
Diversity and Distribution of Mesozooplankton in the Coastal Southwestern Mediterranean Alboran Sea, during Summer: What Are the Driving Factors?
 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
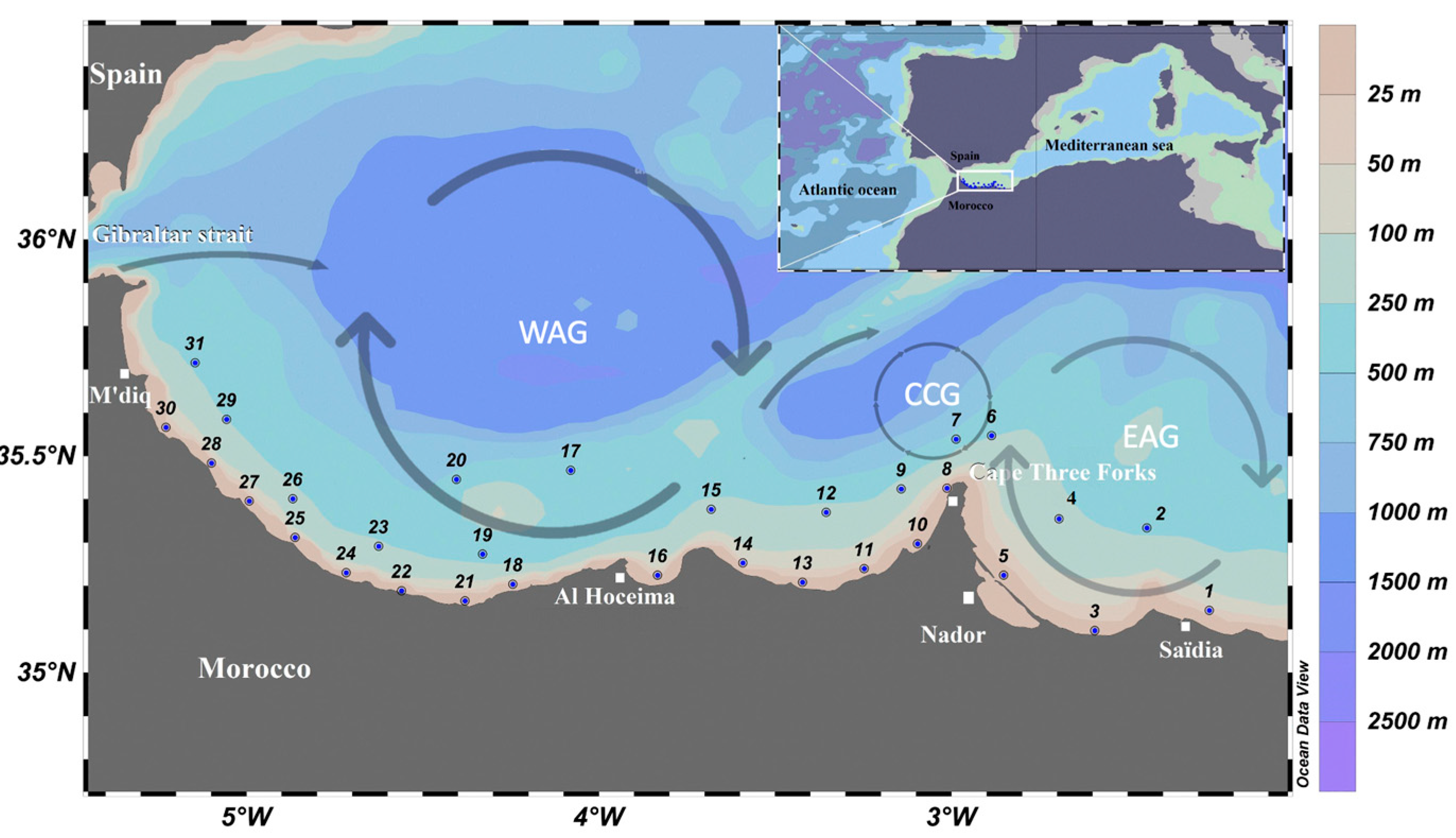
2.1. Study Area, Sampling, and Measurements
2.2. Data Analysis
- -
- The specific richness (S), which is the number of encountered species.
- -
- The relative abundance of species i in a sample:
- -
- The Shannon–Weaver diversity index (H’; [45]) is used to characterize species diversity in a community. It establishes the link between the number of species (ni) and the number of individuals (N) within the ecosystem or a community. It is measured with the following formula:
- -
- The Pielou index (J; [46]), a measure of the equitability (or equidistribution) of the species, is calculated according to the following formula:
2.3. Statistical Analysis
3. Results
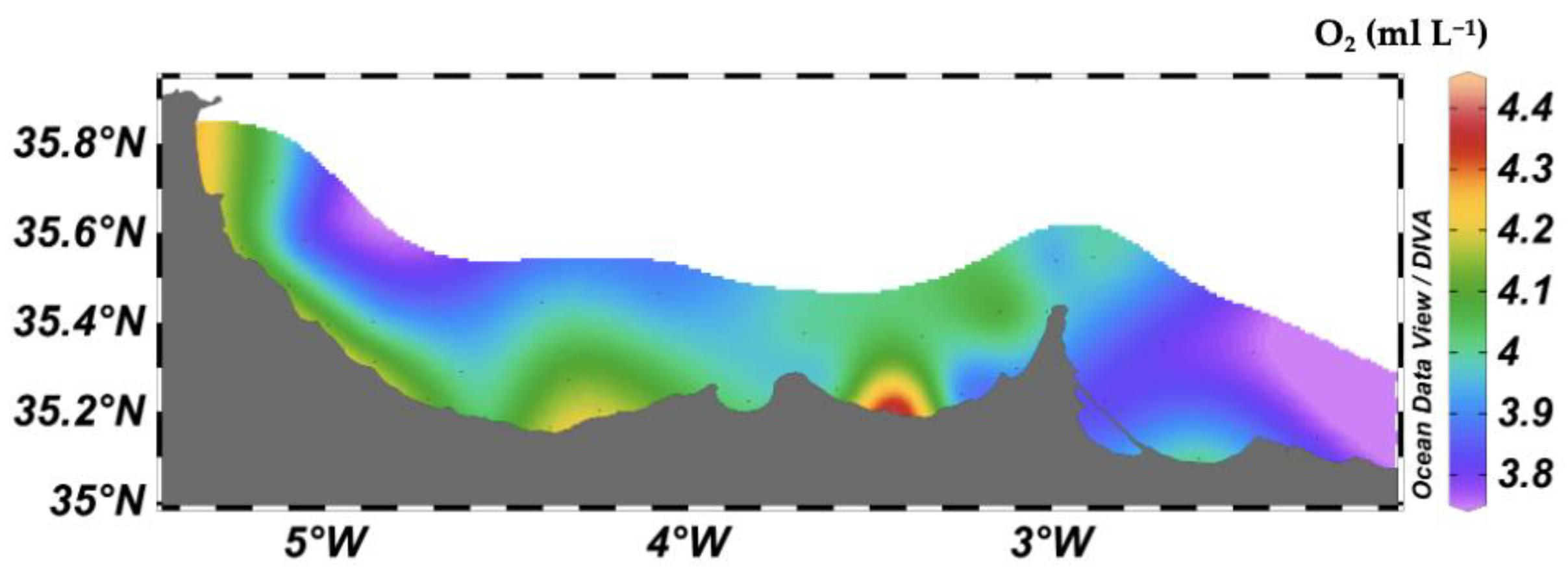
3.1. Environmental Parameters
3.2. Vertical Distribution of Environmental Parameters from T–S Diagrams
3.3. Relationships between Mesozooplankton Biomass and Environmental Factors
3.4. Diversity, Biomass, and Abundance of Mesozooplankton
3.5. Mesozooplankton Community Structure
3.6. Relationships between Mesozooplankton Community and Environmental Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Beaugrand, G.; Edwards, M.; Legendre, L. Marine biodiversity, ecosystem functioning, and carbon cycles. Proc. Natl. Acad. Sci. USA 2010, 107, 10120–10124. [Google Scholar] [CrossRef] [PubMed]
- Brander, K. Impacts of climate change on fisheries. J. Mar. Syst. 2010, 79, 389–402. [Google Scholar] [CrossRef]
- Burrows, M.T.; Schoeman, D.S.; Buckley, L.B.; Moore, P.; Poloczanska, E.S.; Brander, K.M.; Brown, C.; Bruno, J.F.; Duarte, C.M.; Halpern, B.S.; et al. The Pace of Shifting Climate in Marine and Terrestrial. Ecosyst. Sci. 2011, 334, 652–655. [Google Scholar] [CrossRef]
- Steinberg, D.K.; Landry, M.R. Zooplankton and the Ocean Carbon Cycle. Annu. Rev. Mar. Sci. 2017, 9, 413–444. [Google Scholar] [CrossRef] [PubMed]
- Keister, J.E.; Bonnet, D.; Chiba, S.; Johnson, C.L.; Mackas, D.L.; Escribano, R. Zooplankton population connections, community dynamics, and climate variability. ICES J. Mar. Sci. 2012, 69, 347–350. [Google Scholar] [CrossRef]
- Rice, E.; Stewart, G. Decadal changes in zooplankton abundance and phenology of long and Island Sound reflect interacting changes in temperature and community composition. J. Mar. Environ. Res. 2016, 120, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Garrido, J.C.; Lafuente, J.G.; Fanjul, E.; Sotillo, M.G.; Santos, F.J.D.L. «What Does Cause the Collapse of the Western Alboran Gyre, Results of an Operational Ocean Model». J. Prog. Oceanogr. 2013, 116, 142–153. [Google Scholar] [CrossRef]
- Rohling, E.J.; Abu-Zied, R. The Marine Environment: Present and Past. In The Physical Geography of the Mediterranean; Jamie, W., Ed.; Oxford Academic: Oxford, UK, 2009. [Google Scholar] [CrossRef]
- Bergamasco, A.; Malanotte-Rizzoli, P. The circulation of the Mediterranean Sea: A historical review of experimental investigations. Adva. Oceanogr. Limn. 2010, 1, 11–28. [Google Scholar] [CrossRef]
- Vargas-Yáñez, M.; Plaza, F.; García-Lafuente, J.; Sarhan, T.; Vargas, J.M.; Vélez-Belchí, P. About the seasonal variability of the Alboran Sea circulation. J. Mar. Syst. 2002, 35, 229–248. [Google Scholar] [CrossRef]
- Seguin, G.; Gaudy, R.; Errhif, A.; Thibault, D. Observations on the abundance, taxonomic composition and ecological affinities of pelagic copepods collected from the Almeria-Oran frontal region. J. Mar. Life 1993, 3, 19–29. [Google Scholar]
- Gaudy, R.; Youssara, F. Variations of zooplankton metabolism and feeding in the frontal area of the Alboran Sea (western Mediterranean) in winter. Oceanologica Acta 2003, 26, 179–189. [Google Scholar] [CrossRef][Green Version]
- Riandey, V. Etude de la Structure de Taille des Populations Zooplanctoniques Dans Différents Écosystèmes Marins. Apports des Nouveaux Appareils Automatisés D’observation. Ph.D. Thesis, Université de la Méditerranée—Aix Marseille II, Marseille, France, 2005; p. 372. [Google Scholar]
- Gómez, F.; Gorsky, G.; Striby, L.; Vargas, J.M.; Gonzalez, N.; Picheral, M.; García-Lafuente, J.; Varela, M.; Goutx, M. Small-scale temporal variations in biogeochemical features in the Strait of Gibraltar, Mediterranean side—The role of NACW and the interface oscillation. J. Mar. Syst. 2001, 30, 207–220. [Google Scholar] [CrossRef]
- Tintoré, J.; Gomis, D.; Alonso, S.; Parrilla, G. Mesoscale dynamics and vertical l motion in the Alborán Sea. J. Phys. Oceanogr. 1991, 21, 811–823. [Google Scholar] [CrossRef]
- García Lafuente, J.; Delgado, J. The Meandering Path of a Drifter around the Western Alboran Gyre. J. Phys. Oceanogr. 2004, 34, 685–692. [Google Scholar] [CrossRef]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcala, M.; Vaqué, D.; Zingone, A. Plankton in the open Mediterranean Sea: A review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef]
- Mazzocchi, M.G.; Siokou, I.; Tirelli, V.; Bandelj, V.; Fernandez de Puelles, M.L.; Ak Örek, Y.; de Olazabal, A.; Gubanova, A.; Kress, N.; Protopapa, M.; et al. Regional and seasonal characteristics of epipelagic mesozooplankton in the Mediterranean Sea based on an artificial neural network analysis. J. Mar. Syst. 2014, 135, 64–80. [Google Scholar] [CrossRef]
- Zakaria, H.Y.; Hassan, A.M.; Abo-Senna, F.M.; El-Naggar, H.A. Abundance, distribution, diversity and zoogeography of epipelagic copepods off the Egyptian Coast (Mediterranean Sea). Egypt. J. Aquat. Res. 2016, 42, 459–473. [Google Scholar] [CrossRef]
- Khelifi-Touhami, M.; Semroud, R.; Hamdi, H.; Ounissi, M.; Haridi, A.; Nejib Daly, M.; Aissa, P. The planktonic copepod communi-ties from the southern Mediterranean Sea (Algeria, Tunisia) with a re-description of Paracalanus indicus Wolfen-den 1905 (Copepoda: Calanoida). Cah. Biol. Mar. 2007, 48, 327–337. [Google Scholar]
- Berraho, A.; Abdelouahab, H.; Charib, S.; Essarraj, S.; Larissi, J.; Abdellaoui, B.; Christou, E.D. Copepod community along the Mediterranean coast of Morocco (Southwestern Alboran Sea) during spring. Med. Mar. Sci. 2016, 17, 661–665. [Google Scholar] [CrossRef][Green Version]
- Ounissi, M.; Laskri, H.; Khélifi-Touhami, M. Net-zooplankton abundance and biomass from Annaba Bay (SW Mediterranean Sea) under estuarine influences. Mediterr. Mar. Sci. 2016, 17, 519–532. [Google Scholar] [CrossRef]
- Alcaraz, M.; Calbet, A.; Estrada, M.; Marrasé, C.; Saiz, E.; Trepat, I. Physical control of zooplankton communities in the Catalan Sea. J. Prog. Oceanogr. 2007, 74, 294–312. [Google Scholar] [CrossRef]
- Fernandez de Puelles, M.L.; Alemany, F.; Jansá, J. Zooplankton time-series in the Balearic Sea (Western Mediterranean): Variability during the decade 1994–2003. J. Prog. Oceanogr. 2007, 74, 329–354. [Google Scholar] [CrossRef]
- Raybaud, V.; Nival, P.; Mousseau, L.; Gubanova, A.; Altukhov, D.; Khvorov, S.; Ibañez, F.; Andersen, V. Short term changes in zooplankton com-munity during the summer-autumn transition in the open NW Mediterranean Sea: Species composition, abundance and diversity. J. Biogeosciences. 2008, 5, 1765–1782. [Google Scholar] [CrossRef]
- García-Comas, C.; Stemmann, L.; Ibanez, F.; Berline, L.; Mazzocchi, M.G.; Gasparini, S.; Picheral, M.; Gorsky, G. Zooplankton long-term changes in the NW Mediterranean Sea: Decadal periodicity forced by winter hydrographic conditions related to large-scale atmospheric change. J. Mar. Syst. 2011, 87, 216–226. [Google Scholar] [CrossRef]
- Mazzocchi, M.G.; Licandro, P.; Dubroca, L.; Di Capua, I.; Saggiomo, V. Zooplankton associations in a Mediterranean long-term time-series. J. Plankton Res. 2011, 33, 1163–1181. [Google Scholar] [CrossRef]
- Zerouali, F.; et Melhaoui, M. Diversité Spécifique et Biomasse des Copépodes Pélagiques de la Région de Ras Kebdana (Côte Méditerranéenne Marocaine); Actes Institut Agronomique Vétérinaire: Maroc, North Africa, 2002; Volume 22, pp. 91–102. [Google Scholar]
- Zaafa, A.; Ettahiri, O.; Berraho, A.; Elkhiati, N.; Somoue, L.; Zizah, S.; Ramdani, M.; Blaghen, M.; Flower, R. A comparative study of marine zooplankton communities in the Tangier and M’Diq (Gibraltar strait) regions. Hydroécologie Appliqué 2012, 18, 67–80. [Google Scholar] [CrossRef]
- Furnestin, M.L. Chaetognathes et zooplancton du secteur atlantique Marocain. Rev. Trav. L‘institut Tech. Sci. Pêches Marit. 1957, 21, 1–356. [Google Scholar]
- Furnestin, M.L.; et Belfequih, M. Les copépodes du plateau continental marocain et du détroit canarien. II-Les espèces au cours d’un cycle annuel dans les zones d’upwelling. J. Cons. Int. Explor. Mer. 1976, 9, 22–46. [Google Scholar]
- Blackburn, N. Distribution of plankton biomass in an upwelling area off northwest Africa: Preliminary results from expedition joint–I. Coast. Upw. Ecosys. Anal (CUEA) News Lett. 1976, 5, 13–19. [Google Scholar]
- Belfequih, M. Les Copépodes du Plateau Atlanto-Marocain. Un Cycle Annuel Dans les Zones D’upwelling. Ph.D. Thesis, University Aix-Marseille I, Marseille, France, 1980; 126p. [Google Scholar]
- Boucher, J. Peuplement de copépodes des upwellings côtiers nord-ouest africains. II Maintien de la localisation spatiale. Oceanol. Acta 1982, 5, 199–207. [Google Scholar]
- Chiahou, B.; et Ramdani, M. Observations sur les variations de la densité et de la biomasse des copépodes pélagiques des côtes atlantiques marocaines (région d’El Jadida) au cours d’un cycle annuel. Mar. Life 1996, 6, 21–25. [Google Scholar]
- Somoue, L.; Elkhiati, N.; Ramdani, M.; Thong, L.H.; Ettahiri, O.; Berraho, A.; Thang, D.C. Abundance and structure of copepod communities along the Atlantic coast of Southern Morocco. Acta Adr. 2005, 46, 63–76. [Google Scholar]
- Berraho, A.; Somoue, L.; Hernandez-Leon, S.; Valdés, L. Zooplankton in the Canary Current Large Marine Ecosystem. Oceanographic and Biological features in the Canary Current Large Marine Ecosystem. IOC UNESCO Tech. Ser. 2015, 115, 183–195. [Google Scholar]
- Murphy, E. Spatial structure of the Southern Ocean ecosystem: Predator-prey linkages in Southern Ocean food webs. J. Anim. Ecol. 1995, 64, 333–347. [Google Scholar] [CrossRef]
- Robin, R.S.; Srinivasan, M.; Chandrasekar, K. Distribution of zooplankton from Arabian Sea, along Southern Kerala (Southwest Coast of India) during the cruise. Curr. Res. J. Biol Sci. 2009, 1, 155–159. [Google Scholar]
- Linder, S. A proposal for the use of standardized methods for chlorophyll determinations in ecological and eco-physiological investigations. Physiol. Plant 1974, 32, 154–156. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis. Fish. Res. Board Canada 1972, 167, 405. [Google Scholar] [CrossRef]
- Aminot, A.; Chaussepied, M. Manuel des analyses chimiques en milieu marin. Cent. Natl. Pour L’exploitation Océans 1983, 551, 5–395. [Google Scholar]
- Motoda, S. Devices of simple plankton apparatus. Mem. Fac. Fish. Hokkaido Univ. 1959, 7, 73–94. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 117. [Google Scholar]
- Pielou, E.C. Species diversity and pattern diversity in the study of ecological succession. J. Theoret. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analyses and Interpretation; Natural Environment Research Council: Plymouth, UK, 1994. [Google Scholar]
- Anderson, T. The Theory and Practice of Online Learning; Athabasca University Press: Athabasca, AB, Canada, 2008. [Google Scholar] [CrossRef]
- Schlitzer, R. Ocean Data View. Available online: https://odv.awi.de (accessed on 1 May 2022).
- Yebra, L.; Herrera, I.; Mercado, J.M.; Cortés, D.; Gómez-Jakobsen, F.; Alonso, A.; Sánchez, A.; Salles, S.; Valcárcel-Pérez, N. Zooplankton production and carbon export flux in the western Alboran Sea gyre (SW Mediterranean). J. Prog. Oceanogr. 2018, 167, 64–77. [Google Scholar] [CrossRef]
- Johnson, W.S.; Allen, D.M. Zooplankton of the Atlantic and Gulf Coast; Johns Hopkins University Press: Baltimore, MA, USA, 2005. [Google Scholar]
- Idmoussi, H.; Somoue, L.; Peters, F.; Demarcq, H.; Laabir, M.; Estrada, M.; Arin, L.; Makaoui, A.; Ettahiri, O.; Baibai, T.; et al. Patterns of microphytoplankton biomass and diversity in the southern Alboran Sea. J. Med. Mar. Sci. 2021, 23, 103–114. [Google Scholar] [CrossRef]
- METAP. Le Programme D’Assistance Technique Pour la Protection de L’Environnement de la Méditerranée (Metap): Seize Années de Succès Dans L’Amélioration de L’Environnement Dans Les Pays du Maghreb. La Lettre D’information Trimestrielle du Groupe de la Banque Mondiale au Maghreb—Septembre 2006—N°3; 2006. Available online: http://hdl.handle.net/123456789/30291 (accessed on 1 July 2023).
- Bryden, H.L.; Stommel, H.M. Origin of the Mediterranean outflow. J. Mar. Res. 1982, 40, 55–71. [Google Scholar]
- Naranjo, C.; Lafuente, J.G.; Garrido, J.C.S.; Román, A.S.; Cabello, J.D. The Western Alboran Gyre helps ventilate the Western Mediterranean Deep Water through Gibraltar. Deep. Sea Res. Part I J. Oceanogr. Res. Rap. 2012, 63, 157–163. [Google Scholar] [CrossRef]
- García-Lafuente, J.; Naranjo, C.; Sammartino, S.; Sánchez-Garrido, J.C.; Delgado, J. The Mediterranean outflow in the Strait of Gibraltar and its connection with upstream conditions in the Alborán Sea. Ocean Sci. 2017, 13, 195–207. [Google Scholar] [CrossRef]
- Youssara, F.; Gaudy, R. Variations of zooplankton in the frontal area of the Alboran sea (Mediterranean Sea) in winter 1997. J. Oceanol. Acta 2001, 24, 361–376. [Google Scholar] [CrossRef][Green Version]
- Chen, C.-T.; Bănaru, D.; Carlotti, F.; Faucheux, M.; Harmelin-Vivien, M. Seasonal variation in biochemical and energy content of size-fractionated zooplankton in the Bay of Marseille (North-Western Mediterranean Sea). J. Mar. Syst. 2019, 199, 103223. [Google Scholar] [CrossRef]
- Protopapa, M.; Zervoudaki, S.; Tsangaris, C.; Velaoras, D.; Koppelmann, R.; Psarra, S.; Möllmann, C. Zooplankton distribution, growth and respiration in the Cretan Passage, Eastern Mediterranean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2019, 164, 156–169. [Google Scholar] [CrossRef]
- Somoue, L. Structure des Communautés Planctoniques de L’écosystème Pélagique de L’atlantique sud Marocain Entre Cap Boujdor et Cap Blanc (26°30′–21° N). Ph.D. Thesis, Nationale, Faculty of Science Ain Chock, University Hassan II, Casablanca, Morocco, 2004; p. 211. [Google Scholar]
- El Arraj, L.; Tazi, O.; Somoue, L.; Hilmi, K.; Serghini, M.; Ettahiri, O. Diversity and Copepods’composition off Moroccan Atlantic Coast (Northwest Africa): A Review. Eur. Sci. J. 2017, 13, 272. [Google Scholar] [CrossRef][Green Version]
- Gaudy, R. Features and peculiarities of zooplankton communities from the western Mediterranean. In Mediterranean Marine Ecosystems; Moraitou-Apostolopoulou, M., Kiortsis, V., Eds.; Plenum: New York, NY, USA, 1985; pp. 279–301. [Google Scholar] [CrossRef]
- González-Quirós, R.; Checkley, D.M.; Silva, N.; Freire, J.; Valdés, L. Effects of environmental conditions on the seasonal distribution of phytoplankton biomass in the upwelling ecosystem off central-southern Chile. J. Mar. Syst. 2010, 80, 1–19. [Google Scholar]
- Ramírez-Romero, E.; Palomera, I.; Sabatés, A. Environmental drivers of mesozooplankton biomass variability in the North-Western Mediterranean Sea. Prog. Oceanogr. 2018, 166, 227–240. [Google Scholar]
- Gallienne, C.P.; Robins, D.B.; Woodd-Walker, R.S. Abundance, variance and diversity of zooplankton in the sub-tropical Atlantic: A comparison of nepheloid layer and water column assemblages at the Mid-Atlantic Ridge. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2004, 51, 365–382. [Google Scholar]
- Millot, C. Circulation in the Western Mediterranean Sea. J. Mar. Syst. 1999, 20, 423–442. [Google Scholar] [CrossRef]
- Gubanova, A.; Altukhov, D.; Mukhanov, V. Diversity and structure of zooplankton communities in the southeastern Black Sea in relation to environmental factors. Turk. J. Fish. Aquat. Sci. 2011, 11, 375–385. [Google Scholar]
- Kiørboe, T.P.; Sabatini, M.; Tiselius, P. The influence of viscosity on swimming and feeding of marine copepods. J. Exp. Mar. Biol. Ecol. 2002, 281, 61–75. [Google Scholar]
- Hsieh, C.H.; Reiss, C.S.; Hunter, J.R.; Beddington, J.R.; May, R.M.; Sugihara, G. Fishing elevates variability in the abundance of exploited species. Nature 2004, 427, 692–695. [Google Scholar] [CrossRef]
- Chiba, S.; Tadokoro, K.; Saino, T.; Nakata, H. Vertical distribution and community structure of mesozooplankton in the central Arctic Ocean in summer. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 663–681. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Pi |
|---|---|
| Amphipoda | 0.07 |
| Annelida | 0.1 |
| Appendicularia | 5.3 |
| Chaetognata | 3.6 |
| Cladocera | 35.6 |
| Copepoda | 48.5 |
| Meroplankton larvae of crustaceans | 1.5 |
| Foraminifera | 0.004 |
| Echinodermata larvae | 0.01 |
| Gastropod Veliger | 1.6 |
| Ostracoda | 0.1 |
| Radiolaria | 0.3 |
| Salpida | 1.9 |
| Siphonophora | 1.4 |
| Species | Pi |
|---|---|
| Evadne nordmanni (Lovén, 1836) | 13.51 |
| Penilia avirostris (Dana, 1849) | 81.93 |
| Podon leuckartii (G.O. Sars, 1862) | 4.55 |
| Species | Pi |
|---|---|
| Acartia clausi (Giesbrecht, 1889) | 3.07 |
| Calanus helgolandicus (Claus, 1863) | 0.10 |
| Calocalanus contractus (Farran, 1926) | 0.12 |
| Calocalanus pavo (Farran, 1926) | 0.10 |
| Centropages typicus (Krøyer, 1849) | 0.08 |
| Clausocalanus arcuicornis (Dana, 1849) | 0.43 |
| Clausocalanus furcatus (Brady, 1883) | 8.25 |
| Eucalanus elongatus (Dana, 1848) | 3.44 |
| Euchaeta acuta (Giesbrecht, 1893) | 0.93 |
| Euterpina acutifrons (Dana, 1847) | 6.40 |
| Farranula rostrata (Claus, 1863) | 1.82 |
| Macrosetella gracilis (Dana, 1846) | 0.12 |
| Microsetella norvegica (Boeck, 1865) | 0.24 |
| Oithona nana (Giesbrecht, 1893) | 2.71 |
| Oithona plumifera (Baird, 1843) | 6.71 |
| Oncaea curta (Sars G.O., 1916) | 1.59 |
| Oncaea venusta (Philippi, 1843) | 12.71 |
| Paracalanus parvus (Claus, 1863) | 31.18 |
| Pseudocalanus elongatus (Brady, 1865) | 3.65 |
| Rhincalanus nasutus (Giesbrecht, 1888) | 0.40 |
| Sapphirina intestinata (Giesbrecht, 1891) | 0.65 |
| Saphirina sp. (Thompson, J.V. 1829) | 0.21 |
| Spinocalanus magnus (Wolfenden, 1904) | 0.12 |
| Subeucalanus crassus (Giesbrecht, 1888) | 0.02 |
| Subeucalanus monachus (Giesbrecht, 1888) | 0.02 |
| Temora longicornus (Müller, 1785) | 0.78 |
| Temora stylifera (Dana, 1849) | 14.00 |
| Variable | Pseudo-F | P | Prop |
|---|---|---|---|
| LongE | 6.4673 | 0.001 | 0.18235 |
| Depth | 1.4828 | 0.134 | 4.8643 × 10−2 |
| TSS | 1.6235 | 0.094 | 5.3013 × 10−2 |
| SSS | 5.4289 | 0.001 | 0.15768 |
| O2 | 2.6177 | 0.088 | 5.2836 × 10−2 |
| Chl-a | 1.7809 | 0.058 | 5.7859 × 10−2 |
| Group East | Group West | |||
|---|---|---|---|---|
| Variable | Av. Value | Av. Value | Contrib% | Cum.% |
| Penilia avirostris | 117 | 663 | 60.90 | 60.90 |
| Pracalanus parvus | 95.2 | 307 | 15.19 | 76.09 |
| Evadne nordmanni | 29 | 101 | 5.30 | 81.39 |
| Appendicularia | 25.7 | 117 | 3.78 | 85.17 |
| Oncaea venusta | 13.7 | 146 | 3.46 | 88.63 |
| Temora stylifera | 43.2 | 137 | 3.36 | 91.99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benallal, M.R.; Errhif, A.; Somoue, L.; Laabir, M.; Demarcq, H.; Idrissi, M.; Agouzouk, A.; Goliat, Y.; Idmoussi, H.; Makaoui, A.; et al. Diversity and Distribution of Mesozooplankton in the Coastal Southwestern Mediterranean Alboran Sea, during Summer: What Are the Driving Factors? J. Mar. Sci. Eng. 2024, 12, 674. https://doi.org/10.3390/jmse12040674
Benallal MR, Errhif A, Somoue L, Laabir M, Demarcq H, Idrissi M, Agouzouk A, Goliat Y, Idmoussi H, Makaoui A, et al. Diversity and Distribution of Mesozooplankton in the Coastal Southwestern Mediterranean Alboran Sea, during Summer: What Are the Driving Factors? Journal of Marine Science and Engineering. 2024; 12(4):674. https://doi.org/10.3390/jmse12040674
Chicago/Turabian StyleBenallal, Mohamed Reda, Ahmed Errhif, Laila Somoue, Mohamed Laabir, Hervé Demarcq, Mohammed Idrissi, Aziz Agouzouk, Yassine Goliat, Hajar Idmoussi, Ahmed Makaoui, and et al. 2024. "Diversity and Distribution of Mesozooplankton in the Coastal Southwestern Mediterranean Alboran Sea, during Summer: What Are the Driving Factors?" Journal of Marine Science and Engineering 12, no. 4: 674. https://doi.org/10.3390/jmse12040674
APA StyleBenallal, M. R., Errhif, A., Somoue, L., Laabir, M., Demarcq, H., Idrissi, M., Agouzouk, A., Goliat, Y., Idmoussi, H., Makaoui, A., & Ettahiri, O. (2024). Diversity and Distribution of Mesozooplankton in the Coastal Southwestern Mediterranean Alboran Sea, during Summer: What Are the Driving Factors? Journal of Marine Science and Engineering, 12(4), 674. https://doi.org/10.3390/jmse12040674