
Genetic Polymorphism of Zinc Transporter-8 Gene (SLC30A8), Serum Zinc Concentrations, and Proteome Profiles Related to Type 2 Diabetes in Elderly

 , , , , ,
, , , , ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Biochemical Analysis
2.3. Genotyping
2.4. Proteomic Analysis
2.5. Statistical Analysis
3. Results
3.1. Analyses of Demographic, Clinical, and Biochemical Characteristics of Study Groups
3.2. Associations Between SLC30A8 SNPs, HbA1C Level, Metabolic Syndrome, and Serum Zinc Tertiles in Nondiabetic and Prediabetic/Diabetic Groups
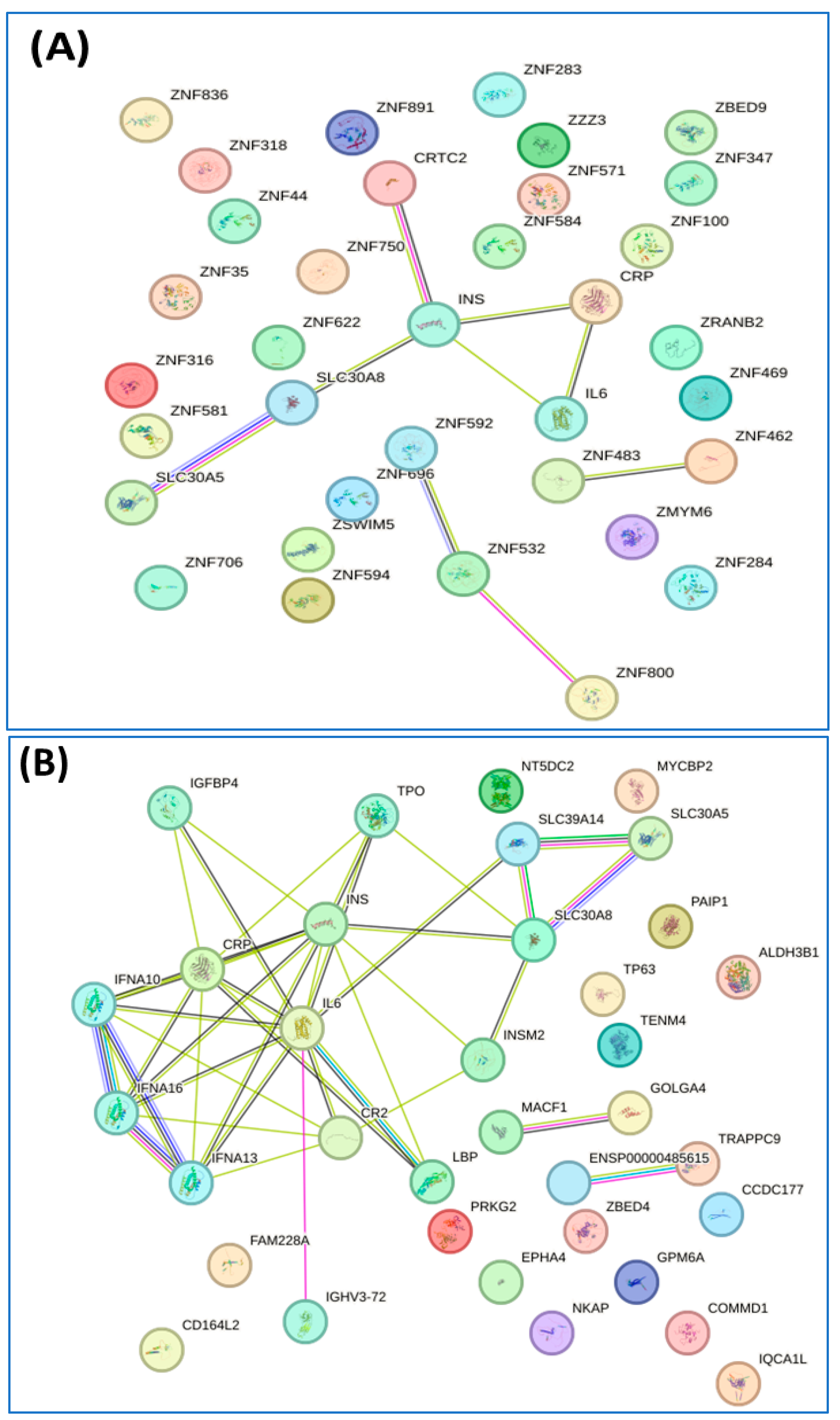
3.3. Proteomic Profiles Related to Diabetes and Metabolic Syndrome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiouri, D.P.; Tsoupra, E.; Peana, M.; Perlepes, S.P.; Stefanidou, M.E.; Chasapis, C.T. Multifunctional role of zinc in human health: An update. Excli. J. 2023, 22, 809–827. [Google Scholar] [PubMed]
- Costa, M.I.; Sarmento-Ribeiro, A.B.; Gonçalves, A.C. Zinc: From biological functions to therapeutic potential. Int. J. Mol. Sci. 2023, 24, 4822. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Shaju, R.; Atfi, A.; Razzaque, M.S. Zinc and diabetes: A connection between micronutrient and metabolism. Cells 2024, 13, 1359. [Google Scholar] [CrossRef]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144–157. [Google Scholar]
- Nagao, H.; Cai, W.; Brandão, B.B.; Wewer Albrechtsen, N.J.; Steger, M.; Gattu, A.K.; Pan, H.; Dreyfuss, J.M.; Wunderlich, F.T.; Mann, M.; et al. Leucine-973 is a crucial residue differentiating insulin and IGF-1 receptor signaling. J. Clin. Investig. 2023, 133, e161472. [Google Scholar] [CrossRef]
- Gleason, C.E.; Lu, D.; Witters, L.A.; Newgard, C.B.; Birnbaum, M.J. The role of AMPK and mTOR in nutrient sensing in pancreatic beta-cells. J. Biol. Chem. 2007, 282, 10341–10351. [Google Scholar] [CrossRef]
- Safarzad, M.; Jazi, M.S.; Kiaei, M.; Asadi, J. Lower serum zinc level is associated with higher fasting insulin in type 2 diabetes mellitus (T2DM) and relates with disturbed glucagon suppression response in male patients. Prim. Care Diabetes 2023, 17, 493–498. [Google Scholar] [CrossRef]
- Sonkar, S.K.; Parmar, K.S.; Ahmad, M.K.; Sonkar, G.K.; Gautam, M. An observational study to estimate the level of essential trace elements and its implications in type 2 diabetes mellitus patients. J. Family Med. Prim. Care 2021, 10, 2594–2599. [Google Scholar] [CrossRef]
- Fernández-Cao, J.C.; Warthon-Medina, M.; Hall Moran, V.; Arija, V.; Doepking, C.; Lowe, N.M. Dietary zinc intake and whole blood zinc concentration in subjects with type 2 diabetes versus healthy subjects: A systematic review, meta-analysis and meta-regression. J. Trace Elem. Med. Biol. 2018, 49, 241–251. [Google Scholar] [CrossRef]
- Yi, B.; Huang, G.; Zhou, Z. Different role of zinc transporter 8 between type 1 diabetes mellitus and type 2 diabetes mellitus. J. Diabetes Investig. 2016, 7, 459–465. [Google Scholar] [CrossRef]
- Goyal, Y.; Verma, A.K.; Joshi, P.C.; Dev, K. Contemplating the role of genetic variants of HHEX, CDKAL1, WFS1 and SLC30A8 genes of TYPE-2 diabetes in Asians ethnic groups. Gene Rep. 2019, 17, 100465. [Google Scholar] [CrossRef]
- Sladek, R.; Rocheleau, G.; Rung, J.; Dina, C.; Bell, C.G.; Paré, G.; Hudson, T.J.; Hirschhorn, J.N.; Rioux, J.D.; Lathrop, G.M.; et al. A Genome-Wide Association Study Identifies Novel Risk Loci for Type 2 Diabetes. Nature 2007, 445, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Zeggini, E.; Weedon, M.N.; Lindgren, C.M.; Frayling, T.M.; Elliott, K.S.; Lango, H.; Timpson, N.J.; Perry, J.R.; Rayner, N.W.; Freathy, R.M.; et al. Replication of Genome-Wide Association Signals in UK Samples Reveals Risk Loci for Type 2 Diabetes. Science 2007, 316, 1336–1341. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Bao, W.; Zhang, Y.; Rong, Y.; Wang, X.; Jin, Y.; Song, Y.; Yao, P.; Sun, C.; Hu, F.B.; et al. Interactions Between Zinc Transporter-8 Gene (SLC30A8) and Plasma Zinc Concentrations for Impaired Glucose Regulation and Type 2 Diabetes. Diabetes 2014, 63, 1796–1803. [Google Scholar] [CrossRef]
- Mashal, S.; Khanfar, M.; Al-Khalayfa, S.; Srour, L.; Mustafa, L.; Hakooz, N.M.; Zayed, A.A.; Khader, Y.S.; Azab, B.A. SLC30A8 Gene Polymorphism rs13266634 Associated with Increased Risk for Developing Type 2 Diabetes Mellitus in Jordanian Population. Gene 2021, 768, 145279. [Google Scholar]
- Yanasegaran, K.; Ng, J.Y.E.; Chua, E.W.; Nawi, A.M.; Ng, P.Y.; Abdul Manaf, M.R. Single Nucleotide Polymorphisms (SNPs) That Are Associated with Obesity and Type 2 Diabetes Among Asians: A Systematic Review and Meta-Analysis. Sci. Rep. 2024, 14, 20062. [Google Scholar] [CrossRef]
- Guan, H.; Zhao, S.; Li, J.; Wang, Y.; Niu, P.; Zhang, Y.; Zhang, Y.; Fang, X.; Miao, R.; Tian, J. Exploring the Design of Clinical Research Studies on the Efficacy Mechanisms in Type 2 Diabetes Mellitus. Front. Endocrinol. 2024, 15, 1363877. [Google Scholar] [CrossRef]
- Nimer, R.M.; Alfaqih, M.A.; Shehabat, E.R.; Mujammami, M.; Abdel Rahman, A.M. Label-Free Quantitative Proteomics Analysis for Type 2 Diabetes Mellitus Early Diagnostic Marker Discovery Using DIA-MS. Sci. Rep. 2023, 13, 20880. [Google Scholar] [CrossRef]
- Ngo, D.; Benson, M.D.; Long, J.Z.; Chen, Z.Z.; Wang, R.; Nath, A.K.; Keyes, M.J.; Shen, D.; Sinha, S.; Kuhn, E.; et al. Proteomic Profiling Reveals Biomarkers and Pathways in Type 2 Diabetes Risk. JCI Insight 2021, 6, e144392. [Google Scholar] [CrossRef]
- Grider, A.; Wickwire, K.; Ho, E.; Chung, C.S.; King, J. Dietary Zinc Depletion and Repletion Affects Plasma Proteins: An Analysis of the Plasma Proteome. Biometals 2013, 26, 133–140. [Google Scholar]
- Wang, M.; Fang, L.; Wang, W.; Cui, H.; Zhang, H.; Fan, Z.; Ji, F.; Tang, H. Integrated Multi-Omics Uncovers Reliable Potential Biomarkers and Adverse Effects of Zinc Deficiency. Clin. Nutr. 2021, 40, 2683–2696. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Obesity: Preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ. Tech. Rep. Ser. 2000, 894, 1–253.
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2014, 37 (Suppl. S1), S81–S90. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C.; et al. Diagnosis and Management of the Metabolic Syndrome: An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef] [PubMed]
- Krachler, M.; Irgolic, K.J. The Potential of ICP-MS for the Simultaneous Determination of Trace Elements in Whole Blood, Plasma, and Serum. J. Trace Elem. Med. Biol. 1999, 13, 157–169. [Google Scholar] [CrossRef]
- Miller, S.S.; Davis, P. A Simple Salting Out Procedure for Extracting DNA from Human Nucleated Cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Kerdsaeng, N.; Roytrakul, S.; Chanprasertyothin, S.; Charernwat, P.; Chansirikarnjana, S.; Sritara, P.; Sirivarasai, J. Serum Glycoproteomics and Identification of Potential Mechanisms Underlying Alzheimer’s Disease. Behav. Neurol. 2021, 2021, 1434076. [Google Scholar]
- Tansakul, N.; Rattanasrisomporn, J.; Roytrakul, S. Proteomics Analysis of Serum Protein Patterns in Ducks During Aflatoxin B1 Exposure. Vet. World 2019, 12, 1499–1505. [Google Scholar]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jVenn: An Interactive Venn Diagram Viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar]
- Ortega, Á.; Berná, G.; Rojas, A.; Martín, F.; Soria, B. Gene-Diet Interactions in Type 2 Diabetes: The Chicken and Egg Debate. Int. J. Mol. Sci. 2017, 18, 1188. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, L.; Yu, D.; Ding, G. The Prevalence and Risk Factors of Dyslipidemia in Different Diabetic Progression Stages Among Middle-Aged and Elderly Populations in China. PLoS ONE 2018, 13, e0205709. [Google Scholar] [CrossRef]
- Stanimirovic, J.; Radovanovic, J.; Banjac, K.; Obradovic, M.; Essack, M.; Zafirovic, S.; Gluvic, Z.; Gojobori, T.; Isenovic, E.R. Role of C-Reactive Protein in Diabetic Inflammation. Mediators Inflamm. 2022, 2022, 3706508. [Google Scholar] [CrossRef]
- Feng, X.; Xu, Y. Hyperhomocysteinemia as a Metabolic Risk Factor for Glucose Intolerance Among High-Risk Groups of Chinese Adults. Med. Sci. Monit. 2017, 23, 2775–2781. [Google Scholar] [CrossRef] [PubMed]
- Dicker-Brown, A.; Fonseca, V.A.; Fink, L.M.; Kern, P.A. The Effect of Glucose and Insulin on the Activity of Methylene Tetrahydrofolate Reductase and Cystathionine-β-Synthase: Studies in Hepatocytes. Atherosclerosis 2001, 158, 297–301. [Google Scholar]
- Nandi, S.; Sivaprasad, M.V. Homocysteine Thiolactone Inhibits Insulin Signaling, and Glutathione Has a Protective Effect. J. Mol. Endocrinol. 2001, 27, 85–91. [Google Scholar]
- Lemaire, K.; Chimienti, F.; Schuit, F. Zinc Transporters and Their Role in the Pancreatic β-Cell. J. Diabetes Investig. 2012, 3, 202–211. [Google Scholar]
- Palmiter, R.D.; Huang, L. Efflux and Compartmentalization of Zinc by Members of the SLC30 Family of Solute Carriers. Pflugers Arch. 2004, 447, 744–751. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, R.; Xu, Y.; Wang, X.; Pan, Z.; Jin, X.; Bai, Y.; Zhang, Z.; Li, F.; Gao, T.; et al. Glucose Potentiates β-Cell Function by Inducing Tph1 Expression in Rat Islets. FASEB J. 2017, 31, 5342–5355. [Google Scholar] [CrossRef]
- Koo, S.H.; Flechner, L.; Qi, L.; Zhang, X.; Screaton, R.A.; Jeffries, S.; Hedrick, S.; Xu, W.; Boussouar, F.; Brindle, P.; et al. The CREB Coactivator TORC2 Is a Key Regulator of Fasting Glucose Metabolism. Nature 2005, 437, 1109–1111. [Google Scholar]
- Bensellam, M.; Laybutt, D.R.; Jonas, J.C. Emerging Roles of Metallothioneins in Beta-Cell Pathophysiology: Beyond and Above Metal Homeostasis and Antioxidant Response. Biology 2021, 10, 176. [Google Scholar] [CrossRef]
- Chung, L. Metallothionein as an Adaptive Protein Prevents Diabetes and Its Toxicity. Nonlinearity Biol. Toxicol. Med. 2004, 2, 89–103. [Google Scholar]
- Rodríguez-Zavala, J.S.; Calleja, L.F.; Moreno-Sánchez, R.; Yoval-Sánchez, B. Role of Aldehyde Dehydrogenases in Physiopathological Processes. Chem. Res. Toxicol. 2019, 32, 405–420. [Google Scholar] [CrossRef]
- Wu, Z.; Cao, A.; Zhang, G.; Lin, C.; Yang, S.; Shen, R.; Ma, J.; Gao, T.; Wang, Y.; Liu, J.; et al. ALDH3B1 Protects Interfollicular Epidermal Cells Against Lipid Peroxidation via the NRF2 Pathway. Cell Stress Chaperones 2022, 27, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; D’Souza, D. Role of NF-κB in the Pathogenesis of Diabetes and Its Associated Complications. Pharmacol. Rep. 2009, 61, 595–603. [Google Scholar] [CrossRef]
- Zhou, X.; Xing, A.; Yao, C.; Wang, S.; Jin, H.; Li, F. The Crucial Role and Mechanism of Insulin Resistance in Metabolic Disease. Front. Endocrinol. 2023, 14, 1149239. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Bizzarri, M. Inositols in Insulin Signaling and Glucose Metabolism. Int. J. Endocrinol. 2018, 2018, 1968450. [Google Scholar]
- Shimizu, I.; Yoshida, Y.; Suda, M.; Minamino, T. DNA Damage Response and Metabolic Disease. Cell Metab. 2014, 20, 967–977. [Google Scholar]
- Ruz, M.; Carrasco, F.; Rojas, P.; Basfi-Fer, K.; Hernández, M.C.; Pérez, A. Nutritional Effects of Zinc on Metabolic Syndrome and Type 2 Diabetes: Mechanisms and Main Findings in Human Studies. Biol. Trace Elem. Res. 2019, 188, 177–188. [Google Scholar]
- Pfeifer, A.; Kilić, A.; Hoffmann, L.S. Regulation of Metabolism by cGMP. Pharmacol. Ther. 2013, 140, 81–91. [Google Scholar]
- Muzurović, E.; Kraljević, I.; Solak, M.; Dragnić, S.; Mikhailidis, D.P. Homocysteine and Diabetes: Role in Macrovascular and Microvascular Complications. J. Diabetes Complicat. 2021, 35, 107834. [Google Scholar] [CrossRef]
- Vakilian, M.; Tahamtani, Y.; Ghaedi, K. A Review on Insulin Trafficking and Exocytosis. Gene 2019, 706, 52–61. [Google Scholar]
- Varney, M.J.; Benovic, J.L.; Michel, M. The Role of G Protein-Coupled Receptors and Receptor Kinases in Pancreatic β-Cell Function and Diabetes. Pharmacol. Rev. 2024, 76, 267–299. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total (n = 265) | Non-Diabetes Group (n = 116) | Prediabetes/Diabetes Group (n = 149) |
|---|---|---|---|
| Age (years) | 67.38 ± 3.88 | 67.21 ± 3.47 | 67.52 ± 4.18 |
| Male (n, %) | 183 (69.1%) | 78 (67.2%) | 105 (70.5%) |
| BMI (kg/m2) | 23.45 ± 3.68 | 22.37 ± 3.40 | 24.28 ± 3.69 a |
| Waist circumference (cm) | 85.33 ± 10.55 | 82.19 ± 9.73 | 87.69 ± 10.56 a |
| Waist–hip ratio | 94.36 ± 7.26 | 92.10 ± 6.68 | 96.08 ± 7.22 a |
| Systolic blood pressure (mmHg) | 132.77 ± 20.92 | 132.40 ± 23.52 | 133.05 ± 18.74 |
| Diastolic blood pressure (mmHg) | 78.51 ± 11.65 | 76.94 ± 11.53 | 79.73 ± 11.64 |
| Fasting plasma glucose (mg/dL) | 93.65 ± 22.37 | 87.86 ± 8.08 | 119.68 ± 8.31 a |
| HbA1C (%) | 5.92 ± 1.03 | 5.38 ± 0.23 | 6.71 ± 0.79 a |
| Triglyceride (mg/dL) | 117.04 ± 76.00 | 111.37 ± 95.39 | 158.25 ± 47.59 a |
| Total cholesterol (mg/dL) | 229.26 ± 43.08 | 224.16 ± 43.53 | 233.24 ± 42.45 |
| Low-density lipoprotein cholesterol (mg/dL) | 129.37 ± 32.68 | 123.25 ± 31.21 | 135.18 ± 38.77 a |
| High-density lipoprotein cholesterol (mg/dL) | 64.13 ± 17.41 | 66.97 ± 18.82 | 61.93 ± 15.95 a |
| Albumin (g/dL) | 4.51 ± 0.25 | 4.49 ± 0.25 | 4.53 ± 0.24 |
| Alanine transaminase (U/L) | 19.73 ± 9.78 | 18.80 ± 8.55 | 20.46 ± 10.60 |
| Aspartate transaminase (U/L) | 22.799 ± 8.92 | 22.47 ± 7.22 | 23.04 ± 10.07 |
| Creatinine (mg/dL) | 0.89 ± 0.16 | 0.86 ± 0.11 | 0.92 ± 0.14 |
| Blood urea nitrogen (mg/dL) | 13.09 ± 2.78 | 12.92 ± 2.82 | 13.22 ± 2.74 |
| Uric acid (mg/dL) | 5.70 ± 1.30 | 5.53 ± 1.17 | 5.84 ± 1.28 |
| hs-C-reactive protein * (mg/L) | 1.31 ± 0.96 | 1.06 ± 0.89 | 2.35 ± 1.13 a |
| Homocysteine (mmol/L) | 16.89 ± 3.74 | 13.08 ± 3.64 | 17.62 ± 3.97 a |
| Serum zinc * (µg/L) | 708.09 ± 126.57 | 742.03 ± 109.25 | 602 ± 171.39 a |
| SNP1 (SLC30A8 rs13266634) Allele Frequencies | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| All subjects (n = 265) | Non-diabetes (n = 116) | Prediabetes/diabetes (n = 149) | |||||||
| Allele | Count | Proportion | Count | Proportion | Count | Proportion | |||
| T | 305 | 0.58 | 125 | 0.54 | 180 | 0.6 | |||
| C | 225 | 0.42 | 107 | 0.46 | 118 | 0.4 | |||
| Genotype | |||||||||
| T/T | 91 | 0.34 | 36 | 0.31 | 55 | 0.37 | |||
| T/C | 123 | 0.46 | 53 | 0.46 | 70 | 0.47 | |||
| C/C | 51 | 0.19 | 27 | 0.23 | 24 | 0.16 | |||
| SNP1 exact test for Hardy–Weinberg equilibrium (n = 265) | |||||||||
| N11 | N12 | N22 | N1 | N2 | p-value | ||||
| All subjects | 91 | 123 | 51 | 305 | 225 | 0.45 | |||
| Non-diabetes | 36 | 53 | 27 | 125 | 107 | 0.45 | |||
| Prediabetes/diabetes | 55 | 70 | 24 | 180 | 118 | 0.86 | |||
| SNP1 association with HbA1C (n = 265, adjusted by group) | |||||||||
| Model | Genotype | n | Response mean (s.e.) | Difference (95% CI) | p-value | AIC | BIC | ||
| Codominant | T/T | 91 | 5.85 (0.07) | 0.00 | < 0.0001 | 583.2 | 601.1 | ||
| T/C | 123 | 5.7 (0.03) | −0.12 (−0.31–0.08) | ||||||
| C/C | 51 | 6.3 (0.23) | 0.57 (0.32–0.81) | ||||||
| Dominant | T/T | 91 | 5.85 (0.07) | 0.00 | 0.42 | 612.3 | 626.6 | ||
| T/C-C/C | 174 | 5.88 (0.07) | 0.08 (−0.11–0.27) | ||||||
| Recessive | T/T-T/C | 214 | 5.77 (0.03) | 0.00 | < 0.0001 | 582.6 | 596.9 | ||
| C/C | 51 | 6.3 (0.23) | 0.63 (0.41–0.86) | ||||||
| Overdominant | T/T-C/C | 142 | 6.01 (0.09) | 0.00 | 0.0005 | 600.8 | 615.2 | ||
| T/C | 123 | 5.7 (0.03) | −0.32 (−0.50–−0.14) | ||||||
| Interaction analysis with covariate serum zinc tertiles (n = 265) | |||||||||
| T1 | T2 | T3 | |||||||
| n | HbA1C mean (s.e.) | Difference (95% CI) | n | HbA1C mean (s.e.) | Difference (95% CI) | n | HbA1C mean (s.e.) | Difference (95% CI) | |
| T/T | 23 | 5.95 (0.14) | 0.00 | 34 | 5.76 (0.07) | −0.11 (−0.48–0.26) | 34 | 5.89 (0.14) | −0.03 (−0.40–0.34) |
| T/C | 42 | 5.8 (0.06) | −0.14 (−0.49–0.21) | 40 | 5.65 (0.05) | −0.19 (−0.55–0.17) | 41 | 5.66 (0.05) | −0.19 (−0.55–0.17) |
| C/C | 23 | 6.92 (0.47) | 1.01 (0.61–1.41) | 15 | 5.94 (0.16) | 0.09 (−0.36–0.55) | 13 | 5.62 (0.13) | 0.09 (−0.39–0.57) |
| Interaction p-value: 0.0048 | |||||||||
| SNP Association with Metabolic Syndrome (n = 265), Adjusted by Prediabetes/Diabetes Status, HbA1C, TC, TG, HDL, LDL, FBG, Hcy, hsCRP, Serum Zinc, and Tertile of Serum Zinc | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Model | Genotype | No MS | MS | OR (95% CI) | p-Value | AIC | BIC | ||
| Codominant | T/T | 66 (33%) | 25 (38.5%) | 1.00 | 0.0056 | 214.4 | 268.1 | ||
| T/C | 103 (51.5%) | 20 (30.8%) | 0.73 (0.30–1.79) | ||||||
| C/C | 31 (15.5%) | 20 (30.8%) | 4.21 (1.39–12.78) | ||||||
| Dominant | T/T | 66 (33%) | 25 (38.5%) | 1.00 | 0.62 | 222.5 | 272.7 | ||
| T/C-C/C | 134 (67%) | 40 (61.5%) | 1.22 (0.55–2.72) | ||||||
| Recessive | T/T-T/C | 169 (84.5%) | 45 (69.2%) | 1.00 | 0.0016 | 212.9 | 263 | ||
| C/C | 31 (15.5%) | 20 (30.8%) | 5.00 (1.82–13.71) | ||||||
| Overdominant | T/T-C/C | 97 (48.5%) | 45 (69.2%) | 1.00 | 0.055 | 219.1 | 269.2 | ||
| T/C | 103 (51.5%) | 20 (30.8%) | 0.46 (0.21–1.03) | ||||||
| Interaction analysis of SNP with tertiles of serum zinc level in association with metabolic syndrome (n = 265), adjusted by prediabetes/diabetes status, HbA1C, TC, TG, HDL, LDL, FBG, Hcy, hsCRP, and serum zinc) | |||||||||
| Tertile 1 of serum zinc | Tertile 2 of serum zinc | Tertile 3 of serum zinc | |||||||
| No Ms | MS | OR (95% CI) | No MS | MS | OR (95% CI) | No MS | MS | OR (95% CI) | |
| T/T | 16 | 7 | 1.00 | 25 | 9 | 6.58 (0.99–43.67) | 25 | 9 | 1.46 (0.16–13.18) |
| T/C | 30 | 12 | 4.51 (0.85–23.91) | 39 | 1 | 0.18 (0.01–3.06) | 34 | 7 | 2.03 (0.21–19.87) |
| C/C | 14 | 9 | 18.92 (2.47–145.08) | 10 | 5 | 5.12 (0.56–47.23) | 10 | 3 | 4.52 (0.33–62.35) |
| Interaction p-value: 0.0052 | |||||||||
| ↑ Upregulated Proteins (n = 40) | ↓ Downregulated Proteins (n = 29) | |||||
|---|---|---|---|---|---|---|
| Protein ID | Protein Name | log2 (FC) | Protein ID | Protein Name | log2 (FC) | |
| 1 | Q6ZNA1 | Zinc finger protein 836 | 2.522 | P08069 | Insulin-like growth factor 1 receptor | −1.126 |
| 2 | A0A1U9X8W9 | ZBED9 | 2.506 | B4DUR8 | T-complex protein 1 subunit gamma | −1.209 |
| 3 | Q5VUA4 | Zinc finger protein 318 | 2.506 | A0A0G2JLC4 | Lipopolysaccharide-induced TNF factor | −1.296 |
| 4 | Q9P0T4 | Zinc finger protein 581 | 2.505 | C7DUW4 | mitogen-activated protein kinase kinase | −1.337 |
| 5 | Q96JF6 | Zinc finger protein 594 | 2.505 | A0A087WWY0 | deleted | −1.406 |
| 6 | D3DSZ2 | deleted | 2.505 | A0A087X2D4 | Aldehyde dehydrogenase 3 family member B1 | −2.573 |
| 7 | A0A494C1V2 | Zinc finger protein 891 | 2.502 | Q9NQR7 | Coiled coil domain-containing protein 177 | −2.580 |
| 8 | K7EQN0 | Zinc finger protein 532 | 2.502 | Q6UWJ8 | CD164 sialomucin-like 2 protein | −2.581 |
| 9 | Q9P217 | Zinc finger SWIM domain-containing protein 5 | 2.501 | H7C169 | Copper metabolism domain containing 1 | −2.585 |
| 10 | O95218 | Zinc finger Ran-binding domain-containing protein 2 | 2.497 | Q5SR47 | Complement C3d receptor 2 | −2.585 |
| 11 | A0A0G2JMF9 | Zinc finger protein 705G | 2.497 | F5GZZ5 | Receptor protein-tyrosine kinase | −2.590 |
| 12 | C9J283 | Zinc finger ZZ-type containing 3 | 2.497 | H7C4B8 | Family with sequence similarity 228 member A | −2.591 |
| 13 | K7ELU5 | Zinc finger protein 571 | 2.496 | A0A7U3JVZ5 | Fibroblast growth factor | −2.592 |
| 14 | X6RCN5 | Zinc finger MYM-type containing 6 | 2.496 | C9JHJ5 | Golgin A4 | −2.594 |
| 15 | Q5VZN3 | Zinc finger protein 483 | 2.495 | D6R9D2 | Neuronal membrane glycoprotein M6-a | −2.594 |
| 16 | P15621 | Zinc finger protein 44 | 2.494 | A0A087WVA7 | IQ motif containing with AAA domain 1 like | −2.598 |
| 17 | H3BS19 | Zinc finger protein 469 | 2.493 | E9PNZ4 | Microtubule actin crosslinking factor 1 | −2.601 |
| 18 | C9JGR2 | Zinc finger protein 35 | 2.493 | A0A499FJI4 | RCR-type E3 ubiquitin transferase | −2.604 |
| 19 | M0R2W6 | Zinc finger protein 584 | 2.491 | Q8N5F7 | NF-kappa-B-activating protein | −2.606 |
| 20 | A6NEH8 | ZNF503-AS2 | 2.490 | A0A024R250 | deleted | −2.606 |
| 21 | Q969S3 | Cytoplasmic 60S subunit biogenesis factor ZNF622 | 2.490 | Q9H857 | 5′-nucleotidase domain-containing protein 2 | −2.607 |
| 22 | Q8IYN0 | Zinc finger protein 100 | 2.490 | D6REB4 | Poly(A) binding protein interacting protein 1 | −2.608 |
| 23 | M0QZE2 | Zinc finger protein 347 | 2.488 | A0A024R930 | deleted | −2.612 |
| 24 | A0A7P0N7C4 | Zinc finger protein 142 | 2.487 | A0A140VJM3 | cGMP-dependent protein kinase | −2.612 |
| 25 | H3BLX4 | Zinc finger protein 462 | 2.486 | B7ZLP5 | SAFB protein | −2.617 |
| 26 | A0A494C0U8 | Zinc finger protein 283 | 2.483 | Q6N022 | Teneurin−4 (Ten−4) | −2.634 |
| 27 | Q32MQ0 | Zinc finger protein 750 | 2.482 | C9D7D0 | Cellular tumor antigen p53 | −2.643 |
| 28 | B9EH69 | ZNF658 protein | 2.480 | A0A0J9YWK7 | Trafficking protein particle complex subunit 9 | −2.644 |
| 29 | B3VRW5 | Tryptophan hydroxylase 1 | 2.477 | O75132 | Zinc finger BED domain-containing protein 4 | −2.737 |
| 30 | Q2TB10 | Zinc finger protein 800 | 2.477 | |||
| 31 | Q5T4K5 | CREB regulated transcription coactivator 2 | 2.476 | |||
| 32 | Q92610 | Zinc finger protein 592 | 2.474 | |||
| 33 | H0YC70 | Zinc finger protein 706 | 2.470 | |||
| 34 | A6NFI3 | Zinc finger protein 316 | 2.470 | |||
| 35 | Q2VY69 | Zinc finger protein 284 | 2.464 | |||
| 36 | E5RG39 | Zinc finger protein 696 | 2.462 | |||
| 37 | B2RN90 | Zinc finger protein 776 | 2.453 | |||
| 38 | P47944 | Metallothionein-4 (MT-4) | 1.236 | |||
| 39 | I1Y8W7 | Sirtuin 1 | 1.213 | |||
| 40 | A0A2R8Y7I7 | Glutathione synthetase | 1.002 | |||
| ↑ Upregulated Proteins (n = 19) | ↓ Downregulated Proteins (n = 31) | |||||
|---|---|---|---|---|---|---|
| Protein ID | Protein Names | log2 (FC) | Protein ID | Protein Names | log2 (FC) | |
| 1 | A0A087X2D4 | Aldehyde dehydrogenase 3 family member B1 | 2.839 | M0R1K5 | NOP2/Sun RNA methyltransferase 4 | −2.459 |
| 2 | Q6UWJ8 | CD164 sialomucin-like 2 protein | 2.633 | A0A6Q8PHP9 | Phospholipase C epsilon 1 | −2.460 |
| 3 | C9D7D0 | Cellular tumor antigen p53 | 2.631 | H0YER2 | Activating signal cointegrator 1 complex subunit 1 | −2.468 |
| 4 | A0A140VJM3 | cGMP-dependent protein kinase | 2.630 | A6N6J7 | [histone H3]-trimethyl-L-lysine(4) demethylase | −2.469 |
| 5 | Q9NQR7 | Coiled-coil domain-containing protein 177 | 2.627 | F2Z3J2 | Proteasome 26S subunit, non-ATPase 5 | −2.471 |
| 6 | H7C169 | Copper metabolism domain-containing 1 | 2.625 | A0A087WTR4 | Acyl-CoA synthetase medium chain family member 5 | −2.474 |
| 7 | F5GZZ5 | Receptor protein-tyrosine kinase | 2.614 | E7EVL1 | Adenylate cyclase type 8 | −2.477 |
| 8 | A0A7U3JVZ5 | Fibroblast growth factor (FGF) | 2.612 | F8WDK8 | Ribosomal protein L22 like 1 | −2.477 |
| 9 | C9JHJ5 | Golgin A4 | 2.610 | Q96HN2 | Adenosylhomocysteinase 3 (AdoHcyase 3) | −2.477 |
| 10 | A0A087WVA7 | IQ motif containing with AAA domain 1 like | 2.606 | Q5T0Y8 | Sphingomyelin phosphodiesterase acid like 3B | −2.478 |
| 11 | E9PNZ4 | Microtubule actin crosslinking factor 1 | 2.601 | X2CV47 | AKT1m transcript variant 3 | −2.486 |
| 12 | D6R9D2 | Neuronal membrane glycoprotein M6-a | 2.599 | Q8NFB6 | AID | −2.487 |
| 13 | D6REB4 | Poly(A) binding protein interacting protein 1 | 2.595 | D6RB24 | NECAP endocytosis associated 2 | −2.488 |
| 14 | H7C4B8 | Family with sequence similarity 228 member A | 2.594 | Q00722 | Phosphoinositide phospholipase C-beta-2) | −2.489 |
| 15 | A0A499FJI4 | RCR-type E3 ubiquitin transferase | 2.590 | Q9Y573 | Actin-binding protein IPP | −2.492 |
| 16 | Q658V8 | Uncharacterized protein DKFZp666C182 | 2.577 | E7EMD6 | A-kinase anchoring protein 10 | −2.493 |
| 17 | O94763 | Protein phosphatase 1 regulatory subunit 19 | 2.576 | P36896 | Activin receptor type-1B | −2.494 |
| 18 | O75132 | Zinc finger BED domain-containing protein 4 | 2.573 | H0Y3V3 | Adhesion G protein-coupled receptor L2 | −2.494 |
| 19 | B4DUR8 | T-complex protein 1 subunit gamma | 1.200 | A0A7P0MKV3 | Mitochondrial ribosomal protein S22 | −2.494 |
| 20 | A0A1W2PR84 | Adhesion G protein-coupled receptor V1 | −2.494 | |||
| 21 | E5RIU2 | ADP ribosylation factor GTPase activating protein 1 | −2.494 | |||
| 22 | Q9BRH5 | Diacylglycerol O-acyltransferase | −2.495 | |||
| 23 | A0A2R8YG22 | Abhydrolase domain containing 5, lysophosphatidic acid acyltransferase | −2.497 | |||
| 24 | P62701 | Small ribosomal subunit protein eS4, X isoform | −2.498 | |||
| 25 | Q9P212 | 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase epsilon-1 | −2.500 | |||
| 26 | C9JFR9 | Cytochrome P450 family 8 subfamily B member 1 | −2.506 | |||
| 27 | F8WER2 | ADP ribosylation factor like GTPase 5A | −2.508 | |||
| 28 | Q6LBH1 | ACPP (Acid phosphatase) | −2.512 | |||
| 29 | J3KNJ4 | Activating signal cointegrator 1 complex subunit 3 | −2.518 | |||
| 30 | F8VRL1 | Actin related protein 6 | −2.522 | |||
| 31 | Q4G170 | ACACB protein | −2.532 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sirivarasai, J.; Tristitworn, P.; Shantavasinkul, P.C.; Roytrakul, S.; Chansirikarnjana, S.; Ruangritchankul, S.; Chanprasertyothin, S.; Charernwat, P.; Panpunuan, P.; Sura, T.; et al. Genetic Polymorphism of Zinc Transporter-8 Gene (SLC30A8), Serum Zinc Concentrations, and Proteome Profiles Related to Type 2 Diabetes in Elderly. J. Clin. Med. 2025, 14, 790. https://doi.org/10.3390/jcm14030790
Sirivarasai J, Tristitworn P, Shantavasinkul PC, Roytrakul S, Chansirikarnjana S, Ruangritchankul S, Chanprasertyothin S, Charernwat P, Panpunuan P, Sura T, et al. Genetic Polymorphism of Zinc Transporter-8 Gene (SLC30A8), Serum Zinc Concentrations, and Proteome Profiles Related to Type 2 Diabetes in Elderly. Journal of Clinical Medicine. 2025; 14(3):790. https://doi.org/10.3390/jcm14030790
Chicago/Turabian StyleSirivarasai, Jintana, Pimvaree Tristitworn, Prapimporn Chattranukulchai Shantavasinkul, Sittiruk Roytrakul, Sirintorn Chansirikarnjana, Sirasa Ruangritchankul, Suwannee Chanprasertyothin, Piangporn Charernwat, Pachara Panpunuan, Thanyachai Sura, and et al. 2025. "Genetic Polymorphism of Zinc Transporter-8 Gene (SLC30A8), Serum Zinc Concentrations, and Proteome Profiles Related to Type 2 Diabetes in Elderly" Journal of Clinical Medicine 14, no. 3: 790. https://doi.org/10.3390/jcm14030790
APA StyleSirivarasai, J., Tristitworn, P., Shantavasinkul, P. C., Roytrakul, S., Chansirikarnjana, S., Ruangritchankul, S., Chanprasertyothin, S., Charernwat, P., Panpunuan, P., Sura, T., & Sritara, P. (2025). Genetic Polymorphism of Zinc Transporter-8 Gene (SLC30A8), Serum Zinc Concentrations, and Proteome Profiles Related to Type 2 Diabetes in Elderly. Journal of Clinical Medicine, 14(3), 790. https://doi.org/10.3390/jcm14030790