
Arbuscular Mycorrhizal Fungi Inoculation Improves Flower Yield and Postharvest Quality Component of Gerbera Grown under Different Salinity Levels



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and AMF Inoculation
2.2. Treatments
2.3. Measurements
2.4. Statistical Analysis
3. Results and Discussion
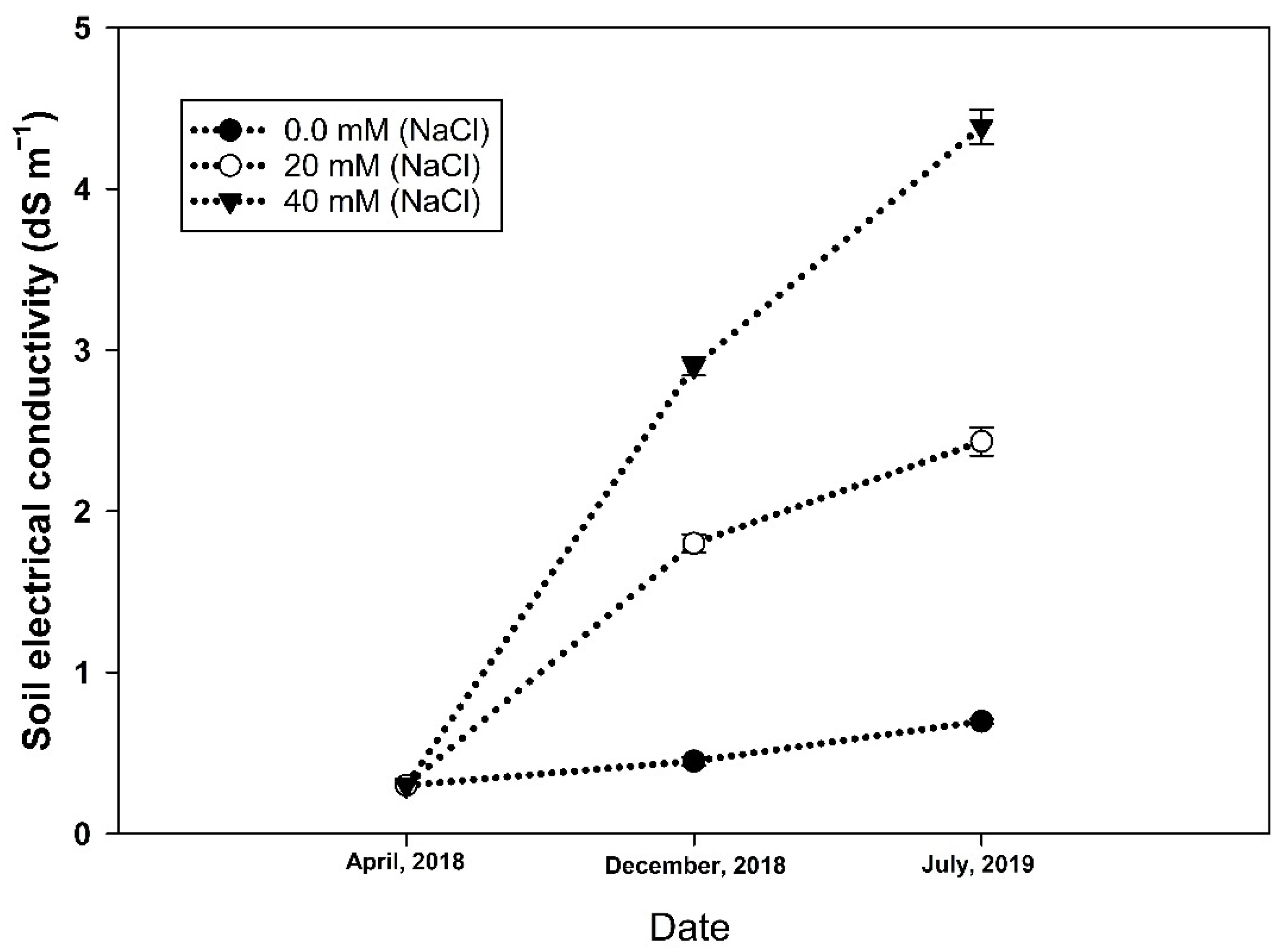
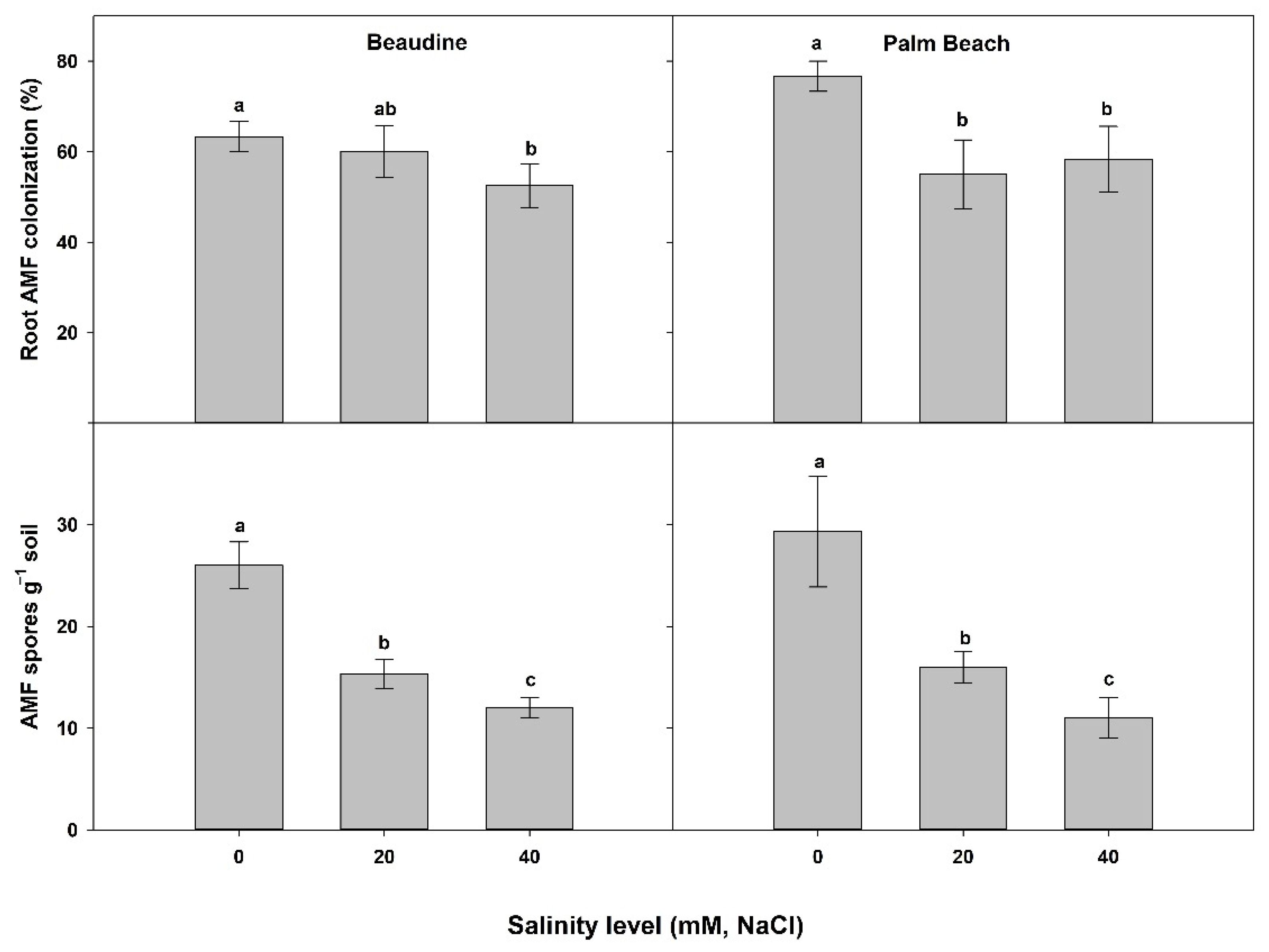
3.1. Salinity Increment, Root Colonization, and Sporulation
3.2. Leaf Physiology
3.3. Flower Yield and Quality
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Naqa, A.; Al-Shayeb, A. Groundwater protection and management strategy in Jordan. Water Resour. Manag. 2009, 23, 2379–2394. [Google Scholar] [CrossRef]
- Al-Karaki, G.; Al-Ajmi, A.; Othman, Y. Response of soilless grown sweet pepper cultivars to salinity. Acta Hortic. 2009, 807, 227–232. [Google Scholar] [CrossRef]
- Al-Karaki, G. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic. 2006, 109, 1–7. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mineral nutrition, toxic element accumulation and water relations of arbuscular mycorrhizal plants. In Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 145–148. [Google Scholar]
- Hack, C.; Porta, M.; Schäufele, R.; Rimoldi, A. Arbuscular mycorrhiza mediated effects on growth, mineral nutrition and biological nitrogen fixation of Melilotus alba Med. in a subtropical grassland soil. Appl. Soil Ecol. 2019, 134, 38–44. [Google Scholar] [CrossRef]
- Clark, R.B. Arbuscular mycorrhizal adaptation, spore germination, root colonization, and host plant growth and mineral acquisition at Low pH. Plant Soil 1997, 192, 15–22. [Google Scholar] [CrossRef]
- Al-Karaki, G.; Clark, R.B. Growth mineral acquistion, and water use by mycorrhizal wheat grown under water stress. J. Plant Nutr. 1998, 21, 263–276. [Google Scholar] [CrossRef]
- Scrase, F.; Sinclair, F.; Farrar, J.; Pavinato, P.; Jones, D. Mycorrhizas improve the absorption of non-available phosphorus by the green manure Tithonia diversifolia in poor soils. Rhizosphere 2019, 9, 27–33. [Google Scholar] [CrossRef]
- Berta, G.; Fusconi, A.; Trotta, A. VA mycorrhizal infection and the morphology of root systems. Environ. Exp. Bot. 1993, 33, 159–173. [Google Scholar] [CrossRef]
- Dugassa-Gobena, D.; Von Allen, H.; Schönbeck, F. Effect of arbuscular mycorrhiza (AM) on health of Linum usitatissimum L. infected by fungal pathogen. Plant Soil 1996, 185, 173–182. [Google Scholar] [CrossRef]
- Al-Raddad, A. Response of beans, broad bean and chickpea plant to inoculation with Glomus species. Sci. Hortic. 1991, 46, 195–200. [Google Scholar] [CrossRef]
- Hooker, J.E.; Munro, M.; Atkinson, D. Vesicular arbuscular fungi induced alteration in poplar root system morphology. Plant Soil 1992, 145, 207–214. [Google Scholar] [CrossRef]
- Othman, Y.; Al-Ajlouni, M.; A’saf, T.; Sawalha, H.; Bany Hani, M. Influence of gibberellic acid on the physiology and flower quality of gerbera and lily cut flowers. Int. J. Agric. Nat. Resour. 2021, 48, 21–33. [Google Scholar] [CrossRef]
- RFH. (2021): Royal FloraHolland Annual-Report-2021. Available online: https://jaarverslag.royalfloraholland.com/wp-content/uploads/2022/03/RFH-Annual-report-2021.pdf (accessed on 6 June 2022).
- Akat Saraçoğlu, Ö.; Meriç, M.K.; Tüzel, İ.H.; Kukul Kurttaş, Y.S. Responses of Gerbera jamesonii plants to different salinity levels and leaching ratios when grown in soilless culture. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019; pp. 357–379. [Google Scholar] [CrossRef]
- Al-Karaki, G.; Abu-Qubah, N.; Othman, Y. Influence of mycorrhizal fungi and water stress on growth and yield of two onion cultivars. Arab. Gulf J. Sci. Res. 2006, 24, 206–214. [Google Scholar]
- Hajiboland, R. Role of Arbuscular Mycorrhiza in Amelioration of Salinity. In Salt Stress in Plants; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Tahat, M.; Alananbeh, K.; Othman, Y.; Leskovar, D. Soil health and sustainable agriculture. Sustainability 2020, 12, 4859. [Google Scholar] [CrossRef]
- Saini, I.; Yadav, K.; Aggarwal, A. Response of arbuscular mycorrhizal fungi along with Trichoderma viride and Pseudomonas fluorescens on the growth, biochemical attributes and vase life of Chrysanthemum indicum. J. Environ. Biol. 2019, 40, 183–191. [Google Scholar] [CrossRef]
- Daft, M.; Okusanya, B. Effect of endogone mycorrhiza on plant growth. New Phytol. 1973, 72, 1333–1339. [Google Scholar] [CrossRef]
- Deljou, N.; Marouf, A.; Hamedan, J. Effect of inoculation with arbuscular mycorrhizal fungi (AMF) on gerbera cut flower (Gerbera jamesonii) production in soilless cultivation. Acta Hortic. 2014, 1034, 417–422. [Google Scholar] [CrossRef]
- Garmendia, I.; Mangas, V. Application of arbuscular mycorrhizal fungi on the production of cut flower roses under commercial-like conditions. Span. J. Agric. Res. 2012, 10, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Tahat, M.; Abo-Farag, K.; Alananbeh, K.; Al-Momany, A. The efficacy of Glomus mosseae and olive cake to control a chili pepper (Capsicum annuum) damping off disease. Fresenius Environ. Bull. 2020, 29, 9863–9871. [Google Scholar]
- Tahat, M.M.; Kamaruzaman, S.; Alananbeh, K.M. Exploring the use of legumes as host plant species in Glomus mosseae sporulation. Legume Res. 2018, 41, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Leskovar, D.; Othman, Y. Direct Seeding and transplanting influence root dynamics, morpho-physiology, yield, and head quality of globe artichoke. Plants 2021, 10, 899. [Google Scholar] [CrossRef]
- Phillips, J.; Hayman, D.S. Improved procedure for clearing roots and staining parasitic and vesicular mycorrizal fungi for rapid assessment of infection. Trans. Brit. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques to measure vesicular-arbuscular infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Pacioni, G. Wet-sieving and decanting technique for spores extraction. In Methods in Microbiology; Varma, A., Trappe, J., Fortin, J., Norris, J., Eds.; Academic Press: London, UK, 1991; pp. 317–322. [Google Scholar]
- Wu, Q.; Srivastava, A.; Zou, Y.; Malhotra, S. Mycorrhizas in citrus: Beyond soil fertility and plant nutrition. Indian J. Agric. Sci. 2017, 87, 427–443. [Google Scholar]
- Zou, Y.; Wu, Q. Sodium chloride stress induced changes in leaf osmotic adjustment of trifoliate orange (Poncirus trifoliata) seedlings inoculated with mycorrhizal fungi. Not. Bot. Horti Agrobot. 2011, 39, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Khalil, H.; Eissa, A.; El-Shazly, S.; Aboul-Nasr, A. Improved growth of salinity-stressed citrus after inoculation with mycorrhizal fungi. Sci. Hortic. 2011, 130, 624–632. [Google Scholar] [CrossRef]
- Mcgee, P.A.; Pattinson, G.S.; Heath, R.A.; Newman, C.A.; Allen, S.J. Survival of propagules of arbuscular mycorrhizal fungi in soils in eastern Australia used to grow cotton. New Phytol. 1997, 135, 773–780. [Google Scholar] [CrossRef]
- Saggin-Júnior, O.; Siqueira, J.O. Micorrizas arbusculares em cafeeiro. In Avanços em Fundamentos e Aplicação de Micorrizas; Siqueira, J.O., Ed.; UFLA-DCS e DCF: Lavras, Brasil, 1996; pp. 203–254. [Google Scholar]
- Wen, Y.; Su, S.; Ma, L.; Wang, X. Effects of gibberellic acid on photosynthesis and endogenous hormones of Camellia oleifera Abel. in 1st and 6th leaves. J. For. Res. 2018, 23, 309–317. [Google Scholar] [CrossRef]
- Bany Hani, M.; Othman, Y.; Al-Ajlouni, M.; A’saf, T. Deep planting improved stem root growth, flower yield and quality of Lilium cultivars. Hortic. Bras. 2022, 40, 143–150. [Google Scholar] [CrossRef]
- Ayad, J.; Othman, Y.; Al-Ajlouni, M.; Alsmirat, N. Photosynthesis, gas exchange and yield of two strawberry (Fragaria × ananassa Duch.) cultivars in response to gibberellic acid. Fresenius Environ. Bull. 2018, 27, 9127–9134. [Google Scholar]
- Chrysargyris, A.; Tzionis, A.; Xylia, P.; Tzortzakis, N. Effects of salinity on tagetes growth, physiology, and shelf life of edible flowers stored in passive modified atmosphere packaging or treated with ethanol. Front. Plant Sci. 2018, 9, 1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Akhter, M.; Noreen, S.; Mahmood, S.; Athar, H.; Ashraf, M.; Alsahli, A.; Ahmad, P. Influence of salinity stress on PSII in barley (Hordeum vulgare L.) genotypes, probed by chlorophyll-a fluorescence. J. King Saud Univ. Sci. 2021, 33, 101239. [Google Scholar] [CrossRef]
- Ding, Y.; Fan, Q.; He, J.; Wu, H.; Zou, Y.; Wu, Q.; Kuča, K. Effects of mycorrhizas on physiological performance and root TIPs expression in trifoliate orange under salt stress. Arch. Agron. Soil Sci. 2019, 66, 182–192. [Google Scholar] [CrossRef]
- Woodson, W. Gene Expression and Flower Senescence. In Genetics and Breeding of Ornamentals Species; Harding, J., Singh, F., Mol, J., Eds.; Kluwer Academic Pubishers: Amsterdam, Switzerland, 1991; pp. 317–381. [Google Scholar]
- Oki, L.; Mattson, N.; Lieth, J. Predicting stem length of cut flower roses at harvest using stem elongation rates in relationship to developmental events. Acta Hortic. 2006, 718, 113–120. [Google Scholar] [CrossRef]
- Akat, O.; Tüzel, I.; Özzambak, M. The effects of different salinity levels and leaching fractions on yield and water consumption of gerbera plants. Acta Hortic. 2009, 807, 233–238. [Google Scholar] [CrossRef]
- Singh, A.; Thakur, A.; Sharma, S.; Gill, P.; Kalia, A. Bio-inoculants enhance growth, nutrient uptake, and buddability of citrus plants under protected nursery conditions. Commun. Soil Sci. Plant Anal. 2018, 49, 2571–2586. [Google Scholar] [CrossRef]
- Navarro, J.M.; Pérez-Tornero, O.; Morte, A. Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J. Plant Physiol. 2014, 171, 76–85. [Google Scholar] [CrossRef]
- Perik, R.; Razé, D.; Harkema, H.; Zhong, Y.; van Doorn, W. Bending in cut Gerbera jamesonii flowers relates to adverse water relations and lack of stem sclerenchyma development, not to expansion of the stem central cavity or stem elongation. Postharvest Biol. Technol. 2012, 74, 11–18. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Treatment | SPAD | Pn (µmol m−2 s−1) | gs (mol m−2 s−1) | E (mmol m−2 s−1) | Leaf Temperature (°C) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | |
| Salinity | ||||||||||
| 0.0 mM | 44.7 a | 52.3 a | 12.4 a | 12.0 a | 0.14 a | 0.16 a | 4.74 | 4.94 a | 31.9 | 31.9 |
| 20 mM | 43.5 ab | 42.8 b | 12.0 a | 12.1 a | 0.10 b | 0.13 a | 4.29 | 4.71 a | 32.4 | 32.0 |
| 40 mM | 38.3 b | 36.8 c | 10.4 b | 9.80 b | 0.07 b | 0.08 b | 3.71 | 2.86 b | 32.9 | 32.3 |
| Mycorrhiza (AMF) | ||||||||||
| With AMF | 42.8 | 45.3 | 12.2 | 11.4 | 0.12 | 0.12 | 4.28 | 4.31 | 32.3 | 32.1 |
| No AMF | 41.1 | 42.7 | 11.3 | 11.3 | 0.10 | 0.13 | 4.19 | 3.97 | 32.5 | 31.9 |
| Significance | ||||||||||
| Salinity | * | *** | ** | ** | ** | * | NS | ** | NS | NS |
| AMF | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Salinity × AMF | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Treatment | SPAD | Pn (µmol m−2 s−1) | gs (mol m−2 s−1) | E (mmol m−2 s−1) | Leaf Temperature (°C) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | |
| Salinity | ||||||||||
| 0.0 mM | 48.4 a | 49.8 a | 9.17 a | 9.30 a | 0.08 | 0.22 a | 1.46 | 2.16 a | 18.3 | 19.1 |
| 20 mM | 43.2 a | 38.5 b | 6.68 b | 6.52 b | 0.07 | 0.11 b | 1.30 | 1.45 b | 19.0 | 18.5 |
| 40 mM | 33.6 b | 32.7 c | 5.32 b | 3.36 c | 0.07 | 0.06 c | 0.97 | 1.04 c | 19.4 | 17.8 |
| Mycorrhiza (AMF) | ||||||||||
| With AMF | 43.3 a | 42.3 a | 7.43 | 7.07 | 0.09 | 0.15 | 1.27 | 1.76 a | 19.0 | 18.4 |
| No AMF | 39.1 b | 38.3 b | 7.38 | 6.42 | 0.07 | 0.13 | 1.25 | 1.47 b | 18.8 | 18.6 |
| Significance | ||||||||||
| Salinity | *** | *** | *** | *** | NS | *** | NS | *** | NS | NS |
| AMF | * | * | NS | NS | NS | NS | NS | * | NS | NS |
| Salinity × AMF | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Treatment | Flower Yield (No Plant−1) | Flower Diameter (cm) | Pedicel Diameter (cm) | Pedicel Length (cm) | Vase Life (day) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | |
| Salinity | ||||||||||
| 0.0 mM | 8.42 a | 9.92 a | 9.55 | 8.87 a | 5.93 a | 5.84 a | 42.4 | 41.7 | 13.3 a | 11.8 a |
| 20 mM | 6.50 b | 6.42 b | 9.60 | 8.75 ab | 5.56 b | 5.60 a | 42.9 | 44.1 | 12.4 a | 10.6 b |
| 40 mM | 4.75 c | 3.67 c | 9.53 | 8.38 b | 5.13 c | 5.28 b | 42.4 | 40.4 | 11.2 b | 9.36 c |
| Mycorrhiza (AMF) | ||||||||||
| With AMF | 7.50 a | 7.83 a | 9.67 | 8.84 a | 5.86 a | 6.02 a | 42.3 | 42.8 | 13.3 a | 11.6 a |
| No AMF | 5.61 b | 5.50 b | 9.45 | 8.50 b | 5.06 b | 5.13 b | 42.8 | 41.3 | 11.2 b | 9.56 b |
| Significance | ||||||||||
| Salinity | *** | *** | NS | * | *** | *** | NS | NS | *** | *** |
| AMF | *** | *** | NS | * | *** | *** | NS | NS | *** | *** |
| Salinity × AMF | NS | NS | NS | NS | * | NS | NS | NS | NS | NS |
| Treatment | Flower Yield (No Plant−1) | Flower Diameter (cm) | Pedicel Diameter (cm) | Pedicel Length (cm) | Vase Life (day) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | Beaudine | Palm Beach | |
| Salinity | ||||||||||
| 0.0 mM | 7.67 a | 7.92 a | 9.45 | 8.55 a | 5.83 a | 5.93 a | 36.2 | 38.8 | 12.8 a | 11.5 a |
| 20 mM | 5.42 b | 5.25 b | 9.63 | 8.58 a | 5.57 b | 5.51 b | 36.0 | 41.0 | 12.4 a | 10.3 b |
| 40 mM | 3.08 c | 2.50 c | 9.21 | 8.18 b | 5.08 c | 5.05 c | 38.7 | 39.0 | 11.3 b | 8.73 c |
| Mycorrhiza (AMF) | ||||||||||
| With AMF | 6.28 a | 6.17 a | 9.63 a | 8.62 a | 5.84 a | 5.89 a | 37.5 | 40.1 | 13.7 a | 11.2 a |
| No AMF | 4.50 b | 4.28 b | 9.23 b | 8.25 b | 5.12 b | 5.00 b | 36.0 | 39.1 | 10.7 b | 9.18 b |
| Significance | ||||||||||
| Salinity | *** | *** | NS | * | *** | *** | NS | NS | ** | *** |
| AMF | *** | *** | * | * | *** | *** | NS | NS | *** | *** |
| Salinity × AMF | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Othman, Y.A.; Tahat, M.; Alananbeh, K.M.; Al-Ajlouni, M. Arbuscular Mycorrhizal Fungi Inoculation Improves Flower Yield and Postharvest Quality Component of Gerbera Grown under Different Salinity Levels. Agriculture 2022, 12, 978. https://doi.org/10.3390/agriculture12070978
Othman YA, Tahat M, Alananbeh KM, Al-Ajlouni M. Arbuscular Mycorrhizal Fungi Inoculation Improves Flower Yield and Postharvest Quality Component of Gerbera Grown under Different Salinity Levels. Agriculture. 2022; 12(7):978. https://doi.org/10.3390/agriculture12070978
Chicago/Turabian StyleOthman, Yahia A., Monther Tahat, Kholoud M. Alananbeh, and Malik Al-Ajlouni. 2022. "Arbuscular Mycorrhizal Fungi Inoculation Improves Flower Yield and Postharvest Quality Component of Gerbera Grown under Different Salinity Levels" Agriculture 12, no. 7: 978. https://doi.org/10.3390/agriculture12070978
APA StyleOthman, Y. A., Tahat, M., Alananbeh, K. M., & Al-Ajlouni, M. (2022). Arbuscular Mycorrhizal Fungi Inoculation Improves Flower Yield and Postharvest Quality Component of Gerbera Grown under Different Salinity Levels. Agriculture, 12(7), 978. https://doi.org/10.3390/agriculture12070978