Abstract
Background: Ceftazidime-avibactam was approved by the FDA to treat infections caused by Enterobacterales carrying blaKPC-2. However, variants of KPC-2 with amino acid substitutions at position 179 have emerged and confer resistance to ceftazidime-avibactam. Methods: The activity of imipenem-relebactam was assessed against a panel of 19 KPC-2 D179 variants. KPC-2 and the D179N and D179Y variants were purified for biochemical analyses. Molecular models were constructed with imipenem to assess differences in kinetic profiles. Results: All strains were susceptible to imipenem–relebactam, but resistant to ceftazidime (19/19) and ceftazidime-avibactam (18/19). KPC-2 and the D179N variant hydrolyzed imipenem, but the D179N variant’s rate was much slower. The D179Y variant was unable to turnover imipenem. All three β-lactamases hydrolyzed ceftazidime at varying rates. The acylation rate of relebactam for the D179N variant was ~2.5× lower than KPC-2. Poor catalytic turnover by the D179Y variant precluded the determination of inhibitory kinetic parameters. Acyl-complexes with imipenem and ceftazidime were less prevalent with the D179N variant compared to the D179Y variant, supporting the kinetic observations that the D179Y variant was not as active as the D179N variant. Relebactam was slower to form an acyl-complex with the D179Y variant compared to avibactam. The D179Y model with imipenem revealed that the catalytic water molecule was shifted, and the carbonyl of imipenem was not within the oxyanion hole. Conversely in the D179N model, imipenem was oriented favorably for deacylation. Conclusions: Imipenem–relebactam overcame the resistance of the D179 variants, suggesting that this combination will be active against clinical isolates harboring these derivatives of KPC-2.
Keywords:
KPC-2; carbapenemase; diazabicyclooctane; beta-lactam; beta-lactamase; relebactam; imipenem; ceftazidime; avibactam 1. Introduction
The pace of β-lactamase evolution is outcompeting scientific advances in the development of novel therapeutics [1]. Serine β-lactamases that hydrolyze carbapenems are a significant threat to our antibiotic armamentarium [2,3]. KPC enzymes are the most prevalent carbapenemases in the United States and are most often found in Enterobacterales [4]. Prior to the approval of ceftazidime-avibactam, meropenem–vaborbactam, and imipenem–relebactam by the US Food and Drug Administration (FDA), clinicians had limited options to treat patients who acquired infections by a KPC-2/3 producing Enterobacterales [5]. Polymyxins, more toxic antimicrobials, were often employed. Thus, the approval of ceftazidime-avibactam, meropenem–vaborbactam, and imipenem–relebactam was a renaissance in the treatment of these infections. The β-lactamase inhibitor partners, avibactam, vaborbactam, and relebactam potently inactivated KPC-2/3, unlike the preceding β-lactamase inhibitors approved in the 1980/90s, clavulanic acid, sulbactam and tazobactam, which were hydrolyzed by KPC [6,7,8,9,10].
However, with the more frequent clinical use of these novel β-lactam-β-lactamase inhibitor combinations, there were increasing reports of ceftazidime-avibactam-resistant KPC-producing Enterobacterales isolates [1,3,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43]. Alarmingly, resistance to ceftazidime-avibactam often emerged during treatment. In these instances, many clinicians switched therapies to meropenem–vaborbactam or imipenem–relebactam. Unlike vaborbactam, which is a monocyclic boronate, relebactam is similar in structure to avibactam. One of the most common resistance mechanisms that KPC-producing Enterobacterales acquire is a single amino acid substitution at Ambler position 179. These variants of KPC possess the characteristics of extended-spectrum β-lactamases (ESBLs) with susceptibility to carbapenems [24,35].
The D179Y amino acid substitution in KPC is one of the most prevalent substitutions found in ceftazidime-avibactam-resistant clinical isolates to date [11,13,14,21,23,35,36,37,39,42,43]. However, the D179N amino acid substitution in KPCs is more problematic as strains that carry this substitution maintain resistance towards penicillins, cephalosporins, aztreonam, narrow-spectrum β-lactam-β-lactamase inhibitor combinations (e.g., ampicillin-clavulanic) and carbapenems [44,45,46]. Unlike the other D179 variants, the D179N variant of KPC-2 is of particular interest because it confers resistance to imipenem, and ceftazidime, in addition to ceftazidime-avibactam; the clinical implications of this “heightened” resistance are alarming [44,47]. The impact of 179 substitutions in KPC on meropenem–vaborbactam was previously evaluated and found to not alter the activity of this combination [7]. Herein, imipenem–relebactam, which is effective at treating Enterobacterales producing KPCs as well as AmpCs and ESBLs, was evaluated against ceftazidime-avibactam resistant KPC-2 variants.
2. Results
2.1. D179 Variants Are Susceptible to Imipenem–Relebactam When Expressed in Escherichia coli
Susceptibility testing was conducted using imipenem (IMI), imipenem–relebactam (IMI-REL), ceftazidime (CAZ), and ceftazidime-avibactam (CAZ-AVI) using isogenic Escherichia coli strains expressing KPC-2 as well as D179 variants possessing the 19 other amino acid substitutions (Table 1). Only wild-type KPC-2 and the D179N variant tested resistant to imipenem alone (MIC ≥ 4 µg/mL). Notably, the addition of relebactam to imipenem restored susceptibility to all strains. The same strains were highly resistant to ceftazidime and ceftazidime-avibactam with all D179 variants testing resistant to ceftazidime and only one strain (E. coli DH10B pBC SK(+) blaKPC-2 D179E; MIC = 8 µg/mL) being susceptible to ceftazidime-avibactam.

Table 1.
Susceptibility testing results for Ambler position D179 site-saturation blaKPC-2 mutants expressed in E. coli DH10B.
2.2. The D179Y Variant Is Less Catalytically Active Compared to the D179N Variant and KPC-2
KPC-2 (blue line) and the D179N variant (yellow line) both hydrolyzed imipenem though the overall rate of hydrolysis by the D179N variant was much slower (Figure 1a). The D179Y variant was unable to turnover imipenem (orange line) and was comparable to the background breakdown of imipenem alone (gray line) (Figure 1a). Conversely, all three β-lactamases hydrolyzed ceftazidime. KPC-2 hydrolyzed 25 µM ceftazidime within 300 sec (blue line) (Figure 1b). Conversely, the two variants had slower rates of hydrolysis for ceftazidime, but the rates were similar (yellow and orange lines); the background breakdown of ceftazidime is shown by a gray line (Figure 1b).
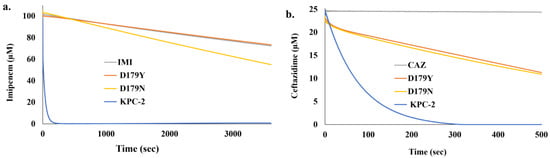
Figure 1.
Progress curves for the hydrolysis of (a) 100 µM imipenem (IMI) and (b) 25 µM ceftazidime (CAZ) by KPC-2 and the D179Y and D179N variants (2 µM enzyme) at 25 °C obtained using a stopped-flow apparatus.
The D179N variant possessed a similar Ki app value for relebactam, as compared to KPC-2, 3.4 ± 0.3 µM and 2.3 ± 0.3 µM, respectively (Table 2). However, the acylation rate of relebactam for the D179N variant was ~2.5x lower than KPC-2. The partition ratios or kinact/kcat values for KPC-2 and the D179N variant were both 1. Due to the poor catalytic turnover by the D179Y variant, inhibition of this variant was not able to be assessed.
Table 2.
Kinetic inhibition constants of the D179N variant compared to wild-type KPC-2.
2.3. Timed Mass Spectrometry Supports the Kinetic Observations That the D179Y Variant Is Slower to Deacylate Imipenem and Ceftazidime, but Also Slower to Be Acylated by Relebactam
Time-based mass spectrometry was used to compare the D179N and D179Y variants upon reaction with imipenem, ceftazidime, relebactam, and avibactam. The presence of acyl-complexes with imipenem and ceftazidime with the D179N variant was captured less often compared to the D179Y variant (Figure 2).
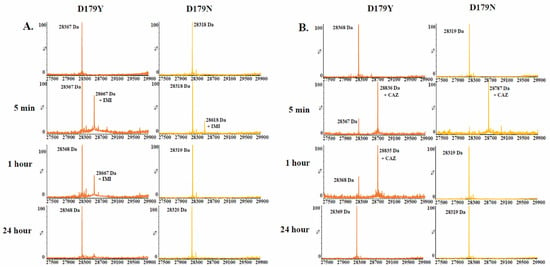
Figure 2.
Time-based mass spectrometry (A) 1:1 ratio of the D179Y and D179N variants to imipenem. (B) 1:50 ratio of variant to ceftazidime.
Comparing relebactam and avibactam using time-based mass spectrometry revealed that relebactam is slower to form an acyl complex with the D179Y variant compared to avibactam (Figure 3). Relebactam and avibactam behaved similarly vs. the D179N variant.
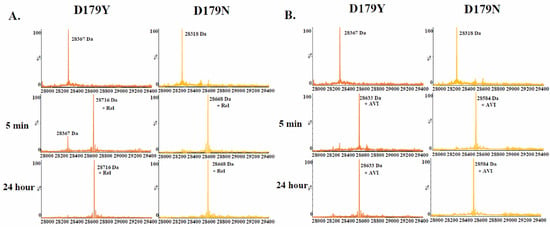
Figure 3.
Time-based mass spectrometry (A) 1:1 ratio of the D179Y and D179N variants to relebactam. (B) 1:1 ratio of variant to avibactam.
2.4. Molecular Modeling
To assess the differences in the biochemical analysis conducted with the purified D179Y and D179N variants, molecular models were constructed with imipenem bound to the nucleophile, S70 when modeling using parameters previously employed [46,49,50]. The D179Y variant revealed that the carbonyl of imipenem is positioned outside of the oxyanion hole, toward S130 (Figure 4a) but inside the oxyanion hole for the D179N variant (Figure 4b). Moreover, the catalytic water is displaced by the ethoxy group of imipenem in the D179Y variant, but not in D179N.
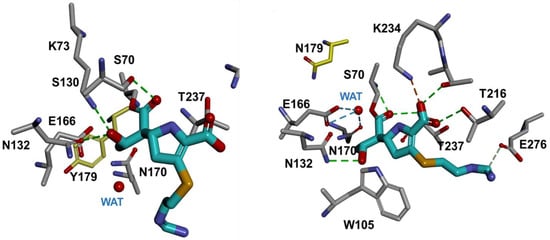
Figure 4.
The acyl–enzyme complexes of the D179Y variant (left) and the D179N variant (right) with imipenem (cyan).
3. Discussion
Imipenem–relebactam was shown to demonstrate antimicrobial activity against ceftazidime-avibactam-resistant D179 variants of KPC-2, including the imipenem-resistant D179N variant. The limitations of our study because of technical challenges, included the susceptibility testing of three blaKPC-2 mutants in a different vector as well as the inability to measure inhibition kinetics with the D179Y variant.
Kinetic and time-based mass spectrometry revealed that the D179N variant hydrolyzed imipenem albeit at a slower rate than KPC-2; however, the D179Y variant was unable to turnover imipenem. Both variants hydrolyzed ceftazidime at similar rates, but not as robustly as KPC-2. Despite similar Ki app values between KPC-2 and the D179N variant for relebactam, the acylation rate was ~2.5-fold lower for the D179N variant compared to KPC-2. Thus, the D179N variant is not as active as KPC-2 vs. β-lactams and is also less inhibited by relebactam. Time-based mass spectrometry showed that the D179Y variant had an even slower relebactam acylation rate as at 5 min apo-enzyme could still be detected. Thus, the mass spectrometry supported the kinetic observations that the D179Y variant was not as active as the D179N variant. Likely the acylation rate of relebactam for the D179Y variant is even lower than that observed for the D179N variant. Overall, these biochemical parameters translate to imipenem resistance and imipenem–relebactam susceptibility when the D179N variant is expressed in E. coli, while the D179Y variant is susceptible to both agents.
Molecular modeling provided insights into the varying phenotypes of the D179N and D179Y variants vs. imipenem. Upon docking and acylation of imipenem into the active sites of both variants, imipenem is well positioned for hydrolysis in the D179N variant, but not the D179Y variant (i.e., carbonyl positioned outside of the oxyanion hole). Structurally, the D179N variant is more poised for efficient catalysis of imipenem, which supports the increased imipenem hydrolysis by D179N compared to the D179Y variant. The molecular modeling of imipenem is reminiscent of crystal structures of carbapenems bound to class A non-carbapenemases, such as SHV-1 and TEM-1 [51,52]. With SHV-1, the carbonyl of meropenem was found outside and inside the oxyanion hole, while the carbonyl of imipenem was outside the oxyanion hole in TEM-1; both structures support slow hydrolysis of or inhibition by carbapenems. In conclusion, these studies complement previous investigations by elucidating how ceftazidime–avibactam resistant variants at Ambler position 179 (e.g., D179Y) test susceptible to imipenem.
4. Materials and Methods
4.1. Bacterial Strains and Reagents
The K. pneumoniae isolates carrying blaKPC-2 and pBR322-catI-blaKPC-2 were kind gifts from Fred Tenover of the Centers for Disease Control and Prevention (Atlanta, GA, USA) [53]. Site saturation mutagenesis at the nucleotides corresponding to position 179 in blaKPC-2 in the pBR322-catI plasmid was previously described [44]. Due to technical difficulties, the blaKPC-2 D179E, D179I, or D179S genes were purchased from Celtek Genes Service a subdivision of Celtek Bioscience, LLC (Franklin, TN, USA), and cloned into the pBC SK(+) phagemid, as previously described [44]. All plasmids were maintained into E. coli DH10B. For protein purification, the construction of pET24a(+)blaKPC-2 and pET24a(+)blaKPC-2 D179N plasmids was previously described [44,48] and the pET24a(+)blaKPC-2 D179Y plasmid was created using a similar approach. These plasmids were maintained into E. coli DH10B and transformed into E. coli Origami™2 DE3 before protein production.
Imipenem and relebactam were provided by Merck (Merck, Sharp & Dohme, Kenilworth, NJ, USA). Nitrocefin was purchased from Oxoid (ThermoFisher Scientific, Basingstoke, UK). Chloramphenicol was obtained from Sigma-Aldrich (St. Louis, MI, USA). Ceftazidime was procured from Sigma-Aldrich and Research Products International and used interchangeably throughout the experimentation. Avibactam was purchased from Advanced ChemBlocks (cat # R16073).
4.2. Susceptibility Testing
Cation-adjusted Mueller-Hinton (MH) agar-dilution minimum inhibitory concentration (MIC) measurements were performed according to Clinical and Laboratory Standards Institute (CLSI) guidelines [47,54]. A Steers replicator was used to deliver 10-μL of each bacterial inoculum containing 104 CFU per spot directly onto the MH agar plates. Plates were incubated for 16–20 h at 37 °C and MIC values were assessed based on a 100% decrease in growth on the drug plates compared to the MH-only control plate. Relebactam and avibactam were tested at a constant 4 µg/mL. MIC measurements were performed in at least triplicate.
4.3. Protein Expression and Purification
The KPC-2, D179N, and D179Y variant β-lactamases were purified from E. coli Origami™2 DE3 (Novagen, Burlington, ON, USA), as previously described [44,48]. Briefly, cells were grown in super optimal broth (SOB) at 37 °C to an optical density at λ600 nm (OD600) of approximately 0.6–0.8 and induced with 0.5 mM isopropyl β-d-1-thiogalactopyranoside (IPTG) for a minimum of three hours to express the β-lactamase. Pelleted cells were frozen and lysed with lysozyme and the supernatants were further purified by preparative isoelectric focusing and fast protein liquid chromatography (FPLC) using a HiTrap Q anion-exchange chromatography column. Proteins were frozen with 25% glycerol and stored at −20 °C. The purity of the proteins was assessed by mass spectrometry. Protein concentrations were determined by measuring absorbance at a wavelength of λ280 nm and using the protein’s extinction coefficient (Δε 39,545 M−1 cm−1) obtained using the ProtParam tool at ExPASy Bioinformatics Resource Portal [55].
4.4. Kinetics
For progress curves of imipenem and ceftazidime hydrolysis, 2.0 μM of KPC-2 and the D179Y and D179N variants were incubated with either 100 μM imipenem or 25 μM ceftazidime in sterile 10 mM PBS pH 7.4 at 25 °C. Data were collected using an Applied Photophysics SX20 Stopped Flow spectrophotometer (260 nm) with the ProData SX software version 1.0.
Kinetic inhibition assays were conducted using an Agilent 8453 diode array spectrophotometer in 10 mM phosphate-buffered saline, pH 7.4 at room temperature. Determination of the values of the kinetic constants apparent Ki (Ki app), k2/K, and kcat/kinact, was previously described [56].
4.5. Timed Mass Spectrometry
The D179Y and D179N variants were incubated with imipenem, relebactam, or avibactam at an enzyme/compound molar ratio of 1:1 and ceftazidime at a molar ratio of 1:50 in sterile 10 mM phosphate-buffered saline (PBS) at pH 7.4 for 5 min and 24 h. Reactions were stopped by adding 10 μL acetonitrile and samples were transferred into 0.1% formic acid in water. Samples were analyzed using Q-TOF Waters Synapt-G2-Si and Waters Acquity UPLC BEH C18 1.7 µm column (2.1 mm × 50 mm), as previously described [44].
4.6. Molecular Modeling
The crystal coordinates of KPC-2 (PDB:2OV5) were used to generate structural representations of the D179Y and D179N variants using Discovery Studio (Acclerys Inc., San Diego, CA, USA) as previously described [49,50,57]. The crystallographic water molecules were maintained during modeling. The KPC-2 β-lactamase structure and the variant model were solvated and minimized to an RMS of 0.03 Å using the Conjugate Gradient method. Acyl–imipenem was constructed using the Fragment Builder tools and minimized using a Standard Dynamics Cascade protocol of Discovery Studio. The acylated imipenem was automatically docked into the active site of the D179Y and D179N variants using the CDOCKER module of Discovery Studio.
5. Conclusions
These studies also complement previous investigations [44,45,46,47] by proposing hypotheses to explain why variants such as KPC-2 D179Y became susceptible to imipenem when expressed in E. coli. Our data suggest that imipenem adopts a catalytically unfavorable conformation in the active site reminiscent of studies done with other carbapenems in non-carbapenemase β-lactamases [51,52]. Thus, imipenem serves as a “β-lactamase inhibitor.” Importantly, there also seems to be a reduction in the ability of relebactam to inactivate D179Y compared to avibactam, although this is not evident by susceptibility testing. The implication of these paradoxical findings only further compels us to investigate these complex β-lactamases to anticipate future development of resistance as new combinations are introduced into the clinic. Moreover, novel KPC variants that confer ceftazidime–avibactam resistance will pose even more challenges as they become more prevalent [58].
Author Contributions
Conceptualization, K.M.P.-W., M.D.B., K.Y. and R.A.B.; methodology, M.D.B., J.D.R., C.R.B., E.T.Z. and M.A.T.; formal analysis, K.M.P.-W., M.D.B. and M.A.T.; resources, R.A.B.; writing—original draft preparation, K.M.P.-W.; writing—review and editing, K.M.P.-W., M.D.B. and R.A.B.; supervision, K.M.P.-W. and R.A.B.; project administration, K.M.P.-W. and R.A.B.; funding acquisition, R.A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by funds and/or facilities provided by an investigator grant from Merck, Sharp & Dohme, Kenilworth, NJ USA; MISP #57394 to RAB. Research reported in this publication was supported in part by funds and/or facilities provided by the Cleveland Department of Veterans Affairs to KMPW and RAB, the Veterans Affairs Merit Review Program Award 1I01BX002872 to KMPW and the Veterans Affairs Merit Review Program Award 1I01BX001974 to RAB from the Biomedical Laboratory Research & Development Service of the VA Office of Research and Development and the Geriatric Research Education and Clinical Center VISN 10 to RAB. The contents do not represent the views of the U.S. Department of Veterans Affairs or the U.S. Government.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available upon request.
Conflicts of Interest
K.Y. is an employee of Merck Sharpe and Dohme. All other authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bush, K.; Page, M.G. What we may expect from novel antibacterial agents in the pipeline with respect to resistance and pharmacodynamic principles. J. Pharmacokinet. Pharmacodyn. 2017, 44, 113–132. [Google Scholar] [CrossRef]
- Drawz, S.M.; Papp-Wallace, K.M.; Bonomo, R.A. New β-lactamase inhibitors: A therapeutic renaissance in an MDR world. Antimicrob. Agents Chemother. 2014, 58, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Bonomo, R.A. Ceftazidime-avibactam and carbapenem-resistant enterobacteriaceae: “We’re Gonna Need a Bigger Boat”. Clin. Infect. Dis. 2016, 63, 1619–1621. [Google Scholar] [CrossRef]
- Sader, H.S.; Mendes, R.E.; Duncan, L.; Kimbrough, J.H.; Carvalhaes, C.G.; Castanheira, M. Ceftazidime-avibactam, meropenem-vaborbactam, and imipenem-relebactam activities against multidrug-resistant Enterobacterales from United States Medical Centers (2018–2022). Diagn. Microbiol. Infect. Dis. 2023, 106, 115945. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Bonomo, R.A. New beta-lactamase inhibitors in the clinic. Infect. Dis. Clin. N. Am. 2016, 30, 441–464. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Barnes, M.D.; Alsop, J.; Taracila, M.A.; Bethel, C.R.; Becka, S.A.; van Duin, D.; Kreiswirth, B.N.; Kaye, K.S.; Bonomo, R.A. Relebactam is a potent inhibitor of the KPC-2 β-lactamase and restores imipenem susceptibility in KPC-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e00174-18. [Google Scholar] [CrossRef] [PubMed]
- Tsivkovski, R.; Lomovskaya, O. Potency of vaborbactam is less affected than that of avibactam in strains producing KPC-2 mutations that confer resistance to ceftazidime-avibactam. Antimicrob. Agents Chemother. 2020, 64, e01936-19. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Bethel, C.R.; Distler, A.M.; Kasuboski, C.; Taracila, M.; Bonomo, R.A. Inhibitor resistance in the KPC-2 beta-lactamase, a preeminent property of this class A beta-lactamase. Antimicrob. Agents Chemother. 2010, 54, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Ehmann, D.E.; Jahic, H.; Ross, P.L.; Gu, R.F.; Hu, J.; Durand-Reville, T.F.; Lahiri, S.; Thresher, J.; Livchak, S.; Gao, N.; et al. Kinetics of avibactam inhibition against Class A, C, and D beta-lactamases. J. Biol. Chem. 2013, 288, 27960–27971. [Google Scholar] [CrossRef]
- Tsivkovski, R.; Lomovskaya, O. Biochemical activity of vaborbactam. Antimicrob. Agents Chemother. 2020, 64, e01935-19. [Google Scholar] [CrossRef]
- Athans, V.; Neuner, E.A.; Hassouna, H.; Richter, S.S.; Keller, G.; Castanheira, M.; Brizendine, K.D.; Mathers, A.J. Meropenem-vaborbactam as salvage therapy for ceftazidime-avibactam-resistant Klebsiella pneumoniae bacteremia and abscess in a liver transplant recipient. Antimicrob. Agents Chemother. 2019, 63, e01551-18. [Google Scholar] [CrossRef] [PubMed]
- Bianco, G.; Boattini, M.; Comini, S.; Iannaccone, M.; Bondi, A.; Cavallo, R.; Costa, C. In vitro activity of cefiderocol against ceftazidime-avibactam susceptible and resistant KPC-producing Enterobacterales: Cross-resistance and synergistic effects. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Guzman-Puche, J.; Garcia-Gutierrez, M.; Caston, J.J.; Gracia-Ahufinger, I.; Perez-Nadales, E.; Recio, M.; Natera, A.M.; Marfil-Perez, E.; Martinez-Martinez, L.; et al. Use of carbapenems in the combined treatment of emerging ceftazidime/avibactam-resistant and carbapenem-susceptible KPC-producing Klebsiella pneumoniae infections: Report of a case and review of the literature. J. Glob. Antimicrob. Resist. 2020, 22, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Arends, S.J.R.; Davis, A.P.; Woosley, L.N.; Bhalodi, A.A.; MacVane, S.H. Analyses of a ceftazidime-avibactam-resistant Citrobacter freundii isolate carrying blaKPC-2 reveals a heterogenous population and reversible genotype. mSphere 2018, 3, e00408-18. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Polo, J.A.; Hernandez-Garcia, M.; Morosini, M.I.; Perez-Viso, B.; Soriano, C.; De Pablo, R.; Canton, R.; Ruiz-Garbajosa, P. Outbreak by KPC-62-producing ST307 Klebsiella pneumoniae isolates resistant to ceftazidime/avibactam and cefiderocol in a university hospital in Madrid, Spain. J. Antimicrob. Chemother. 2023, 78, 1259–1264. [Google Scholar] [CrossRef]
- Cavallini, S.; Unali, I.; Bertoncelli, A.; Cecchetto, R.; Mazzariol, A. Ceftazidime/avibactam resistance is associated with different mechanisms in KPC-producing Klebsiella pneumoniae strains. Acta Microbiol. Immunol. Hung. 2021, 68, 235–239. [Google Scholar] [CrossRef]
- Compain, F.; Arthur, M. Impaired inhibition by avibactam and resistance to the ceftazidime-avibactam combination due to the D(179)Y substitution in the KPC-2 beta-lactamase. Antimicrob. Agents Chemother. 2017, 61, e00451-17. [Google Scholar] [CrossRef]
- Corcione, S.; De Benedetto, I.; Shbaklo, N.; Torsello, G.; Lupia, T.; Bianco, G.; Cavallo, R.; Brazzi, L.; Montrucchio, G.; De Rosa, F.G. Ceftazidime-avibactam (C/A) resistant, meropenem sensitive KPC-producing Klebsiella pneumoniae in ICU setting: We are what we are treated with? Int. J. Mol. Sci. 2023, 24, 4767. [Google Scholar] [CrossRef]
- Di Bella, S.; Giacobbe, D.R.; Maraolo, A.E.; Viaggi, V.; Luzzati, R.; Bassetti, M.; Luzzaro, F.; Principe, L. Resistance to ceftazidime/avibactam in infections and colonisations by KPC-producing Enterobacterales: A systematic review of observational clinical studies. J. Glob. Antimicrob. Resist. 2021, 25, 268–281. [Google Scholar] [CrossRef]
- Fontana, C.; Favaro, M.; Campogiani, L.; Malagnino, V.; Minelli, S.; Bossa, M.C.; Altieri, A.; Andreoni, M.; Sarmati, L. Ceftazidime/avibactam-resistant Klebsiella pneumoniae subsp. pneumoniae isolates in a tertiary Italian hospital: Identification of a new mutation of the carbapenemase type 3 (KPC-3) gene conferring ceftazidime/avibactam resistance. Microorganisms 2021, 9, 2356. [Google Scholar] [CrossRef]
- Gaibani, P.; Re, M.C.; Campoli, C.; Viale, P.L.; Ambretti, S. Bloodstream infection caused by KPC-producing Klebsiella pneumoniae resistant to ceftazidime/avibactam: Epidemiology and genomic characterization. Clin. Microbiol. Infect. 2020, 26, 516.e511–516.e514. [Google Scholar] [CrossRef]
- Galani, I.; Karaiskos, I.; Angelidis, E.; Papoutsaki, V.; Galani, L.; Souli, M.; Antoniadou, A.; Giamarellou, H. Emergence of ceftazidime-avibactam resistance through distinct genomic adaptations in KPC-2-producing Klebsiella pneumoniae of sequence type 39 during treatment. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Gottig, S.; Frank, D.; Mungo, E.; Nolte, A.; Hogardt, M.; Besier, S.; Wichelhaus, T.A. Emergence of ceftazidime/avibactam resistance in KPC-3-producing Klebsiella pneumoniae in vivo. J. Antimicrob. Chemother. 2019, 74, 3211–3216. [Google Scholar] [CrossRef] [PubMed]
- Haidar, G.; Clancy, C.J.; Shields, R.K.; Hao, B.; Cheng, S.; Nguyen, M.H. Mutations in blaKPC-3 that confer ceftazidime-avibactam resistance encode novel KPC-3 variants that function as extended-spectrum β-lactamases. Antimicrob. Agents Chemother. 2017, 61, e02534-16. [Google Scholar] [CrossRef] [PubMed]
- Hobson, C.A.; Cointe, A.; Jacquier, H.; Choudhury, A.; Magnan, M.; Courroux, C.; Tenaillon, O.; Bonacorsi, S.; Birgy, A. Cross-resistance to cefiderocol and ceftazidime-avibactam in KPC beta-lactamase mutants and the inoculum effect. Clin. Microbiol. Infect. 2021, 27, 1172.e7. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Sun, B.; Huang, Y.; Liu, C.; Wang, Y.; Ren, Y.; Zhang, Y.; Wang, Y.; Mu, D. Diversity of ceftazidime-avibactam resistance mechanism in KPC2-producing Klebsiella pneumoniae under antibiotic selection pressure. Infect. Drug Resist. 2022, 15, 4627–4636. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, K.; Dong, H.; Ren, D.; Gong, D.; Jiang, F.; Shi, C.; Li, J.; Zhang, Q.; Yan, W.; et al. Ceftazidime-avibactam resistance in Klebsiella pneumoniae wequence type 11 due to a mutation in plasmid-borne blaKPC-2 to blaKPC-33, in Henan, China. Infect. Drug Resist. 2021, 14, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Deng, J.; Feng, Y.; Zhang, W.; Wu, S.; Liu, Y.; Che, H.; Xie, Y. Emergence of ceftazidime-avibactam resistance due to a novel blaKPC-2 mutation during treatment of carbapenem-resistant Klebsiella pneumoniae infections. J. Infect. Public Health 2022, 15, 545–549. [Google Scholar] [CrossRef]
- Mendes, G.; Ramalho, J.F.; Bruschy-Fonseca, A.; Lito, L.; Duarte, A.; Melo-Cristino, J.; Caneiras, C. First eescription of ceftazidime/avibactam resistance in a ST13 KPC-70-producing Klebsiella pneumoniae strain from Portugal. Antibiotics 2022, 11, 167. [Google Scholar] [CrossRef]
- Moreira, N.K.; Caierao, J. Ceftazidime-avibactam: Are we safe from class A carbapenemase producers’ infections? Folia. Microbiol. 2021, 66, 879–896. [Google Scholar] [CrossRef]
- Nichols, W.W.; Bradford, P.A.; Stone, G.G. The primary pharmacology of ceftazidime/avibactam: Microbiology from clinical studies, and development of resistance during treatment. J. Antimicrob. Chemother. 2023, 78, 871–892. [Google Scholar] [CrossRef] [PubMed]
- Nichols, W.W.; Lahiri, S.D.; Bradford, P.A.; Stone, G.G. The primary pharmacology of ceftazidime/avibactam: Resistance in vitro. J. Antimicrob. Chemother. 2023, 78, 569–585. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Mack, A.R.; Taracila, M.A.; Bonomo, R.A. Resistance to novel beta-lactam-beta-lactamase inhibitor combinations: The “price of progress”. Infect. Dis. Clin. N. Am. 2020, 34, 773–819. [Google Scholar] [CrossRef]
- Shi, Q.; Yin, D.; Han, R.; Guo, Y.; Zheng, Y.; Wu, S.; Yang, Y.; Li, S.; Zhang, R.; Hu, F. Emergence and recovery of ceftazidime-avibactam resistance in blaKPC-33-harboring Klebsiella pneumoniae sequence type 11 Isolates in China. Clin. Infect. Dis. 2020, 71, S436–S439. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Chen, L.; Cheng, S.; Chavda, K.D.; Press, E.G.; Snyder, A.; Pandey, R.; Doi, Y.; Kreiswirth, B.N.; Nguyen, M.H.; et al. Emergence of ceftazidime-avibactam resistance due to plasmid-borne blaKPC-3 mutations during treatment of carbapenem-resistant Klebsiella pneumoniae infections. Antimicrob. Agents Chemother. 2017, 61, e02097-16. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Chen, L.; Kreiswirth, B.N.; Clancy, C.J. In vitro selection of meropenem resistance among ceftazidime-avibactam-resistant, meropenem-susceptible Klebsiella pneumoniae isolates with variant KPC-3 carbapenemases. Antimicrob. Agents Chemother. 2017, 61, e00079-17. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Potoski, B.A.; Haidar, G.; Hao, B.; Doi, Y.; Chen, L.; Press, E.G.; Kreiswirth, B.N.; Clancy, C.J.; Nguyen, M.H. Clinical outcomes, drug toxicity, and emergence of ceftazidime-avibactam resistance among patients treated for carbapenem-resistant Enterobacteriaceae infections. Clin. Infect. Dis. 2016, 63, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Tiseo, G.; Falcone, M.; Leonildi, A.; Giordano, C.; Barnini, S.; Arcari, G.; Carattoli, A.; Menichetti, F. Meropenem-vaborbactam as salvage therapy for ceftazidime-avibactam-, cefiderocol-resistant ST-512 Klebsiella pneumoniae-producing KPC-31, a D179Y variant of KPC-3. Open Forum. Infect. Dis. 2021, 8, ofab141. [Google Scholar] [CrossRef]
- Venditti, C.; Nisii, C.; D’Arezzo, S.; Vulcano, A.; Capone, A.; Antonini, M.; Ippolito, G.; Di Caro, A. Molecular and phenotypical characterization of two cases of antibiotic-driven ceftazidime-avibactam resistance in blaKPC-3-harboring Klebsiella pneumoniae. Infect. Drug Resist. 2019, 12, 1935–1940. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, J.; Liu, Z.; Sun, A.; Sun, L.; Li, B.; Lu, B.; Liu, Y.; Cao, B. In vivo selection of imipenem resistance among ceftazidime-avibactam-resistant, imipenem-susceptible Klebsiella pneumoniae isolate with KPC-33 carbapenemase. Front. Microbiol. 2021, 12, 727946. [Google Scholar] [CrossRef]
- Wang, L.; Shen, W.; Zhang, R.; Cai, J. Identification of a novel ceftazidime-avibactam-resistant KPC-2 Variant, KPC-123, in Citrobacter koseri following ceftazidime-avibactam treatment. Front. Microbiol. 2022, 13, 930777. [Google Scholar] [CrossRef]
- Wilson, W.R.; Kline, E.G.; Jones, C.E.; Morder, K.T.; Mettus, R.T.; Doi, Y.; Nguyen, M.H.; Clancy, C.J.; Shields, R.K. Effects of KPC variant and porin genotype on the in vitro activity of meropenem-vaborbactam against carbapenem-resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2019, 63, e02048-18. [Google Scholar] [CrossRef]
- Zhang, P.; Shi, Q.; Hu, H.; Hong, B.; Wu, X.; Du, X.; Akova, M.; Yu, Y. Emergence of ceftazidime/avibactam resistance in carbapenem-resistant Klebsiella pneumoniae in China. Clin. Microbiol. Infect. 2020, 26, 124.e1. [Google Scholar] [CrossRef]
- Barnes, M.D.; Winkler, M.L.; Taracila, M.A.; Page, M.G.; Desarbre, E.; Kreiswirth, B.N.; Shields, R.K.; Nguyen, M.H.; Clancy, C.; Spellberg, B.; et al. Klebsiella pneumoniae carbapenemase-2 (KPC-2), substitutions at Ambler position Asp179, and resistance to ceftazidime-avibactam: Unique antibiotic-resistant phenotypes emerge from β-lactamase protein engineering. mBio 2017, 8, e00528-17. [Google Scholar] [CrossRef]
- Alsenani, T.A.; Viviani, S.L.; Kumar, V.; Taracila, M.A.; Bethel, C.R.; Barnes, M.D.; Papp-Wallace, K.M.; Shields, R.K.; Nguyen, M.H.; Clancy, C.J.; et al. Structural characterization of the D179N and D179Y variants of KPC-2 beta-lactamase: Omega-loop destabilization as a mechanism of resistance to ceftazidime-avibactam. Antimicrob. Agents Chemother. 2022, 66, e0241421. [Google Scholar] [CrossRef]
- Taracila, M.A.; Bethel, C.R.; Hujer, A.M.; Papp-Wallace, K.M.; Barnes, M.D.; Rutter, J.D.; VanPelt, J.; Shurina, B.A.; van den Akker, F.; Clancy, C.J.; et al. Different conformations revealed by NMR underlie resistance to ceftazidime/avibactam and susceptibility to meropenem and imipenem among D179Y variants of KPC beta-lactamase. Antimicrob. Agents Chemother. 2022, 66, e0212421. [Google Scholar] [CrossRef]
- Winkler, M.L.; Papp-Wallace, K.M.; Bonomo, R.A. Activity of ceftazidime/avibactam against isogenic strains of Escherichia coli containing KPC and SHV β-lactamases with single amino acid substitutions in the omega-loop. J. Antimicrob. Chemother. 2015, 70, 2279–2286. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Taracila, M.; Wallace, C.J.; Hujer, K.M.; Bethel, C.R.; Hornick, J.M.; Bonomo, R.A. Elucidating the role of Trp105 in the KPC-2 β-lactamase. Protein Sci. 2010, 19, 1714–1727. [Google Scholar] [CrossRef]
- Systèmes, D. BIOVIA, Dassault Systèmes, [Discovery Studio 2020 Client]; Dassault Systèmes: San Diego, CA, USA, 2020. [Google Scholar]
- Papp-Wallace, K.M.; Taracila, M.A.; Smith, K.M.; Xu, Y.; Bonomo, R.A. Understanding the molecular determinants of substrate and inhibitor specificities in the carbapenemase KPC-2: Exploring the roles of Arg220 and Glu276. Antimicrob. Agents Chemother. 2012, 56, 4428–4438. [Google Scholar] [CrossRef]
- Maveyraud, L.; Mourey, L.; Kotra, L.P.; Pedelacq, J.D.; Guillet, V.; Mobashery, S.; Samama, J.P. Structural basis for clinical longevity of carbapenem antibiotics in the face of challenge by the common class A β-lactamases from the antibiotic-resistant bacteria. J. Am. Chem. Soc. 1998, 120, 9748–9752. [Google Scholar] [CrossRef]
- Nukaga, M.; Bethel, C.R.; Thomson, J.M.; Hujer, A.M.; Distler, A.; Anderson, V.E.; Knox, J.R.; Bonomo, R.A. Inhibition of class A beta-lactamases by carbapenems: Crystallographic observation of two conformations of meropenem in SHV-1. J. Am. Chem. Soc. 2008, 130, 12656–12662. [Google Scholar] [CrossRef] [PubMed]
- Yigit, H.; Queenan, A.M.; Rasheed, J.K.; Biddle, J.W.; Domenech-Sanchez, A.; Alberti, S.; Bush, K.; Tenover, F.C. Carbapenem-resistant strain of Klebsiella oxytoca harboring carbapenem-hydrolyzing beta-lactamase KPC-2. Antimicrob. Agents Chemother. 2003, 47, 3881–3889. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Thirty-Secondth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the Expasy Server; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Papp-Wallace, K.M.; Winkler, M.L.; Gatta, J.A.; Taracila, M.A.; Chilakala, S.; Xu, Y.; Johnson, J.K.; Bonomo, R.A. Reclaiming the efficacy of β-lactam-β-lactamase inhibitor combinations: Avibactam restores the susceptibility of CMY-2-producing Escherichia coli to ceftazidime. Antimicrob. Agents Chemother. 2014, 58, 4290–4297. [Google Scholar] [CrossRef] [PubMed]
- Levitt, P.S.; Papp-Wallace, K.M.; Taracila, M.A.; Hujer, A.M.; Winkler, M.L.; Smith, K.M.; Xu, Y.; Harris, M.E.; Bonomo, R.A. Exploring the role of a conserved class A residue in the omega-loop of KPC-2 β-lactamase: A mechanism for ceftazidime hydrolysis. J. Biol. Chem. 2012, 287, 31783–31793. [Google Scholar] [CrossRef] [PubMed]
- Hobson, C.A.; Pierrat, G.; Tenaillon, O.; Bonacorsi, S.; Bercot, B.; Jaouen, E.; Jacquier, H.; Birgy, A. Klebsiella pneumoniae carbapenemase variants resistant to ceftazidime-avibactam: An evolutionary overview. Antimicrob. Agents Chemother. 2022, 66, e0044722. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).