A Novel Method That Allows SNP Discrimination with 160:1 Ratio for Biosensors Based on DNA-DNA Hybridization



{kind=link}
{kind=link}
{kind=link}
Abstract
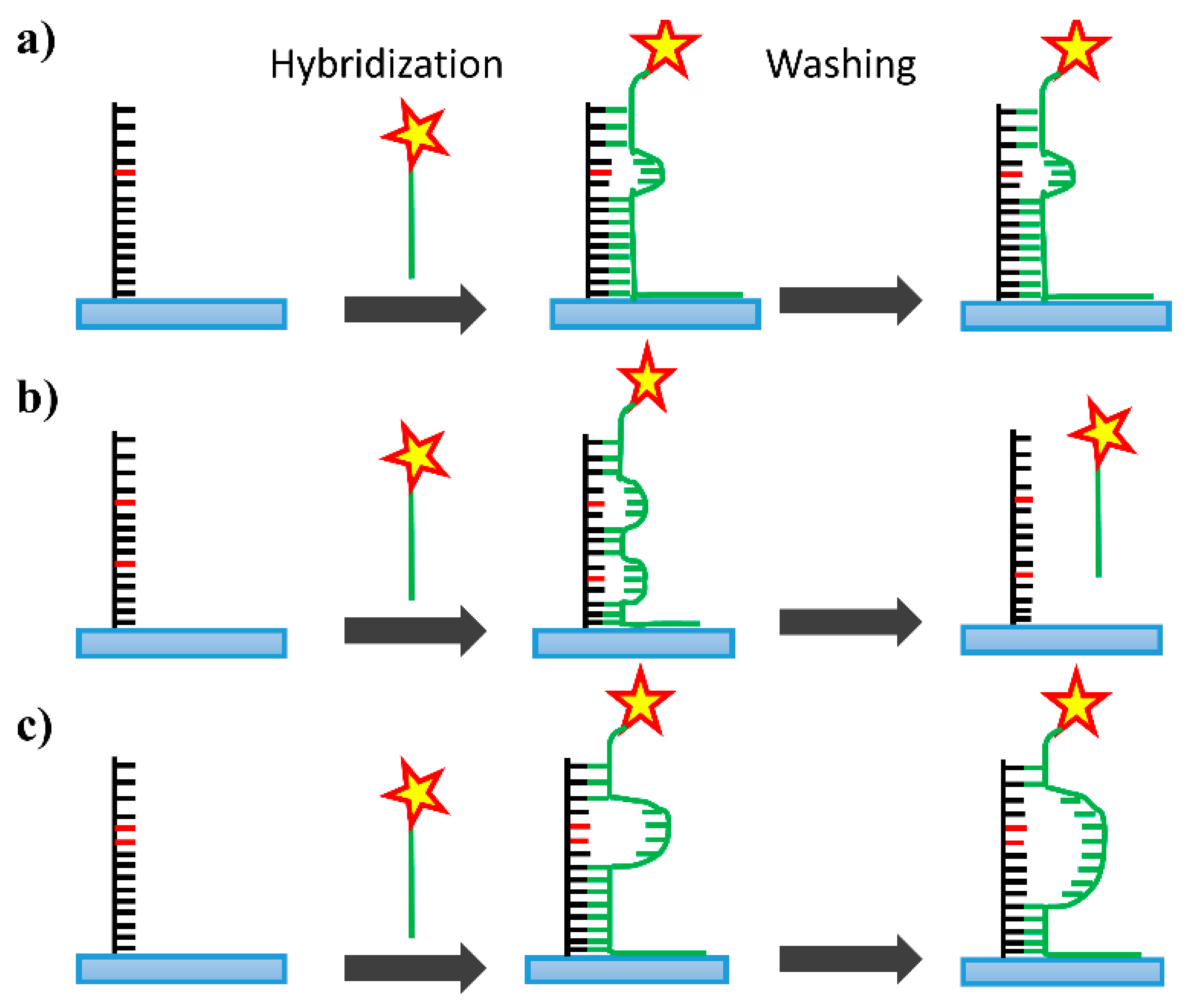
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Composition of Different Solutions
2.3. Typical Method for the Preparation of TB-NTM 9G DNAChip
2.4. Typical Hybridization and Washing Method
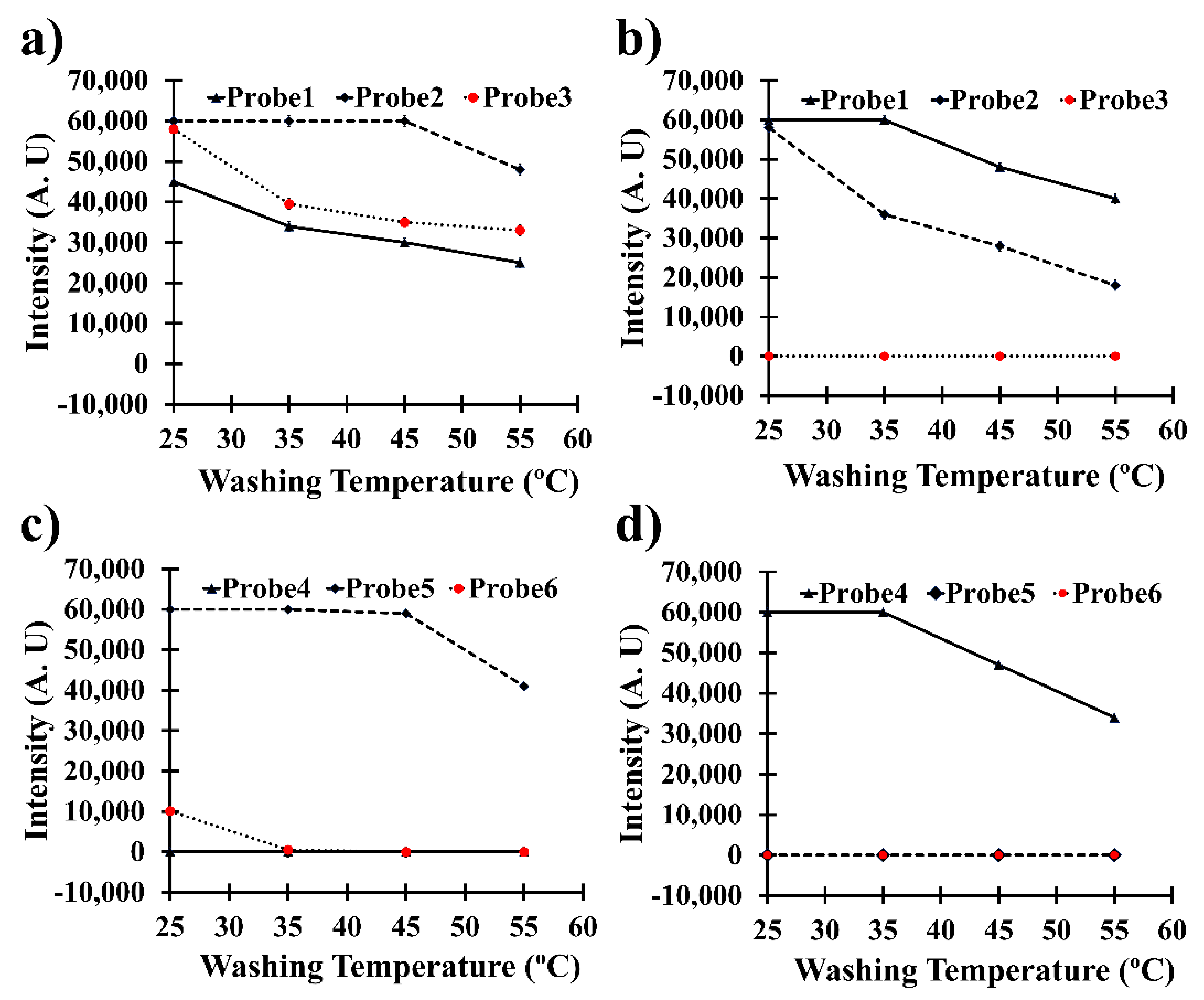
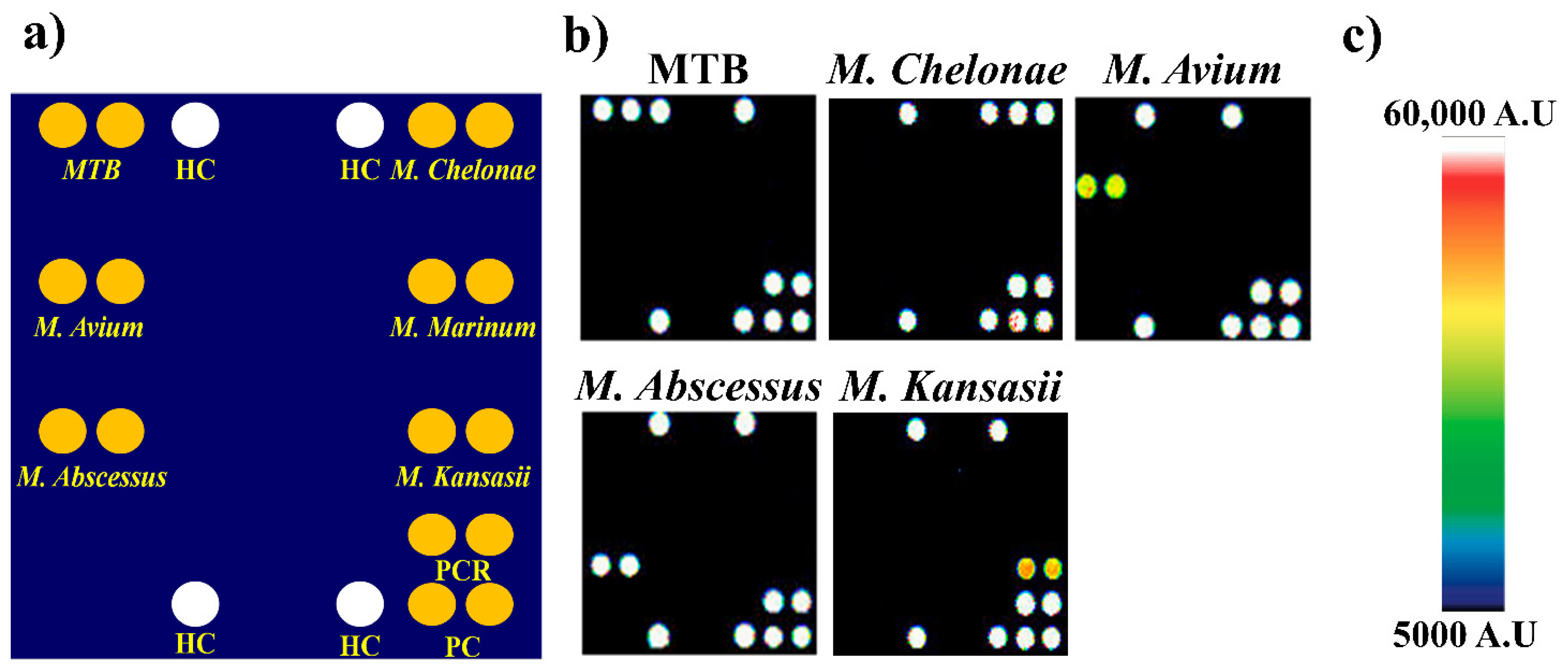
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hwang, S.M.; Lim, M.S.; Hong, Y.J.; Kim, T.S.; Park, K.U.; Song, J.; Lee, J.H.; Kim, E.C. Simultaneous detection of Mycobacterium tuberculosis complex and nontuberculous mycobacteria in respiratory specimens. Tuberculosis 2013, 93, 642–646. [Google Scholar] [CrossRef]
- Yasmin, R.; Zhu, H.; Chen, Z.; Montagna, R.A. A modifiable microarray-based universal sensor: Providing sample-to-results automation. Heliyon 2016, 2, e00179. [Google Scholar] [CrossRef][Green Version]
- Tang, C.; He, Z.; Liu, H.; Xu, Y.; Huang, H.; Yang, G.; Xiao, Z.; Li, S.; Liu, H.; Deng, Y.; et al. Application of magnetic nanoparticles in nucleic acid detection. J. Nanobiotechnol. 2020, 18, 62. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Song, K.; Sonawane, M.D.; Sayyed, D.R.; Kim, T. Immobilization techniques for microarray: Challenges and applications. Sensors 2014, 14, 22208–22229. [Google Scholar] [CrossRef]
- He, Z.; Zhou, J. Empirical evaluation of a new method for calculating signal-to-noise ratio for microarray data analysis. Appl. Environ. Microbiol. 2008, 74, 2957–2966. [Google Scholar] [CrossRef] [PubMed]
- Comtet-Marre, S.; Chaucheyras-Durand, F.; Bouzid, O.; Mosoni, P.; Bayat, A.R.; Peyret, P.; Forano, E. FibroChip, a Functional DNA Microarray to Monitor Cellulolytic and Hemicellulolytic Activities of Rumen Microbiota. Front. Microbiol. 2018, 9, 215. [Google Scholar] [CrossRef]
- Baaj, Y.; Magdelaine, C.; Ubertelli, V.; Valat, C.; Mousseau, Y.; Qiu, H.; Funalot, B.; Vallat, J.M.; Sturtz, F.G. Multiplex detection and genotyping of point mutations involved in charcot-marie-tooth disease using a hairpin microarray-based assay. Res. Lett. Biochem. 2009, 2009, 960560. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Tai, S.M.; Lee, E.C.; Mak, T.S.; Liu, T.K.; Tang, V.W.; Poon, W.T. Rapid identification of pathogenic variants in two cases of charcot-marie-tooth disease by gene-panel sequencing. Int. J. Mol. Sci. 2017, 18, 770. [Google Scholar] [CrossRef]
- Choi, Y.-D.; Jung, W.-W.; Nam, J.-H.; Choi, H.-S.; Park, C.-S. Detection of HPV genotypes in cervical lesions by the HPV DNA Chip and sequencing. Gynecol. Oncol. 2005, 98, 369–375. [Google Scholar] [CrossRef]
- Nimse, S.B.; Song, K.S.; Kim, J.; Sayyed, D.R.; Kim, T. 9G DNAChip Technology: Self-Assembled Monolayer (SAM) of ssDNA for ultra-sensitive detection of biomarkers. Int. J. Mol. Sci. 2013, 14, 5723–5733. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.J.; Sunkara, V.; Park, J.W. DNA microarrays on nanoscale-controlled surface. Nucleic Acids Res. 2005, 33, e106. [Google Scholar] [CrossRef]
- Liu, H.; Li, S.; Liu, L.; Tian, L.; He, N. Multiplex single nucleotide polymorphisms genotyping using solid-phase single base extension on magnetic nanoparticles. Anal. Biochem. 2009, 386, 126–128. [Google Scholar] [CrossRef]
- Yang, W.; Lai, R.Y. Effect of diluent chain length on the performance of the electrochemical DNA sensor at elevated temperature. Analyst 2011, 136, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Zimenkov, D.V.; Kulagina, E.V.; Antonova, O.V.; Krasnova, M.A.; Chernyaeva, E.N.; Zhuravlev, V.Y.; Kuz’min, A.V.; Popov, S.A.; Zasedatelev, A.S.; Gryadunov, D.A. Evaluation of a low-density hydrogel microarray technique for mycobacterial species identification. J. Clin. Microbiol. 2015, 53, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Pozhitkov, A.E.; Boube, I.; Brouwer, M.H.; Noble, P.A. Beyond Affymetrix arrays: Expanding the set of known hybridization isotherms and observing pre-wash signal intensities. Nucleic Acids Res. 2010, 38, e28. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.K.; Kim, J.; Mathies, R.A. Microfluidic hydrogel arrays for direct genotyping of clinical samples. Biosens. Bioelectron. 2016, 79, 371–378. [Google Scholar] [CrossRef]
- Baeissa, A.; Dave, N.; Smith, B.D.; Liu, J. DNA-functionalized monolithic hydrogels and gold nanoparticles for colorimetric DNA detection. ACS Appl. Mater. Interfaces 2010, 2, 3594–3600. [Google Scholar] [CrossRef]
- Kajiyama, T.; Miyahara, Y.; Kricka, L.J.; Wilding, P.; Graves, D.J.; Surrey, S.; Fortina, P. Genotyping on a thermal gradient DNA chip. Genome Res. 2003, 13, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Pozhitkov, A.E.; Stedtfeld, R.D.; Hashsham, S.A.; Noble, P.A. Revision of the nonequilibrium thermal dissociation and stringent washing approaches for identification of mixed nucleic acid targets by microarrays. Nucleic Acids Res. 2007, 35, e70. [Google Scholar] [CrossRef] [PubMed]
- Vasiliskov, V.A.; Chudinov, A.V.; Chechetkin, V.R.; Surzhikov, S.A.; Zasedatelev, A.S.; Mikhailovich, V.M. Separate production of single-stranded DNA is not necessary: Circuit denaturation of double-stranded DNA followed by hybridization of single strands on oligonucleotide microchips. J. Biomol. Struct. Dyn. 2009, 27, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Gryadunov, D.; Nicot, F.; Dubois, M.; Mikhailovich, V.; Zasedatelev, A.; Izopet, J. Hepatitis C virus genotyping using an oligonucleotide microarray based on the NS5B sequence. J. Clin. Microbiol. 2010, 48, 3910–3917. [Google Scholar] [CrossRef][Green Version]
- Abbey, D.; Hickman, M.; Gresham, D.; Berman, J. High-Resolution SNP/CGH microarrays reveal the accumulation of loss of heterozygosity in commonly used candida albicans strains. G3 2011, 1, 523–530. [Google Scholar] [CrossRef]
- Divne, A.-M.; Allen, M. A DNA microarray system for forensic SNP analysis. Forensic Sci. Int. 2005, 154, 111–121. [Google Scholar] [CrossRef]
- McGrath, E.E.; McCabe, J.; Anderson, P.B. Guidelines on the diagnosis and treatment of pulmonary non-tuberculous mycobacteria infection. Int. J. Clin. Pract. 2008, 62, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Jeon, B.Y.; Shim, T.S.; Jin, H.; Cho, S.N.; Lee, H. Development of a highly sensitive one-tube nested real-time PCR for detecting Mycobacterium tuberculosis. Diagn. Microbiol. Infect. Dis. 2014, 80, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.H.; Shan, J.Y. Nanomechanical behaviors of microcantilever-based single-stranded DNA chips induced by counterion osmotic effects. Biomech. Modeling Mechanobiol. 2011, 10, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Kim, J.; Ta, V.-T.; Kim, H.-S.; Song, K.-S.; Jung, C.-Y.; Nguyen, V.-T.; Kim, T. New water-soluble iminecalix [4] arene with a deep hydrophobic cavity. Tetrahedron Lett. 2009, 50, 7346–7350. [Google Scholar] [CrossRef]
- Nimse, S.B.; Kim, T. Biological applications of functionalized calixarenes. Chem. Soc. Rev. 2013, 42, 366–386. [Google Scholar] [CrossRef]
- Song, K.S.; Nimse, S.B.; Kim, J.; Kim, J.; Nguyen, V.T.; Ta, V.T.; Kim, T. 9G DNAChip: Microarray based on the multiple interactions of 9 consecutive guanines. Chem. Commun. 2011, 47, 7101–7103. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Song, K.S.; Kim, J.; Ta, V.T.; Nguyen, V.T.; Kim, T. A generalized probe selection method for DNA chips. Chem. Commun. 2011, 47, 12444–12446. [Google Scholar] [CrossRef]
- Melting Temperature (Tm) Calculations. Available online: http://www.basic.northwestern.edu/biotools/oligocalc.html (accessed on 25 November 2020).
- Sugimoto, N.; Nakano, S.; Yoneyama, M.; Honda, K. Improved thermodynamic parameters and helix initiation factor to predict stability of DNA duplexes. Nucleic Acids Res. 1996, 24, 4501–4505. [Google Scholar] [CrossRef]
- Wallace, R.B.; Shaffer, J.; Murphy, R.F.; Bonner, J.; Hirose, T.; Itakura, K. Hybridization of synthetic oligodeoxyribonucleotides to phi chi 174 DNA: The effect of single base pair mismatch. Nucleic Acids Res. 1979, 6, 3543–3557. [Google Scholar] [CrossRef] [PubMed]
- Michikawa, Y.; Suga, T.; Ohtsuka, Y.; Matsumoto, I.; Ishikawa, A.; Ishikawa, K.; Iwakawa, M.; Imai, T. Visible genotype sensor array. Sensors 2008, 8, 2722–2735. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.M.; Abendroth, J.M.; Nakatsuka, N.; Zhu, B.; Yang, Y.; Andrews, A.M.; Weiss, P.S. Detecting DNA and RNA and Differentiating Single-Nucleotide Variations via Field-Effect Transistors. Nano Lett. 2020, 20, 5982–5990. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nimse, S.B.; Song, K.-S.; Warkad, S.D.; Kim, T. A Novel Method That Allows SNP Discrimination with 160:1 Ratio for Biosensors Based on DNA-DNA Hybridization. Biosensors 2021, 11, 265. https://doi.org/10.3390/bios11080265
Nimse SB, Song K-S, Warkad SD, Kim T. A Novel Method That Allows SNP Discrimination with 160:1 Ratio for Biosensors Based on DNA-DNA Hybridization. Biosensors. 2021; 11(8):265. https://doi.org/10.3390/bios11080265
Chicago/Turabian StyleNimse, Satish Balasaheb, Keum-Soo Song, Shrikant Dashrath Warkad, and Taisun Kim. 2021. "A Novel Method That Allows SNP Discrimination with 160:1 Ratio for Biosensors Based on DNA-DNA Hybridization" Biosensors 11, no. 8: 265. https://doi.org/10.3390/bios11080265
APA StyleNimse, S. B., Song, K.-S., Warkad, S. D., & Kim, T. (2021). A Novel Method That Allows SNP Discrimination with 160:1 Ratio for Biosensors Based on DNA-DNA Hybridization. Biosensors, 11(8), 265. https://doi.org/10.3390/bios11080265