
The Endogenous Metabolic Response of Tribolium castaneum under a High Concentration of CO2

Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. CO2 Treatments
2.3. RNA Extraction and cDNA Synthesis
2.4. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.5. Determination of Carbohydrate Content and Enzyme Activity
2.6. Statistical Analyses
3. Results
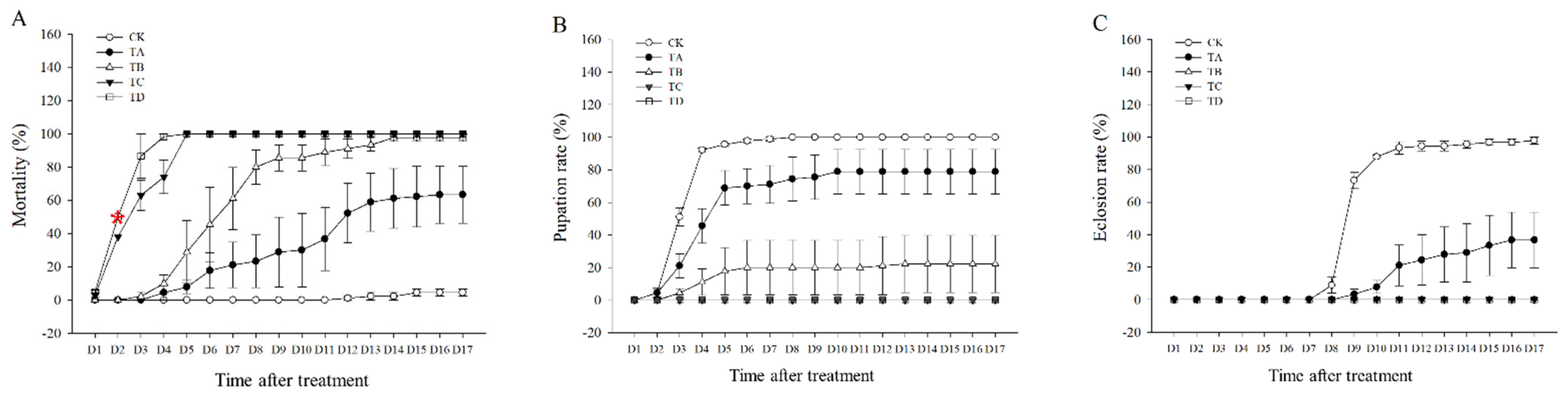
3.1. Mortality, Pupation Rate, and Eclosion Rate of T. castaneum under Different CO2 Concentrations
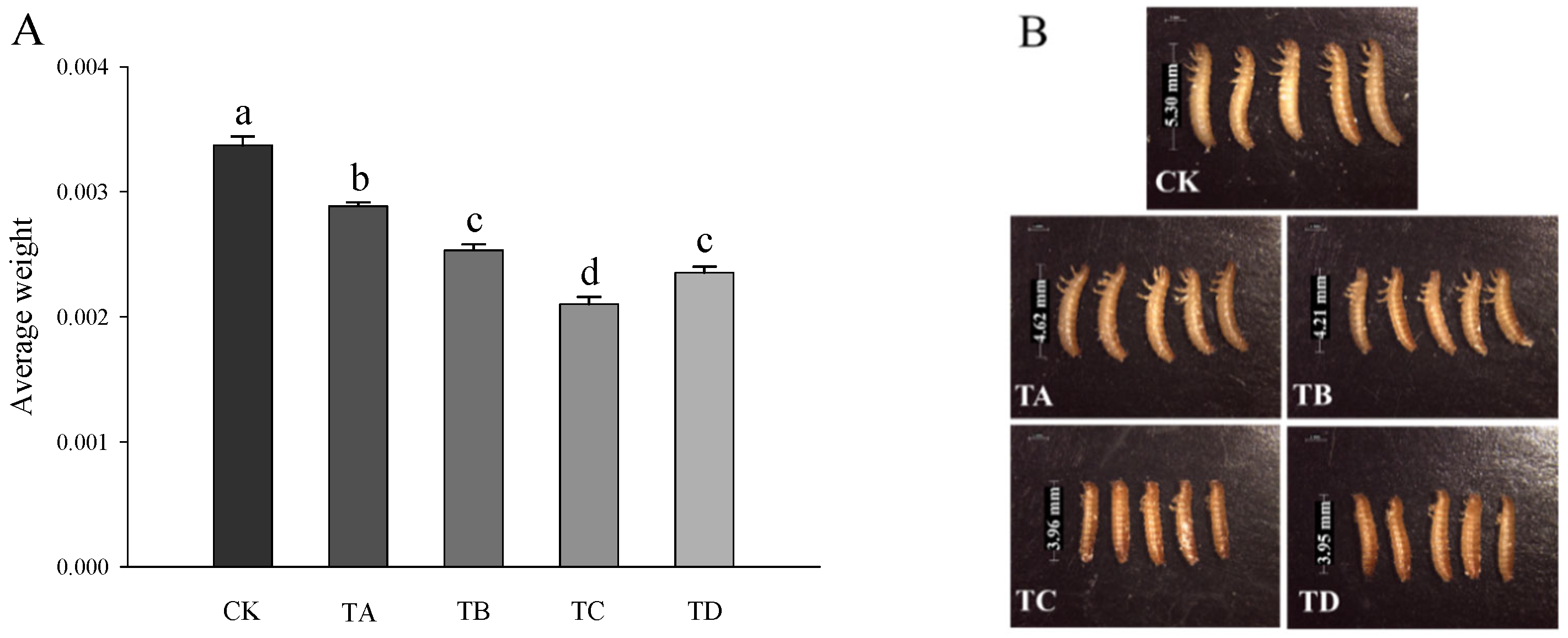
3.2. Changes in Body Weight and Morphological Features of T. castaneum under Different Concentrations of CO2
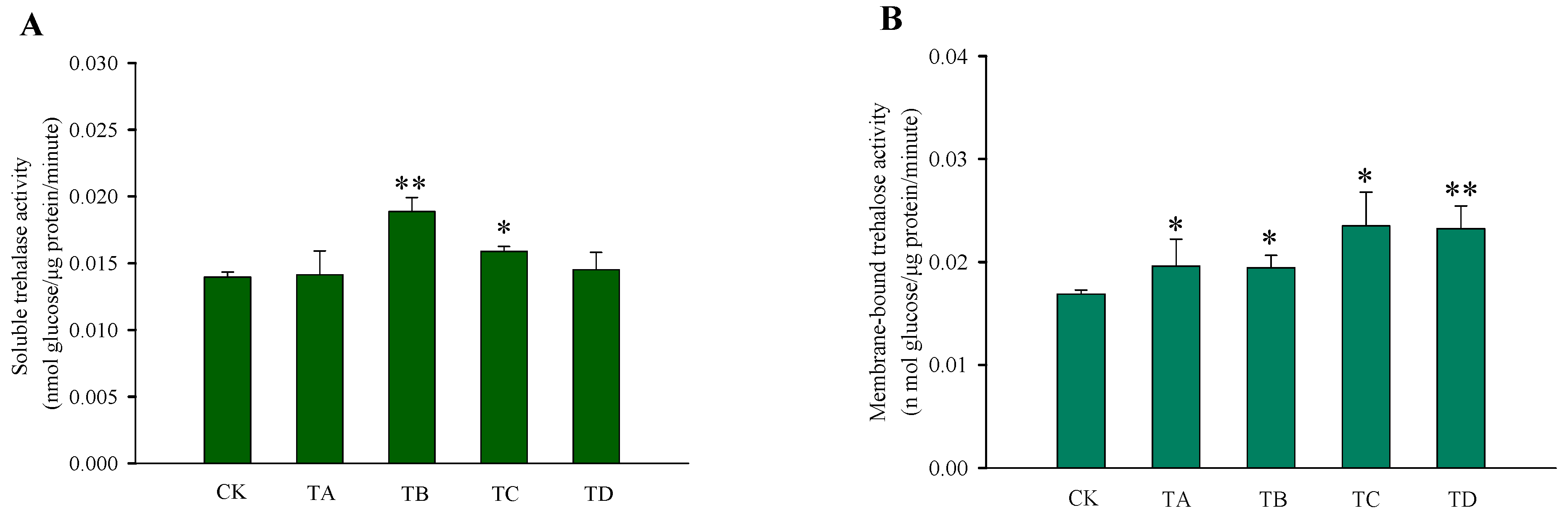
3.3. Changes in Carbohydrate Content and Enzymatic Activity in T. castaneum under Different Concentrations of CO2
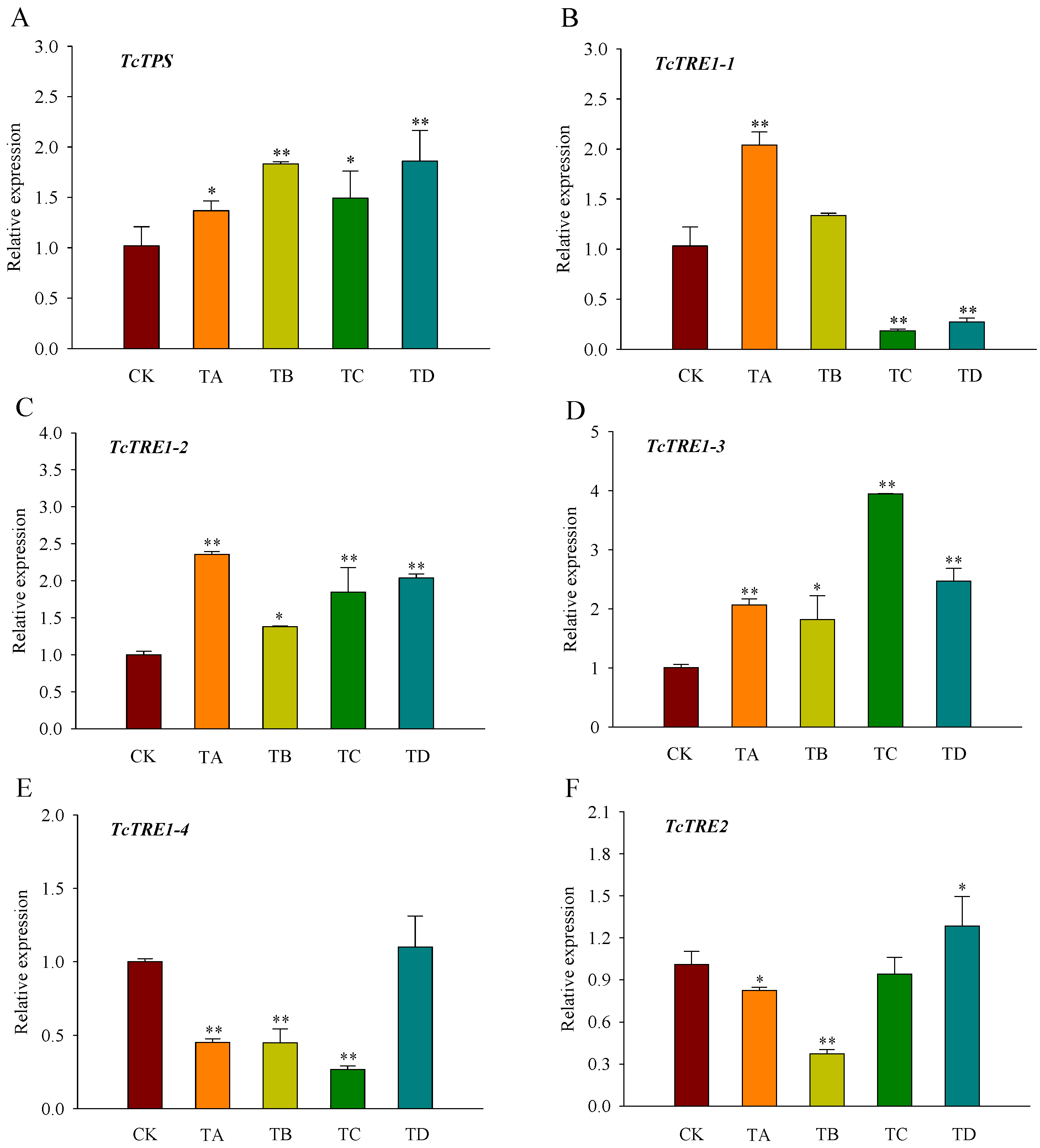
3.4. Expression of Trehalose Metabolism Pathway Genes in T. castaneum under Different CO2 Concentrations
3.5. Gene Expression of Insulin Signaling Pathway Genes in T. castaneum under Different Concentrations of CO2
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mehmood, K.; Husain, M.; Aslam, M.; Ahmedani, M.S.; Aulahk, A.M.; Shaheen, F.A. Changes in the nutritional composition of maize flour due to Tribolium castaneum infestation and application of carbon dioxide to manage this pest. Environ. Sci. Pollut. Res. 2018, 25, 18540–18547. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Lyu, Z.H.; Wang, C.Y.; Cheng, J.; Lin, T. RNA interference of a trehalose-6-phosphate synthase gene reveals its roles in the biosynthesis of chitin and lipids in Heortia vitessoides (Lepidoptera: Crambidae). Insect Sci. 2018, 27, 212–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, G.G. Field estimates of population growth rates of Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.) (Coleoptera: Tenebrionidae and Bostrychidae) in bulk wheat. J. Stored Prod. Res. 1988, 24, 13–22. [Google Scholar] [CrossRef]
- Lis, Ł.B.; Bakuła, T.; Baranowski, M.; Czarnewicz, A. The carcinogenic effects of benzoquinones produced by the flour beetle. Pol. J. Vet. Sci. 2011, 14, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubert, J.; Stejskal, V.; Athanassiou, C.G.; Throne, J.E. Health Hazards Associated with Arthropod Infestation of Stored Products. Annu. Rev. Entomol. 2018, 63, 553–573. [Google Scholar] [CrossRef]
- Zettler, L.J.; Cuperus, G.W. Pesticide resistance in Tribolium castaneum (Coleoptera: Tenebrionidae) and Rhyzopertha dominica (Coleoptera: Bostrichidae) in wheat. J. Econ. Entomol. 1990, 83, 1677–1681. [Google Scholar] [CrossRef]
- Lee, B.H.; Lee, S.E.; Annis, P.C.; Pratt, S.J.; Park, B.S.; Tumaalii, F. Fumigant toxicity of essential oils and monoterpenes against the red flour beetle, Tribolium castaneum Herbst. J. Asia-Pac. Entomol. 2002, 5, 237–240. [Google Scholar] [CrossRef]
- Epidi, T.E.; Odili, E.O. Biocidal activity of selected plant powders against Tribolium castaneum Herbst in stored groundnut (Arachis hypogaea L.). Afr. J. Environ. Sci. Technol. 2009, 3, 1–5. [Google Scholar] [CrossRef]
- Stejskal, V.; Vendl, T.; Aulicky, R.; Athanassiou, C. Synthetic and Natural Insecticides: Gas, Liquid, Gel and Solid Formulations for Stored-Product and Food-Industry Pest Control. Insects 2021, 12, 590. [Google Scholar] [CrossRef] [PubMed]
- Stejskal, V.; Vendl, T.; Li, Z.; Aulicky, R. Minimal Thermal Requirements for Development and Activity of Stored Product and Food Industry Pests (Acari, Coleoptera, Lepidoptera, Psocoptera, Diptera and Blattodea): A Review. Insects 2019, 10, 149. [Google Scholar] [CrossRef] [Green Version]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.A.; Faroni, L.R.; Silva, F.H.; Batista, M.D.; Guedes, R.N. Spread of phosphine resistance among Brazilian populations of three species of stored product insects. Neotrop. Èntomol. 2010, 39, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkin, L.C.; Oppert, B. Gene expression in Tribolium castaneum life stages: Identifying a species-specific target for pest control applications. PeerJ 2019, 7, e6946. [Google Scholar] [CrossRef] [Green Version]
- Davyt-Colo, B.; Girotti, J.R.; González, A.; Pedrini, N. Secretion and Detection of Defensive Compounds by the Red Flour Beetle Tribolium castaneum Interacting with the Insect Pathogenic Fungus Beauveria bassiana. Pathogens 2022, 11, 487. [Google Scholar] [CrossRef]
- Joop, G.; Vilcinskas, A. Coevolution of parasitic fungi and insect hosts. Zoology 2016, 119, 350–358. [Google Scholar] [CrossRef]
- Cao, Y.; Xu, K.K.; Zhu, X.; Bai, Y.; Yang, W.; Li, C. Role of Modified Atmosphere in Pest Control and Mechanism of Its Effect on Insects. Front. Physiol. 2019, 10, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunasekaran, N.; Rajendran, S. Toxicity of carbon dioxide to drugstore beetle Stegobium paniceum and cigarette beetle Lasioderma serricorne. J. Stored Prod. Res. 2005, 41, 283–294. [Google Scholar] [CrossRef]
- Wong, F.J.; Castane, C.; Riudavets, J. Lethal effects of CO2-modified atmospheres for the control of three Bruchidae species. J. Stored Prod. Res. 2013, 55, 62–67. [Google Scholar] [CrossRef]
- Helenius, I.T.; Krupinski, T.; Turnbull, D.W.; Gruenbaum, Y.; Silverman, N.; Johnson, E.A.; Sporn, P.H.S.; Sznajder, J.I.; Beitel, G.J. Elevated CO2 suppresses specific Drosophila innate immune responses and resistance to bacterial infection. Proc. Natl. Acad. Sci. USA 2009, 106, 18710–18715. [Google Scholar] [CrossRef] [Green Version]
- Bretscher, A.J.; Busch, K.E.; de Bono, M. A carbon dioxide avoidance behavior is integrated with responses to ambient oxygen and food in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2008, 105, 8044–8049. [Google Scholar] [CrossRef] [Green Version]
- Hallem, E.A.; Sternberg, P.W. Acute carbon dioxide avoidance in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2008, 105, 8038–8043. [Google Scholar] [CrossRef] [Green Version]
- Beall, C.M. Two routes to functional adaptation: Tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. USA 2007, 104, 8655–8660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCue, M.D.; De Los Santos, R. Upper thermal limits of insects are not the result of insufficient oxygen delivery. Physiol. Biochem. Zool. 2013, 86, 257–265. [Google Scholar] [CrossRef]
- Harrison, J.F.; Haddad, G.G. Effects of oxygen on growth and size: Synthesis of molecular, organismal, and evolutionary studies with Drosophila melanogaster. Annu Rev. Physiol. 2011, 73, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Wang, L.; Qiu, J.; Liu, Z.; Geng, X. Comparative metabolomics analysis of Callosobruchus chinensis larvae under hypoxia, hypoxia/hypercapnia and normoxia. Pest Manag. Sci. 2017, 73, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Friedlander, A.; Navarro, S. Effect of controlled atmospheres on the sorbitol pathway in Ephestia cautella (Walker) pupae. Experientia 1983, 39, 744–746. [Google Scholar] [CrossRef]
- Feron, O. Pyruvate into lactate and back: From the Warburg effect to symbiotic energy fuel exchange in cancer cells. Radiother Oncol. 2009, 92, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Boardman, L.; Sørensen, J.G.; Johnson, S.A.; Terblanche, J.S. Interactions between Controlled Atmospheres and Low Temperature Tolerance: A Review of Biochemical Mechanisms. Front. Physiol. 2011, 2, 92. [Google Scholar] [CrossRef] [Green Version]
- Badre, N.H.; Martin, M.E.; Cooper, R.L. The physiological and behavioral effects of carbon dioxide on Drosophila melanogaster larvae. Comp. Biochem. Physiol. A-Mol. Integr. Physiol. 2005, 140, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Navarro, S. Modified Atmospheres for the Control of Stored-Product Insects and Mites. In Insect Management for Food Storage and Processing; Department of Food Science, Agricultural Research Organization: Bet Dagan, Israel, 2006; pp. 105–146. [Google Scholar] [CrossRef]
- Tunc, I.; Navarro, S. Sensitivity of Tribolium castaneum eggs to modified atmospheres. Entomol. Exp. Appl. 1983, 34, 221–226. [Google Scholar] [CrossRef]
- Kharel, K.; Mason, L.J.; Murdock, L.L.; Baributsa, D. Efficacy of Hypoxia Against Tribolium castaneum (Coleoptera: Tenebrionidae) Throughout Ontogeny. J. Econ. Entomol. 2019, 112, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.Y.; Xu, K.K.; Luo, Y.J.; Wang, Y.Y.; Zhou, M.; Li, C.; Tang, B. The sequence characteristics and functions on regulating trehalose metabolism of two PTP genes in Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2020, 89, 101692. [Google Scholar] [CrossRef]
- Lord, J.C.; Hartzer, K.; Toutges, M.; Oppert, B. Evaluation of quantitative PCR reference genes for gene expression studies in Tribolium castaneum after fungal challenge. J. Microbiol. Methods 2010, 80, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qiu, L.Y.; Yang, H.L.; Wang, H.J.; Zhou, M.; Wang, S.G.; Tang, B. Study on the Effect of Wing Bud Chitin Metabolism and Its Developmental Network Genes in the Brown Planthopper, Nilaparvata lugens, by Knockdown of TRE Gene. Front. Physiol. 2017, 8, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoback, W.W.; Stanley, D.W. Insects in hypoxia. J. Insect Physiol. 2001, 47, 533–542. [Google Scholar] [CrossRef]
- Kaiser, A.; Klok, C.J.; Socha, J.J.; Lee, W.K.; Quinlan, M.C.; Harrison, J.F. Increase in tracheal investment with beetle size supports hypothesis of oxygen limitation on insect gigantism. Proc. Natl. Acad. Sci. USA 2007, 104, 13198–13203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; Lei, J.; Ahn, J.E.; Wang, Y.; Lei, C.; Zhu-Salzman, K. CO2 enhances effects of hypoxia on mortality, development, and gene expression in cowpea bruchid, Callosobruchus maculatus. J. Insect Physiol. 2013, 59, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Husain, M.; Sukirno, S.; Mehmood, K.; Tufail, M.; Rasool, K.G.; Alwaneen, W.S.; Aldawood, A.S. Effectiveness of carbon dioxide against different developmental stages of Cadra cautella and Tribolium castaneum. Environ. Sci. Pollut Res. Int. 2017, 24, 12787–12795. [Google Scholar] [CrossRef] [PubMed]
- Bourne-Murrieta, L.R.; Iturralde-García, R.D.; Wong-Corral, F.J.; Castañé, C.; Riudavets, J. Effect of packaging chickpeas with CO2 modified atmospheres on mortality of Callosobruchus chinensis (Coleoptera: Chrysomelidae). J. Stored Prod. Res. 2021, 94, 101894. [Google Scholar] [CrossRef]
- Nystul, T.G.; Roth, M.B. Carbon monoxide-induced suspended animation protects against hypoxic damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2004, 101, 9133–9136. [Google Scholar] [CrossRef] [Green Version]
- Laere, A.V. Trehalose, reserve and/or stress metabolite? FEMS Microbiol. Lett. 1989, 63, 201–209. [Google Scholar] [CrossRef]
- Crowe, J.H. Trehalose as a “chemical chaperone”: Fact and fantasy. In Molecular Aspects of the Stress Response: Chaperones, Membranes and Networks; Csermely, P., Vígh, L., Eds.; Springer: New York, NY, USA, 2007; Volume 594, pp. 143–158. [Google Scholar]
- Xu, J.; Sheng, Z.; Palli, S.R. Juvenile hormone and insulin regulate trehalose homeostasis in the red flour beetle, Tribolium castaneum. PLoS Genet. 2013, 9, e1003535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Xu, Q.; Zou, Q.; Fang, Q.; Wang, S.; Ye, G. Sequencing and characterization of glycogen synthase and glycogen phosphorylase genes from Spodoptera exigua and analysis of their function in starvation and excessive sugar intake. Arch. Insect Biochem. Physiol. 2012, 80, 42–62. [Google Scholar] [CrossRef]
- Becker, A.; Schlöder, P.; Steele, J.E.; Wegener, G. The regulation of trehalose metabolism in insects. Experientia 1996, 52, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ma, E.; Behar, K.L.; Xu, T.; Haddad, G.G. Role of trehalose phosphate synthase in anoxia tolerance and development in Drosophila melanogaster. J. Biol. Chem. 2002, 277, 3274–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizcano, J.M.; Alessi, D.R. The insulin signalling pathway. Curr. Biol. 2002, 12, R236–R238. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.; Liu, G.; Hou, L.; Gui, W.; Chen, B.; Kang, L. Genetic variation in PTPN1 contributes to metabolic adaptation to high-altitude hypoxia in Tibetan migratory locusts. Nat. Commun. 2018, 9, 4991. [Google Scholar] [CrossRef]
- Han, B.; Zhang, T.; Feng, Y.; Liu, X.; Zhang, L.; Chen, H.; Zeng, F.; Wang, M.; Liu, C.; Li, Y.; et al. Two insulin receptors coordinate oogenesis and oviposition via two pathways in the green lacewing, Chrysopa pallens. J. Insect Physiol. 2020, 123, 104049. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
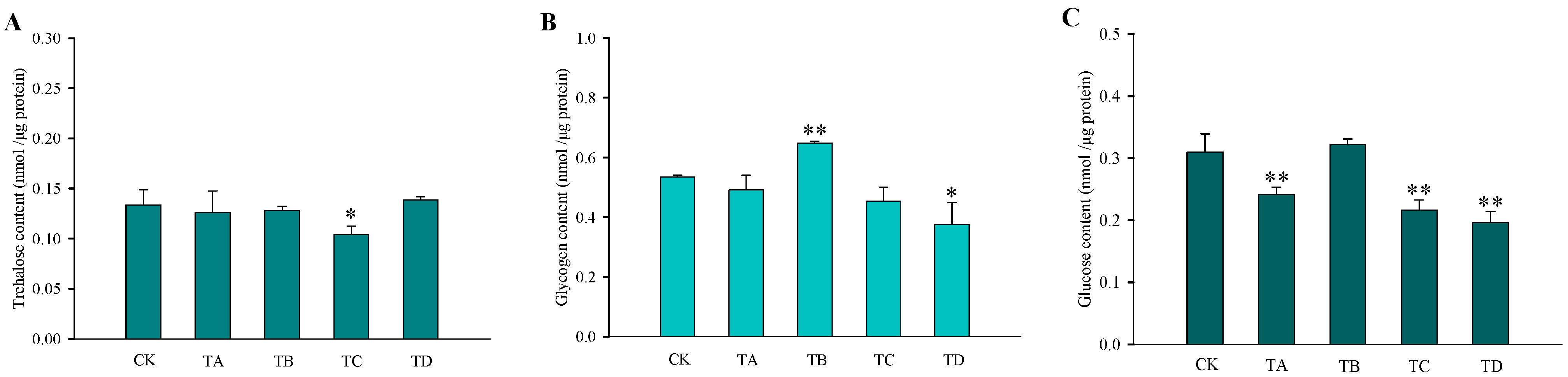{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Number | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Length (bp) |
|---|---|---|---|---|
| QTcRPL13a [34] | XM_969211 | ACCATATGACCGCAGGAAAC | GGAAACCTTGTTACGACTT | |
| QTcTre1-1 | XM_968798 | AACGACTCGCAATGGCTGG | CGGAGGCGTAGTGGAATAGAG | 127 |
| QTcTre1-2 | XM 968883 | GTGCCCAATGGGTTTATCG | CAACCACAACACTTCCTTCG | 261 |
| QTcTre1-3 | XM_968859 | CCTCTCATTCGTCACAAGCG | AAGCGTTTGATTTCTTTGCG | 205 |
| QTcTre1-4 | XM_968826 | ACGGTGCCCGCATCTACTA | GTGTAGGTGGTCCCGTTCTTG | 187 |
| QTcTre2 | EFA11183 | CTCAGCCTGGCCCTTAGTTG | GGAGTCCTCGTAGATGCGTT | 120 |
| QTcTPS | XM_970683 | CGATTCGTACTACAACGGCTGC | GTGGTGTAGCATTGCCAGTGC | 105 |
| QtcInR1 | KF922648.1 | CGGCCGTCTAACACTTACGT | GCAGGGTCATACACTTCGGC | 601 |
| QtcInR2 | KF922649.1 | GGGCTGCAAACAGTTGTGTT | AAGTGCCGTTCAGCATCGTC | 287 |
| QTcIlp1 | EFA02918.1 | TTACGTCTGGTCTTCACCGCACAT | TGGTTGGGTTTGGATTCGGAGAGT | 97 |
| QTcIlp2 | EFA02796.2 | TGGCCGGAATACACACTTGTAGGA | TCTTCTTCCGCAGTAGACCGCTTT | 152 |
| QTcIlp3 | EEZ99258.2 | AAAGTCTGCTTCACCTTGCTCCTC | AATAGCGCACAGTTCGGTGAGAGT | 143 |
| QTcIlp4 | KYB29166.1 | TCGGAGAAGCGACCAAAGCTACAA | TCTGAAGTGATAGAAGCGCCAGCA | 131 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Zhang, X.; Pan, B.; Zhu, J.; Qian, X.; Li, X.; Xu, K.; Tang, B.; Li, C. The Endogenous Metabolic Response of Tribolium castaneum under a High Concentration of CO2. Agriculture 2022, 12, 979. https://doi.org/10.3390/agriculture12070979
Zhou M, Zhang X, Pan B, Zhu J, Qian X, Li X, Xu K, Tang B, Li C. The Endogenous Metabolic Response of Tribolium castaneum under a High Concentration of CO2. Agriculture. 2022; 12(7):979. https://doi.org/10.3390/agriculture12070979
Chicago/Turabian StyleZhou, Min, Xinyu Zhang, Biying Pan, Jiaqi Zhu, Xiaoxiao Qian, Xian Li, Kangkang Xu, Bin Tang, and Can Li. 2022. "The Endogenous Metabolic Response of Tribolium castaneum under a High Concentration of CO2" Agriculture 12, no. 7: 979. https://doi.org/10.3390/agriculture12070979
APA StyleZhou, M., Zhang, X., Pan, B., Zhu, J., Qian, X., Li, X., Xu, K., Tang, B., & Li, C. (2022). The Endogenous Metabolic Response of Tribolium castaneum under a High Concentration of CO2. Agriculture, 12(7), 979. https://doi.org/10.3390/agriculture12070979