S.; Bouffartigues, E.; Tortuel, D.; David, A.; Tahrioui, A.; Labbé, C.; Barreau, M.; Tareau, A.-S.; Louis, M.; Lesouhaitier, O.; et al. Cell Envelope Stress Response in Pseudomonas aeruginosa. Adv. Exp. Med. Biol. 2022, 1386, 147–184. [Google Scholar] [CrossRef]
- Coleman, S.R.; Blimkie, T.; Falsafi, R.; Hancock, R.E.W. Multidrug adaptive resistance of Pseudomonas aeruginosa swarming cells. Antimicrob. Agents Chemother. 2020, 64, e01999-19. [Google Scholar] [CrossRef]
- Petrou, V.I.; Herrera, C.M.; Schultz, K.M.; Clarke, O.B.; Vendome, J.; Tomasek, D.; Banerjee, S.; Rajashankar, K.R.; Dufrisne, M.B.; Kloss, B.; et al. Structures of aminoarabinose transferase ArnT suggest a molecular basis for lipid A glycosylation. Science 2016, 351, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Fernández, L.; Jenssen, H.; Bains, M.; Wiegand, I.; Gooderham, W.J.; Hancock, R.E.W. The two-component system CprRS senses cationic peptides and triggers adaptive resistance in Pseudomonas aeruginosa independently of ParRS. Antimicrob. Agents Chemother. 2012, 56, 6212–6222. [Google Scholar] [CrossRef] [PubMed]
- Pichereau, V.; Bourot, S.; Flahaut, S.; Blanco, C.; Auffray, Y.; Bernard, T. The osmoprotectant glycine betaine inhibits salt-induced cross-tolerance towards lethal treatment in Enterococcus faecalis. Microbiology 1999, 145, 427–435. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Takahashi, J.; Komatsuzawa, H.; Yamada, S.; Nishida, T.; Labischinski, H.; Fujiwara, T.; Ohara, M.; Yamagishi, J.-I.; Sugai, M. Molecular Characterization of an atl Null Mutant of Staphylococcus aureus. Microbiol. Immunol. 2002, 46, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Szweda, P.; Schielmann, M.; Kotlowski, R.; Gorczyca, G.; Zalewska, M.; Milewski, S. Peptidoglycan hydrolases-potential weapons against Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2012, 96, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Takeuchi, F.; Kuroda, M.; Yuzawa, H.; Aoki, K.-I.; Oguchi, A.; Nagai, Y.; Iwama, N.; Asano, K.; Naimi, T.; et al. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet 2002, 359, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.M.C.; Seemann, T.; Bulach, D.M.; Gladman, S.L.; Chen, H.; Haring, V.; Moore, R.J.; Ballard, S.; Grayson, M.L.; Johnson, P.D.R.; et al. Comparative analysis of the first complete Enterococcus faecium genome. J. Bacteriol. 2012, 194, 2334–2341. [Google Scholar] [CrossRef]
- Rolain, J.-M.; Diene, S.M.; Kempf, M.; Gimenez, G.; Robert, C.; Raoult, D. Real-Time Sequencing to Decipher the Molecular Mechanism of Resistance of a Clinical Pan-Drug-Resistant Acinetobacter baumannii Isolate from Marseille, France. Antimicrob. Agents Chemother. 2013, 57, 592. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef]
- Abad, L.; Josse, J.; Tasse, J.; Lustig, S.; Ferry, T.; Diot, A.; Laurent, F.; Valour, F. Antibiofilm and intraosteoblastic activities of rifamycins against Staphylococcus aureus: Promising in vitro profile of rifabutin. J. Antimicrob. Chemother. 2020, 75, 1466–1473. [Google Scholar] [CrossRef]
- Macia, M.D.; Rojo-Molinero, E.; Oliver, A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin. Microbiol. Infect. 2014, 20, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Tahrioui, A.; Ortiz, S.; Azuama, O.C.; Bouffartigues, E.; Benalia, N.; Tortuel, D.; Maillot, O.; Chemat, S.; Kritsanida, M.; Feuilloley, M.; et al. Membrane-Interactive Compounds from Pistacia lentiscus L. Thwart Pseudomonas aeruginosa Virulence. Front. Microbiol. 2020, 11, 543891. [Google Scholar] [CrossRef] [PubMed]
- Azuama, O.C.; Ortiz, S.; Quirós-Guerrero, L.; Bouffartigues, E.; Tortuel, D.; Maillot, O.; Feuilloley, M.; Cornelis, P.; Lesouhaitier, O.; Grougnet, R.; et al. Tackling Pseudomonas aeruginosa Virulence by Mulinane-Like Diterpenoids from Azorella atacamensis. Biomolecules 2020, 10, 1626. [Google Scholar] [CrossRef] [PubMed]
- Khader, R.; Tharmalingam, N.; Mishra, B.; Felix, L.; Ausubel, F.M.; Kelso, M.J.; Mylonakis, E. Characterization of Five Novel Anti-MRSA Compounds Identified Using a Whole-Animal Caenorhabditis elegans/Galleria mellonella Sequential-Screening Approach. Antibiotics 2020, 9, 449. [Google Scholar] [CrossRef] [PubMed]
- Sauvage, S.; Gaviard, C.; Tahrioui, A.; Coquet, L.; Le, H.; Alexandre, S.; Ben Abdelkrim, A.; Bouffartigues, E.; Lesouhaitier, O.; Chevalier, S.; et al. Impact of Carbon Source Supplementations on Pseudomonas aeruginosa Physiology. J. Proteome Res. 2022, 21, 1392–1407. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Kessler, B.M. Gel-aided sample preparation (GASP)—A simplified method for gel-assisted proteomic sample generation from protein extracts and intact cells. Proteomics 2015, 15, 1224–1229. [Google Scholar] [CrossRef] [PubMed]
- Pintacuda, G.; Lassen, F.H.; Hsu, Y.-H.H.; Kim, A.; Martín, J.M.; Malolepsza, E.; Lim, J.K.; Fornelos, N.; Eggan, K.C.; Lage, K. Genoppi is an open-source software for robust and standardized integration of proteomic and genetic data. Nat. Commun. 2021, 12, 2580. [Google Scholar] [CrossRef]
- Hibbert, T.M.; Whiteley, M.; Renshaw, S.A.; Neill, D.R.; Fothergill, J.L. Emerging strategies to target virulence in Pseudomonas aeruginosa respiratory infections. Crit. Rev. Microbiol. 2023, 10, 1–16. [Google Scholar] [CrossRef]
- Hotinger, J.A.; Morris, S.T.; May, A.E. The Case against Antibiotics and for Anti-Virulence Therapeutics. Microorganisms 2021, 9, 2049. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).