Abstract
This work aimed to reveal the heterotrophic nitrification and aerobic denitrification activities in a salt-tolerant strain, Tritonibacter mobile HN1, isolated from mariculture sludge by using a pure culture method. Strain HN1 showed the removal rates of ammonia, nitrite, nitrate, and total nitrogen of 98.22%, 100%, 95.71%, and 86.63%, respectively, with the carbon source of sodium citrate or sodium succinate, ratio of carbon to nitrogen of 15, salinity of 3%, temperature of 30 °C, shaking speed of 120 rpm, and pH of 8. The genes of narG, narH, narI, nirS, norB, norC, nosZ, glnA, and GltB were found in the genome sequence of strain HN1, confirming the pathways of nitrogen assimilation, heterotrophic nitrification, and aerobic denitrification. In addition, two nitrifying genes, amo and nap, were missing in the genome of Tritonibacter mobile HN1, indicating that HN1 may have novel genes for this function. This study showed that HN1 had the potential to remove nitrogen contaminants in saline environments and was the first Tritonibacter mobilis strain with heterotrophic nitrification and aerobic denitrification capabilities.
1. Introduction
In recent years, exogenous nitrogen-containing pollutants substantially increased with the acceleration of urbanization and industrialization, as well as the continuous development of intensive aquaculture. The excessive discharge of nitrogen-containing pollutants caused eutrophication and frequent occurrence of red tides in marine environments, leading to an ecological imbalance and threatening human health [1]. Aerobic autotrophic nitrification and anaerobic heterotrophic denitrification were included in traditional biological denitrification processes to remove the pollution [2]. Nitrification was the sequential oxidation of ammonia to nitrite and nitrate by autotrophic microorganisms under aerobic conditions, and denitrification was the process in which heterotrophic microorganisms reduced nitrate to nitrogen under strict anaerobic conditions [2].
In the traditional understanding, the two processes of nitrification and denitrification occurred in different bacteria [2]. Recently, heterotrophic nitrification–aerobic denitrification (HN-AD) bacteria were found to achieve synchronous nitrification–denitrification (SND) in aerobic environments, which broke through the traditional denitrification theories [3,4,5,6]. Compared to the traditional denitrification process, HN-AD strains simultaneously performed the nitrification and denitrification processes, thereby simplifying the process flow and reducing the operating costs for removing the nitrogen pollution from the environment [7,8,9]. However, the low tolerance of HN-AD bacteria to special environments, such as high salinity environments, still limited the application of these organisms in industrial practice, therefore becoming a hot research topic in recent years [10,11].
The current work isolated an HN-AD bacterium, Tritonibacter mobilis HN1, from the sediments of a sewage-treatment plant and aimed to reveal the nitrogen removal ability of Tritonibacter mobilis HN1 in a saline environment and related metabolic pathways, which provided a potential strain for denitrification in saline wastewater and contributed to a better understanding and exploration of Tritonibacter mobilis strains.
2. Materials and Methods
2.1. Preparation of Culture Medium
The modified Stephenson culture medium used in this experiment was purchased from Shandong Tuopu Biotechnology Co., Ltd. (Yantai, Shangdong, China). The modified Stephenson culture medium contained the following ingredients: 0.01 g of MnSO4·4H2O, 0.25 g of NaH2PO4, 0.75 g of K2HPO4, 0.5 g of CaCO3, 0.03 g of MgSO4·7H2O, 2.0 g of (NH4)2SO4, and 1 L of artificial seawater (pH 8.2 ± 0.2). The bacteria were enriched and isolated using HNM1 medium, which contained 21.74 g of CH3COONa in 1 L modified Stephenson culture medium.
The medium used for screening experiments that can simultaneously conduct nitrification and denitrification included 0.3607 g of KNO3, 0.2357 g of (NH4)2SO4, 0.25 g of NaH2PO4, 0.03 g of MgSO4·7H2O, 0.01 g of MnCl2·4H2O, 5.125 g of CH3COONa, and 0.75 g of K2HPO4 in 1 L of artificial seawater.
The heterotrophic nitrification medium (HNM2) contained the following components: 0.75 g of K2HPO4, 0.4714g of (NH4)2SO4, 0.25 g of NaH2PO4, 0.01g of MnCl2·4H2O, 0.03 g of MgSO4·7H2O, 6.125 g of Na3C6H5O7·2H2O, and 1 L of artificial seawater, which was also used as a general purpose medium for the rapid growth of bacteria.
The composition of the denitrifying media DM2 and DM1 was the same as HNM2, except that the artificial seawater contained 0.7214 g of KNO3 and 0.4929 g of NaNO2 per liter to replace (NH4)2SO4. The artificial seawater used in the experiment was prepared from sea salt produced by Guangzhou Yier BE Co., Ltd. (Guangzhou, China). The initial pH of all experimental culture media was adjusted to 7.0 ± 0.05 before autoclaving at 121 °C for 20 min. A total of 2% (v/v) agar was added to the above liquid culture medium before sterilization to prepare solid culture medium.
2.2. Isolation and Identification
Sediment samples were obtained from the secondary sedimentation tank of Zhujiajian Water Co., Ltd., a wastewater treatment plant for treating urban sewage located in Zhoushan City, Zhejiang Province, China. A total of 10 mL of sample and 90 mL of sterile HNM1 medium were mixed in a 250 mL conical flask. The flask was cultivated at 30 °C in a rotary shaker for 3 days (120 rpm). After 3 days of incubation, the enrichment solution was diluted to different concentrations (10−4, 10−5, 10−6, and 10−7) with sterile artificial seawater, and the above dilutions were spread on HNM1 solid plates in 3 parallel samples for each gradient. After that, the solid plates were incubated in a 30 °C incubator until the appearance of colonies was visible to the naked eye, the coated plates were observed, and different single colonies were selected by inoculation rings using colony morphology, degree of protrusion, color and other characteristics, and the above procedure was repeated until a pure colony was isolated. The pure colonies were incubated at 120 rpm and 30 °C for 72 h. The denitrification abilities of isolates were measured by continuously monitoring the concentrations of total nitrogen (TN) and ammonium (see Section 2.6). The isolate with the highest denitrification efficiency was mixed with sterile glycerol at the final concentration of 20% (v/v) and stored at −80 °C for further experiments.
DNA extraction kit (Omega Bio tek, Mogadore, OH, USA) was used to extract DNA from the bacterium according to the manufacturer’s instructions for identification. The purified DNA was amplified using polymerase chain reaction (PCR) for the 16S rRNA gene with primers 27F (5′-AGGTTTGATCCTCAG-3′) and 1492R (5′-GGTACTTACGACT-3′) [12]. DNA sequencing was performed through Majorbio, and online BLAST program was used for homology comparison (EZBioCloud; https://www.ezbiocloud.net; accessed on 16 January 2021). In mega7, a phylogenetic tree was constructed using Neighbor-Joining method with a bootstrap value of 1000 [13].
2.3. Mensuration of Ammonium Removal Rate of Strain HN1
Strain HN1 with highest nitrogen removal activity among the isolates was cultured in 100 mL sterile HNM2 medium at a temperature of 30 °C, a rotational speed of 120 rpm, and an inoculum size of 1% (v/v). Samples were collected and measured every 12 h to determine the concentrations of NO3−-N, NO2−-N, NH4+-N, TN, and OD600 by the methods in Section 2.6. In addition, the intracellular nitrogen content was calculated by subtracting the TN of the centrifuged medium (5000 r/min at 10 °C for 15 min) from that of non-centrifuged medium based on previous research results [14].
2.4. Influence of Various Factors on HN-AD Capabilities of Strain HN1
Single-factor experiments were conducted in DM1, DM2, and HNM2, respectively, with appropriate adjustments including carbon sources, C/N, temperature, pH, salinity, and shaking speed, to determine the influence of different factors on the heterotrophic nitrification and aerobic denitrification capability and growth of strain HN1.
Sodium acetate, sodium citrate, D-(+)-glucose, sodium succinate, and sucrose were selected for the optimization of carbon sources. The nitrogen source concentration was set as 100 mg/L, and ratio of carbon to nitrogen (5, 10, 15, 20, and 25) was controlled by adjusting the concentrations of carbon sources for further optimization. The pH values were adjusted to 5.0, 6.0, 7.0, 8.0, and 9.0 by using HCl (1 mol/L) and NaOH (1 mol/L). Temperature was adjusted to 20 °C, 25 °C, 30 °C, 35 °C, and 40 °C. Shaking speed in DO test was adjusted to 60, 90, 120, 150, and 180 rpm. The salinity was adjusted to 1%, 2%, 3%, 4% and 5%.
To optimize the initial C/N ratio, sodium citrate was selected as the carbon source. Regarding inoculation, the strain HN1 cultured in HNM2 medium was first washed twice with physiological saline to eliminate the impact of residual medium on the results and then inoculated at a concentration of 1% (v/v). All the bacterial strains were incubated at 30 °C and 120 rpm. The concentrations of NH4+-N, NO2−-N, NO3−-N, and OD600 in the 48 h carbon source experiment samples were measured using the method described in Section 2.6.
2.5. Whole Genome Sequencing and Genome Annotation of Strain HN1
The whole genome sequencing was conducted by Majorbio Biotechnology (Shanghai, China), using Illumina and PacBio sequencing techniques to sequence the genome of strain HN1. Gene annotation and functional prediction were completed in the following six databases: NR, Swiss-Prot, Pfam, EggNOG, GO, and KEGG pathway.
2.6. The Concentration Determination of Nitrogen Compounds and Statistical Analysis
The contents of NH4+-N, NO2−-N, NO3−-N, and TN were determined according to the method in former work [15]. In brief, NH4+-N was measured by using Nessler’s reagent photometry at 420 nm. NO2−-N was measured by using N-(1-naphthalene)-diaminoethane spectrophotometry method (540 nm). NO3−-N was determined by using the phenol disulfonic acid method. The alkaline potassium persulfate digestion–ultraviolet spectrophotometric method was performed to measure TN. OD600 was measured using a UV spectrophotometer, which can determine the growth status of the strain. The calculation method for nitrogen balance was proposed based on previous research [16]. Statistical analysis was performed using one-way ANOVA and Tukey’s HSD test in SPSS 23.0 software (p < 0.05). The visualization of data is achieved through the software Origin 2021.
3. Results
3.1. Identification of Strain HN1
The strain with efficient denitrification ability obtained after purification isolation was named HN1. HN1 colonies were obtained on HNM1 plates after 48 h, under 30 °C incubation. Through characterization morphological observation, the obtained colonies were irregular in form, highly raised with smooth surface, pale yellow in color, and slightly transparent. The 16S rRNA gene sequence of strain HN1 had been deposited in GenBank (accession no. OR668337). The highest similarity of the 16S rRNA gene sequence of strain HN1 was 99.05% (Tritonbacter mobilis subsp. pelagius NBRC 102038; accession no. LNWZ01000042). The Neighbor-Joining phylogenetic tree (Figure 1) also showed that the 16S rRNA gene sequence of Tritonibacter mobilis subsp. pelagius strain NBRC 102038 had the closest phylogenetic relationship with that of strain HN1.
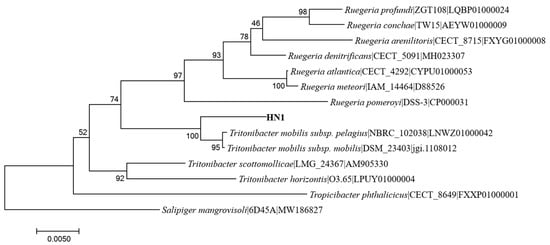
Figure 1.
The phylogenetic tree, derived from partial 16S rRNA gene sequences by Neighbor-Joining analysis.
3.2. The Performance of Nitrogen Removal
This experiment was carried out by using a 3.0% salinity HNM2 medium, and the growth of HN1 and the changes of nitrogenous compounds such as NO3−-N, NO2−-N, NH4+-N, and TN were determined during 96 h. The growth curve and denitrification of strain HN1 are shown in Figure 2. After 12 h of cultivation, the strain HN1 entered the logarithmic phase, and then the OD600 increased from 0.160 to 0.996 within 12 to 48 h. The negative correlation trend between NH4+-N and OD600 indicated a close relationship between the removal of NH4+-N and the growth of HN1. The NH4+-N dropped sharply to 2.38 mg/L at 48 h, and the average removal rate was 4.13 mg/L/h. NO3−-N decreased from 14.68 mg/L to 1.46 mg/L in the meantime. During the whole incubation process, the significant accumulation of NO2−-N was not detected. The changes in TN content were similar to the change rule of NH4+-N. At 96 h, the removal rate of NH4+-N was 98.22%, and the removal rate of TN was 86.63%, of which 42.91% was absorbed by the strain. The strain absorbed 42.91% into intracellular nitrogen, 44.51% of nitrogen became gaseous, and the remaining portion was converted into other forms of nitrogen.
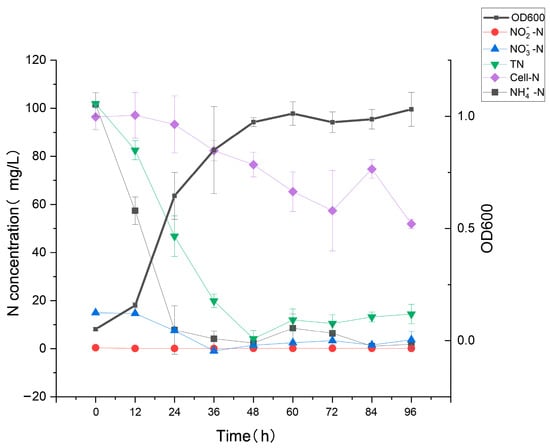
Figure 2.
The concentrations of various nitrogen forms during the growth of strain HN1. The error bars mean the standard deviations.
3.3. The Effect of Various Factors on the Nitrogen Removal over Strain HN1
Carbon sources provide energy for bacteria and serve electrons in the nitrogen removal process. The NH4+-N removal rate of strain HN1 was lower when using glucose compared to other carbon sources (74.79–96.93%) at 72 h. Strain HN1 exhibited excellent NO2−-N (96.70% and 100%, respectively, at 48 h) and NO3−-N (93.60% and 95.71%, respectively, at 72 h) removal efficiencies when sodium citrate and sodium succinate were used as the sole carbon source. In addition, HN1 exhibited a higher nitrogen removal rate with less biomass when sodium citrate served as the sole carbon source. In conclusion, sodium citrate was selected as the optimal carbon source for subsequent experiments (Figure 3).
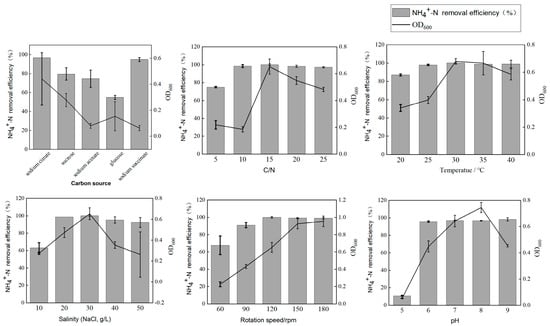
Figure 3.
Growth and NH4+-N removal of strain HN1 under different conditions. The error bars mean the standard deviations.
During the nitrogen removal process, the C/N ratio influenced the efficiency of HN-AD by affecting electron transfer. The removal rate of NH4+-N exceeds 97.62% when the C/N is greater than 10. However, there was no obvious difference between the C/N ratios of 15, 20, and 25. The biomass of the strain varied with the C/N ratio; the biomass of strain HN1 reached the highest value (OD600 = 0.657) when the C/N ratio was 15. Consequently, the optimal C/N ratio of strain HN1 was 15 (Figure 3).
The removal rate of NH4+-N exceeded 92.81% in the salinity range of 2–5%. With the change in salinity, the NH4+-N removal rate showed a rising and then a falling trend. When the salinity was 3%, the NH4+-N removal rate reached its maximum value (99.92%). Meanwhile, the trend of biomass change was also similar to the change in the NH4+-N removal rate. Therefore, the optimal salinity for selecting strain HN1 was 3% (Figure 3).
For temperature, the nitrogen removal efficiency of strain HN1 at 20–40 °C was above 85.69%, and there was little change in nitrogen removal efficiency above 25 °C (97.81–99.92%). The trend of changes in cell biomass was similar to that of the nitrogen removal process. As the temperature increased, the biomass of strain HN1 reached its maximum at 30 °C and then gradually decreased as the temperature increased. After careful consideration, 30 °C was the optimal nitrogen removal temperature for strain HN1 (Figure 3).
This study adjusted DO by changing the rotational speed of the shaking table from 60 rpm to 180 rpm. The NH4+-N removal rate reached 99.92% at 120 rpm. When the rotational speed exceeds 120 rpm, there is no obvious change in the nitrogen removal rate (average 99.35%). At the same time, strain HN1 achieved extremely high biomass without significant differences at 150 rpm and 180 rpm (OD600 = 0.955 and OD600 = 0.955, respectively). To sum up, considering economic benefits, 120 rpm was selected as the optimal rotational speed for nitrogen removal of strain HN1 (Figure 3).
The nitrogen removal efficiency of strain HN1 exceeded 96% within the initial pH range of 6–9 and there is no significant difference. In terms of biomass, the maximum biomass occurred at pH 8 (OD600 = 0.712). Therefore, the optimal initial pH for nitrogen removal efficiency was 8 (Figure 3).
3.4. Genome and Functional Genes Analysis of Strain HN1
The complete genome sequence of HN1 was 4,736,451 bp and was deposited in the GenBank database under the BioProject Number of PRJNA1025341. Among them, GC content accounts for 61.38% (Figure 4). A total of five genomic islands (GIs) and 4239 genes were forecasted in the genome of HN1. In order to thoroughly clarify the metabolic pathway of nitrogen removal in HN1, the coding genes of all key enzymes related to the nitrogen metabolism process were counted; the results are shown in Table 1. For genes encoding key enzymes involved in the HN-AD process, narG, nirS, norB, nosZ, nirB, and nasA genes were annotated in strain HN1 with the denitrification process gene intact. In contrast, the genes encoding ammonia monooxygenase (AMO) and hydroxylamine oxidase (Hao) have not been discovered, and the nitrification process was missing, presumably as a possible novel pathway (Figure 5). Meanwhile, catalyzed by glnA and gltB genes, ammonia was converted to glutamate, which enters the glutamate cycle. Combined with nitrogen balance analysis, half of the external nitrogen source was assimilated into intracellular nitrogen. Therefore, the expression of key genes indicates that there may be two possible nitrogen removal methods for strain HN1: heterotrophic nitrification–aerobic denitrification and nitrogen assimilation.
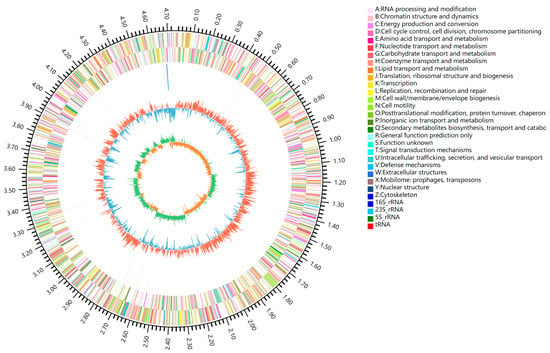
Figure 4.
The circle genomic map of HN1.

Table 1.
Nitrogen metabolism-related genes detected by genomic annotation.
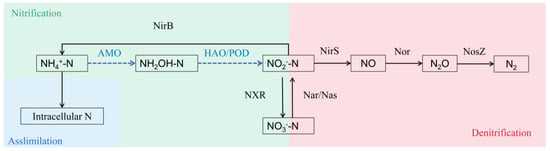
Figure 5.
Prediction of nitrogen metabolism pathway of strain HN1.
4. Discussion
Strain HN1 was identified as a Tritonibacter mobilis bacterium, which currently is the only strain of this species with HN-AD capability. Therefore, it is of significance to understand the N degradation capacity and removal pathway of this new strain.
In this experiment, ammonium salt was the sole nitrogen source to explore HN1 growth and nitrogen utilization. The experimental results showed that the removal rate of NH4+-N was closely related to the growth of HN1, and its average deconditioning is similar to Vibrio diabolicus SF16 (4.50 mg/L/h) [9] but much lower than Pseudomonas mosselii K17 (9.7 mg/L/h) [17]. The ammonium accumulation between 48 and 60 h may be mainly due to the depletion of carbon sources and the lysis of dead cells at the later stage of culture [18,19]. No significant NO2−-N accumulation was detected throughout the entire cultivation process, indicating that strain HN1 exhibits excellent HN-AD performance similar to Pseudomonas tolaasii Y-11 [20]. Changes in TN content indicate that this strain had a certain TN removal ability. The strain HN1 effectively removed 98.22% NH4+-N with a salinity of 3%, which provides a novel bio-tool for nitrogen pollution removal in high-salinity sewage or mariculture wastewater.
Bacteria tend to use carbon sources with simple structures and small molecules. Strain HN1 showed extremely low nitrogen removal efficiency with glucose as the sole carbon source but showed extremely high efficiency when using sodium succinate and sodium citrate. This phenomenon probably occurs because the rate of nitrification is faster when reducing substances such as organic acids are used as substrates, and glucose will inhibit nitrification by increasing the stimulation of cell synthesis. During the process of removing NH4+-N, the biomass of sodium citrate as the sole carbon source was much lower than that of sodium succinate. This might be attributed to the weak anabolism of NH4+-N, and a little content of nitrogen was transformed into intracellular nitrogen. Many strains have shown similar results, such as the optimal carbon source for strain Photobacterium sp. NNA4 was sodium succinate, while the denitrification performance was the lowest when glucose was used as a carbon source [21], and the optimal carbon source for the Halomonas venusta SND-01 strain was sodium citrate [22].
When the C/N ratio is below 5, carbon sources might not provide sufficient electron donors and energy to bacteria, and an excessive C/N ratio could lead to organic pollution [23]. Interestingly, when the C/N ratio was higher than 15, the biomass decreased with the increase in the C/N instead, which might have been due to the lack of raw materials used for synthesizing proteins and nucleic acids, leading to the inability of cell multiplication. Most research results have shown that an excessively high or low C/N ratio can inhibit the nitrogen removal process, such as in strains Vibrio diabolicus SF16 [9] and Pseudomonas sp. DM02 [24]. When the C/N is less than 10, the denitrification efficiency is below 80%. The C/N ratio tolerance range of Bacillus litoralis N13 was wide, with a nitrogen removal efficiency of up to 90% in the C/N ratio range of 5–20 [25].
High salinity can inhibit the aerobic denitrification process and change the morphology of cells [26]. The optimal salinity for nitrogen removal by strain HN1 was 3%, and according to Kushner’s classification, it should belong to moderate halophilic bacteria [27]. Different types of HN-AD bacteria have different levels of salt tolerance, and the optimal salinity range for nitrogen removal by strains Vibrio sp. AB15 [28] and Bacillus litoralis N31 [25] were similar to strain HN1. However, the strain Zobellella B307 has high salt tolerance and can reach NH4+-N removal rates of over 98% at salinity ranges of 5–65 ‰ [10].
A high temperature can promote carbon metabolism processes, thereby generating more energy to indirectly promote the nitrogen removal process. Whether the temperature is too high or too low can affect the enzyme activity during the denitrification process, affecting the denitrification rate. Many HN-AD bacteria previously studied were mesophilic bacteria (with a suitable temperature of 20 °C–40 °C), such as strains Marinobacter sp. F6 [29] and Acinetobacteri sp. JR1 [30]. This study indicates that a low temperature can reduce the biological activity of strain HN1 and inhibit both the growth rate and nitrogen removal rate. At the same time, a high temperature can also inhibit the growth of strain HN1, which may be related to a decrease in the activity of related enzymes. Recently, there have also been many HN-AD strains isolated from extreme environments, such as Bacillus simplex H-b [31], which can achieve a denitrification efficiency of over 90% at 10 °C–35 °C, and Priestiary abhattai KX-3 can achieve denitrification efficiency of 80% at 5 °C–37 °C [32].
Dissolved oxygen (DO) is the more important electron acceptor in aerobic denitrification compared to NO3−-N and has a significant impact on the nitrogen removal process [33]. DO participates in the aerobic respiration and aerobic denitrification process of HN-AD organisms [34], so it is very important for bacterial growth and the nitrogen removal process. From this study, it was found that increasing the rotational speed to a certain extent would increase the NH4+-N removal rate. This might be owing to the increase in rotational speed causing an increase in DO and increasing the contact between bacteria and substrates to increase the reaction rate. However, an excessive increase in rotational speed did not have too much impact. Similar to the results of strain Vibrio sp. LY1024 [35], its denitrification efficiency remained at 98% at speeds greater than 130 rpm. The results of strain Acinetobacter sp. T1 were different from those of this study. The NH4+-N removal rate of strain Acinetobacter sp. T1 reached 83% at 160 rpm after 48 h, but the nitrogen removal rate began to decrease when the speed was greater than 160 rpm [36].
pH is an important factor in the environment, and both nitrification and denitrification processes can cause changes in pH, and extreme pH environments can inhibit bacterial metabolism and cell growth [37]. The result of this study was similar to strain Rhizobium sp.WS7, which has a higher nitrogen removal rate in the pH range of 6–10, with an initial pH of 7 for nitrogen removal [38]. Strain HN1 could be applied to slightly acidic to slightly alkaline water bodies, with a relatively wide range of applications. It is generally believed that most HN-AD bacteria are suitable for growth under neutral or slightly alkaline conditions [39,40]. However, the strain Acinetobacter sp. JR1 can achieve a 90% ammonia removal rate in the pH range of 4.5–10 [30].
The denitrification genes, including narG, nirS, norB, and nosZ, were detected in this study, which could explain the complete denitrification pathway (NO3−-N→NO2−-N→NO→N2O→N2). Previous studies have shown that the reduction of NO2−-N to NO can be catalyzed by two enzymes, the cytochrome cd1 nitrite reductase (cd1-NIR) and Cu-containing nitrite reductase (Cu-NIR) encoding by nirS and nirK genes, respectively. Only the nirS gene was annotated in the HN1 genome sequence, which is similar to the detection results of functional genes in Pseudomonas bauzanensis DN13-1 [41] and Acinetobacter sp. JR1 [30]. This is consistent with a previous study that showed the nirS and nirK genes were usually not possessed by a single strain simultaneously [42]. Contrary to the result of Sphingopyxis sp. CY-10 [43], which possessed both the nirK and nirS genes.
The key genes involved in nitrogen removal have been detected to further confirm the HN-AD capacity of HN1 and establish its nitrogen metabolism pathway. Key genes corresponding to hydroxylamine oxidoreductase (HAO) and ammonia monooxygenase (AMO) were absent from the genome of strain HN1, which is similar to other reported HN-AD strains, such as Stutzerimonas frequens TF18 [44], Acinetobacter oleivorans AHP123 [45], and Halomonas venusta SND-01 [22], resulting in the prevention of the complete nitrification pathway. However, the presence of the nxrAB proved that HN1 could convert nitrite to nitrate. Thus, it is assumed that an unknown enzyme with a similar function to the AMO and HAO enzymes is used by strain HN1 to conduct HN-AD via the nitrification pathway (NH4+→NH2OH→NO2−-N→NO3−-N).
5. Conclusions
The bacteria with HN-AD activity in the salt environment have been the research hot spots due to their potential in nitrogen pollution remediation. In the current work, the HN-AD bacterium Tritonibacter mobilis HN1 was isolated and cultured from mariculture sludge with a high ammonium removal efficiency. The total nitrogen removal rate of strain HN1 reached 86.63%, of which 42.91% was absorbed as intracellular organic nitrogen, and 44.51% was converted into gaseous nitrogen. The rest was lost as other forms of nitrogen at the salinity of 3.0% within 96 h. The total ammonia nitrogen, nitrate, and nitrite removal efficiencies of the strain were 96.93%, 100%, and 95.71%, respectively, with the carbon source being sodium citrate or sodium succinate, a C/N ratio of 15, a salinity of 3%, a temperature of 30 °C, a shaking speed of 120 rpm, and a pH of 8. This study shows that HN1 has a greater potential for degrading nitrogen contaminants in salt environments, such as nitrogen-containing wastewater from the ocean, and it is the first strain in Tritonibacter mobilis to have both heterotrophic nitrification and aerobic denitrification capabilities.
The whole genome analysis showed the presence of key denitrification genes such as nar, nirS, norB, norC, nosZ, glnA, GltB, and GltD in strain HN1 that confirmed methods for heterotrophic nitrification–aerobic denitrification and nitrogen assimilation. In addition, the theory that HN1 may have a different HN-AD pathway than other genes has been demonstrated by the absence of some common HN-AD genes (amo, nap). Nitrogen balance analysis was used to confirm the aerobic denitrification pathway. Therefore, strain HN1 may be capable of nitrogen assimilation and possess a new HN-AD pathway, which provides novel bacterial sources for basic research and industrial production.
Author Contributions
Methodology, R.Q., J.W. (Jingyu Wei), H.X., Y.L. and J.X.; software, R.Q. and Y.Z.; validation, R.Q., Y.Z. and J.W. (Jianxin Wang); formal analysis, R.Q., Y.Z., W.Q. and J.W. (Jianxin Wang); data curation, R.Q. and Y.Z.; writing—original draft preparation, R.Q. and Y.Z.; writing—review and editing, Y.Z., W.Q. and J.W. (Jianxin Wang); visualization, R.Q.; supervision, W.Q. and J.W. (Jianxin Wang); project administration, Y.Z., W.Q., and J.W. (Jianxin Wang); funding acquisition, J.W. (Jianxin Wang). All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Province Key Research and Development Program of Zhejiang (grant number 2021C02047), Science and Technology Bureau Projects of Zhoushan (grant numbers 2022C13048 and 2019C21011), and the National Key Research and Development Program (grant number 2019YFD090305).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are included within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hu, X.; Su, J.; Ali, A.; Wang, Z.; Wu, Z. Heterotrophic nitrification and biomineralization potential of Pseudomonas sp. HXF1 for the simultaneous removal of ammonia nitrogen and fluoride from groundwater. Bioresour. Technol. 2021, 323, 124608. [Google Scholar] [CrossRef] [PubMed]
- Hutchins, D.A.; Capone, D.G. The marine nitrogen cycle: New developments and global change. Nat. Rev. Microbiol. 2022, 20, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Dawn, E.; Holmes, Y.D.; Jessica, A. Smith. Chapter Four—Nitrogen cycling during wastewater treatment. Adv. Appl. Microbiol. 2019, 106, 113–192. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Pajares, S.; Ramos, R. Processes and Microorganisms Involved in the Marine Nitrogen Cycle: Knowledge and Gaps. Front. Mar. Sci. 2019, 6, 739. [Google Scholar] [CrossRef]
- Yu, T.; Li, M.; Niu, M.; Fan, X.; Liang, W.; Wang, F. Difference of nitrogen-cycling microbes between shallow bay and deep-sea sediments in the South China Sea. Appl. Microbiol. Biotechnol. 2018, 102, 447–459. [Google Scholar] [CrossRef]
- Robertson, L.A.; Kuenen, J.G. Thiosphaera pantotropha gen. nov. sp. nov., a facultatively anaerobic, facultatively autotrophic sulphur bacterium. Microbiology 1983, 129, 2847–2855. [Google Scholar] [CrossRef]
- Cui, Y.; Cui, Y.W.; Huang, J.L. A novel halophilic Exiguobacterium mexicanum strain removes nitrogen from saline wastewater via heterotrophic nitrification and aerobic denitrification. Bioresour. Technol. 2021, 333, 125189. [Google Scholar] [CrossRef]
- Duan, J.; Fang, H.; Su, B.; Chen, J.; Lin, J. Characterization of a halophilic heterotrophic nitrification-aerobic denitrification bacterium and its application on treatment of saline wastewater. Bioresour. Technol. 2015, 179, 421–428. [Google Scholar] [CrossRef]
- Xiang, Z.; Chen, X.; Bai, J.; Li, B.; Li, H.; Huang, X. Bioaugmentation performance for moving bed biofilm reactor (MBBR) treating mariculture wastewater by an isolated novel halophilic heterotrophic nitrification aerobic denitrification (HNAD) strain (Zobellella B307). J. Environ. Manag. 2023, 325, 116566. [Google Scholar] [CrossRef]
- Huang, F.; Pan, L.; He, Z.; Zhang, M.; Zhang, M. Heterotrophic nitrification-aerobic denitrification characteristics and antibiotic resistance of two bacterial consortia from Marinomonas and Halomonas with effective nitrogen removal in mariculture wastewater. J. Environ. Manag. 2021, 279, 111786. [Google Scholar] [CrossRef] [PubMed]
- Osborne, C.A.; Galic, M.; Sangwan, P.; Janssen, P.H. PCR-generated artefact from 16S rRNA gene-specific primers. FEMS Microbiol. Lett. 2005, 248, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Li, X.; Fan, W.; Wang, J. Heterotrophic nitrification and aerobic denitrification by a novel Acinetobacter sp. ND7 isolated from municipal activated sludge. Bioresour. Technol. 2020, 301, 122749. [Google Scholar] [CrossRef]
- Yu, X.; Lingguang, H.; Ligang, X. Characteristics of diffuse source N pollution in Lean River catchment. Procedia Environ. Sci. 2011, 10, 2437–2443. [Google Scholar] [CrossRef]
- Dong, L.; Ge, Z.; Qu, W.; Fan, Y.; Dai, Q.; Wang, J. Characteristics and mechanism of heterotrophic nitrification/aerobic denitrification in a novel Halomonas piezotolerans strain. J. Basic. Microbiol. 2022, 62, 124–134. [Google Scholar] [CrossRef]
- Hu, J.; Yang, X.; Deng, X.; Liu, X.; Yu, J.; Chi, R.; Xiao, C. Isolation and Nitrogen Removal Efficiency of the Heterotrophic Nitrifying-Aerobic Denitrifying Strain K17 from a Rare Earth Element Leaching Site. Front. Microbiol. 2022, 13, 905409. [Google Scholar] [CrossRef]
- Guo, L.; Chen, Q.; Fang, F.; Hu, Z.; Wu, J.; Miao, A.; Xiao, L.; Chen, X.; Yang, L. Application potential of a newly isolated indigenous aerobic denitrifier for nitrate and ammonium removal of eutrophic lake water. Bioresour. Technol. 2013, 142, 45–51. [Google Scholar] [CrossRef]
- Ren, Y.-X.; Yang, L.; Liang, X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresour. Technol. 2014, 171, 1–9. [Google Scholar] [CrossRef]
- He, T.; Li, Z.; Sun, Q.; Xu, Y.; Ye, Q. Heterotrophic nitrification and aerobic denitrification by Pseudomonas tolaasii Y-11 without nitrite accumulation during nitrogen conversion. Bioresour. Technol. 2016, 200, 493–499. [Google Scholar] [CrossRef]
- Liu, Y.; Ai, G.M.; Wu, M.R.; Li, S.S.; Miao, L.L.; Liu, Z.P. Photobacterium sp. NNA4, an efficient hydroxylamine-transforming heterotrophic nitrifier/aerobic denitrifier. J. Biosci. Bioeng. 2019, 128, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.Q.; Cui, Y.W.; Yang, H.J.; Xu, M.J.; Cui, Y.; Chen, Z. A halophilic aerobic-heterotrophic strain Halomonas venusta SND-01: Nitrogen removal by ammonium assimilation and heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2023, 374, 128758. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.; Zhou, X.; Arslan, M.; Luo, Z.; Wei, J.; Wu, Z.; Gamal El-Din, M. Heterotrophic nitrification and aerobic denitrification process: Promising but a long way to go in the wastewater treatment. Sci. Total Environ. 2022, 805, 150212. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Zhao, X.; Senbati, Y.; Song, K.; He, X. Nitrogen removal by heterotrophic nitrifying and aerobic denitrifying bacterium Pseudomonas sp. DM02: Removal performance, mechanism and immobilized application for real aquaculture wastewater treatment. Bioresour. Technol. 2021, 322, 124555. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Pan, L.; Lv, N.; Tang, X. Characterization of novel Bacillus strain N31 from mariculture water capable of halophilic heterotrophic nitrification-aerobic denitrification. J. Biosci. Bioeng. 2017, 124, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Gui, M.; Chen, Q.; Ni, J. Effect of NaCl on aerobic denitrification by strain Achromobacter sp. GAD-3. Appl. Microbiol. Biotechnol. 2017, 101, 5139–5147. [Google Scholar] [CrossRef] [PubMed]
- Ventosa, A.; Nieto, J.N.J.; Oren, A. Biology of Moderately Halophilic Aerobic Bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [CrossRef]
- Dou, L.; Chen, W.; Pan, L.; Huang, F. Characterization of Vibrio sp. strain AB15 and Pseudomonas fluorescens strain NB14 from the biofloc of shrimp culture ponds capable of high ammonia and nitrite removal efficiency. J. World Aquac. Soc. 2021, 52, 843–858. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Liu, Y.; Gao, X.Y.; Ai, G.M.; Miao, L.L.; Liu, Z.P. Characterization of a marine origin aerobic nitrifying-denitrifying bacterium. J. Biosci. Bioeng. 2012, 114, 33–37. [Google Scholar] [CrossRef]
- Yang, J.R.; Wang, Y.; Chen, H.; Lyu, Y.K. Ammonium removal characteristics of an acid-resistant bacterium Acinetobacter sp. JR1 from pharmaceutical wastewater capable of heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2019, 274, 56–64. [Google Scholar] [CrossRef]
- Yang, Q.; Yang, T.; Shi, Y.; Xin, Y.; Zhang, L.; Gu, Z.; Li, Y.; Ding, Z.; Shi, G. The nitrogen removal characterization of a cold-adapted bacterium: Bacillus simplex H-b. Bioresour. Technol. 2021, 323, 124554. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Zhao, X.; Song, X.; Wang, D.; Shi, G.; Duan, X.; Chen, X.; Shen, G. Nitrogen removal by a novel strain Priestia aryabhattai KX-3 from East Antarctica under alkaline pH and low-temperature conditions. Process Biochem. 2023, 130, 674–684. [Google Scholar] [CrossRef]
- Patureau, D.; Bernet, N.; Delgenès, J.P.; Moletta, R. Effect of dissolved oxygen and carbon-nitrogen loads on denitrification by an aerobic consortium. Appl. Microbiol. Biotechnol. 2000, 54, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Wu, Y.; Li, J.; Yin, M.; Li, D.; Pei, H.; Chang, S.; Guo, W. Effect of dissolved oxygen on high C/N wastewater treatment in moving bed biofilm reactors based on heterotrophic nitrification and aerobic denitrification: Nitrogen removal performance and potential mechanisms. Bioresour. Technol. 2022, 365, 128147. [Google Scholar] [CrossRef]
- Wang, L.; Fu, Y.; Wang, S.; Ye, F.; Cui, E.; Sun, Q. Domesticating a Halotolerant Bacterium of Vibrio sp. LY1024 with Heterotrophic Nitrification–Aerobic Denitrification Property for Efficient Nitrogen Removal in Mariculture Wastewater Treatment. Coatings 2022, 12, 1786. [Google Scholar] [CrossRef]
- Chen, S.; He, S.; Wu, C.; Du, D. Characteristics of heterotrophic nitrification and aerobic denitrification bacterium Acinetobacter sp. T1 and its application for pig farm wastewater treatment. J. Biosci. Bioeng. 2019, 127, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Huang, J.J.; Hua, B.; Champagne, P. Nitrogen removal bacterial strains, MSNA-1 and MSD4, with wide ranges of salinity and pH resistances. Bioresour. Technol. 2020, 310, 123309. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Luo, X.; Ma, W.; Lv, P. Biological nitrogen removal and metabolic characteristics of a novel cold-resistant heterotrophic nitrification and aerobic denitrification Rhizobium sp. WS7. Bioresour. Technol. 2022, 362, 127756. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, W.; Zhu, S.; Liu, F.; Qin, L.; Xu, C.; Wang, Z. Biological nitrogen and phosphorus removal by a phosphorus-accumulating bacteria Acinetobacter sp. strain C-13 with the ability of heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2021, 322, 124507. [Google Scholar] [CrossRef]
- Lou, L.; Luo, H.; Fang, J.; Liu, G. The advance of heterotrophic nitrification aerobic denitrification microorganisms in wastewater treatment. Bioresour. Technol. Rep. 2023, 22, 101495. [Google Scholar] [CrossRef]
- Zhang, M.; Li, A.; Yao, Q.; Wu, Q.; Zhu, H. Nitrogen removal characteristics of a versatile heterotrophic nitrifying-aerobic denitrifying bacterium, Pseudomonas bauzanensis DN13-1, isolated from deep-sea sediment. Bioresour. Technol. 2019, 305, 122626. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Feng, L.; Pi, S.; Cui, D.; Ma, F.; Zhao, H.-P.; Li, A. A critical review of aerobic denitrification: Insights into the intracellular electron transfer. Sci. Total Environ. 2020, 731, 139080. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhang, F.; Zhang, L.; Liu, H.; Zhang, Q.; Xing, Z.; Zhao, T. Characterization of a novel salt-tolerant strain Sphingopyxis sp. CY-10 capable of heterotrophic nitrification and aerobic denitrification. Bioresour. Technol. 2022, 358, 127353. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wang, B.; Liu, M.; Jiang, K.; Wang, L. Stutzerimonas frequens strain TF18 with superior heterotrophic nitrification-aerobic denitrification ability for the treatment of aquaculture effluent. Process Biochem. 2023, 130, 156–165. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y.; Tan, X.; Sheng, Y.; Li, Y.; Zhang, Q.; Xu, J.; Shi, Z. Genomics and nitrogen metabolic characteristics of a novel heterotrophic nitrifying-aerobic denitrifying bacterium Acinetobacter oleivorans AHP123. Bioresour. Technol. 2023, 375, 128822. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).