A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate Inhibits the Basal and Hsp90-Stimulated Migration and Invasion of Tumor Cells

Abstract
1. Introduction
2. Results
2.1. The 2,5-DHBA–Gelatin Conjugate Exhibited No Cytotoxic and Antiproliferative Activities Against Tumor Cell Lines
2.2. The 2,5-DHBA–Gelatin Conjugate Decreased the Basal Migration and Invasion of Cells In Vitro
2.3. The 2,5-DHBA–Gelatin Conjugate Detached Hsp90α And Hsp90β from the Cell Surface
2.4. The Migration and Invasion of Cells Stimulated by Exogenous Hsp90 Was Reduced by the 2,5-DHBA–Gelatin Conjugate
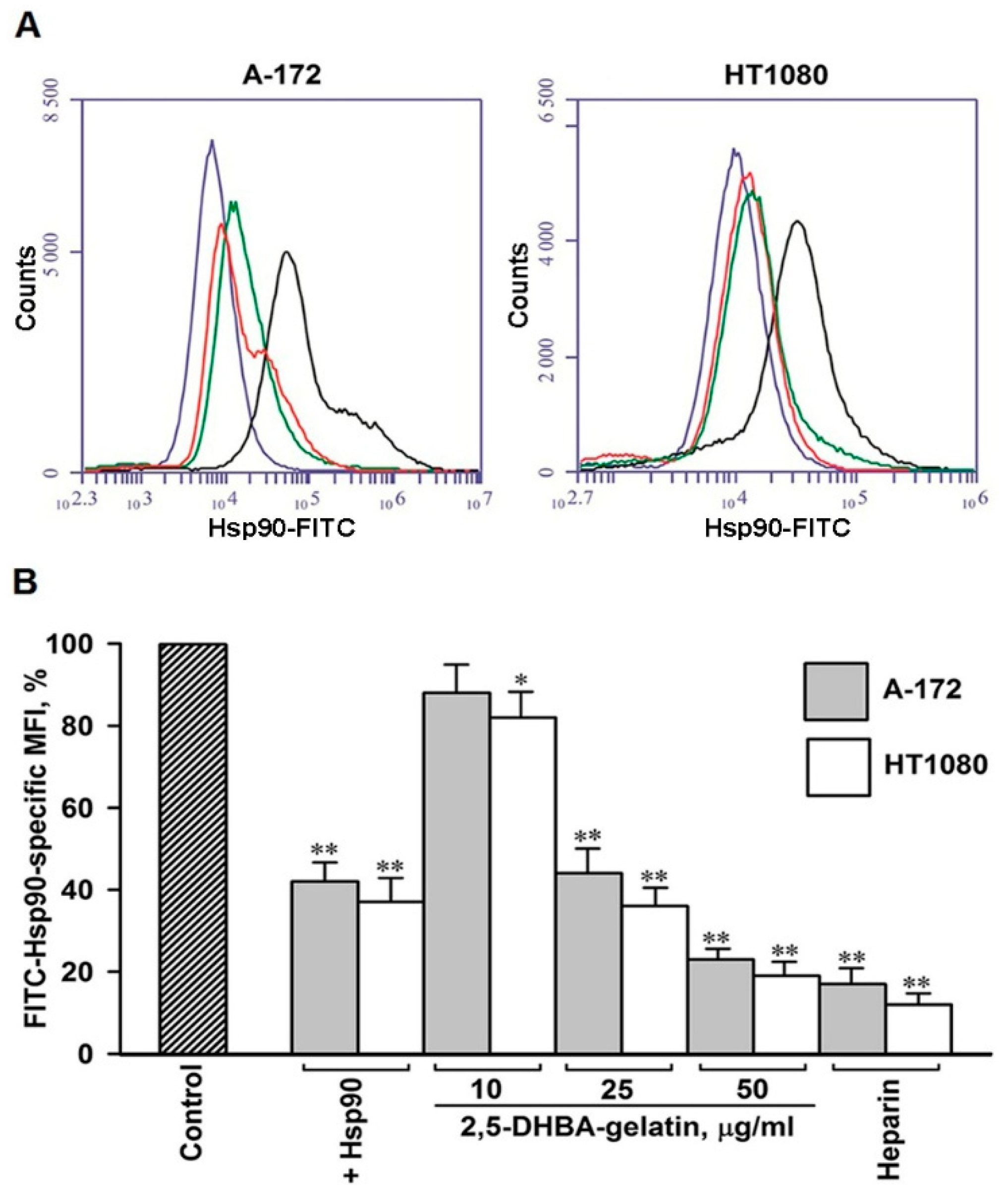
2.5. The 2,5-DHBA–Gelatin Conjugate Specifically Inhibited the Binding of Soluble Hsp90 to the Cell Surface
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cells and Virus
4.3. Preparation of the 2,5-DHBA–Gelatin Conjugate
4.4. Purification and Fluorescent Labeling of Hsp90
4.5. Determination of Cytotoxicity and Antiproliferative Activity of 2,5-DHBA–Gelatin
4.6. Evaluation of Basal Cell Migration and Invasion
4.7. Hsp90-Stimulated Migration and Invasion of Cells
4.8. Determination of the Level of Membrane-Associated Hsp90α and Hsp90β
4.9. Binding of Hsp90 to Cells in the Presence of the 2,5-DHBA–Gelatin Conjugate
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Wells, A.; Grahovac, J.; Wheeler, S.; Ma, B.; Lauffenburger, D. Targeting tumor cell motility as a strategy against invasion and metastasis. Trends Pharmacol. Sci. 2013, 34, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef]
- Gallagher, J. Fell-Muir lecture: Heparan sulphate and the art of cell regulation: A polymer chain conducts the protein orchestra. Int. J. Exp. Pathol. 2015, 96, 203–231. [Google Scholar] [CrossRef] [PubMed]
- Morla, S. Glycosaminoglycans and glycosaminoglycan mimetics in cancer and inflammation. Int. J. Mol. Sci. 2019, 20, 1963. [Google Scholar] [CrossRef]
- Zacharski, L.R. Heparin as an anticancer therapeutic. Expert Opin. Investig. Drugs 2008, 17, 1029–1037. [Google Scholar] [CrossRef]
- Borsig, L. Heparin as an inhibitor of cancer progression. Prog. Mol. Biol. Transl. Sci. 2010, 93, 335–349. [Google Scholar]
- Bochenek, J.; Püsküllüoglu, M.; Krzemieniecki, K. The antineoplastic effect of low-molecular-weight heparins-A literature review. Contemp. Oncol. 2013, 17, 6–13. [Google Scholar] [CrossRef]
- Zhou, H.; Roy, S.; Cochran, E.; Zouaoui, R.; Chu, C.L.; Duffner, J.; Zhao, G.; Smith, S.; Galcheva-Gargova, Z.; Karlgren, J.; et al. M402, a novel Heparan sulfate mimetic, targets multiple pathways implicated in tumor progression and metastasis. PLoS ONE 2011, 6, e21106. [Google Scholar] [CrossRef]
- Ritchie, J.P.; Ramani, V.C.; Ren, Y.; Naggi, A.; Torri, G.; Casu, B.; Penco, S.; Pisano, C.; Carminati, P.; Tortoreto, M.; et al. SST0001, a chemically modified heparin, inhibits myeloma growth and angiogenesis via disruption of the heparanase/syndecan-1 axis. Clin. Cancer Res. 2011, 17, 1382–1393. [Google Scholar] [CrossRef]
- Norrby, K.; Nordenhem, A. Dalteparin, a low-molecular-weight heparin, promotes angiogenesis mediated by heparin-binding VEGF-A in vivo. APMIS 2010, 118, 949–957. [Google Scholar] [CrossRef]
- Bereczky, B.; Gilly, R.; Rásó, E.; Vágó, A.; Tímár, J.; Tóvári, J. Selective antimetastatic effect of heparins in preclinical human melanoma models is based on inhibition of migration and microvascular arrest. Clin. Exp. Metastasis 2005, 22, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Smorenburg, S.M.; Van Noorden, C.J. The complex effects of heparins on cancer progression and metastasis in experimental studies. Pharmacol. Rev. 2001, 53, 93–105. [Google Scholar] [PubMed]
- Varki, N.M.; Varki, A. Heparin inhibition of selectin-mediated interactions during the hematogenous phase of carcinoma metastasis: Rationale for clinical studies in humans. Semin. Thromb. Hemost. 2002, 28, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.; Harvey, J.R.; Murphy, K.J.; Pye, D.; O’Boyle, G.; Lennard, T.W.; Kirby, J.A.; Ali, S. Modulatory effects of heparin and short-length oligosaccharides of heparin on the metastasis and growth of LMD MDA-MB 231 breast cancer cells in vivo. Br. J. Cancer 2007, 97, 761–768. [Google Scholar] [CrossRef]
- Stevenson, J.L.; Choi, S.H.; Varki, A. Differential metastasis inhibition by clinically relevant levels of heparins-correlation with selectin inhibition, not antithrombotic activity. Clin. Cancer Res. 2005, 11, 7003–7011. [Google Scholar] [CrossRef]
- Li, J.; Buchner, J. Structure, function and regulation of the hsp90 machinery. Biomed. J. 2013, 36, 106–117. [Google Scholar]
- Becker, B.; Multhoff, G.; Farkas, B.; Wild, P.J.; Landthaler, M.; Stolz, W.; Vogt, T. Induction of Hsp90 protein expression in malignant melanomas and melanoma metastases. Exp. Dermatol. 2004, 13, 27–32. [Google Scholar] [CrossRef]
- Cheng, C.; Li, W. Secretion of heat shock protein-90 (Hsp90) by normal cells under stress or by tumor cells during invasion: Why? Cancer Ther. 2008, 6, 765–772. [Google Scholar]
- Eustace, B.K.; Sakurai, T.; Stewart, J.K.; Yimlamai, D.; Unger, C.; Zehetmeier, C.; Lain, B.; Torella, C.; Henning, S.W.; Beste, G.; et al. Functional proteomic screens reveal an essential extracellular role for hsp90 alpha in cancer cell invasiveness. Nat. Cell Biol. 2004, 6, 507–514. [Google Scholar] [CrossRef]
- McCready, J.; Sims, J.D.; Chan, D.; Jay, D.G. Secretion of extracellular hsp90α via exosomes increases cancer cell motility: A role for plasminogen activation. BMC Cancer 2010, 10, 294. [Google Scholar] [CrossRef]
- Sidera, K.; Gaitanou, M.; Stellas, D.; Matsas, R.; Patsavoudi, E. A critical role for HSP90 in cancer cell invasion involves interaction with the extracellular domain of HER-2. J. Biol. Chem. 2008, 283, 2031–2041. [Google Scholar] [CrossRef] [PubMed]
- Stellas, D.; Karameris, A.; Patsavoudi, E. Monoclonal antibody 4C5 immunostains human melanomas and inhibits melanoma cell invasion and metastasis. Clin. Cancer Res. 2007, 13, 1831–1838. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Zhuo, W.; Fu, Y.; Shi, H.; Liang, Y.; Tong, M.; Chang, G.; Luo, Y. The regulatory mechanism of HSP90α secretion and its function in tumor malignancy. Proc. Natl. Acad. Sci. USA 2009, 106, 21288–21293. [Google Scholar] [CrossRef]
- Tsutsumi, S.; Scroggins, B.; Koga, F.; Lee, M.J.; Trepel, J.; Felts, S.; Carreras, C.; Neckers, L. A small molecule cell-impermeant Hsp90 antagonist inhibits tumor cell motility and invasion. Oncogene 2008, 27, 2478–2487. [Google Scholar] [CrossRef]
- McCready, J.; Wong, D.S.; Burlison, J.A.; Ying, W.; Jay, D.G. An impermeant ganetespib analog inhibits extracellular Hsp90-mediated cancer cell migration that involves lysyl oxidase 2-like protein. Cancers (Basel) 2014, 6, 1031–1046. [Google Scholar] [CrossRef] [PubMed]
- Sahu, D.; Zhao, Z.; Tsen, F.; Cheng, C.F.; Park, R.; Situ, A.J.; Dai, J.; Eginli, A.; Shams, S.; Chen, M.; et al. A potentially common peptide target in secreted heat shock protein-90α for hypoxia-inducible factor-1α-positive tumors. Mol. Biol. Cell 2012, 23, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.A.; Zohn, I.E. Hectd1 regulates intracellular localization and secretion of Hsp90 to control cellular behavior of the cranial mesenchyme. J. Cell Biol. 2012, 196, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wang, X.; Zhuo, W.; Shi, H.; Feng, D.; Sun, Y.; Liang, Y.; Fu, Y.; Zhou, D.; Luo, Y. The regulatory mechanism of extracellular Hsp90α on matrix metalloproteinase-2 processing and tumor angiogenesis. J. Biol. Chem. 2010, 285, 40039–40049. [Google Scholar] [CrossRef]
- Snigireva, A.V.; Vrublevskaya, V.; Afanasyev, N.; Morenkov, O. Cell surface heparan sulfate proteoglycans are involved in the binding of Hsp90α and Hsp90β to the cell plasma membrane. Cell Adhes. Migr. 2015, 9, 460–468. [Google Scholar] [CrossRef]
- Snigireva, A.V.; Vrublevskaya, V.V.; Skarga, Y.Y.; Morenkov, O.S. Cell surface heparan sulfate proteoglycans are involved in the extracellular Hsp90-stimulated migration and invasion of cancer cells. Cell Stress Chaperones 2019, 24, 309–322. [Google Scholar] [CrossRef]
- Lisov, A.; Vrublevskaya, V.; Lisova, Z.; Leontievsky, A.; Morenkov, O. A 2,5-dihydroxybenzoic acid-gelatin conjugate: The synthesis, antiviral activity and mechanism of antiviral action against two alphaherpesviruses. Viruses 2015, 7, 5343–5360. [Google Scholar] [CrossRef] [PubMed]
- Workman, P. Combinatorial attack on multistep oncogenesis by inhibiting the Hsp90 molecular chaperone. Cancer Lett. 2004, 206, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Hsu, Y.M.; Chen, C.C.; Chen, L.L.; Lee, C.C.; Huang, T.S. Secreted heat shock protein 90α induces colorectal cancer cell invasion through CD91/LRP-1 and NF-kappaB-mediated integrin αV expression. J. Biol. Chem. 2010, 285, 25458–25466. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.F.; Fan, J.; Fedesco, M.; Guan, S.; Li, Y.; Bandyopadhyay, B.; Bright, A.M.; Yerushalmi, D.; Liang, M.; Chen, M.; et al. Transforming growth factor alpha (TGFalpha)-stimulated secretion of HSP90alpha: Using the receptor LRP-1/CD91 to promote human skin cell migration against a TGFbeta-rich environment during wound healing. Mol. Cell Biol. 2008, 28, 3344–3358. [Google Scholar] [CrossRef] [PubMed]
- Gopal, U.; Bohonowych, J.E.; Lema-Tome, C.; Liu, A.; Garrett-Mayer, E.; Wang, B.; Isaacs, J.S. A novel extracellular Hsp90 mediated co-receptor function for LRP1 regulates EphA2 dependent glioblastoma cell invasion. PLoS ONE 2011, 6, e17649. [Google Scholar] [CrossRef]
- Hance, M.W.; Dole, K.; Gopal, U.; Bohonowych, J.E.; Jezierska-Drutel, A.; Neumann, C.A.; Liu, H.; Garraway, I.P.; Isaacs, J.S. Secreted Hsp90 is a novel regulator of the epithelial to mesenchymal transition (EMT) in prostate cancer. J. Biol. Chem. 2012, 287, 37732–37744. [Google Scholar] [CrossRef]
- Tsen, F.; Bhatia, A.; O’Brien, K.; Cheng, C.F.; Chen, M.; Hay, N.; Stiles, B.; Woodley, D.T.; Li, W. Extracellular heat shock protein 90 signals through subdomain II and the NPVY motif of LRP-1 receptor to Akt1 and Akt2: A circuit essential for promoting skin cell migration in vitro and wound healing in vivo. Mol. Cell Biol. 2013, 33, 4947–4959. [Google Scholar] [CrossRef]
- Thuringer, D.; Hammann, A.; Benikhlef, N.; Fourmaux, E.; Bouchot, A.; Wettstein, G.; Solary, E.; Garrido, C. Transactivation of the epidermal growth factor receptor by heat shock protein 90 via toll-like receptor 4 contributes to the migration of glioblastoma cells. J. Biol. Chem. 2011, 286, 3418–3428. [Google Scholar] [CrossRef]
- Li, W.; Li, Y.; Guan, S.; Fan, J.; Cheng, C.F.; Bright, A.M.; Chinn, C.; Chen, M.; Woodley, D.T. Extracellular heat shock protein-90alpha: Linking hypoxia to skin cell motility and wound healing. EMBO J. 2007, 26, 1221–1233. [Google Scholar] [CrossRef]
- Li, W.; Tsen, F.; Sahu, D.; Bhatia, A.; Chen, M.; Multhoff, G.; Woodley, D.T. Extracellular Hsp90 (eHsp90) as the actual target in clinical trials: Intentionally or unintentionally. Int. Rev. Cell Mol. Biol. 2013, 303, 203–235. [Google Scholar]
- Chen, W.S.; Chen, C.C.; Chen, L.L.; Lee, C.C.; Huang, T.S. Secreted heat shock protein 90α (Hsp90α) induces nuclear factor-κb-mediated TCF12 protein expression to down-regulate E-cadherin and to enhance colorectal cancer cell migration and invasion. J. Biol. Chem. 2013, 288, 9001–9010. [Google Scholar] [CrossRef] [PubMed]
- Puoci, F.; Morelli, C.; Cirillo, G.; Curcio, M.; Parisi, O.I.; Maris, P.; Sisci, D.; Picci, N. Anticancer activity of a quercetin-based polymer towards HeLa cancer cells. Anticancer Res. 2012, 32, 2843–2847. [Google Scholar]
- Seymour, L. Synthetic polymers with intrinsic anticancer activity. J. Bioact. Compat. Polym. 1991, 6, 178–216. [Google Scholar] [CrossRef]
- Hatice, K.C.; Dogan, A.L.; Zakir, M.O.; Uner, A.G.; Guner, A. Synthesis, characterization, and antitumor activity of poly(maleic anhydride-co-vinyl acetate-co-acrylic acid). J. Appl. Polym. Sci. 2006, 100, 3425–3432. [Google Scholar]
- Cirillo, G.; Kraemer, K.; Fuessel, S.; Puoci, F.; Curcio, M.; Spizzirri, U.G.; Altimari, I.; Iemma, F. Biological activity of a gallic acid-gelatin conjugate. Biomacromolecules 2010, 11, 3309–3315. [Google Scholar] [CrossRef] [PubMed]
- Andrijevic, L.; Radotic, K.; Bogdanovic, J.; Mutavdzic, D.; Bogdanovic, G. Antiproliferative effect of synthetic lignin against human breast cancer and normal fetal lung cell lines. Potency of low molecular weight fractions. J. BUON 2008, 13, 241–244. [Google Scholar] [PubMed]
- Ottenbrite, R.M. Antitumor activity of polycarboxylic acid polymers. J. Macromol. Sci. Chem. 1985, 22, 819–832. [Google Scholar] [CrossRef]
- Nikitovic, D.; Assouti, M.; Sifaki, M.; Katonis, P.; Krasagakis, K.; Karamanos, N.K.; Tzanakakis, G.N. Chondroitin sulfate and heparan sulfate-containing proteoglycans are both partners and targets of basic fibroblast growth factor-mediated proliferation in human metastatic melanoma cell lines. Int. J. Biochem. Cell Biol. 2008, 40, 72–83. [Google Scholar] [CrossRef]
- Fthenou, E.; Zafiropoulos, A.; Tsatsakis, A.; Stathopoulos, A.; Karamanos, N.K.; Tzanakakis, G.N. Chondroitin sulfate A chains enhance platelet derived growth factor-mediated signalling in fibrosarcoma cells. Int. J. Biochem. Cell Biol. 2006, 38, 2141–2150. [Google Scholar] [CrossRef]
- Kocak, F.E.; Erdoğan, E.; Ozyıgıt, F.; Yerlıkaya, A. Evaluation of antiproliferative and antimetastatic effects of heparin and erythropoietin on B16f10 melanoma cell line. Cell Mol. Biol. (Noisy Grand) 2013, 59, 1894–1898. [Google Scholar]
- Chatzinikolaou, G.; Nikitovic, D.; Berdiaki, A.; Zafiropoulos, A.; Katonis, P.; Karamanos, N.K.; Tzanakakis, G.N. Heparin regulates colon cancer cell growth through p38 mitogen-activated protein kinase signalling. Cell Prolif. 2012, 43, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, M.K.; Hoerster, J.C.; Bundschuh, D.; Boemicke, P.; Luedders, D.W.; Köster, F. Growth-modulatory effects of heparin and VEGF165 on the choriocarcinoma cell-line JEG-3 and its expression of heparanase. Anticancer Res. 2014, 34, 7071–7076. [Google Scholar] [PubMed]
- Friedl, P.; Gilmour, D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2009, 10, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Sidera, K.; Samiotaki, M.; Yfanti, E.; Panayotou, G.; Patsavoudi, E. Involvement of cell surface HSP90 in cell migration reveals a novel role in the developing nervous system. J. Biol. Chem. 2004, 279, 45379–45388. [Google Scholar] [CrossRef] [PubMed]
- Dreyfuss, J.L.; Regatieri, C.V.; Jarrouge, T.R.; Cavalheiro, R.P.; Sampaio, L.O.; Nader, H.B. Heparan sulfate proteoglycans: Structure, protein interactions and cell signaling. An. Acad. Bras. Cienc. 2009, 81, 409–429. [Google Scholar] [CrossRef]
- Lisova, Z.A.; Lisov, A.V.; Leontievsky, A.A. Two laccase isoforms of the basidiomycete Cerrena unicolor VKMF-3196. Induction, isolation and properties. J. Basic Microbiol. 2010, 50, 72–82. [Google Scholar] [CrossRef]
- Skarga, Y.; Vrublevskaya, V.; Evdokimovskaya, Y.; Morenkov, O. Purification of the 90 kDa heat shock protein (hsp90) and simultaneous purification of hsp70/hsc70, hsp90 and hsp96 from mammalian tissues and cells using thiophilic interaction chromatography. Biomed. Chromatogr. 2009, 23, 1208–1216. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell viability assays. In Assay Guidance Manual [Internet]; Sittampalam, G.S., Grossman, A., Brimacombe, K., Eds.; Eli Lilly & Company: Indianapolis, IN, USA; National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Concentration, µg/mL | Cytotoxicity (%) (a) | Antiproliferative Activity (%) (a) | ||
|---|---|---|---|---|---|
| HT1080 | A-172 | HT1080 | A-172 | ||
| 2,5-DHBA–gelatin | 1000 | 104 ± 15 | 97 ± 12 | 102 ± 16 | 102 ± 13 |
| 300 | 96 ± 12 | 96 ± 14 | 99 ± 14 | 101 ± 12 | |
| 100 | 103 ± 16 | 100 ± 13 | 104 ± 17 | 94 ± 17 | |
| 30 | 95 ± 13 | 93 ± 14 | 96 ± 17 | 96 ± 16 | |
| 10 | 97 ± 15 | 99 ± 16 | 93 ± 14 | 96 ± 15 | |
| Heparin | 1000 | 104 ± 17 | 97 ± 14 | 95 ± 15 | 92 ± 15 |
| 300 | 101 ± 13 | 99 ± 14 | 94 ± 12 | 98 ± 14 | |
| 100 | 101 ± 12 | 105 ± 15 | 99 ± 13 | 98 ± 14 | |
| 30 | 93 ± 14 | 96 ± 15 | 103 ± 15 | 102 ± 16 | |
| 10 | 98 ± 13 | 94 ± 13 | 101 ± 15 | 95 ± 11 | |
| Chondroitin sulfate | 1000 | 100 ± 16 | 99 ± 12 | 91 ± 12 | 94 ± 16 |
| 300 | 100 ± 12 | 103 ± 13 | 95 ± 14 | 92 ± 15 | |
| 100 | 104 ± 16 | 105 ± 13 | 93 ± 12 | 95 ± 15 | |
| 30 | 94 ± 14 | 101 ± 16 | 99 ± 12 | 100 ± 13 | |
| 10 | 99 ± 16 | 95 ± 16 | 100 ± 13 | 101 ± 13 | |
| Dermatan sulfate | 1000 | 99 ± 17 | 102 ± 15 | 92 ± 13 | 91 ± 15 |
| 300 | 103 ± 14 | 97 ± 13 | 96 ± 14 | 93 ± 14 | |
| 100 | 104 ± 18 | 100 ± 14 | 101 ± 13 | 95 ± 14 | |
| 30 | 101 ± 16 | 101 ± 15 | 95 ± 14 | 104 ± 14 | |
| 10 | 97 ± 14 | 96 ± 14 | 103 ± 15 | 107 ± 17 | |
| Geldanamycin (b) | 5.6 | 5 ± 3 | 0 | 0 | 0 |
| 0.56 | 43 ± 9 | 3 ± 3 | 13 ± 7 | 0 | |
| 0.056 | 94 ± 11 | 13 ± 4 | 55 ± 12 | 2 ± 2 | |
| 0.0056 | 98 ± 15 | 78 ± 14 | 100 ± 16 | 57 ± 8 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snigireva, A.V.; Morenkov, O.S.; Skarga, Y.Y.; Lisov, A.V.; Lisova, Z.A.; Leontievsky, A.A.; Zhmurina, M.A.; Petrenko, V.S.; Vrublevskaya, V.V. A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate Inhibits the Basal and Hsp90-Stimulated Migration and Invasion of Tumor Cells. J. Funct. Biomater. 2020, 11, 39. https://doi.org/10.3390/jfb11020039
Snigireva AV, Morenkov OS, Skarga YY, Lisov AV, Lisova ZA, Leontievsky AA, Zhmurina MA, Petrenko VS, Vrublevskaya VV. A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate Inhibits the Basal and Hsp90-Stimulated Migration and Invasion of Tumor Cells. Journal of Functional Biomaterials. 2020; 11(2):39. https://doi.org/10.3390/jfb11020039
Chicago/Turabian StyleSnigireva, Anastasiya V., Oleg S. Morenkov, Yuri Y. Skarga, Alexander V. Lisov, Zoya A. Lisova, Alexey A. Leontievsky, Mariya A. Zhmurina, Viktoria S. Petrenko, and Veronika V. Vrublevskaya. 2020. "A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate Inhibits the Basal and Hsp90-Stimulated Migration and Invasion of Tumor Cells" Journal of Functional Biomaterials 11, no. 2: 39. https://doi.org/10.3390/jfb11020039
APA StyleSnigireva, A. V., Morenkov, O. S., Skarga, Y. Y., Lisov, A. V., Lisova, Z. A., Leontievsky, A. A., Zhmurina, M. A., Petrenko, V. S., & Vrublevskaya, V. V. (2020). A 2,5-Dihydroxybenzoic Acid–Gelatin Conjugate Inhibits the Basal and Hsp90-Stimulated Migration and Invasion of Tumor Cells. Journal of Functional Biomaterials, 11(2), 39. https://doi.org/10.3390/jfb11020039