Femtosecond Spectroscopy of Au Hot-Electron Injection into TiO2: Evidence for Au/TiO2 Plasmon Photocatalysis by Bactericidal Au Ions and Related Phenomena

 , ,
, ,  , ,
, , 
 ,
, 
Abstract
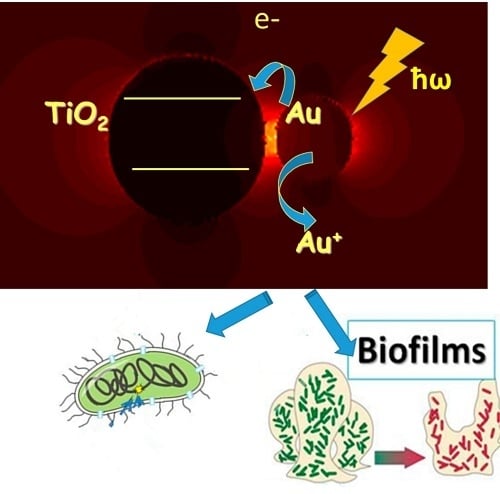
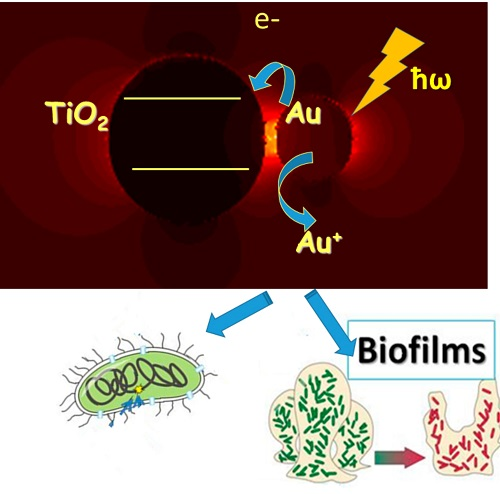
1. Introduction
2. Materials and Methods
2.1. Preparation of Au/TiO2 and Au Nanoparticles
2.2. Irradiation Procedures on the Au/TiO2, Electron Microscopy, and Laser Femtosecond Experimental Setup
2.2.1. Electron Microscopy
2.2.2. Femtosecond Laser Photolysis Setup
2.3. Preparation of Bacterial Strains, Evaluation of the Bacterial Inactivation Kinetics, and Bacterial Growth Conditions
2.3.1. Minimum Inhibitory Concentrations (MICs) and Biofilm Formation
2.3.2. Biofilm Formation
2.3.3. Effect of AuHCl4 on the E. coli AB1157 Biofilm and Effect of Au Ions on Bacterial Survival
2.3.4. Effect of Au Ions on E. coli AB1157 Cell Survival in Stationary and Exponential Growth Phases
2.3.5. Statistical Analysis
3. Results and Discussions
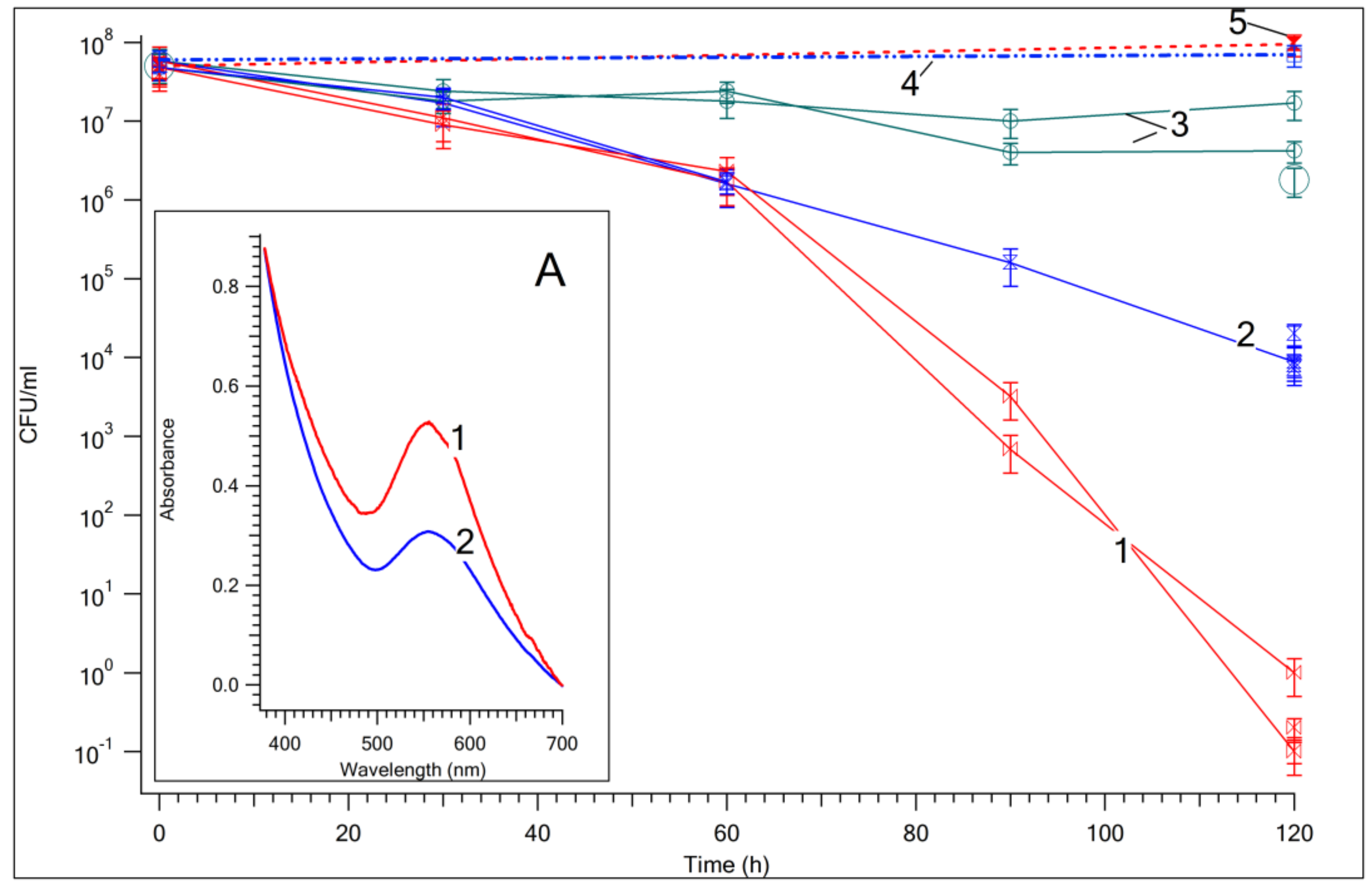
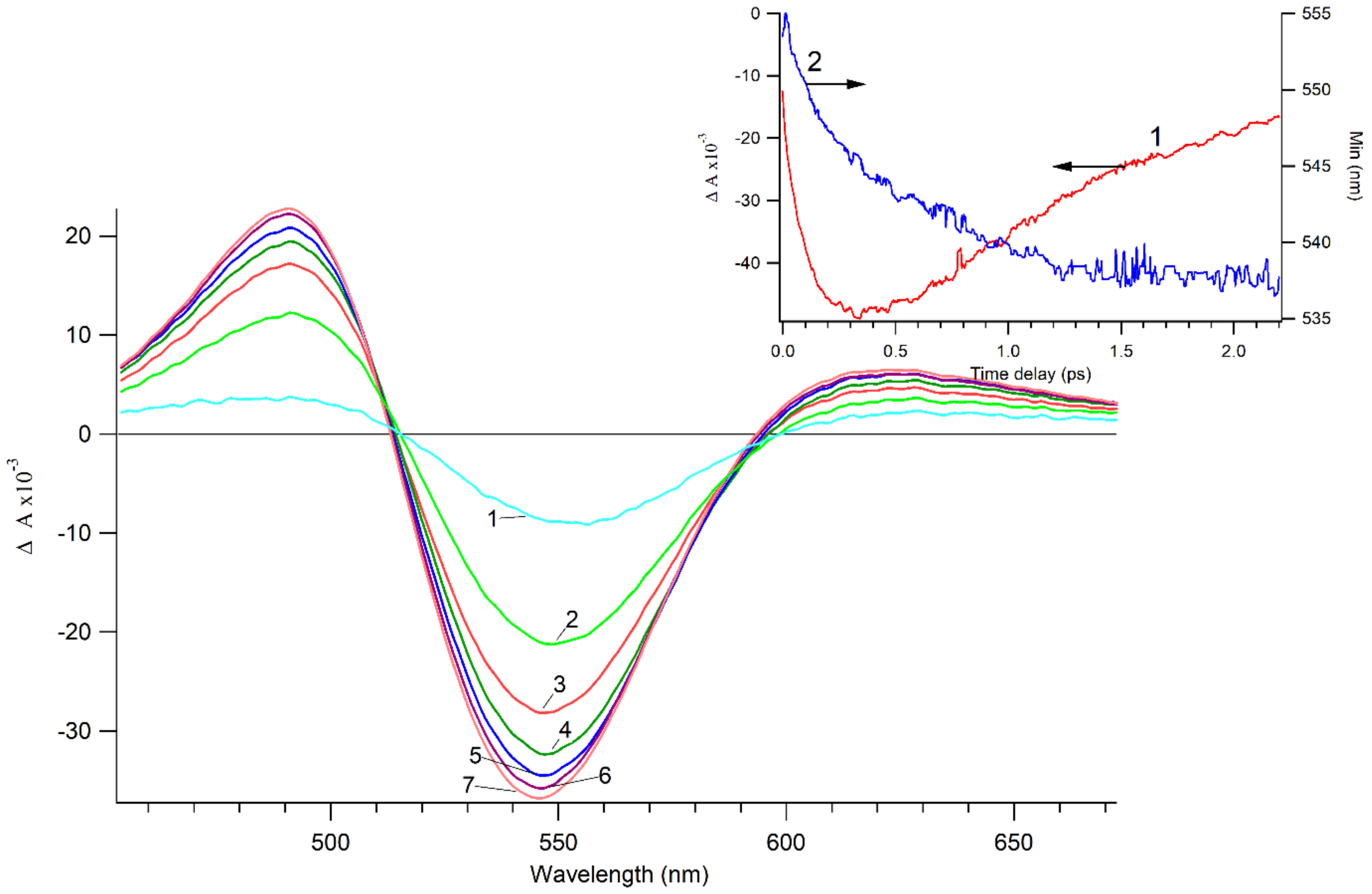
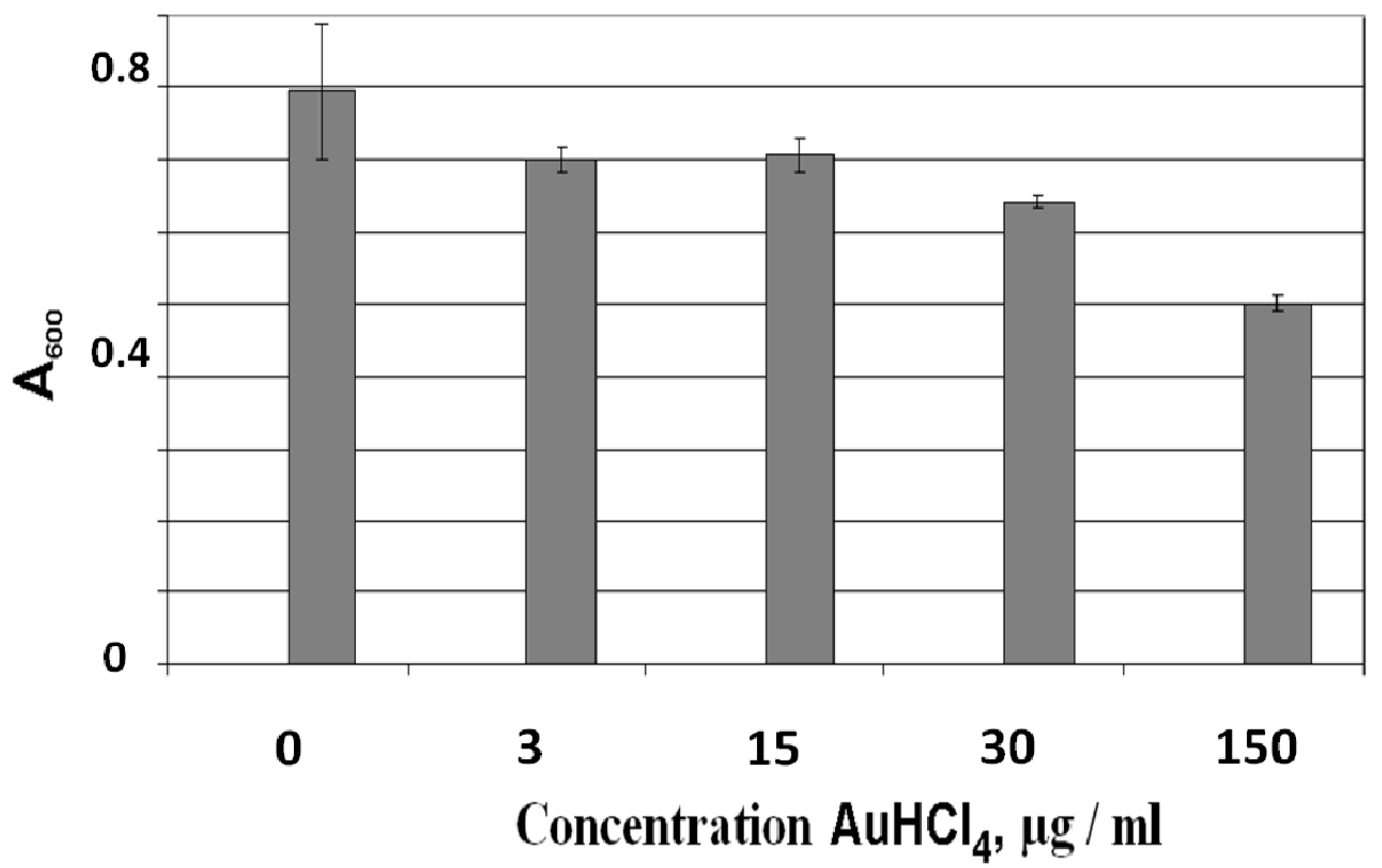
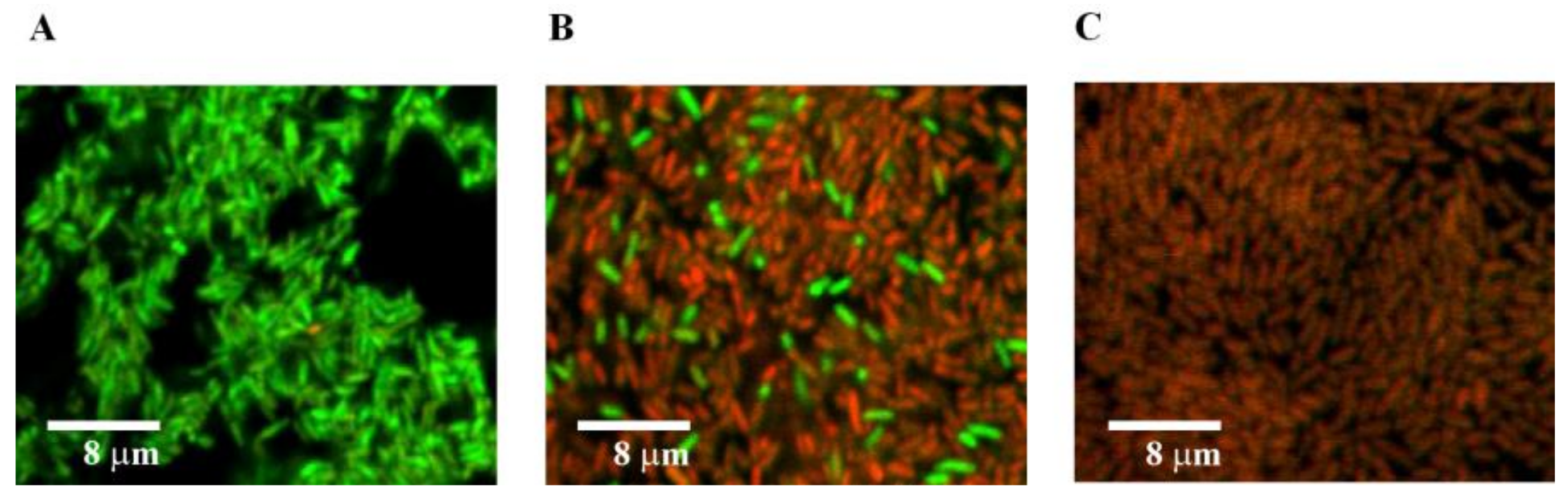
3.1. Photocatalytic Bactericidal Effect and Femtosecond Laser Spectroscopy of Au/TiO2 Suspensions
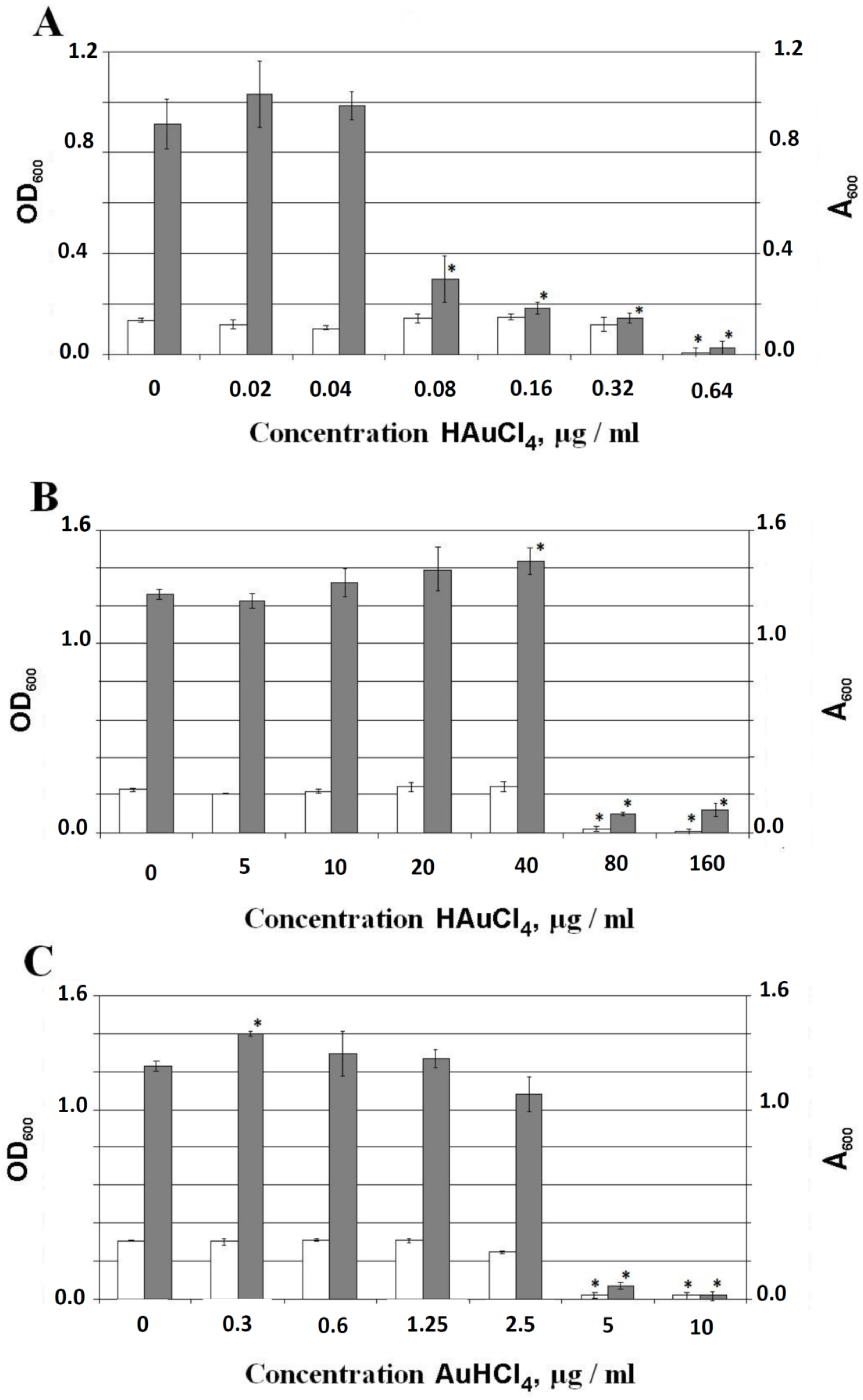
3.2. MICs of AuHCl4 for Different Gram-Type Bacteria and Effect of AuHCl4 on the Biofilm Formation
3.2.1. MICs of AuHCl4 of Different Gram-Type Bacteria
3.2.2. Effect of AuHCl4 on the Formation of Biofilms
3.2.3. Effect of AuHCl4 on Mature Biofilms of E. coli AB1157
3.3. The Effect of Phase of Growth on the Sensitivity of E. coli AB1157 Cells to AuHCl4
3.4. About the Effect of the Au Ions’ Interaction with Bacteria
3.5. The Effect of the Au Ions’ Interaction with Bacteria
3.6. Benefits of the Information Reported in This Study: Outlook and Future Work
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jalvo, B.; Faraldos, M. Antimicrobial and antibiofilm efficacy of self-cleaning surfaces functionalized by TiO2 photocatalytic nanoparticles against Staphylococcus aureus and Pseudomonas putida. J. Hazard. Mater. 2017, 340, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Yadav, H.M.; Kim, J.S. Developments in photocatalytic antibacterial activity of nano TiO2: A review. Korean J. Chem. Eng. 2016, 33, 1989–1998. [Google Scholar] [CrossRef]
- Fagan, R.; McCormack, D.E. A review of solar and visible light active TiO2 photocatalysis for treating bacteria, cyanotoxins and contaminants of emerging concern. Semicond. Process. 2016, 42, 2–14. [Google Scholar] [CrossRef]
- Wilke, C.M.; Gaillard, J.F. The critical role of light in moderating microbial stress due to mixtures of engineered nanomaterials. Environ. Sci. Nano 2018, 5, 96–102. [Google Scholar] [CrossRef]
- Armelao, L.; Barreca, D. Photocatalytic and antibacterial activity of TiO2 and Au/TiO2 nano-systems. Nanotechnology 2007, 18, 375709. [Google Scholar] [CrossRef]
- Pan, Y.; Leifert, A. Gold nanoparticles of diameter 1.4 nm trigger necrosis by oxidative stress and mitochondrial damage. Small 2009, 5, 2067–2076. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lawson, R. Toxic effects of gold nanoparticles on Salmonella typhimurium bacteria. Toxicol. Health 2011, 27, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Kong, Y. Antibacterial activities of gold and silver nanoparticles against Escherichia coli and bacillus Calmette-Guérin. J. Nanobiotechnol. 2012, 10, 19. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, Y. The molecular mechanism of action of bactericidal gold nanoparticles on Escherichia coli. Biomaterials 2012, 33, 2327–2333. [Google Scholar] [CrossRef]
- Bresee, J.; Maier, K.E. Growth inhibition of Staphylococcus aureus by mixed monolayer gold nanoparticles. Small 2011, 7, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.E.; Mwamuka, J. Gold nanoparticles are taken up by human cells but do not cause acute cytotoxicity. Small 2005, 1, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Rahme, K.; Chen, L. PEGylated gold nanoparticles: Polymer quantification as a function of PEG lengths and nanoparticle dimensions. RSC Adv. 2013, 3, 6085–6094. [Google Scholar] [CrossRef]
- Zhang, W.; Li, Y. Photogeneration of reactive oxygen species on uncoated silver, gold, nickel, and silicon nanoparticles and their antibacterial effects. Langmuir 2013, 29, 4647–4651. [Google Scholar] [CrossRef] [PubMed]
- Silvero, M.J.; Becerra, M.C. Plasmon-induced oxidative stress and macromolecular damage in pathogenic bacteria. RSC Adv. 2016, 6, 100203–100208. [Google Scholar] [CrossRef]
- Zharov, V.P.; Mercer, K.E. Photo-termal nano-therapeutics and nano-diagnostics for selective killing of bacteria targeted with gold nanoparticles. Biophys. J. 2006, 90, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Calixto, G.M.; Bernegossi, J.J. Nanotechnology-Based Drug Delivery Systems for Photodynamic Therapy of Cancer: A Review. Molecules 2016, 21, E342. [Google Scholar] [CrossRef] [PubMed]
- Ngwa, W.; Kumar, R. Targeted radiotherapy with gold nanoparticles: Current status and future perspectives. Nanomed. (Lond.) 2014, 9, 1063–1082. [Google Scholar] [CrossRef]
- Her, S.; Jaffray, D.A. Gold nanoparticles for applications in cancer radiotherapy: Mechanisms and recent advancements. Adv. Drug Deliv. Rev. 2015, S0169-409X, 30014. [Google Scholar] [CrossRef]
- Aiboushev, A.; Gostev, F. Spectral properties of the surface plasmon resonance and electron injection from gold nanoparticles to TiO2 mesoporous film: Femtosecond study. Photochem. Photobiol. Sci. 2013, 12, 631–637. [Google Scholar] [CrossRef]
- Cushing, S.K.; Chen, C.J. Tunable Nonthermal Distribution of Hot Electrons in a Semiconductor Injected from a Plasmonic Gold Nanostructure. ACS Nano 2018, 12, 7117–7126. [Google Scholar] [CrossRef]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Checa, S.K.; Soncini, F.C. Bacterial gold sensing and resistance. Biometals 2011, 24, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Benedek, T.G. The history of gold therapy for tuberculosis. J. Hist. Med. Allied Sci. 2004, 59, 50–89. [Google Scholar] [CrossRef] [PubMed]
- Milacic, V.; Fregona, D. Gold complexes as prospective metal-based anticancer drugs. Histol. Histopathol. 2008, 23, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.F., III. Gold-Based Therapeutic Agents. Chem. Rev. 1999, 99, 2589–2600. [Google Scholar] [CrossRef]
- Radzig, M.A.; Nadtochenko, V.A. Antibacterial effects of silver nanoparticles on Gram-negative bacteria: Influence on the growth and biofilms formation, mechanisms of action. Colloids Surf. B Biointerfaces 2013, 102, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Sau, T.K.; Murphy, C.J. Seeded high yield synthesis of short Au nanorods in aqueous solution. Langmuir 2004, 20, 6414–6420. [Google Scholar] [CrossRef]
- Turkevich, J.; Stevenson, P.C. A study of the nucleation and growth processes in the synthesis of colloidal gold. Discuss. Faraday Soc. 1951, 11, 55–75. [Google Scholar] [CrossRef]
- Nadtochenko, V.; Denisov, N. Ultrafast Spectroscopy of Fano-Like Resonance between Optical Phonon and Excitons in CdSe Quantum Dots: Dependence of Coherent Vibrational Wave-Packet Dynamics on Pump Fluence. Nanomaterials 2017, 7, 371. [Google Scholar] [CrossRef]
- Zaitseva, J.; Granik, V. Effect of nitrofurans and generators on biofilm formation by Pseudomonas aeruginosa PAO1 and Burkholderia Cenocepacia 370. Res. Microbiol. 2009, 160, 353–357. [Google Scholar] [CrossRef]
- Fujishima, A.; Zhang, X. TiO2 photocatalysis and related surface phenomena. Surf. Sci. Rep. 2008, 63, 515–582. [Google Scholar] [CrossRef]
- Tian, Y.; Tatsuma, T. Mechanisms and applications of plasmon-induced charge separation at TiO2 films loaded with gold nanoparticles. J. Am. Chem. Soc. 2005, 127, 7632–7637. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Zhang, Q.M. Escherichia coli Nth and human hNTH1 DNA glycosylases are involved in removal of 8-oxoguanine from 8-oxoguanine/guanine mispairs in DNA. Nucleic Acids Res. 2001, 29, 1975–1981. [Google Scholar] [CrossRef] [PubMed]
- Wyrzykowski, J.; Volkert, M.R. The Escherichia coli methyl-directed mismatch repair system repairs base pairs containing oxidative lesions. J. Bacteriol. 2003, 185, 1701–1704. [Google Scholar] [CrossRef] [PubMed]
- David, S.S.; O’Shea, V.L. Base-excision repair of oxidative DNA damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Hengge-Aronis, R. Signal transduction and regulatory mechanisms involved in control of the σS(rpoS) subunit of RNA Polymerase. Microbiol. Molec. Biol. Revs. 2002, 66, 373–395. [Google Scholar] [CrossRef]
- Nikaido, H. Porins and specific diffusion channels in bacterial outer membranes. J. Biol. Chem. 1994, 269, 3905–3908. [Google Scholar]
- Pugsley, A.P.; Schnaitman, C.A. Outer membrane proteins of Escherichia coli. VII. Evidence that bacteriophage-directed protein 2 functions as a pore. J. Bacteriol. 1978, 133, 1181–1189. [Google Scholar]
- Li, X.Z.; Nikaido, H. Silver-resistant mutants of Escherichia coli display active efflux of Ag+ and are deficient in porins. J. Bacteriol. 1997, 179, 6127–6132. [Google Scholar] [CrossRef]
- Teitzel, G.M.; Parsek, M.R. Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Kirketerp-Moller, K. Silver against Pseudomonas aeruginosa biofilms. APMIS 2007, 115, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Shah, D. Specialized persistent cells and the mechanism of multidrug tolerance in Escherichia coli. J. Bacteriol. 2004, 186, 8172–8180. [Google Scholar] [CrossRef] [PubMed]
- Summers, A.O. Damage control: Regulating defenses against toxic metals and metalloids. Curr. Opin. Microbiol. 2009, 12, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Calamai, P.; Carotti, S. Biological properties of two gold(III) complexes: AuCl3(Hpm) and AuCl2(pm). J. Inorg. Biochem. 1997, 66, 103–109. [Google Scholar] [CrossRef]
- Marino, T.; Russo, N. Theoretical investigation on DNA/RNA base pairs mediated by copper, silver, and gold cations. Dalton Trans. 2012, 41, 1816–1823. [Google Scholar] [CrossRef] [PubMed]
- Volesky, B.; Holan, Z.R. Biosorption of heavy metals. Biotechnol. Progress 1995, 11, 235–250. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC of AuHCl4 (μg/mL)* |
|---|---|
| Escherichia coli AB1157 | 1.0 ± 0.3 |
| Pseudomonas aeruginosa PAO1 | 80 ± 10 |
| Serratia proteamaculans 94 | 1.4 ± 0.3 |
| AuHCl4 (μg/mL) | Exponential Phase of Growth (cells/mL) | Stationary Phase of Growth (cells/mL) | ||
|---|---|---|---|---|
| 0 | 16·× 107 | 100% ** | 22·× 107 | 100% |
| 5 | 1.4·× 107 | 8.8% | 8.2·× 107 | 38% |
| 8.5 | 8·× 105 | 0.5% | 12.6·× 106 | 6% |
| 17 | 4.8·× 105 | 0.3% | 5·× 106 | 2.3% |
| Strains of Escherichia coli K12 | Characteristics | MIC of AuHCl4, μg/mL* |
|---|---|---|
| SBS 1936SBS 2680 | rpo S+ rpo S− | 1.3 ± 0.21.0 ± 0.3 |
| AB 1157AB 1886AB 2463AB 2494 | uvrA+uvrB+recA+lexA+recBC+recF+lon+ uvr A6 rec A13 lex A | 1.0 ± 0.30.5 ± 0.20.7 ± 0.21.0 ± 0.3 |
| MC 4100TK 821MH 1471MH 225 | ompR+ompF+ompC+ompR+ompF−ompC−ompR+ompF−ompC+ompR+ompF+ompC− | 1.0 ± 0.31.3 ± 0.31.0 ± 0.31.0 ± 0.3 |
| BW 25113JW 2703JW 2928JW 3610JW 0097 | rrnB3 ΔlacZ4787 hsdR514 Δ(araBAD)567Δ(rhaBAD)568 rph-1mutS::kanmutY::kanmutM::kanmutT::kan | 1.0 ± 0.30.5 ± 0.20.5 ± 0.21.3 ± 0.31.0 ± 0.3 |
| TK 603TK 610TK 612 | uvr A6umu C36umu C44 | 1.0 ± 0.31.0 ± 0.30.8 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radzig, M.; Koksharova, O.; Khmel, I.; Ivanov, V.; Yorov, K.; Kiwi, J.; Rtimi, S.; Tastekova, E.; Aybush, A.; Nadtochenko, V. Femtosecond Spectroscopy of Au Hot-Electron Injection into TiO2: Evidence for Au/TiO2 Plasmon Photocatalysis by Bactericidal Au Ions and Related Phenomena. Nanomaterials 2019, 9, 217. https://doi.org/10.3390/nano9020217
Radzig M, Koksharova O, Khmel I, Ivanov V, Yorov K, Kiwi J, Rtimi S, Tastekova E, Aybush A, Nadtochenko V. Femtosecond Spectroscopy of Au Hot-Electron Injection into TiO2: Evidence for Au/TiO2 Plasmon Photocatalysis by Bactericidal Au Ions and Related Phenomena. Nanomaterials. 2019; 9(2):217. https://doi.org/10.3390/nano9020217
Chicago/Turabian StyleRadzig, Marina, Olga Koksharova, Inessa Khmel, Vladimir Ivanov, Khursand Yorov, John Kiwi, Sami Rtimi, Elina Tastekova, Arseny Aybush, and Victor Nadtochenko. 2019. "Femtosecond Spectroscopy of Au Hot-Electron Injection into TiO2: Evidence for Au/TiO2 Plasmon Photocatalysis by Bactericidal Au Ions and Related Phenomena" Nanomaterials 9, no. 2: 217. https://doi.org/10.3390/nano9020217
APA StyleRadzig, M., Koksharova, O., Khmel, I., Ivanov, V., Yorov, K., Kiwi, J., Rtimi, S., Tastekova, E., Aybush, A., & Nadtochenko, V. (2019). Femtosecond Spectroscopy of Au Hot-Electron Injection into TiO2: Evidence for Au/TiO2 Plasmon Photocatalysis by Bactericidal Au Ions and Related Phenomena. Nanomaterials, 9(2), 217. https://doi.org/10.3390/nano9020217