Floral Biology and Pollination Efficiency in Yam (Dioscorea spp.)

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Overview on Floral Biology and Pollination in Yam
3. Pollen Viability Assessment
3.1. In Vitro Germination
3.2. Staining Methods
3.3. In Vivo Germination
3.4. Impedance and Optical Flow Cytometry
3.5. Imagery in Pollen Quality Assessment
4. Pollen Storage and Longevity
4.1. Pollen Collection
4.2. Storage Methods
4.3. Factors Affecting Stored Pollen Longevity
4.4. Post-Storage Practices, Pollen Retrieval and Usage
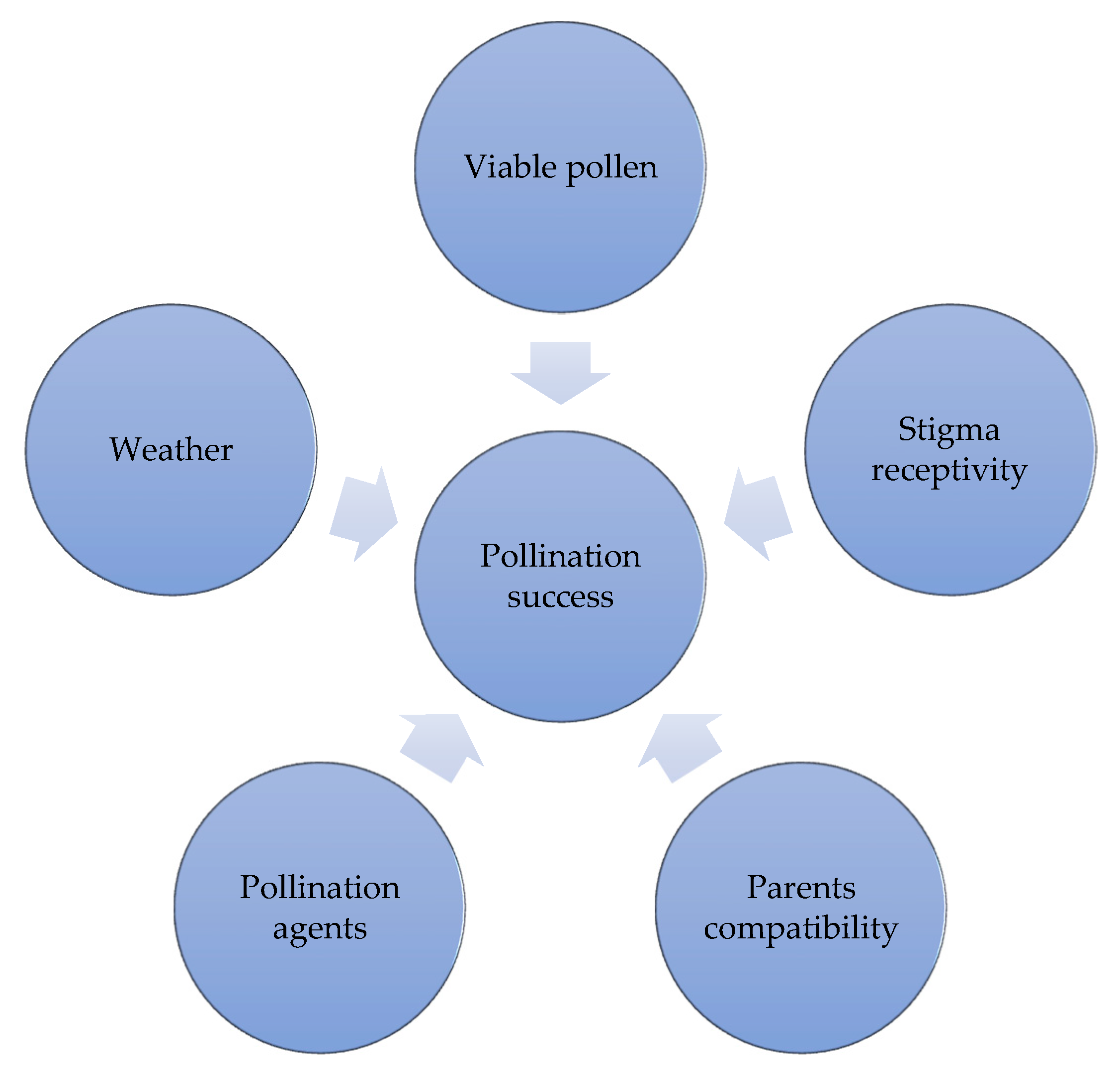
5. Pollination Practices, Constraints and Tips for Improving Efficiency
5.1. Yam Crossing
5.2. Time of Pollination
5.3. Pollination Tools
5.4. Mating Designs and Pollination in Yam Breeding
6. Manipulation of Flowering: Lessons from Other Root and Tuber Crops
7. Insect Pollinators’ Identification and Management
8. Conclusions
- (1)
- The establishment of highly reproducible and cost-effective protocols for quality pollen collection and storage, necessary for breaking the asynchrony and low flowering barriers. These storage efforts should be complemented by agronomic and/or hormonal manipulations to enhance the flowering intensity with long flowering window.
- (2)
- Sites selection promoting optimum flowering, fruit and seed set.
- (3)
- Markers prediction for flower sex’s phenotype and intensity at early growth stage of the plant.
- (4)
- Profiling of genotypes for ploidy status, cross-compatibility and flowering time to identify compatible groups.
- (5)
- Use of biotechnological approaches such as ploidy manipulation, gene editing, androgenesis or gynogenesis induction, embryo rescue, somatic and protoplast fusion, etc. to break intra and inter-specific cross incompatibility.
- (6)
- Proper choice of mating design, rearing insect pollinators, and a continuous training of technicians involved in pollination activities.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilkin, P.; Schols, P.; Chase, M.W.; Chayamarit, K.; Furness, C.A.; Huysmans, S.; Rakotonasolo, F.; Smets, E.; Thapyai, C. A Plastid Gene Phylogeny of the Yam Genus, Dioscorea: Roots, Fruits and Madagascar. Syst. Bot. 2005, 30, 736–749. [Google Scholar] [CrossRef]
- Govaerts, R.; Wilkin, P.; Saunders, R.M.K. World Checklist of Dioscoreales; Yams and Their Allies: The Board of Trustees of the Royal Botanic Gardens: Kew, UK; London, UK, 2007; pp. 1–65. [Google Scholar]
- Darkwa, K.; Olasanmi, B.; Asiedu, R.; Asfaw, A. Review of empirical and emerging breeding methods and tools for yam (Dioscorea spp.) improvement: Status and prospects. Plant Breed. 2019, 139, 474–497. [Google Scholar] [CrossRef]
- Gatarira, C.; Paterne, A.A.; Matsumoto, R.; Edemodu, A.; Adetimirin, V.; Bhattacharjee, R.; Asiedu, R.; Asfaw, A. Genome-Wide Association Analysis for Tuber Dry Matter and Oxidative Browning in Water Yam (Dioscorea alata L.). Plants 2020, 9, 969. [Google Scholar] [CrossRef] [PubMed]
- Coursey, D.G. Yams. An Account of the Nature, Origins, Cultivation and Utilisation of the Useful Members of the Dioscoreaceae; Longmans: London, UK, 1967; p. 230. [Google Scholar]
- Scarcelli, N.; Cubry, P.; Akakpo, R.; Thuillet, A.-C.; Obidiegwu, J.E.; Baco, M.N.; Otoo, E.; Sonké, B.; Dansi, A.; Djedatin, G.; et al. Yam genomics supports West Africa as a major cradle of crop domestication. Sci. Adv. 2019, 5, e1947. [Google Scholar] [CrossRef]
- FAO FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QL (accessed on 1 September 2020).
- Obidiegwu, J.E.; Akpabio, E.M. The geography of yam cultivation in southern Nigeria: Exploring its social meanings and cultural functions. J. Ethn. Foods 2017, 4, 28–35. [Google Scholar] [CrossRef]
- Omohimi, C.I.; Piccirillo, C.; Roriz, M.; Ferraro, V.; Vasconcelos, M.; Sanni, L.O.; Tomlins, K.; Pintado, M.M.; Abayomi, L.A. Study of the proximate and mineral composition of different Nigerian yam chips, flakes and flours. J. Food Sci. Technol. 2017, 55, 42–51. [Google Scholar] [CrossRef]
- McKoy, M.-L.; Thomas, P.-G.; Asemota, H.N.; O Omoruyi, F.; Simon, O. Effects of Jamaican Bitter Yam (Dioscorea polygonoides) and Diosgenin on Blood and Fecal Cholesterol in Rats. J. Med. Food 2014, 17, 1183–1188. [Google Scholar] [CrossRef]
- Ijato, J.; Tedela, P. Phytotoxic Potentials of Cold and Hot Aqueous Extracts of Chromolaena odorata against Fungal Deteriorating Agents of Yam Tubers (Dioscorea rotundata, Poir) after Harvest. Am. J. Exp. Agric. 2015, 5, 262–266. [Google Scholar] [CrossRef]
- Mignouna, D.B.; Akinola, A.A.; Abdoulaye, T.; Alene, A.D.; Manyong, V.; Maroya, N.G.; Aighewi, B.A.; Kumar, L.P.; Balogun, M.; Lopez-Montes, A.; et al. Potential returns to yam research investment in sub-Saharan Africa and beyond. Outlook Agric. 2020, 49, 215–224. [Google Scholar] [CrossRef]
- Norman, P.E.; Paterne, A.A.; Danquah, A.; Tongoona, P.; Danquah, E.Y.; De Koeyer, D.; Ikeogu, U.N.; Asiedu, R.; Asfaw, A. Paternity Assignment in White Guinea Yam (Dioscorea rotundata) Half-Sib Progenies from Polycross Mating Design Using SNP Markers. Plants 2020, 9, 527. [Google Scholar] [CrossRef]
- Di-Giovanni, F.; Kevan, P.G. Factors affecting pollen dynamics and its importance to pollen contamination: A review. Can. J. For. Res. 1991, 21, 1155–1170. [Google Scholar] [CrossRef]
- Lebot, V.; Abraham, K.; Kaoh, J.; Rogers, C.; Molisalé, T. Development of anthracnose resistant hybrids of the Greater Yam (Dioscorea alata L.) and interspecific hybrids with D. nummularia Lam. Genet. Resour. Crop. Evol. 2019, 66, 871–883. [Google Scholar] [CrossRef]
- Akoroda, M.O. Floral biology in relation to hand pollination of white yam. Euphytica 1983, 32, 831–838. [Google Scholar] [CrossRef]
- Bakayoko, Y.; Kouakou, A.M.; Dibi, K.E.B.; Ehounou, A.E.; Essis, B.S.; Nzue, B.; Kouassi, A.B.; Nguetta, A.S.; Adebola, P.O. Effect of the manual pollination period on the fruiting of yams (Dioscorea rotundata Poir; Dioscoreaceae) in Ivory Coast. Eur. Sci. J. 2019, 15, 448–464. [Google Scholar]
- Akakpo, R.; Scarcelli, N.; Chaïr, H.; Dansi, A.; Djedatin, G.; Thuillet, A.-C.; Rhoné, B.; François, O.; Alix, K.; Vigouroux, Y. Molecular basis of African yam domestication: Analyses of selection point to root development, starch biosynthesis, and photosynthesis related genes. BMC Genom. 2017, 18, 782. [Google Scholar] [CrossRef] [PubMed]
- Asfaw, A.; Paterne, A.A.; Nwachukwu, C.; Eolasanmi, B.; Obidiegwu, J.E.; Nwachukwu, E.; Adebola, P.; De Koeyer, D. Sample Preservation and Plant Sex Prediction in White Guinea Yam (Dioscorea rotundata Poir.) Seedlings. J. Appl. Biot. Rep. 2020. [Google Scholar] [CrossRef]
- Sadik, S.; Okereke, O.U. Flowering, Pollen Grain Germination, Fruiting, Seed Germination and Seedling Development of White Yam, Dioscorea rotundata Poir. Ann. Bot. 1975, 39, 597–604. [Google Scholar] [CrossRef]
- Akoroda, M.O. Pollen management for controlled hybridization of white yam. Sci. Hortic. 1985, 25, 201–209. [Google Scholar] [CrossRef]
- Schmid, R.; Kubitzki, K.; Kadereit, J.W. The Families and Genera of Vascular Plants. TAXON 2005, 54, 574. [Google Scholar] [CrossRef]
- Tamiru, M.; Natsume, S.; Takagi, H.; White, B.; Yaegashi, H.; Shimizu, M.; Yoshida, K.; Uemura, A.; Oikawa, K.; Abe, A.; et al. Genome sequencing of the staple food crop white Guinea yam enables the development of a molecular marker for sex determination. BMC Biol. 2017, 15, 1–20. [Google Scholar] [CrossRef]
- Segnou; Fatokun, C.A.; Akoroda, M.O.; Hahn, S.K. Studies on the reproductive biology of white yam (Dioscorea rotundata Poir.). Euphytica 1992, 64, 197–203. [Google Scholar] [CrossRef]
- Zoundjihékpon, J.; Hamon, P.; Noirot, M.; Tio-Touré, B.; Hamon, S. Flowering synchronisation between male and female West African cultivated yams (Dioscorea cayenensis-rotundata complex). Euphytica 1997, 95, 371–375. [Google Scholar] [CrossRef]
- Abraham, K.; Nair, P.G. Floral biology and artificial pollination in Dioscorea alata L. Euphytica 1990, 48, 45–51. [Google Scholar]
- Girma, G.; Natsume, S.; Carluccio, A.V.; Takagi, H.; Matsumura, H.; Uemura, A.; Muranaka, S.; Stavolone, L.; Gedil, M.; Spillane, C.; et al. Identification of candidate flowering and sex genes in white Guinea yam (D. rotundata Poir.) by SuperSAGE transcriptome profiling. PLoS ONE 2019, 14, e0216912. [Google Scholar] [CrossRef]
- Wickens, G.E.; Onwueme, I.C. The Tropical Tuber Crops: Yams, Cassava, Sweet Potato and Cocoyams. Kew Bull. 1979, 34, 418. [Google Scholar] [CrossRef]
- Sadik, S.; Okereke, O.U. A new approach to improvement of yam Dioscorea rotundata. Nat. Cell Biol. 1975, 254, 134–135. [Google Scholar] [CrossRef]
- Abraham, K. Dormancy and viability of Dioscorea alata seeds. J. Root Crop 1992, 18, 142–143. [Google Scholar]
- Lebot, V. Tropical Root and Tuber Crops: Cassava, Sweet Potato, Yams and Aroids; CABI: Wallingford, UK, 2009; p. 413. [Google Scholar]
- Lopez-Montes, A.; Bhattacharjee, R.; Tessema, G. Yam breeding at IITA: Achievements, challenges and prospects. R4D Review Int. Inst. Trop. Agric. 2012, 8, 1–3. [Google Scholar]
- Bai, K.V.; Jos, J.S. Female fertility and seed set in Dioscorea alata L. Trop. Agric. 1986, 63, 7–10. [Google Scholar]
- Hamadina, E.I.; Craufurd, P.Q.; Asiedu, R. Flowering intensity in white yam (Dioscorea rotundata). J. Agric. Sci. 2009, 147, 469–477. [Google Scholar] [CrossRef]
- Akoroda, M.O.; Wilson, J.; Chheda, H.R. The association of sexuality with plant traits and tuber yield in white yam. Euphytica 1984, 33, 435–442. [Google Scholar] [CrossRef]
- Girma, G.; Hyma, K.E.; Asiedu, R.; Mitchell, S.E.; Gedil, M.; Spillane, C. Next-generation sequencing based genotyping, cytometry and phenotyping for understanding diversity and evolution of guinea yams. Theor. Appl. Genet. 2014, 127, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Abraham, K.; Nair, P.G. Polyploidy and sterility in relation to sex in Dioscorea alata L. (Dioscoreaceae). Genetica 1991, 83, 93–97. [Google Scholar] [CrossRef]
- Scarcelli, N.; Tostain, S.; Vigouroux, Y.; Agbangla, C.; Daïnou, O.; Pham, J.L. Farmers’ use of wild relative and sexual reproduction in a vegetatively propagated crop. The case of yam in Benin. Mol. Ecol. 2006, 15, 2421–2431. [Google Scholar] [CrossRef]
- Loko, Y.L.; Dansi, A.; Linsoussi, C.; Assogba, P.; Dansi, M.; Vodouhè, R.; Akoegninou, A.; Sanni, A. Current status and spatial analysis of Guinea yam (Dioscorea cayenensis Lam.-D. rotundata Poir. complex) diversity in Benin. Int. Res. J. Agric. Sci. Soil Sci. 2013, 3, 219–238. [Google Scholar]
- Arnau, G.; Abraham, K.; Sheela, M.; Chaïr, H.; Sartie, A.; Asiedu, R. Yams. Root and Tuber Crops; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2010; pp. 127–148. [Google Scholar]
- Araki, H.; Harada, T.; Yakuwa, T. Some Characteristics of Interspecific Hybrids between Dioscorea japonica Thunb. and Dioscorea opposita Thunb. J. Jpn. Soc. Hortic. Sci. 1983, 52, 153–158. [Google Scholar] [CrossRef]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. Plant Syst. Evol. 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Volk, G.M. Collecting Pollen for Genetic Resources Conservation. In Collecting Plant Genetic Diversity: Technical Guidelines 2011 Update; Guarino, L., Ramanatha, V.R., Goldberg, E., Eds.; Bioversity International: Rome, Italy, 2011; pp. 1–10. [Google Scholar]
- Impe, D.; Reitz, J.; Köpnick, C.; Rolletschek, H.; Börner, A.; Senula, A.; Nagel, M. Assessment of Pollen Viability for Wheat. Front. Plant Sci. 2020, 10, 10. [Google Scholar] [CrossRef]
- Alexander, L.W. Optimizing pollen germination and pollen viability estimates for Hydrangea macrophylla, Dichroa febrifuga, and their hybrids. Sci. Hortic. 2019, 246, 244–250. [Google Scholar] [CrossRef]
- Towill, L.E.; Walters, C. Cryopreservation of Pollen. In Cryopreservation of Tropical Plant Germplasm: Current Research Progress and Application; Englemann, F., Tagaki, H., Eds.; JIRCAS/IPGRI Co-Publication: Tsukuba, Japan, 2000; pp. 115–129. [Google Scholar]
- Brewbaker, J.L.; Kwack, B.H. The essential role of calcium ion in pollen germination and pollen tube growth. Am. J. Bot. 1963, 50, 859–865. [Google Scholar] [CrossRef]
- Akoroda, M. Long-term storage of yam pollen. Sci. Hortic. 1983, 20, 225–230. [Google Scholar] [CrossRef]
- Daniel, I.O.; Ng, N.Q.; Tayo, T.O.; Togun, A.O. Wet–cold preservation of West African yam (Dioscorea spp.) pollen. J. Agric. Sci. 2002, 138, 57–62. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, K.; Zhong, W.-P.; Chen, P.; Fan, X.; Yuan, D. Optimization of in vitro pollen germination and pollen viability tests for Castanea mollissima and Castanea henryi. Sci. Hortic. 2020, 271, 109481. [Google Scholar] [CrossRef]
- Ayenan, M.A.T.; Danquah, A.; Ampomah-Dwamena, C.; Hanson, P.; Asante, I.K.; Danquah, E.Y. Optimizing pollencounter for high throughput phenotyping of pollen quality in tomatoes. MethodsX 2020, 7, 100977. [Google Scholar] [CrossRef]
- Luria, G.; Rutley, N.; Lazar, I.; Harper, J.F.; Miller, G. Direct analysis of pollen fitness by flow cytometry: Implications for pollen response to stress. Plant J. 2019, 98, 942–952. [Google Scholar] [CrossRef]
- Ng, N.Q.; Daniel, I.O. Storage of pollens for long-term conservation of yam genetic resources. In Cryopreservation of Tropical Plant Germplasm: Current Research Progress and Application; Englemann, F., Tagaki, H., Eds.; JIRCAS/IPGRI Co-Publication: Tsukuba, Japan, 2000; pp. 136–139. [Google Scholar]
- Heidmann, I.; Schade-Kampmann, G.; Lambalk, J.; Ottiger, M.; Di Berardino, M. Impedance Flow Cytometry: A Novel Technique in Pollen Analysis. PLoS ONE 2016, 11, e0165531. [Google Scholar] [CrossRef]
- Tello, J.; Montemayor, M.I.; Forneck, A.; Ibáñez, J. A new image-based tool for the high throughput phenotyping of pollen viability: Evaluation of inter- and intra-cultivar diversity in grapevine. Plant Methods 2018, 14, 1–17. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Costa, C.M.; Yang, S. Counting pollen grains using readily available, free image processing and analysis software. Ann. Bot. 2009, 104, 1005–1010. [Google Scholar] [CrossRef]
- Kakui, H.; Yamazaki, M.; Hamaya, N.-B.; Shimizu, K.K. Pollen Grain Counting Using a Cell Counter. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2020; Volume 2160, pp. 1–11. [Google Scholar]
- Dinato, N.B.; Imaculada Santos, I.R.; Zanotto Vigna, B.B.; de Paula, A.F.; Fávero, A.P. Pollen Cryopreservation for Plant Breeding and Genetic Resources Conservation. Cryoletters 2020, 41, 115–127. [Google Scholar]
- Ren, R.; Li, Z.; Zhou, H.; Zhang, L.; Jiang, X.; Liu, Y. Changes in apoptosis-like programmed cell death and viability during the cryopreservation of pollen from Paeonia suffruticosa. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 357–368. [Google Scholar] [CrossRef]
- Funnekotter, B.; Colville, L.; Kaczmarczyk, A.; Turner, S.R.; Bunn, E.; Mancera, R.L. Monitoring of oxidative status in three native Australian species during cold acclimation and cryopreservation. Plant Cell Rep. 2017, 36, 1903–1916. [Google Scholar] [CrossRef]
- Mukherjee, A.; George, J.; Pillai, R.; Chakrabarti, S.K.; Naskar, S.K.; Patro, R.; Nayak, S.; Lebot, V. Development of taro (Colocasia esculenta (L.) Schott) hybrids overcoming its asynchrony in flowering using cryostored pollen. Euphytica 2016, 212, 29–36. [Google Scholar] [CrossRef]
- Streczynski, R.; Clark, H.; Whelehan, L.M.; Ang, S.-T.; Hardstaff, L.K.; Funnekotter, B.; Bunn, E.; Offord, C.A.; Sommerville, K.D.; Mancera, R.L. Current issues in plant cryopreservation and importance for ex situ conservation of threatened Australian native species. Aust. J. Bot. 2019, 67, 1–15. [Google Scholar] [CrossRef]
- Gaudet, D.; Yadav, N.S.; Sorokin, A.; Bilichak, A.; Kovalchuk, I. Development and Optimization of a Germination Assay and Long-Term Storage for Cannabis sativa Pollen. Plants 2020, 9, 665. [Google Scholar] [CrossRef]
- Akoroda, M.O.; Wilson, J.E.; Chheda, H.R. Artificial pollination, pollen viability, and storage in white yam. in Tropical Root Crops: Research strategies for the 1980s. In Proceedings of the 1st Triennial Root Crops Symposium of the International Society for Tropical Root Crops-Africa Branch, Ibadan, Nigeria, 8–12 September 1980; IDRC: Ottawa, ON, Canada, 1981. [Google Scholar]
- Nair, S.G.; Abraham, K.; Sreekumari, M.T. Stigma receptivity in white yam (Dioscorea rotundata, Poir.). In Proceedings of the National Symposium on Production and Utilization of Tropical Tuber Crops, Thiruvananthapuram, Kerala, India, 27–29 November 1985; pp. 71–74. [Google Scholar]
- Bulle-Legrand, M. Etude de la floraison de quatre espèces d’Ignames en vue d’une amélioration par la voie sexuée. Ph.D Thesis, Université de Paris-Sud, Orsay, France, 1983. [Google Scholar]
- Ehounou, A.E.; Kouakou, A.M.; N’zi, J.C.; Dibi, K.B.E.; Bakayoko, Y.; Essis, B.S.; N’Zue, B.; Arnau, G.; Maledon, E.; Asfaw, A.; et al. Production of hybrid seeds by intraspecific crossing in yam Dioscorea alata L. Int. J. Sci. Res. 2019, 8, 1212–1221. [Google Scholar]
- Otoo, E. Yam Breeding in Ghana. J. Agric. Sci. 2017, 9, 122. [Google Scholar] [CrossRef]
- Norman, P.E.; Asfaw, A.; Tongoona, P.B.; Danquah, A.; Danquah, E.Y.; De Koeyer, D.; Asiedu, R. Pollination success in some white yam genotypes under polycross and nested mating designs. Int. J. Biol. Sci. Appl. 2018, 5, 19–28. [Google Scholar]
- Acquaah, G. Principles of Plant Genetics and Breeding; Blackwell Publishing: Malden, MA, USA, 2012. [Google Scholar]
- Denadi, N.; Gandonou, C.; Missihoun, A.A.; Zoundjihékpon, J.; Quinet, M. Plant Sex Prediction Using Genetic Markers in Cultivated Yams (Dioscorea rotundata Poir.) in Benin. Agronomy 2020, 10, 1521. [Google Scholar] [CrossRef]
- Fu, L.; Wang, Q.; Zhang, M.; Zheng, Y.; Wu, M.; Lan, Z.; Pu, J.; Zhang, H.; Chen, F.; Su, W.; et al. Electrochemical Sex Determination of Dioecious Plants Using Polydopamine-Functionalized Graphene Sheets. Front. Chem. 2020, 8, 92. [Google Scholar] [CrossRef]
- Carputo, D. Cytological and breeding behavior of pentaploids derived from 3x × 4x crosses in potato. Theor. Appl. Genet. 2003, 106, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, J.E.; Bonierbale, M. Potatoes. In Root and Tuber Crops; Springer: New York, NY, USA, 2010; pp. 1–52. [Google Scholar]
- Seguí-Simarro, J.M. Androgenesis in Solanaceae. In Methods in Molecular Biology; Springer Science and Business Media LLC: Amsterdam, The Netherlands, 2016; Volume 1359, pp. 209–244. [Google Scholar]
- Lentini, Z.; González, Á.; Tabares, E.; Buitrago, M.E.; Wêdzony, M. Studies on Gynogenesis Induction in Cassava (Manihot esculenta Crantz) Unpollinated Ovule Culture. Front. Plant Sci. 2020, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Lentini, Z.; Restrepo, G.; Buitrago, M.E.; Tabares, E. Protocol for Rescuing Young Cassava Embryos. Front. Plant Sci. 2020, 11, 522. [Google Scholar] [CrossRef] [PubMed]
- Luthra, S.K.; Gupta, V.K.; Tiwari, J.K.; Kumar, V.; Bhardwaj, V.; Sood, S.; Dalamu; Kaur, R.P.; Kumar, R.; Vanishree, G.; et al. Potato Breeding in India; Technical Bulletin No 74; ICAR-Central Potato Research Institute: Shimla, Himachal, India, 2020; p. 214. [Google Scholar]
- Lebot, V. Sweet potato. In Root and Tuber Crops; Springer: New York, NY, USA, 2010; pp. 97–125. [Google Scholar]
- Wen, F.; Su, W.-P.; Zheng, H.; Yu, B.-C.; Ma, Z.-F.; Zhang, P.; Guo, W.-W. Plant regeneration via protoplast electrofusion in cassava. J. Integr. Agric. 2020, 19, 632–642. [Google Scholar] [CrossRef]
- Muthoni, J.; Shimelis, H.; Melis, R.; Kabira, J. Reproductive biology and early generation’s selection in conventional potato breeding. Austr. J. Crop Sci. 2012, 6, 488–497. [Google Scholar]
- Hernan, C.; Jaramillo, J.; Salazar, S.; Pineda, L.; Calle, F.; Setter, T. Induction of flowering in cassava through grafting. J. Plant Breed. Crop. Sci. 2017, 9, 19–29. [Google Scholar] [CrossRef]
- Ceballos, H.; Rojanaridpiched, C.; Phumichai, C.; Becerra, L.A.; Kittipadakul, P.; Iglesias, C.; Gracen, V.E. Excellence in Cassava Breeding: Perspectives for the Future. Crop. Breeding Genet. Genom. 2020, 2, 200008. [Google Scholar] [CrossRef]
- Souza, L.S.; Diniz, R.P.; Neves, R.D.J.; Alves, A.A.C.; De Oliveira, E.J. Grafting as a strategy to increase flowering of cassava. Sci. Hortic. 2018, 240, 544–551. [Google Scholar] [CrossRef]
- Pineda, M.; Morante, N.; Salazar, S.; Cuásquer, J.; Hyde, P.T.; Setter, T.L.; Ceballos, H. Induction of Earlier Flowering in Cassava through Extended Photoperiod. Agronomy 2020, 10, 1273. [Google Scholar] [CrossRef]
- Ye, M.; Peng, Z.; Tang, D.; Yang, Z.; Li, D.; Xu, Y.; Zhang, C.; Huang, S. Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 2018, 4, 651–654. [Google Scholar] [CrossRef]
- Quero-García, J.; Ivancic, A.; Lebot, V. Taro and Cocoyam. In Root and Tuber Crops; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2010; pp. 149–172. [Google Scholar]
- Hyde, P.T.; Guan, X.; Abreu, V.; Setter, T.L. The anti-ethylene growth regulator silver thiosulfate (STS) increases flower production and longevity in cassava (Manihot esculenta Crantz). Plant Growth Regul. 2020, 90, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Adeyemo, O.S.; Chavarriaga, P.; Tohme, J.; Fregene, M.; Davis, S.J.; Setter, T.L. Overexpression of Arabidopsis FLOWERING LOCUS T (FT) gene improves floral development in cassava (Manihot esculenta, Crantz). PLoS ONE 2017, 12, e0181460. [Google Scholar] [CrossRef] [PubMed]
- Odipio, J.; Getu, B.; Chauhan, R.D.; Alicai, T.; Bart, R.S.; Nusinow, D.A.; Taylor, N.J. Transgenic overexpression of endogenous FLOWERING LOCUS T-like gene MeFT1 produces early flowering in cassava. PLoS ONE 2020, 15, e0227199. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, H.; Quynh, D.T.N.; Anh, N.H.; Nhan, P.T.; Matsui, A.; Takahashi, S.; Tanaka, M.; Anh, N.M.; Van Dong, N.; Ham, L.H.; et al. Field transcriptome analysis reveals a molecular mechanism for cassava-flowering in a mountainous environment in Southeast Asia. Plant Mol. Biol. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pineda, M.; Yu, B.; Tian, Y.; Morante, N.; Salazar, S.; Hyde, P.T.; Setter, T.L.; Ceballos, H. Effect of Pruning Young Branches on Fruit and Seed Set in Cassava. Front. Plant Sci. 2020, 11, 1107. [Google Scholar] [CrossRef]
- Lentini, Z.; Tabares, E.; Buitrago, M.E. Vibratome Sectioning and Clearing for Easing Studies of Cassava Embryo Formation. Front. Plant Sci. 2020, 11, 1180. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.W.; Cabanillas, E.; Ortiz, S. Natural pollination, hand pollination and crossability of some Mexican species of Dioscorea. Trop. Agric. 1963, 40, 135–141. [Google Scholar]
- Howlett, B.G.; Gee, M. The potential management of the drone fly (Eristalis tenax) as a crop pollinator in New Zealand. N. Z. Plant Prot. 2019, 72, 221–230. [Google Scholar] [CrossRef]
- Nicholas, S.; Thyselius, M.; Holden, M.; Nordström, K. Rearing and Long-Term Maintenance of Eristalis tenax Hoverflies for Research Studies. J. Vis. Exp. 2018, 135, e57711. [Google Scholar] [CrossRef]
- Doyle, T.; Hawkes, W.L.S.; Massy, R.; Powney, G.D.; Menz, M.H.M.; Wotton, K.R. Pollination by hoverflies in the Anthropocene. Proc. R. Soc. B. Boil. Sci. 2020, 287, 20200508. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Observed Genotypes | % Non Flowering | % Flowering | % Male Flowers | % Female Flowers | % Monoecious | References |
|---|---|---|---|---|---|---|
| 391 | 55.2 | 44.8 | 35.6 | 9.2 | - | [20] |
| 1938 | 44.7 | 55.3 | 43.5 | 11.8 | 2.8 | [27] |
| 2946 | 24.3 | 75.7 | 29.9 | 35.5 | 10.3 | [16] |
| 190 | 46.3 | 53.7 | 33.7 | 15.8 | 4.2 | [19] |
| - | 40.0 | 60.0 | 37.0 | 18.0 | 5.0 | [19] * |
| 367 | 43.0 | 57.0 | 35.0 | 12.0 | 10.0 | [19] ** |
| - | 42.2 | 57.8 | 35.8 | 17.1 | 6.5 | - |
| Country | Species | Optimum Time for Pollination | % Pollination Success | References |
|---|---|---|---|---|
| India | D. rotundata | 9.00–12.00 noon | - | [66] |
| India | D. alata | 2.00 p.m. | - | [33] |
| Nigeria | D. rotundata | 12.00–2.00 p.m. | 28.0 | [16] |
| Côte d’Ivoire | D. rotundata D. cayenensis | 9.00 a.m.–1.00 p.m. | - | [25] |
| Guadeloupe | - | 9 a.m.–1 p.m. | - | [67] |
| India | D. alata | 12.00–3.00 p.m. | 74.5 | [26] |
| Côte d’Ivoire | D. rotundata | 7.30–11.00 a.m. | 8.3 | [17] |
| Côte d’Ivoire | D. alata | 7.30–11.00 a.m. | 19.7 | [68] |
| Nigeria | D. rotundata | 12.00–2.00 p.m. | 35.2 | [21] |
| Practices | Objectives | Species | References |
|---|---|---|---|
| Manipulation of ploidy with due regard to endosperm balance number (EBN) | To reduce cross-incompatibility among species | Solanum tuberosum L. | [74,75] |
| Double haploid technology through in vitro androgenesis or gynogenesis induction | To reduce cross-incompatibility among species. To develop inbred or homozygous lines in one step. | Solanum tuberosum L.; Manihot esculenta Crantz | [75,76,77] |
| Embryo rescue | To secure a hybrid where embryo abortion is due to a defective endosperm | Solanum tuberosum L.; Manihot esculenta Crantz | [75,78,79] |
| Mentor pollination | To achieve fertilization through a second pollination with compatible pollen to help incompatible pollen | Solanum tuberosum L. | [75,79] |
| Somatic (protoplast) fusion | To achieve difficult or impossible sexual hybridizations | Solanum tuberosum L.; Ipomoea batatas (L.) Lam; Manihot esculenta Crantz | [75,80,81] |
| In ploidy manipulation | To achieve maximum heterozygosity | Solanum tuberosum L. | [75] |
| Grafting | To induce intense flowering | Ipomoea batatas (L.) Lam.; Manihot esculenta Crantz; Solanum tuberosum L. | [79,80,82,83,84,85] |
| Reduced nitrogen fertilizer | To favor the flowering over lush and leafy vines | Ipomoea batatas (L.) Lam; Manihot esculenta Crantz | [80,86] |
| Gene editing using CRISPR–Cas9 system | To knock out the self-incompatibility gene S-RNase for potato. To incorporate PTST1 gene for earlier flowering in cassava. | Solanum tuberosum L.; Manihot esculenta Crantz | [84,87] |
| Treatment with growth regulators such as gibberellic acid, auxins, abscisic acid, ethylene, ascorbic acid, etc. | To induce earlier, profuse and long flowering | Colocasia esculenta L.; Manihot esculenta Crantz; Solanum tuberosum L. | [62,82,83,85,88,89] |
| Leaf removal | To induce artificial flowering | Colocasia esculenta L. | [88] |
| Drought and thermal stress induction | To induce artificial flowering | Colocasia esculenta L.; Manihot esculenta Crantz | [83,86,88] |
| Extended photoperiod using red light treatments at night | To induce early and profuse flowering | Manihot esculenta Crantz | [84,86] |
| Genetic transformation | To increase the level of FT genes; FLOWERING LOCUS T-like gene MeFT1 to induce early flowering | Manihot esculenta Crantz | [83,90,91,92] |
| Choice of location with longer photoperiods and cooler temperatures | To induce early and profuse flowering | Manihot esculenta Crantz; Solanum tuberosum L. | [79,86,92] |
| Application of the anti-ethylene growth regulator silver thiosulfate (STS) | To increase flower production and longevity; to reduce early abortion | Manihot esculenta Crantz | [89] |
| Pruning young branches soon after flowering | To prevent the common abortion of first inflorescences | Manihot esculenta Crantz | [93] |
| Spraying of benzyladenine in pruned plants for cassava or foliar spray with GA3 for potato | To promote flower development and feminization of male flowers | Manihot esculenta Crantz; Solanum tuberosum L. | [82,93] |
| Vibratome sectioning and clearing | To develop and optimize a methodology to induce doubled haploids via gynogenesis or from ovules pollinated with irradiated pollen | Manihot esculenta Crantz | [94] |
| Use of pollen mixture | To reduce cross incompatibility barriers among cultivars or species | Solanum tuberosum L. | [79] |
| Identification of matching parental groups | To reduce cross incompatibility barriers among cultivars or species | Solanum tuberosum L. | [79] |
| Increased plant density | To increase flowering intensity and seed weight | Solanum tuberosum L. | [82] |
| Shading of glasshouse to reduce temperature | To increase flowering intensity | Solanum tuberosum L. | [82] |
| Girdling or constriction of the stem | To increase flowering intensity | Solanum tuberosum L. | [79,82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mondo, J.M.; Agre, P.A.; Edemodu, A.; Adebola, P.; Asiedu, R.; Akoroda, M.O.; Asfaw, A. Floral Biology and Pollination Efficiency in Yam (Dioscorea spp.). Agriculture 2020, 10, 560. https://doi.org/10.3390/agriculture10110560
Mondo JM, Agre PA, Edemodu A, Adebola P, Asiedu R, Akoroda MO, Asfaw A. Floral Biology and Pollination Efficiency in Yam (Dioscorea spp.). Agriculture. 2020; 10(11):560. https://doi.org/10.3390/agriculture10110560
Chicago/Turabian StyleMondo, Jean M., Paterne A. Agre, Alex Edemodu, Patrick Adebola, Robert Asiedu, Malachy O. Akoroda, and Asrat Asfaw. 2020. "Floral Biology and Pollination Efficiency in Yam (Dioscorea spp.)" Agriculture 10, no. 11: 560. https://doi.org/10.3390/agriculture10110560
APA StyleMondo, J. M., Agre, P. A., Edemodu, A., Adebola, P., Asiedu, R., Akoroda, M. O., & Asfaw, A. (2020). Floral Biology and Pollination Efficiency in Yam (Dioscorea spp.). Agriculture, 10(11), 560. https://doi.org/10.3390/agriculture10110560