Abstract
The rapidly emerging fertilizer rapeseed used as green manure has wide applications for use. However, there have been few studies on its decomposition and effects on soil nutrients and microorganisms after its decay. In this study, 12 rapeseed lines to be screened were decomposed through a randomized block field design with two green-manure-specific varieties as the controls. The contents of nitrogen, phosphorus, and potassium from the plants, soil nutrients, and microbial changes after degradation were measured. There were substantial differences in the rates of decomposition and cumulative release of nutrients among the different lines after 30 days of rolling. The contents of phosphorus and potassium in the soil were 1.23–2.03 and 3.93–6.32 times those before decomposition, respectively. In addition, there was a significant difference in the relative content of soil microorganisms at the phylum level after the decomposition of different species of rapeseeds. Most of the top 20 bacterial groups significantly correlated with the characteristics of plant decomposition and soil nutrient content, including Proteobacteria, Actinomycetes, Armatimonadetes, Rokubacteria, and Planctomycetes. A principal component analysis showed that the soil microorganisms and nutrients are the leading factors that enable the evaluation of the decomposing characteristics of green manure rapeseed. Numbers 5 (purple leaf mustard) and 8 (Xiafang self-seeding) were more effective than two controls, which can be used as excellent types of germplasm to promote the breeding of green manure rapeseed.
1. Introduction
In parallel with the rapid development of modern industry, the unreasonable application of chemical fertilizer has caused a series of environmental and safety problems [1,2,3], which deters the realization of the target for sustainable development [3]. Planting green fertilizer can reduce the use of chemical fertilizers and pesticides [4], and the use of green manure can increase the contents of soil nutrients and improve the soil fertility and structure of soil microbial community among others [5,6]. It is of substantial significance to cultivate high-quality green manure varieties and develop the green manure industry for the ecological environment and agricultural green sustainable development.
Green manure rapeseed, an organic fertilizer that has rapidly emerged in recent years, can be planted over a wide range of areas and is highly adaptable [7]. In addition to improving the contents of soil nutrients and increasing the number of beneficial bacterial communities in the soil [8,9], it results in the accumulation of large amounts of dry matter, which is twice or more than that of milk vetch (Astragalus sinicus L.) [10]. Moreover, previous studies have shown that the C/N ratio of rapeseed is relatively moderate, which renders it more easily decomposed by soil microorganisms than milk vetch, and the nutrients are easily absorbed by the subsequent crops [11]. Glucosinolate hydrolysate in rapeseed can inhibit weeds, soil pathogens, and soil nematodes [12,13,14], which is one of the important characteristics that renders it different from other green fertilizer crops. In addition, green manure rapeseed can be combined with various farming systems, such as the “rice–rice–rapeseed” [15] and “cotton–rapeseed” crop rotations in the Yangtze River Basin [16], with particular application value and broad development prospects.
The efficiency of decomposed green manure to serve as fertilizer is one of the main goals of the evaluation of green manure germplasm resources. The accumulation of soil organic matter is affected by the type of green manure, time and quantity in which it is returned, and environmental factors, among others [17,18,19]. Studies have shown that the decomposition rate can reach more than 50% within one month by returning rapeseed at the blooming stage [20]. However, the rates of decomposition and patterns of nutrient release of various rapeseed varieties differ [21]. In addition, there are few studies on the effects of rapeseed decomposition on soil microorganisms, and there is a lack of excellent green manure-specific varieties. Based on this, the decomposition regularities of 12 types of rapeseed and their impacts on soil nutrients and microorganisms were studied to screen high-quality green manure germplasm resources and promote the breeding of green manure rapeseed.
2. Materials and Methods
2.1. Overview of the Experimental Field
The experimental site was located in the Yunyuan Base of Hunan Agricultural University, Changsha, Hunan, China (113°07′ E, 28°18′ N) (Figure S1). It is located in a humid subtropical monsoon climate. During the decomposition period (20 March 2019 to 19 April 2019), the cumulative rainfall was 157.8 mm, and the average temperature was 16.5 °C. The experimental field implemented a “rice–oilseed” rotation. The tested soil was yellow-brown loam. Before the treatment of returning green manure to the soil, the soil pH was 5.70, and the content of organic matter, total nitrogen, available phosphorus, and available potassium were 21.60 g·kg−1, 1.29 g·Kg−1, 10.23 mg·kg−1, and 137.76 mg·kg−1, respectively.
2.2. Experimental Design
Field simulation experiments were utilized with three replicates in this study, and the abridged general view is shown in Figure S1. The plants were collected at the blooming stage. The samples were cut into 2–3 cm segments and weighed until 300 g of each had been collected. They were then mixed evenly and packed in nylon mesh bags that were 25 × 35 cm with an aperture of 74 μm. The bags were buried 20 cm below the soil surface with the samples 30 cm apart from each other. The plant residues and soil samples were taken after 30 days. The plant residues were dried to determine their nutrient contents and rate of decomposition of each variety. One part of the soil samples was dried to determine the nutrient content, while the other part was quickly placed in liquid nitrogen and stored at −80 °C to determine their microbial diversity. The basic traits of rapeseed before decomposition are shown in Table 1.

Table 1.
Basic characters of rapeseed before decomposition.
2.3. Test Method
The nutrients of plant samples were determined as follows: The total nitrogen was determined using a SKALAR interval flow analyzer. The total phosphorus was determined by the Mo-Sb-Vc colorimetric method [22], and the total potassium was determined by atomic absorption spectrophotometry [23].
The soil samples were analyzed to determine their contents of nutrients: The total nitrogen was determined using the semi micro Kjeldahl method [24]. The available phosphorus was determined using the Olsen sodium bicarbonate method [25], and the available potassium was determined by NH4OAc (1.0 mol·L−1) extraction atomic absorption spectrophotometry [26].
The experiment was conducted as described by Wang Yingying [23]. Novogene Biotech Co., Ltd. (Beijing, China) was utilized to perform 16S (V3 + V4) regional amplicon sequencing by Single-End based on an IonS5TMXL sequencing platform.
2.4. Data Analysis
Decomposition rate was calculated as follows:
Decomposition rate (%) = (M0 − M30)/M0 × 100
Plant nutrient release rate was calculated as follows:
where M0 is the initial dry matter weight of the test material; M30 is the dry matter weight (residue) of green fertilizer for 30 days; C0 is the initial nutrient concentration of the test material, and C30 is the nutrient concentration of the test material for 30 days.
Plant nutrient release rate (%) = (C0 × M0 − C30 × M30)/C0 × M0 × 100
SPSS 24.0 (IBM, Inc., Armonk, NY, USA) was used for correlation and principal component analyses, and Origin 2019 (OriginLab, Northampton, MA, USA) was used for mapping. The LSD method was used to determine the difference significance test. The difference was considered to be significant at p < 0.05. The bioinformatics analysis of high-throughput data was conducted using Novo Magic (https://magic.novogene.com, accessed on 23 August 2021).
3. Results and Analysis
3.1. Decomposition of Rapeseed Plants and the Influence of Soil Nutrients
The analysis of the plants in Table 2 shows that the rate of decomposition and the cumulative release rate of nitrogen (N) and phosphorus (P) vary substantially among the different materials. The 30-day decomposition rate of each material was between 37.12% and 76.13%. Number 8 decomposed at the highest rate, whereas number 10 decomposed the most slowly. The cumulative release rate of nitrogen (N) was between 17.88% and 54.59%. Numbers 5, 1, and 4 were higher. The cumulative release rate of phosphorus (P) was between 15.28% and 39.37%. Numbers 3 and 1 were higher than those of the two controls. The cumulative release rate of potassium (K) was relatively high, ranging from 88.28% to 95.67%, but there was little difference among the rapeseeds.
Table 2.
Plant decomposition and soil nutrient content of rapeseed.
The analysis of soil nutrients of rapeseeds before and after decomposition is shown in Table 2. The contents of soil nitrogen (N) increased after the decomposition of most rapeseed lines, whereas the difference was not significant. The contents of soil phosphorus (P) increased significantly after the decomposition of rapeseed numbers 12, 2, and 3, which was higher than that of both controls. The content of soil potassium (K) increased significantly and was 3.93- to 6.32-fold higher than that before decomposition. With the exception of 6, 4, 1, and 2, the other strains were higher than both controls.
3.2. The Effects of Rapeseed on Soil Microbial Diversity and Community Structure after Decomposition
3.2.1. The Effect of Rapeseed on Soil Microbial Community Structure after Decomposition
On the decomposed and pre-decomposed soil samples of 14 types of rapeseeds, 16S sequencing was performed. A total of 3,588,932 raw reads were obtained, and each sample obtained an average of 79,754 ± 9571 sequences (Table S1). The sample-based species accumulation box plot in Figure 1a shows that the number of orthologous taxonomic units (OTUs) increased with the number of samples. As the number of OTUs became saturated, the curve was asymptotically stable, and the number of new OTUs added to each sample decreased, indicating that the sequencing depth was accurate enough to represent the composition of soil bacteria. The dilution curve (Figure 1b) shows that with the increase in number of sequences, the bacterial species reached the saturation stage, indicating that the microorganisms in the population captured most of the bacterial members of each strain. After the quality control of the sequences, OTU cluster analysis was conducted using 97% similarity (Figure 1c). The results showed that the common number of OTUs in each sample was 1166, comprising 39.27% of the total OTUs. This indicated that the populations of various varieties (lines) differed somewhat, with the number of specific OTUs of the CK3 totaling 418, comprising 15.31% of the total OTUs. Alternatively, there were differences in the soil microorganisms before and after decomposition. Based on the annotation results of the SILVA database (Figure 1d), the top 10 species of soil bacteria at the phylum level were Proteobacteria, Acidobacteria, Bacteroidetes, Actinobacteria, Chloroflexi, Gemmatimonadetes, Thaumarchaeota, Firmicutes, Verrucomicrobia, and unknown microorganisms. Among them, CK3 and the other strains clustered into two branches. The abundances of Acidobacteria and Bacteroides in the decomposed soil increased, while the abundances of Actinomyces and Chloroflexi decreased. In general, there was no significant difference in the composition of soil microorganisms at the phylum level after the decomposition of different varieties (lines), but the relative content differed somewhat.
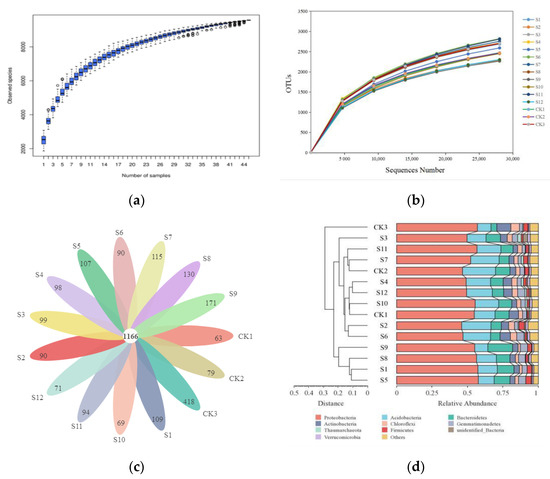
Figure 1.
Differences in soil microorganisms among different strains after decomposition. (a): the species accumulation box plot; (b): the dilution curve; (c): the petal diagram; (d): the clustering stacked histogram.
3.2.2. The Effects of Rapeseed on the Diversity and Function of Soil Microbial Communities after Decomposition
The α diversity of soil bacteria in Table 3 shows that the sequencing depth of samples was >96.50% (coverage index), and there was no significant difference between the original soil (CK3, Table 2) and decomposed soil of the rapeseed plants in the Chao1 value and OTU number, with the exception of number 12. The Shannon (except for number 9) and Simpson indices of the original soil were lower than those of the soil that contained the decomposed tissue. The former did not differ significantly among the decomposed soil of rapeseeds. Overall, the dominance of soil microbial community did not change significantly after the decomposition of rapeseeds. The soil samples of numbers 11 and 6 had a higher abundance and diversity of colonies.
Table 3.
α-Diversity of the bacterial community in different types of rapeseed cultivars.
The functional analysis of each sample community obtained by the FAPROTAX software (Figure 2) showed that the first 15 types of functions were primarily types of nitrogen metabolism. Compared with the original soil, the abundance of genera related to nitrogen metabolism in the decomposed soil was higher, while the abundance of animal parasitic or symbiotic bacteria and animal human intestinal microflora was lower. This differed from the abundance of microbial functional categories for the soil in which the rapeseeds had decomposed. The original soil (CK3, Table 2) and the decomposed soil clustered into two branches, indicating that the main functional groups of soil microorganisms had clearly changed.
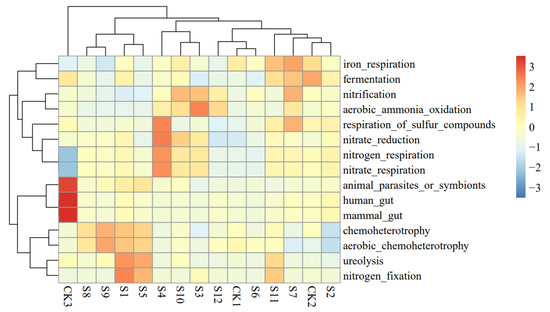
Figure 2.
Cluster diagram of soil microbial community function in different varieties.
3.3. Correlation Analysis and Principal Component Analysis of Environmental Factors, Plant Nutrients, Soil Nutrients, and Microorganisms
3.3.1. Correlation Analysis of Environmental Factors, Plant Nutrients, Soil Nutrients, and Microorganisms
A Spearman analysis was used to analyze the correlation between plant properties before and after decomposition and soil nutrients and microbial properties after decomposition (Figure 3). There were significant positive correlations between the plant cumulative decomposition rates of N and P, whereas there was a significant positive correlation between the dry matter weight and the soil N and P. Most of the top 20 bacterial groups significantly correlated with the decomposition characters of the plants, and the soil nutrient content, Proteobacteria, Actinobacteria, Bacteroidetes, Rokubacteria, and Planctomycetes, extremely significantly correlated with the traits of plants before and after decomposition and soil nutrients in particular.
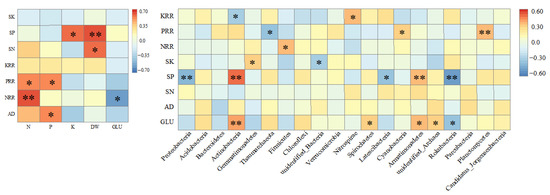
Figure 3.
Correlation analysis between environmental factors and soil nutrients and microorganisms. DW: dry weight; GLU: glucosinolate content; AD: accumulative decomposition rate; NRR, PRR, and KRR: the cumulative release rate of nitrogen, phosphorus, and potassium, respectively; TN, TP, and TK: the contents of soil nitrogen, phosphorus, and potassium, respectively. Asterisks denote statistical significance at the p < 0.05 (*) and p < 0.01 (**) level.
3.3.2. Environmental Factors and Principal Component Analysis of Soil Microbial Community
Based on the analysis of Section 3.3.1, there were positive or negative correlations among the plant nutrient content, soil nutrients, and microorganisms. The nutrient content of plant samples before and after decomposition, the corresponding soil nutrient content after decomposition, and the five types of bacteria with significant differences described above were standardized for a principal component analysis. Table 4 lists the first six principal components, which can represent 87.08% of the nutrient information of 14 types of rapeseed plants. The first principal component contributed the most, comprising 29.86%, and the soil microorganisms and plant nutrients comprised the larger positive load. The second principal component comprised 16.71%, and the contents of soil nutrients comprised a large positive load. In the third principal component, the rate of plant decomposition and nutrient release comprised a large positive load. In the fourth principal component, the content of soil nitrogen and rate of plant decomposition comprised a large positive load. The fifth and sixth principal components that comprised the larger positive load were the plant trait factors before decomposition. The normalized values of plant and soil nutrients and microbial characteristics were multiplied by the eigenvectors of the first six principal components and weighted to obtain the scores and ranking of each principal component (Table 5). Compared with the two controls, numbers 5, 9, 8, and 10 had higher characteristics of soil microbes and nutrients, and numbers 8, 6, 2, and 5 had higher characteristics of plant nutrients and decomposition. The overall analysis indicated that numbers 5 and 8 were more effective than those of CK1 and CK2.
Table 4.
Principal component analysis of plant and soil nutrient characteristics.
Table 5.
Comprehensive comparison of nutrient characteristics via a principal component analysis.
4. Discussion
4.1. Decomposition of Rapeseed
The most effective fertilization can be achieved by returning rapeseed at the blooming stage [27]. Therefore, the materials at this stage were selected for decomposition in this experiment, and the decomposition rate after 30 days was between 37.12% and 76.13%. Except for numbers 2, 9, 10, and 11, the rate of decomposition of other strains exceeded 50%, which was consistent with the findings of previous studies [20]. The decomposition and cumulative rates of release of N and P among the different rapeseeds are highly variable, which may be related to their own characteristics. The high rate of decomposition of number 8 may be owing to its low contents of lignin and cellulose (Table S2).
Alternatively, returning rapeseed to the field plays an important role in improving the soil and farmland systems [7,8,9,10]. In this experiment, the contents of soil P and K increased significantly, and the content of K, in particular, increased by 4–6-fold after the return of rapeseeds, which showed that one month’s decomposition time can effectively meet the growth of the next crops. In addition, although the cumulative rate of release of N was between 17.88% and 54.59%, there was no apparent increase in the soil N. This could be caused by the action of soil denitrifying bacteria, the alkaline soil pH, high content of soil CaCO3, or high temperature [17], which could make a profound study. Although the increase in soil nitrogen is not significant, it can also reduce the use of nitrogen fertilizer. Xiao Xiaojun et al. [28] showed that under the condition of returning rapeseed, only a small amount of nitrogen fertilizer can meet the growth of the subsequent rice crop. In general, utilizing soil nutrients as screening conditions, except for CK1 and CK2, numbers 2 and 3 are suitable for use in soil that is slightly deficient in nitrogen. Numbers 9, 5, 7, and 8 are suitable for phosphorus deficient soil, while all of the strains are suitable for potassium deficient soil.
4.2. Effects of Decomposition on Soil Bacterial Diversity and Community Structure
In addition to increasing the content of soil nutrients and improving the soil physical and chemical properties, the impact on soil microorganisms is a critical question. Based on the annotation results of the SILVA database, there was no significant difference in the composition of soil microorganisms at the phylum level after the decomposition of different rapeseeds, and the Chao index and OTUs of the original soil (CK3) were relatively high, indicating that the abundance and composition of the soil bacterial community in natural soil field were relatively stable in a certain environment, which suggests that the decomposition of rapeseed does not destroy the original microbial structure. After decomposition, the Simpson and Shannon indices increased (with the exception of number 9), indicating that sugars, proteins, amino acids, and other substances may provide nutrition for microorganisms or form part of the soil humus during the decomposition process of rapeseed, which increases the diversity of some soil bacteria [29,30].
Furthermore, a cluster analysis showed that the microbial groups of CK3 and the soil after rapeseed decomposition are divided into two branches, and the abundances of Acidobacteria and Bacteroidetes increased significantly after decomposition. Acidobacteria, known as “ecosystem engineer”, participate in the metabolism and transportation of carbon, nitrogen, and sulfur and are the core community of nitrogen cycle [31]. Simultaneously, it also regulates the biosynthesis of secondary metabolites and the stress and hunger response. In addition, overturning and covering the green manure or adding organic fertilizer are conducive to the growth of Bacteroidetes [32,33]. It is hypothesized that Acidobacteria and Bacteroidetes play important roles in the decomposition of rapeseed. Alternatively, the decrease in the abundance of Actinobacteria and Chloroflexi in the soil after rapeseed decomposition is completely consistent with the research of Li Lina et al. [34]. Although Chloroflexi is the dominant population in soil, there is little understanding of the function of such microorganisms because most of them are not culturable [35]. Some previous studies have shown that some of the members of Chloroflexi, such as GP6, GP16, and GP4, significantly negatively correlate with the content of organic carbon in soil [36], which may be the reason for the relatively high content of Chloroflexi without returning rapeseed in this experiment. Similarly, Actinobacteria are less competitive than other microbial groups in soils that contain a higher level of nutrients [32]. However, although the increase in Acidobacteria and Bacteroidetes and the decrease in Actinobacteria and Chloroflexi can indicate that the populations of soil microorganisms have changed after decomposition, it is not an effective method to evaluate the difference in green manure rapeseed because its changes are similar in all strains.
4.3. Correlation between Soil Nutrients, Microbial Composition, and the Plant Decomposition Index
The decomposition of green manure will affect the soil nutrients, and the latter will affect the structural diversity and abundance of the soil microbial community [5,6,18]. Microorganisms are the main driving force of the soil nutrient material cycle, and the microbial communities in the soil are vulnerable to the influence of the external environment; thus, they react to the soil environment [37,38]. In this study, most of the top 20 bacterial groups significantly correlated with the characteristics of plant decomposition and soil nutrient content, including Proteobacteria, Actinobacteria, Armatimonadetes, Rokubacteria, and Planctomycetes that extremely significantly correlated with soil nutrients and plant properties before and after decomposition. Proteobacteria play an important role in nitrogen fixation and degradation [39,40]. Similar to the Actinobacteria, their abundance reflects their ability to grow in different environments, which can regulate the amount of soil fertilization [41]. Armatimonadetes constitutes a moderately abundant and phylogenetically diverse bacterial phylum [42], which is known to be capable of the metabolism of ammonium [43]. Interestingly, similar to Actinobacteria, it has a significant positive correlation with the contents of plant glucosinolates and soil phosphorus. The establishment of a model to predict their content would help to elucidate their functions. Rokubacteria are known as the “genome giant in uncultured bacteria”, because they have the potential of a large genome (6–8 Mbps), a high GC content (66–71%) [44], and multifunctional and mixed-nutrient metabolism [45,46]. Planctomycetes are widely distributed and abundant components of the soil microbial community. They participate in sugar degradation and are highly significant to the global nitrogen cycle. The changes in soil organic matter, Ca2+ content, and pH will affect their community composition [47,48]. Buckley et al. [47] showed that the abundance of Planctomycetes was related to the spatial heterogeneity of nitrate, which can be used to judge the historical conditions of soil. Therefore, it seems likely that the suitable utilization of these bacteria can promote the fertilization of crops and healthy operation of the agroecosystem. More importantly, taking the relationship between these five bacteria and soil nutrients and the plant decomposition index as the evaluation factor for screening green manure rapeseed is of substantial significance for the development of green manure rapeseed breeding, which merits further study.
5. Conclusions
In summary, compared with the original soil, the decomposition of rapeseed significantly increased the contents of soil nutrients and bacterial community structure and diversity. Proteobacteria, Actinobacteria, Armatimonadetes, Rokubacteria, and Planctomycetes in the soil significantly correlated with soil nutrients and plant properties before and after decomposition. This indicates that they may play an important role in the decomposition of rapeseed. A principal component analysis showed that soil microorganisms and nutrients were the leading factors in the evaluation of the characteristics of decomposition of green manure rapeseed. Compared with the CK1 and CK2, numbers 5 (purple leaf mustard) and 8 (Xiafang self-seeding) performed outstandingly, which renders them valuable for use as high-quality green manure rapeseed materials for further research.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agriculture11121219/s1, Figure S1: Schematic diagram of experimental site, Table S1: Data preprocessing statistics and quality control, Table S2: Basic characters of oilseed rape before decomposition.
Author Contributions
C.G. and M.G. designed the experiment. H.M. conducted the experiment, and X.W. processed and analyzed data and wrote the first draft. C.G. and M.G. revised and edited. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Rapeseed Industrial Technology System (CARS-13) and the Natural Science Foundation of China (grant no. 31130040).
Data Availability Statement
All the data included in this study are available upon request by contact with the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, Q.; Awasthi, M.K.; Zhang, Z.; Wong, J.W.C. Sustainable composting and its environmental implications. In Sustainable Resource Recovery and Zero Waste Approaches, 1st ed.; Taherzadeh, M.J., Bolton, K., Wong, J., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 7, pp. 115–132. [Google Scholar] [CrossRef]
- Ren, F.L.; Zhang, X.B.; Liu, J.; Sun, N.; Wu, L.H.; Xu, M.G. A synthetic analysis of greenhouse gas emissions from manure amended agricultural soils in China. Sci. Rep. 2017, 7, 8123. [Google Scholar] [CrossRef]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of Pesticides Use in Agriculture: Their Benefits and Hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Lin, W.; Lin, M.; Zhou, H.; Wu, H.; Li, Z.; Lin, W. The effects of chemical and organic fertilizer usage on rhizosphere soil in tea orchards. PLoS ONE 2019, 14, e0217018. [Google Scholar] [CrossRef]
- Dong, N.G.; Hu, G.L.; Zhang, Y.Q.; Qi, J.X.; Chen, Y.H.; Hao, Y.B. Effects of green-manure and tillage management on soil microbial community composition, nutrients and tree growth in a walnut orchard. Sci. Rep. 2021, 11, 16882. [Google Scholar] [CrossRef]
- Costerousse, B.; Quattrini, J.; Grüter, R.; Frossard, E.; Thonar, C. Green manure effect on the ability of native and inoculated soil bacteria to mobilize zinc for wheat uptake (Triticum aestivum L.). Plant Soil 2021, 467, 287–309. [Google Scholar] [CrossRef]
- Dong-Hui, F.U.; Jiang, L.Y.; Mason, A.S.; Xiao, M.L.; Zhu, L.R.; Li-Zhi, L.I.; Zhou, Q.H.; Shen, C.J.; Huang, C.H. Research progress and strategies for multifunctional rapeseed: A case study of china. J. Integr. Agric. 2016, 15, 1673–1684. [Google Scholar] [CrossRef]
- Huang, M.; Zhou, X.F.; Xie, X.B.; Chen, J.N.; Zou, Y.B. Increased soil fertility in a long-term rice-oilseed rape cropping system and its potential roles in reducing nitrogen inputs and environmental impacts. In Cropping Systems: Applications, Management and Impact; Hodges, J.G., Ed.; Nova Science Publishers: New York, NY, USA, 2017; pp. 103–113. ISBN 978-63485-888-5. [Google Scholar]
- Li, W.G.; Yang, X.X.; Huang, C.G.; Xue, N.W.; Xia, Q.; Liu, X.L.; Zhang, X.Q.; Yang, S.; Yang, Z.P.; Gao, Z.Q. Effects of rapeseed green manure on soil fertility and bacterial community in dryland wheat field. Agric. Sci. China 2019, 52, 2664–2677. [Google Scholar] [CrossRef]
- Gu, C.M.; Li, Y.S.; Xie, L.H.; Hu, X.J.; Liao, X.; Qin, L. Analysis on application advantages of rapeseed as green manure. Soil Fertil. Sci. China 2019, 01, 180–183. [Google Scholar] [CrossRef]
- Wang, D.Y.; Peng, J.; Xu, C.M.; Zhao, F.; Zhang, X.F. Effects of rape straw manuring on soil fertility and rice growth. Chin. J. Rice Sci. 2011, 26, 85–91. [Google Scholar] [CrossRef]
- Haramoto, E.; Gallandt, E. Brassica cover cropping for weed management: A review. Renew. Agric. Food. Syst. 2004, 19, 187–198. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D.; Velasco, P. Natural control of plant pathogens through glucosinolates: An effective strategy against fungi and oomycetes. Phytochem. Rev. 2020, 19, 1045–1059. [Google Scholar] [CrossRef]
- Johnson, A.W.; Golden, A.M.; Auld, D.L.; Sumner, D.R. Effects of rapeseed and vetch as green manure crops and fallow on nematodes and soil-borne pathogens. J. Nematol. 1992, 24, 117–126. [Google Scholar] [CrossRef]
- Huang, M.; Tian, A.L.; Zhou, X.; Gao, W.; Li, Z.B.; Chen, G.; Li, A.H.; Chen, Y.M.; Liu, L.S.; Yin, X.H.; et al. Yield performance of machine-transplanted double-season rice grown following oilseed rape. Sci. Rep. 2019, 9, 6818. [Google Scholar] [CrossRef]
- Ren, T.; Li, H.; Lu, J.W.; Bu, R.Y.; Li, X.K.; Cong, R.H.; Lu, M.X. Crop rotation-dependent yield responses to fertilization in winter oilseed rape (Brassica napus L.). Crop J. 2015, 3, 396–404. [Google Scholar] [CrossRef][Green Version]
- Edmeades, D.C. The long-term effects of manures and fertilisers on soil productivity and quality: A review. Nutr. Cycl. Agroecosyst. 2003, 66, 165–180. [Google Scholar] [CrossRef]
- Sharma, P.; Laor, Y.; Raviv, M.; Medina, S.; Saadi, I.; Krasnovsky, A.; Vager, M.; Levy, G.J.; Bar-Tal, A.; Borisover, M. Green manure as part of organic management cycle: Effects on changes in organic matter characteristics across the soil profile. Geoderma 2017, 305, 197–207. [Google Scholar] [CrossRef]
- Adekiya, A.O.; Agbede, T.M.; Aboyeji, C.M.; Dunsin, O.; Ugbe, J.O. Green manures and npk fertilizer effects on soil properties, growth, yield, mineral and vitamin c composition of okra (abelmoschus esculentus (L.) moench). J. Saudi Soc. Agric. Sci. 2019, 18, 218–223. [Google Scholar] [CrossRef]
- Liu, X.H.; Zhou, X.; Deng, L.C.; Fan, L.Y.; Qu, L.; Li, M. Decomposition characteristics of rapeseed green manure and effect of nutrient release on soil fertility. Hunan Agric. Sci. 2020, 416, 39–44. [Google Scholar] [CrossRef]
- Kriaučiūnienė, Z.; Čepulienė, R.; Velička, R.; Marcinkevičienė, A.; Lekavičienė, K.; Šarauskis, E. Oilseed Rape Crop Residues: Decomposition, Properties and Allelopathic Effects. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2018; Volume 32. [Google Scholar] [CrossRef]
- Yuan, Y.Y.; Liu, Q.M.; Zhong, Y.S.; Liao, F.P.; Lin, J.R. Mechanism of CP7 antibacterial protein against Aeromonas hydrophila. Microbiol. China 2012, 39, 949–957. [Google Scholar]
- Wang, Y.Y. Evaluation of Different Varieties of Mung Bean Used as Green Manure. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, June 2011. [Google Scholar]
- Fawcett, J.K. The semi-micro Kjeldahl method for the determination of nitrogen. J. Med. Lab. Technol. 1954, 12, 1–22. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U.S. Department of Agriculture: Washington, DC, USA, 1954.
- Bao, S.D. Soil and Agrochemistry Analysis, 3rd ed.; Chinese Agriculture Press: Beijing, China, 2005; pp. 44–107. [Google Scholar]
- Han, P.P.; Hu, X.J.; Liao, X.S.; Xie, L.H.; Li, Y.S.; Gu, C.M.; Qin, L.; Liao, X. Effects of flowering rapeseed-manure returns on soil microorganism and growth of peanut. Chin. J. Oil. Crop. Sci. 2019, 41, 638–642. [Google Scholar] [CrossRef]
- Xiao, X.J.; Lü, W.S.; Yu, P.L.; Zheng, W.; Li, Y.Z.; Hu, L.; Xiao, F.L.; Zhang, S.W.; Huang, T.B.; Xiao, G.B. Effects of nitrogen application rate on yield formation and nitrogen use efficiency of early rice under rape straw returning in triple cropping. Crops 2019, 2, 103–109. [Google Scholar] [CrossRef]
- Corato, U.D. Agricultural waste recycling in horticultural intensive farming systems by on-farm composting and compost-based tea application improves soil quality and plant health: A review under the perspective of a circular economy. Sci. Total Environ. 2020, 738, 139840. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Celano, G.; Piccolo, A.; Villecco, D.; Spaccini, R.; Palese, M.A.; Zaccardelli, M. Effects of on-farm composted tomato residues on soil biological activity and yields in a tomato cropping system. Chem. Biol. Technol. Agric. 2015, 2, 4. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; Suriani, N.L. Recent understanding of soil acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, R.F.; Xue, C.; Xun, W.B.; Sun, L.; Xu, Y.C.; Shen, Q.R. Pyrosequencing reveals contrasting soil bacterial diversity and community structure of two main winter wheat cropping systems in China. Microb. Ecol. 2014, 67, 443–453. [Google Scholar] [CrossRef]
- Becraft, E.D.; Woyke, T.; Jarett, J.; Ivanova, N.; Godoy-Vitorino, F.; Poulton, N.; Brown, J.M.; Brown, J.; Lau, M.C.Y.; Onstott, T.; et al. Rokubacteria: Genomic Giants among the Uncultured Bacterial Phyla. Front. Microbiol. 2017, 8, 2264. [Google Scholar] [CrossRef]
- Li, L.N.; Xi, Y.G.; Chen, E.; He, L.P.; Wang, L.; Xiao, X.J.; Tian, W. Effects of tillage and green manure crop on composition and diversity of soil microbial community. J. Ecol. Rural Environ. 2018, 34, 342–348. [Google Scholar] [CrossRef]
- Yamada, T.; Sekiguchi, Y. Cultivation of uncultured chloroflexi subphyla: Significance and ecophysiology of formerly uncultured chloroflexi ‘Subphylum I’ with natural and biotechnological relevance. Microbes Environ. 2009, 24, 205–216. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition-current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liao, M.; Fang, Z.; Guo, J.; Xie, X.; Xu, C. How silicon fertilizer improves nitrogen and phosphorus nutrient availability in paddy soil? J. Zhejiang Univ. Sci. B 2021, 22, 521–532. [Google Scholar] [CrossRef]
- Chen, W.M.; Moulin, L.; Bontemps, C.; Vandamme, P.; Bena, G.; Boivin-Masson, C. Legume Symbiotic Nitrogen Fixation by Beta-Proteobacteria Is Widespread in Nature. J. Bacteriol. 2003, 185, 7266–7272. [Google Scholar] [CrossRef] [PubMed]
- Aislabie, J.; Deslippe, J.R.; Dymond, J. Soil Microbes and Their Contribution to Soil Services. In Ecosystems Services in New Zealand Conditions and Trends; Dymond, J.R., Ed.; Manaaki Press: Lincoln, New Zealand, 2013; Volume 1, pp. 143–161. ISSN 0478347367. [Google Scholar]
- Lee, K.C.Y.; Dunfield, P.F.; Stott, M.B. The Phylum Armatimonadetes. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Wang, D.P.; Li, T.; Huang, K.L.; He, X.W.; Zhang, X.X. Roles and correlations of functional bacteria and genes in the start-up of simultaneous anammox and denitrification system for enhanced nitrogen removal. Sci. Total Environ. 2019, 655, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Bukar, M.; Sodipo, O.; Dawkins, K.; Ramirez, R.; Esiobu, N. Microbiomes of top and sub-layers of semi-arid soils in north-eastern nigeria are rich in firmicutes and proteobacteria with surprisingly high diversity of rare species. Adv. Microbiol. 2019, 9, 102–118. [Google Scholar] [CrossRef]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczykc, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by culture-independent approach. Appl. Soil. Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Kroeger, M.E.; Delmont, T.O.; Eren, A.M.; Meyer, K.M.; Guo, J.; Kiran, K.; Rodrigues, J.L.M.; Bohannan, B.J.M.; Tringe, S.G.; Borges, C.D. New biological insights into how deforestation in amazonia affects soil microbial communities using metagenomics and metagenome-assembled genomes. Front. Microbiol. 2018, 9, 1635. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, C.N.; Li, Z.; Andeer, P.F.; Spaulding, S.; Banfield, J.F. Proteogenomic analyses indicate bacterial methylotrophy and archaeal heterotrophy are prevalent below the grass root zone. PeerJ 2016, 4, e2687. [Google Scholar] [CrossRef]
- Buckley, D.H.; Huangyutitham, V.; Nelson, T.A.; Rumberger, A.; Thies, J.E. Diversity of planctomycetes in soil in relation to soil history and environmental heterogeneity. Appl. Environ. Microbiol. 2006, 72, 4522–4531. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Ivanova, A.A. Planctomycetes in boreal and subarctic wetlands: Diversity patterns and potential ecological functions. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).