
Carbon Nanotubes Substrates Alleviate Pro-Calcific Evolution in Porcine Valve Interstitial Cells

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. CNTs Synthesis
2.2. Isolation and Culture of Porcine Aortic Valve Interstitial Cells
2.3. Immunofluorescence Assay
2.4. pVICs Lysis to Expose the Surface-Interacting Plasma Membrane
2.5. Evaluation of α-SMA and Vinculin Expression
2.6. Cell Density, Degree of Circularity and Branching Assay
2.7. Sorting of pVICs Morphologies When Grown on CNTs Substrates
2.8. CNTs and Cell Characterization by Field Emission Scanning Electron Microscopy
2.9. Atomic Force Microscopy Imaging and Force Spectroscopy
2.10. Statistical Analysis
3. Results
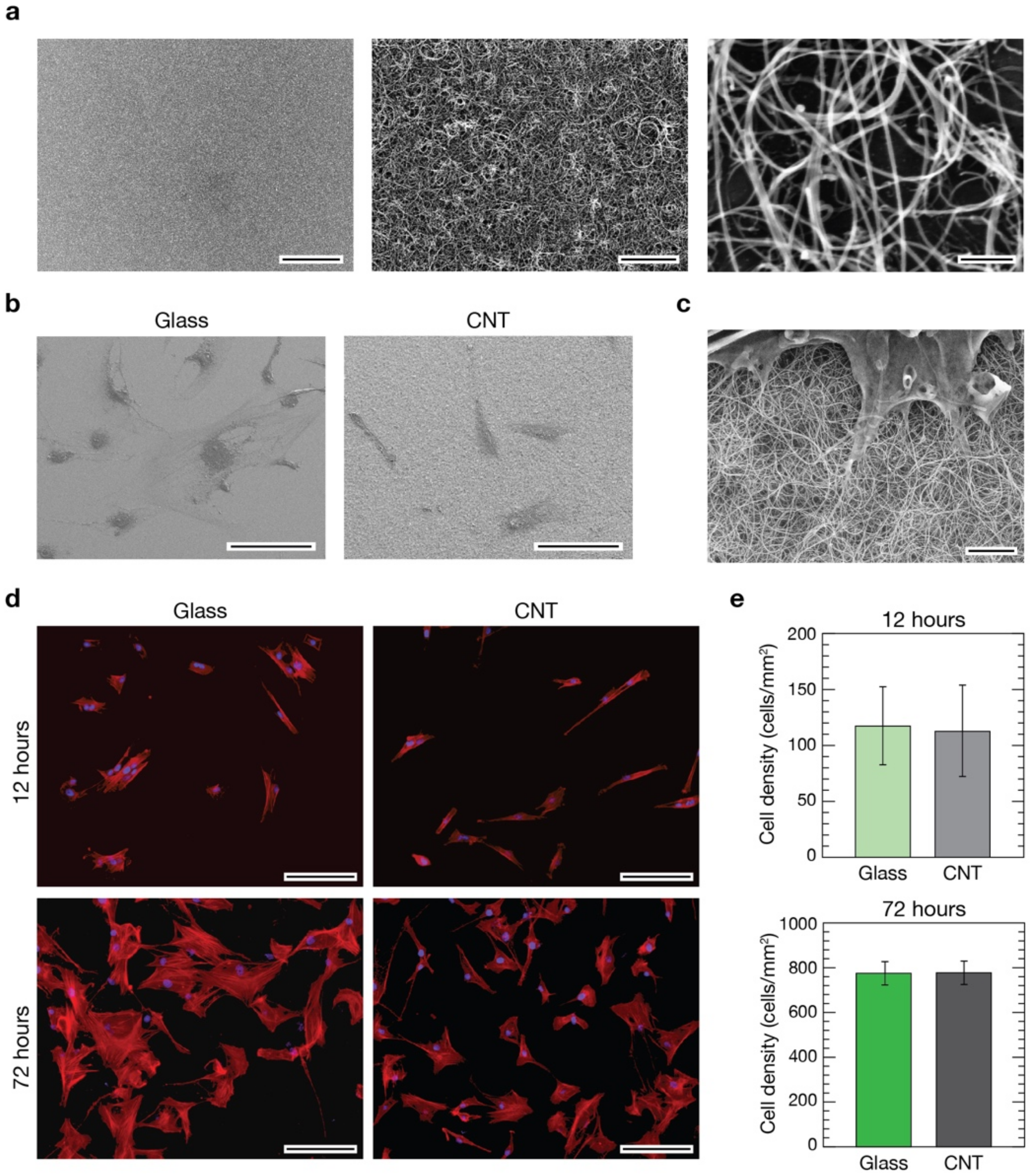
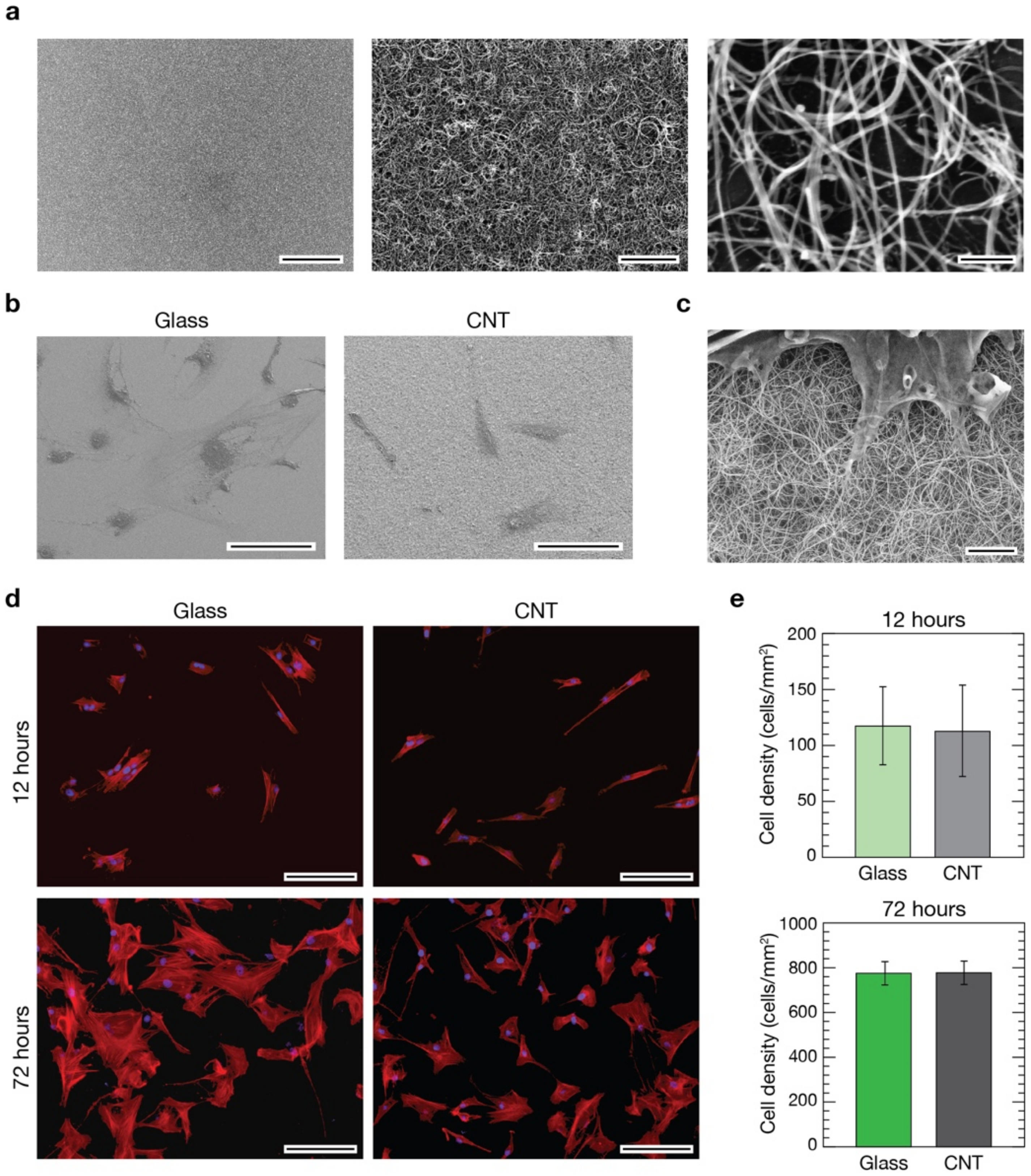
3.1. CNTs Fabrication and Characterization
3.2. Evaluation of pVICs Adhesion and Density on CNTs
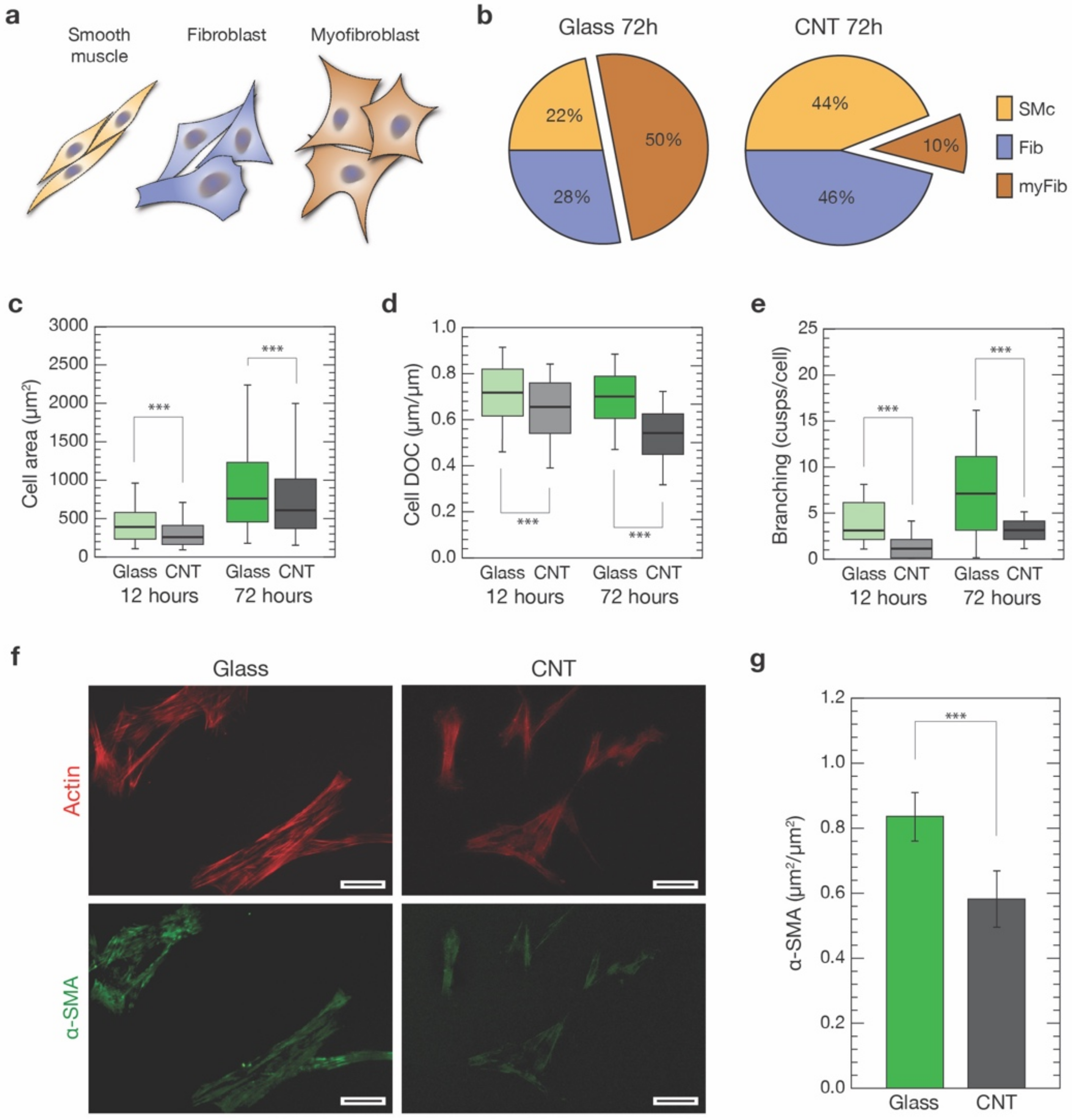
3.3. CNTs Influence on VIC Morphology
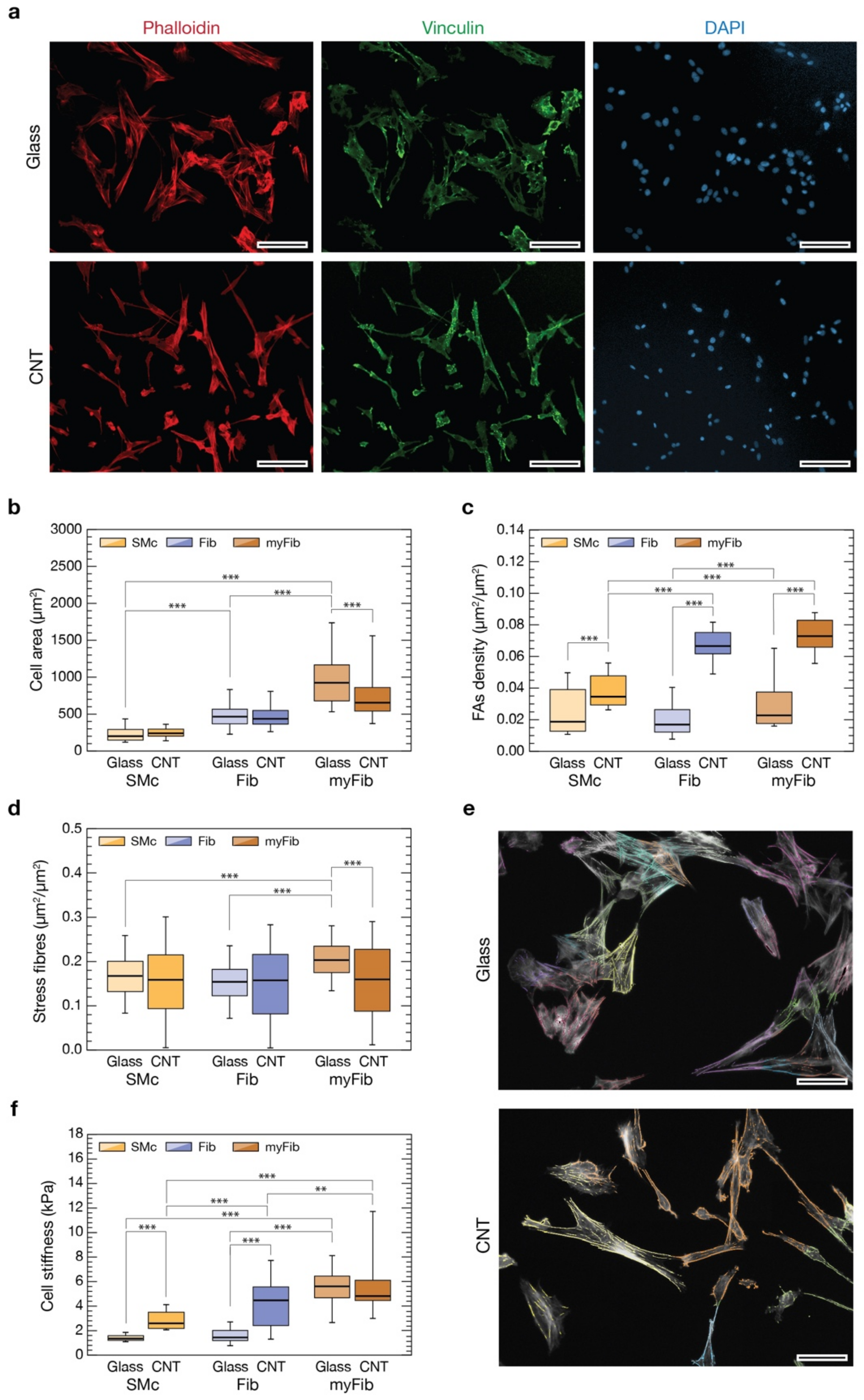
3.4. CNTs Affect Cytoskeletal Organization, Focal Adhesions and Stiffness of VICs
3.5. Cell Membrane Interaction with CNTs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eveborn, G.W.; Schirmer, H.; Heggelund, G.; Lunde, P.; Rasmussen, K. The Evolving Epidemiology of Valvular Aortic Stenosis. The Tromsø Study. Heart 2013, 99, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Rocca, F.; Sartore, S.; Guidolin, D.; Bertiplaglia, B.; Gerosa, G.; Casarotto, D.; Pauletto, P. Cell Composition of the Human Pulmonary Valve: A Comparative Study with the Aortic Valve–the VESALIO∗ Project∗∗Vitalitate Exornatum Succedaneum Aorticum Labore Ingegnoso Obtinebitur. Ann. Thorac. Surg. 2000, 70, 1594–1600. [Google Scholar] [CrossRef]
- Latif, N.; Sarathchandra, P.; Chester, A.H.; Yacoub, M.H. Expression of Smooth Muscle Cell Markers and Co-Activators in Calcified Aortic Valves. Eur. Heart J. 2015, 36, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Porras, A.M.; Van Engeland, N.C.; Marchbanks, E.; McCormack, A.; Bouten, C.V.; Yacoub, M.H.; Latif, N.; Masters, K.S. Robust Generation of Quiescent Porcine Valvular Interstitial Cell Cultures. J. Am. Heart Assoc. 2017, 14, e005041. [Google Scholar] [CrossRef]
- Pho, M.; Lee, W.; Watt, D.R.; Laschinger, C.; Simmons, C.A.; McCulloch, C.A. Cofilin is a marker of myofibroblast differentiation in cells from porcine aortic cardiac valves. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1767–H1778. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.M.; Allen, S.P.; Yacoub, M.H. Phenotypic and functional characterization of interstitial cells from human heart valves, pericardium and skin. J. Heart Valve Dis. 2000, 9, 150–158. [Google Scholar] [PubMed]
- Chen, J.-H.; Simmons, C.A.; Towler, D.A. Cell-Matrix Interactions in the Pathobiology of Calcific Aortic Valve Disease. Circ. Res. 2011, 108, 1510–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajamannan, N.M.; Subramaniam, M.; Rickard, D.; Stock, S.R.; Donovan, J.; Springett, M.; Orszulak, T.; Fullerton, D.A.; Tajik, A.J.; Bonow, R.O.; et al. Human Aortic Valve Calcification Is Associated with an Osteoblast Phenotype. Circulation 2003, 107, 2181–2184. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.L.K.; Simmons, C.A. Lessons from (Patho)Physiological Tissue Stiffness and Their Implications for Drug Screening, Drug Delivery and Regenerative Medicine. Adv. Drug Deliv. Rev. 2011, 63, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-H.; Chen, W.L.K.; Sider, K.L.; Yip, C.Y.Y.; Simmons, C.A. β-Catenin Mediates Mechanically Regulated, Transforming Growth Factor-Β1-Induced Myofibroblast Differentiation of Aortic Valve Interstitial Cells. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Sun, Y.; Simmons, C.A. Determination of Local and Global Elastic Moduli of Valve Interstitial Cells Cultured on Soft Substrates. J. Biomech. 2013, 46, 1967–1971. [Google Scholar] [CrossRef]
- Liu, F.; Mih, J.D.; Shea, B.S.; Kho, A.T.; Sharif, A.S.; Tager, A.M.; Tschumperlin, D.J. Feedback Amplification of Fibrosis through Matrix Stiffening and COX-2 Suppression. J. Cell Biol. 2010, 190, 693–706. [Google Scholar] [CrossRef] [Green Version]
- Wyss, K.; Yip, C.Y.Y.; Mirzaei, Z.; Jin, X.; Chen, J.-H.; Simmons, C.A. The Elastic Properties of Valve Interstitial Cells Undergoing Pathological Differentiation. J. Biomech. 2012, 45, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Merryman, W.D.; Youn, I.; Lukoff, H.D.; Krueger, P.M.; Guilak, F.; Hopkins, R.A.; Sacks, M.S. Correlation between Heart Valve Interstitial Cell Stiffness and Transvalvular Pressure: Implications for Collagen Biosynthesis. Am. J. Physiol.-Heart Circ. Physiol. 2006, 290, H224–H231. [Google Scholar] [CrossRef] [PubMed]
- Barbour, K.; Huang, H.-Y.S. Strain Effects on Collagen Proteolysis in Heart Valve Tissues. Mech. Time-Depend. Mater. 2020, 24, 85–100. [Google Scholar] [CrossRef]
- Gelain, F.; Bottai, D.; Vescovi, A.; Zhang, S. Designer Self-Assembling Peptide Nanofiber Scaffolds for Adult Mouse Neural Stem Cell 3-Dimensional Cultures. PLoS ONE 2006, 1, e119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.-H.; Li, C.-H.; Tang, M.-J.; Liang, J.-I.; Chen, C.-H.; Yeh, M.-L. Migration Speed and Directionality Switch of Normal Epithelial Cells after TGF-Β1-Induced EMT (TEMT) on Micro-Structured Polydimethylsiloxane (PDMS) Substrates with Variations in Stiffness and Topographic Patterning. Cell Commun. Adhes. 2013, 20, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasirci, V.; Kenar, H. Novel Surface Patterning Approaches for Tissue Engineering and Their Effect on Cell Behavior. Nanomedicine 2006, 1, 73–90. [Google Scholar] [CrossRef] [PubMed]
- George, J.H.; Shaffer, M.S.; Stevens, M.M. Investigating the Cellular Response to Nanofibrous Materials by Use of a Multi-Walled Carbon Nanotube Model. J. Exp. Nanosci. 2006, 1, 1–12. [Google Scholar] [CrossRef]
- Cellot, G.; Cilia, E.; Cipollone, S.; Rancic, V.; Sucapane, A.; Giordani, S.; Gambazzi, L.; Markram, H.; Grandolfo, M.; Scaini, D.; et al. Carbon Nanotubes Might Improve Neuronal Performance by Favouring Electrical Shortcuts. Nat. Nanotech. 2009, 4, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, V.; Cellot, G.; Toma, F.M.; Long, C.S.; Caldwell, J.H.; Zentilin, L.; Giacca, M.; Turco, A.; Prato, M.; Ballerini, L.; et al. Carbon Nanotubes Promote Growth and Spontaneous Electrical Activity in Cultured Cardiac Myocytes. Nano Lett. 2012, 12, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Aldinucci, A.; Turco, A.; Biagioli, T.; Toma, F.M.; Bani, D.; Guasti, D.; Manuelli, C.; Rizzetto, L.; Cavalieri, D.; Massacesi, L.; et al. Carbon nanotube scaffolds instruct human dendritic cells: Modulating immune responses by contacts at the nanoscale. Nano Lett. 2013, 13, 6098–6105. [Google Scholar] [CrossRef]
- Rago, I.; Rauti, R.; Bevilacqua, M.; Calaresu, I.; Pozzato, A.; Cibinel, M.; Dalmiglio, M.; Tavagnacco, C.; Goldoni, A.; Scaini, D. Carbon Nanotubes, Directly Grown on Supporting Surfaces, Improve Neuronal Activity in Hippocampal Neuronal Networks. Adv. Biosys. 2019, 3, 1800286. [Google Scholar] [CrossRef] [PubMed]
- Pampaloni, N.P.; Rago, I.; Calaresu, I.; Cozzarini, L.; Casalis, L.; Goldoni, A.; Ballerini, L.; Scaini, D. Transparent Carbon Nanotubes Promote the Outgrowth of Enthorino-dentate Projections in Lesioned Organ Slice Cultures. Develop. Neurobiol. 2020, 80, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Vardharajula, S.; Tiwari, P.; Erdal, E.; Komal, V.; Dennis, V. Ali Functionalized Carbon Nanotubes: Biomedical Applications. Int. J. Nanomed. 2012, 7, 5361. [Google Scholar] [CrossRef] [Green Version]
- Mallakpour, S.; Soltanian, S. Surface Functionalization of Carbon Nanotubes: Fabrication and Applications. RSC Adv. 2016, 6, 109916–109935. [Google Scholar] [CrossRef]
- Latif, N.; Quillon, A.; Sarathchandra, P.; McCormack, A.; Lozanoski, A.; Yacoub, M.H.; Chester, A.H. Modulation of Human Valve Interstitial Cell Phenotype and Function Using a Fibroblast Growth Factor 2 Formulation. PLoS ONE 2015, 10, e0127844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, R.; Consolo, F.; Spiccia, M.; Piola, M.; Kassem, S.; Prandi, F.; Vinci, M.C.; Forti, E.; Polvani, G.; Fiore, G.B.; et al. Feasibility of Pig and Human-Derived Aortic Valve Interstitial Cells Seeding on Fixative-Free Decellularized Animal Pericardium: Seeding of aortic valve interstitial cells in decellularized pericardium. J. Biomed. Mater. Res. 2016, 104, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, U.; Vinckier, A.; Kernen, P.; Zeisel, D.; Biber, J.; Semenza, G.; Murer, H.; Groscurth, P. Preparation of Basal Cell Membranes for Scanning Probe Microscopy. FEBS Lett. 1998, 436, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Curl, C.L.; Bellair, C.J.; Harris, P.J.; Allman, B.E.; Roberts, A.; Nugent, K.A.; Delbridge, L.M. Quantitative phase microscopy: A new tool for investigating the structure and function of unstained live cells. Clin. Exp. Pharmacol. Physiol. 2004, 31, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Chamley, J.H.; Gröschel-Stewart, U.; Campbell, G.R.; Burnstock, G. Distinction between Smooth Muscle, Fibroblasts and Endothelial Cells in Culture by the Use of Fluoresceinated Antibodies against Smooth Muscle Actin. Cell Tissue Res. 1977, 177, 445. [Google Scholar] [CrossRef] [PubMed]
- Hegner, B.; Schaub, T.; Catar, R.; Kusch, A.; Wagner, P.; Essin, K.; Lange, C.; Riemekasten, G.; Dragun, D. Intrinsic Deregulation of Vascular Smooth Muscle and Myofibroblast Differentiation in Mesenchymal Stromal Cells from Patients with Systemic Sclerosis. PLoS ONE 2016, 11, e0153101. [Google Scholar] [CrossRef]
- Schuster, R.; Rockel, J.S.; Kapoor, M.; Hinz, B. The Inflammatory Speech of Fibroblasts. Immunol. Rev. 2021, 302, 126–146. [Google Scholar] [CrossRef]
- Salvarani, N.; Maguy, A.; De Simone, S.A.; Miragoli, M.; Jousset, F.; Rohr, S. TGF-β1 (Transforming Growth Factor-β1) Plays a Pivotal Role in Cardiac Myofibroblast Arrhythmogenicity. Circ. Arrhythm. Electrophysiol. 2017, 10, e004567. [Google Scholar] [CrossRef]
- Thakar, R.G.; Cheng, Q.; Patel, S.; Chu, J.; Nasir, M.; Liepmann, D.; Komvopoulos, K.; Li, S. Cell-Shape Regulation of Smooth Muscle Cell Proliferation. Biophys. J. 2009, 96, 3423–3432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Urso, M.; Kurniawan, N.A. Mechanical and Physical Regulation of Fibroblast–Myofibroblast Transition: From Cellular Mechanoresponse to Tissue Pathology. Front. Bioeng. Biotechnol. 2020, 8, 609653. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Nečas, D.; Klapetek, P. Gwyddion: An Open-Source Software for SPM Data Analysis. Open Phys. 2012, 10, 181. [Google Scholar] [CrossRef]
- Lévy, R.; Maaloum, M. Measuring the Spring Constant of Atomic Force Microscope Cantilevers: Thermal Fluctuations and Other Methods. Nanotechnology 2002, 13, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Sneddon, I.N. The Relation between Load and Penetration in the Axisymmetric Boussinesq Problem for a Punch of Arbitrary Profile. Int. J. Eng. Sci. 1965, 3, 47–57. [Google Scholar] [CrossRef]
- Grimm, K.B.; Oberleithner, H.; Fels, J. Fixed Endothelial Cells Exhibit Physiologically Relevant Nanomechanics of the Cortical Actin Web. Nanotechnology 2014, 25, 215101. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Li, W.Z.; Xie, S.S.; Qian, L.X.; Chang, B.H.; Zou, B.S.; Zhou, W.Y.; Zhao, R.A.; Wang, G. Large-Scale Synthesis of Aligned Carbon Nanotubes. Science 1996, 274, 1701–1703. [Google Scholar] [CrossRef]
- Pampaloni, N.P.; Scaini, D.; Perissinotto, F.; Bosi, S.; Prato, M.; Ballerini, L. Sculpting Neurotransmission during Synaptic Development by 2D Nanostructured Interfaces. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 2521–2532. [Google Scholar] [CrossRef] [Green Version]
- Baum, J.; Duffy, H.S. Fibroblasts and Myofibroblasts: What Are We Talking About? J. Cardiovasc. Pharmacol. 2011, 57, 376–379. [Google Scholar] [CrossRef]
- Porter, K.E.; Turner, N.A. Cardiac Fibroblasts: At the Heart of Myocardial Remodeling. Pharmacol. Ther. 2009, 123, 255–278. [Google Scholar] [CrossRef]
- Phan, S.H. Biology of Fibroblasts and Myofibroblasts. Proc. Am. Thorac. Soc. 2008, 5, 334–337. [Google Scholar] [CrossRef]
- Tarbit, E.; Singh, I.; Peart, J.N.; Rose’Meyer, R.B. Biomarkers for the Identification of Cardiac Fibroblast and Myofibroblast Cells. Heart Fail. Rev. 2019, 24, 1–15. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, M.; Wu, G.; Yao, C.; Zhang, J. Recent Advances in Morphological Cell Image Analysis. Comput. Math. Methods Med. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutkovskiy, A.; Malashicheva, A.; Sullivan, G.; Bogdanova, M.; Kostareva, A.; Stensløkken, K.; Fiane, A.; Vaage, J. Valve Interstitial Cells: The Key to Understanding the Pathophysiology of Heart Valve Calcification. J. Am. Heart Assoc. 2017, 6, e006339. [Google Scholar] [CrossRef]
- Bogdanova, M.; Zabirnyk, A.; Malashicheva, A.; Enayati, K.Z.; Karlsen, T.A.; Kaljusto, M.-L.; Kvitting, J.-P.E.; Dissen, E.; Sullivan, G.J.; Kostareva, A.; et al. Interstitial Cells in Calcified Aortic Valves Have Reduced Differentiation Potential and Stem Cell-like Properties. Sci. Rep. 2019, 9, 12934. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Park, Y.-S.; Park, M.; Jang, M.J.; Kim, S.-M.; Lee, J.; Choi, J.Y.; Jung, D.H.; Chang, Y.-T.; Yoon, M.-H.; et al. Axon-First Neuritogenesis on Vertical Nanowires. Nano Lett. 2016, 16, 675–680. [Google Scholar] [CrossRef]
- Reynolds, A.J.; Chaponnier, C.; Jahoda, C.A.B.; Gabbiani, G. A Quantitative Study of the Differential Expression of Alpha-Smooth Muscle Actin in Cell Populations of Follicular and Non-Follicular Origin. J. Investig. Dermatol. 1993, 101, 577–583. [Google Scholar] [CrossRef]
- Pellegrin, S.; Mellor, H. Actin Stress Fibres. J. Cell Sci. 2007, 120, 3491–3499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.-J.; Dangerfield, A.L.; Rattan, S.G.; Bathe, K.L.; Cunnington, R.H.; Raizman, J.E.; Bedosky, K.M.; Freed, D.H.; Kardami, E.; Dixon, I.M.C. Cardiac Fibroblast to Myofibroblast Differentiation in Vivo and in Vitro: Expression of Focal Adhesion Components in Neonatal and Adult Rat Ventricular Myofibroblasts. Dev. Dyn. 2010, 239, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.T.; Horwitz, A.R.; Schwartz, M.A. Cell Adhesion: Integrating Cytoskeletal Dynamics and Cellular Tension. Nat. Rev. Mol. Cell Biol. 2010, 11, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Burridge, K.; Guilluy, C. Focal Adhesions, Stress Fibers and Mechanical Tension. Exp. Cell Res. 2016, 343, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, S.; Vieira, A.F.; Taubenberger, A.V.; Araújo, M.; Martins, N.P.; Brás-Pereira, C.; Polónia, A.; Herbig, M.; Barreto, C.; Otto, O.; et al. Actin Stress Fiber Organization Promotes Cell Stiffening and Proliferation of Pre-Invasive Breast Cancer Cells. Nat. Commun. 2017, 8, 15237. [Google Scholar] [CrossRef]
- Haase, K.; Pelling, A.E. Investigating Cell Mechanics with Atomic Force Microscopy. J. R. Soc. Interface 2015, 12, 20140970. [Google Scholar] [CrossRef] [PubMed]
- Usprech, J.; Chen, W.L.K.; Simmons, C.A. Heart Valve Regeneration: The Need for Systems Approaches: Systems Approaches for Heart Valve Regeneration. WIREs Syst. Biol. Med. 2016, 8, 169–182. [Google Scholar] [CrossRef]
- Santoro, R.; Scaini, D.; Severino, L.U.; Amadeo, F.; Ferrari, S.; Bernava, G.; Garoffolo, G.; Agrifoglio, M.; Casalis, L.; Pesce, M. Activation of Human Aortic Valve Interstitial Cells by Local Stiffness Involves YAP-Dependent Transcriptional Signaling. Biomaterials 2018, 181, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Pesce, M.; Santoro, R. Feeling the Right Force: How to Contextualize the Cell Mechanical Behavior in Physiologic Turnover and Pathologic Evolution of the Cardiovascular System. Pharmacol. Ther. 2017, 171, 75–82. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulloa, L.S.; Perissinotto, F.; Rago, I.; Goldoni, A.; Santoro, R.; Pesce, M.; Casalis, L.; Scaini, D. Carbon Nanotubes Substrates Alleviate Pro-Calcific Evolution in Porcine Valve Interstitial Cells. Nanomaterials 2021, 11, 2724. https://doi.org/10.3390/nano11102724
Ulloa LS, Perissinotto F, Rago I, Goldoni A, Santoro R, Pesce M, Casalis L, Scaini D. Carbon Nanotubes Substrates Alleviate Pro-Calcific Evolution in Porcine Valve Interstitial Cells. Nanomaterials. 2021; 11(10):2724. https://doi.org/10.3390/nano11102724
Chicago/Turabian StyleUlloa, Luisa Severino, Fabio Perissinotto, Ilaria Rago, Andrea Goldoni, Rosaria Santoro, Maurizio Pesce, Loredana Casalis, and Denis Scaini. 2021. "Carbon Nanotubes Substrates Alleviate Pro-Calcific Evolution in Porcine Valve Interstitial Cells" Nanomaterials 11, no. 10: 2724. https://doi.org/10.3390/nano11102724
APA StyleUlloa, L. S., Perissinotto, F., Rago, I., Goldoni, A., Santoro, R., Pesce, M., Casalis, L., & Scaini, D. (2021). Carbon Nanotubes Substrates Alleviate Pro-Calcific Evolution in Porcine Valve Interstitial Cells. Nanomaterials, 11(10), 2724. https://doi.org/10.3390/nano11102724