
Postnatal Cytoarchitecture and Neurochemical Hippocampal Dysfunction in Down Syndrome


Abstract
1. Introduction
2. Experimental Section
2.1. Subjects and Tissue Samples
2.2. Immunohistochemistry
2.3. Immunofluorescence
2.4. Histochemistry
2.5. Cell Quantitation
2.6. Statistical Analysis
3. Results
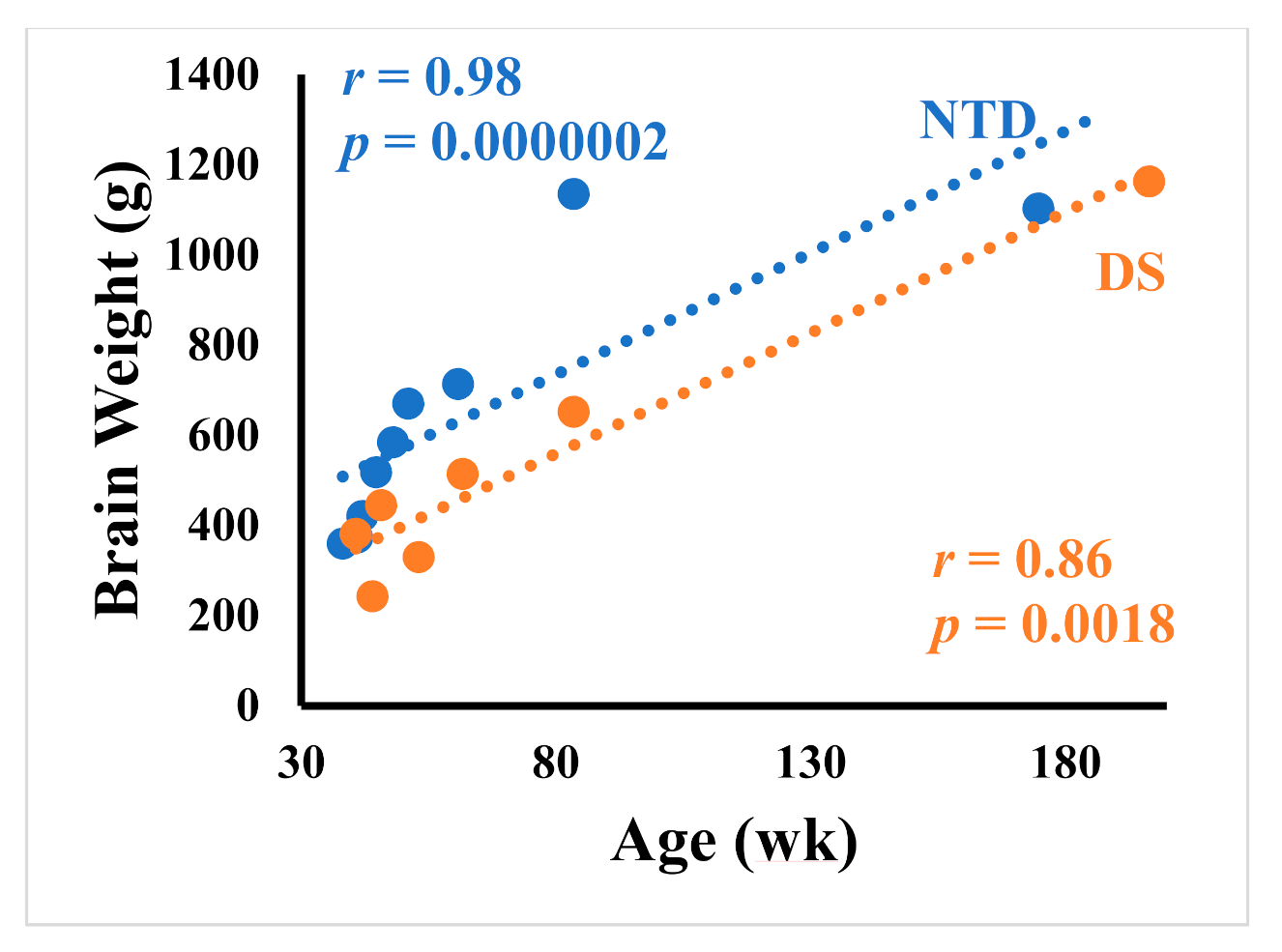
3.1. Case Demographics
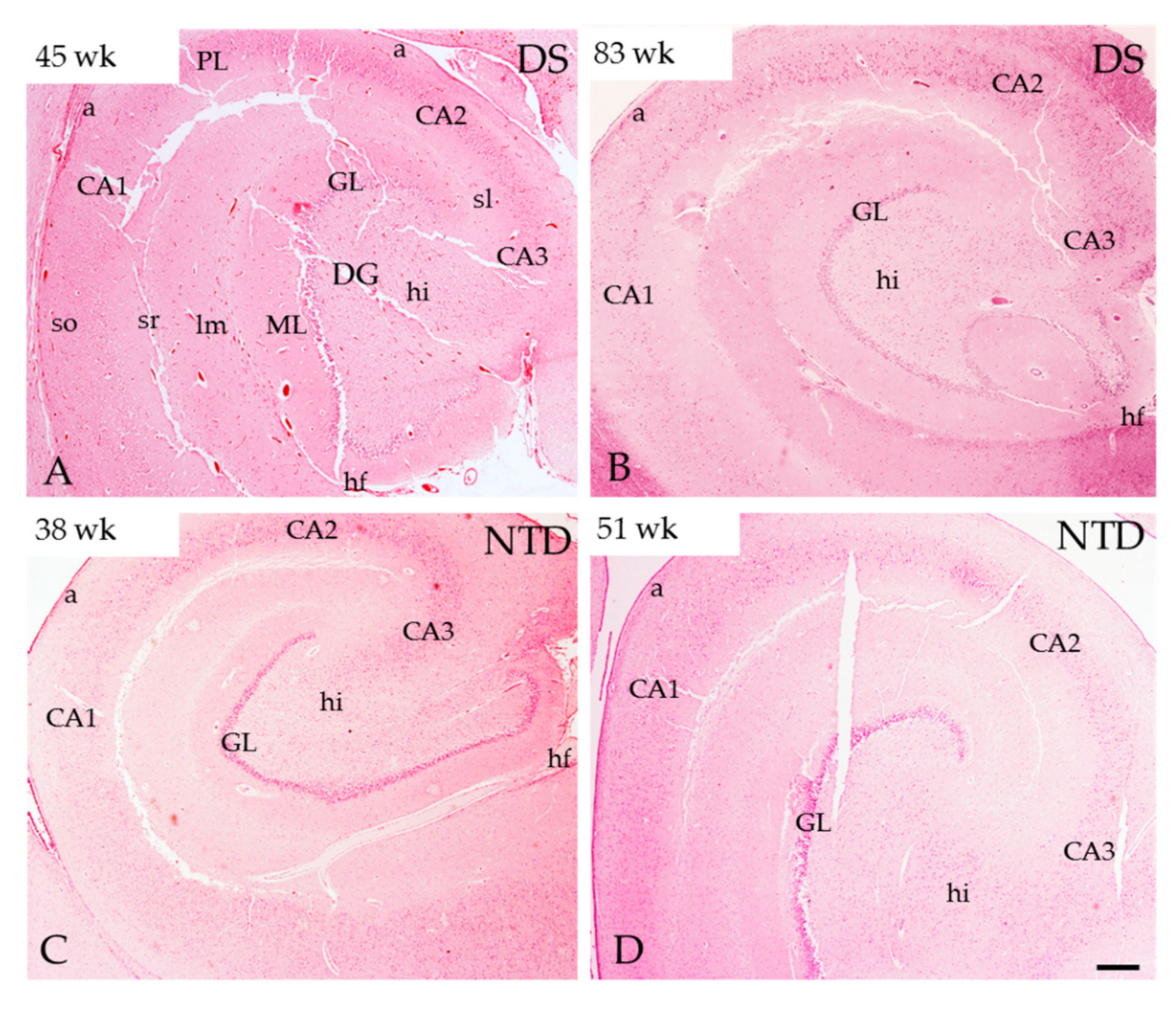
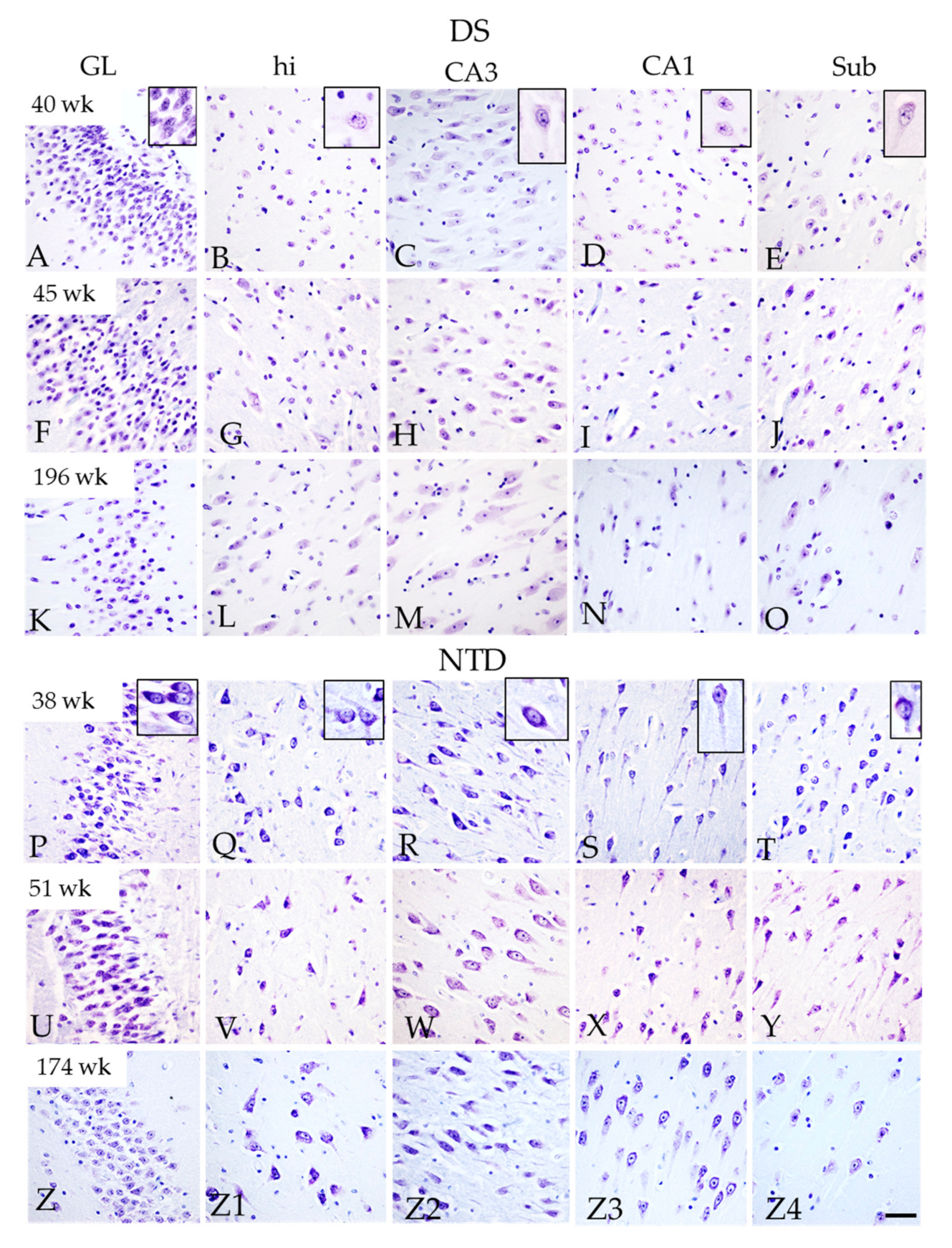
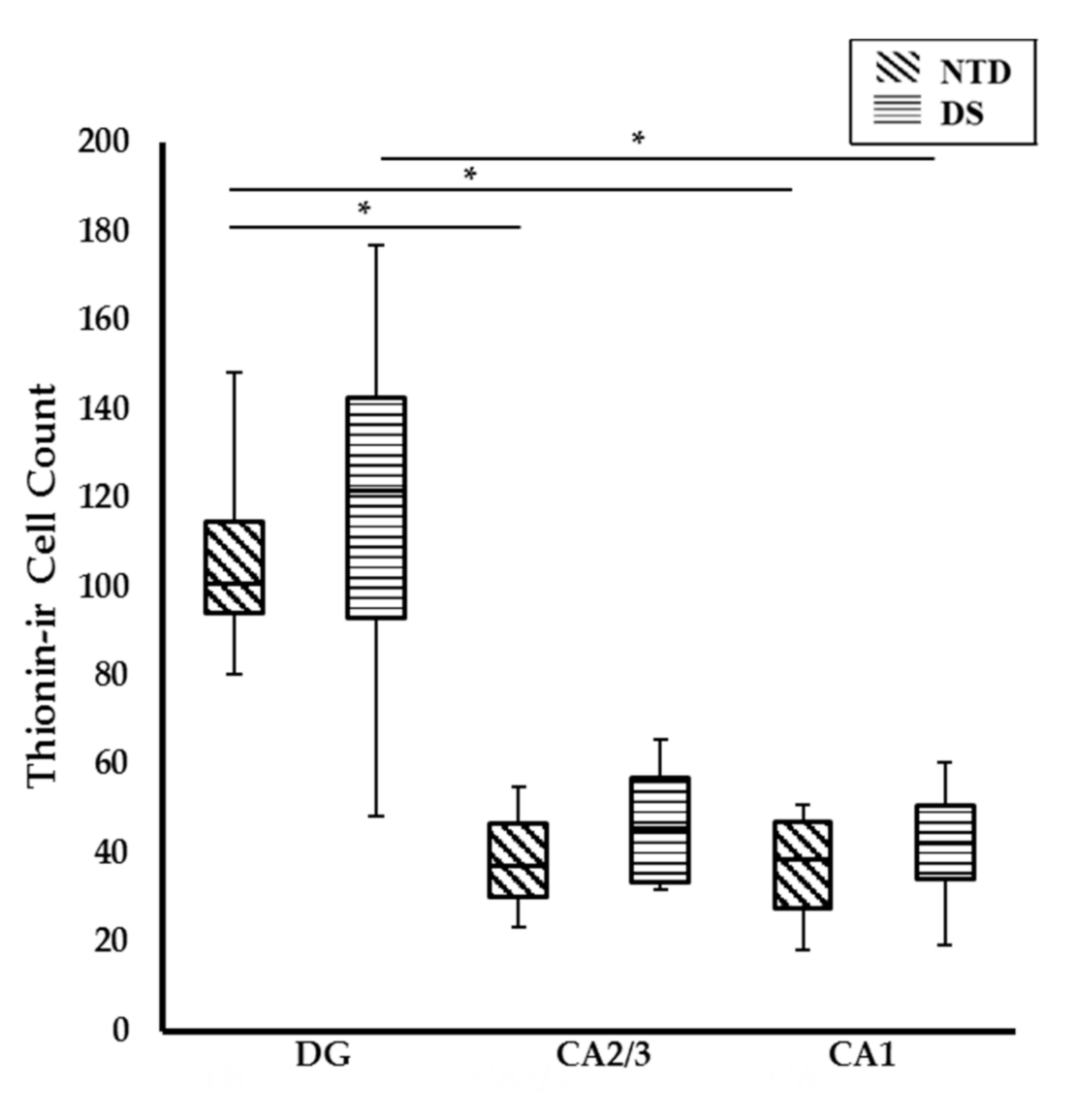
3.2. Postnatal Hippocampal Complex Cytoarchitecture
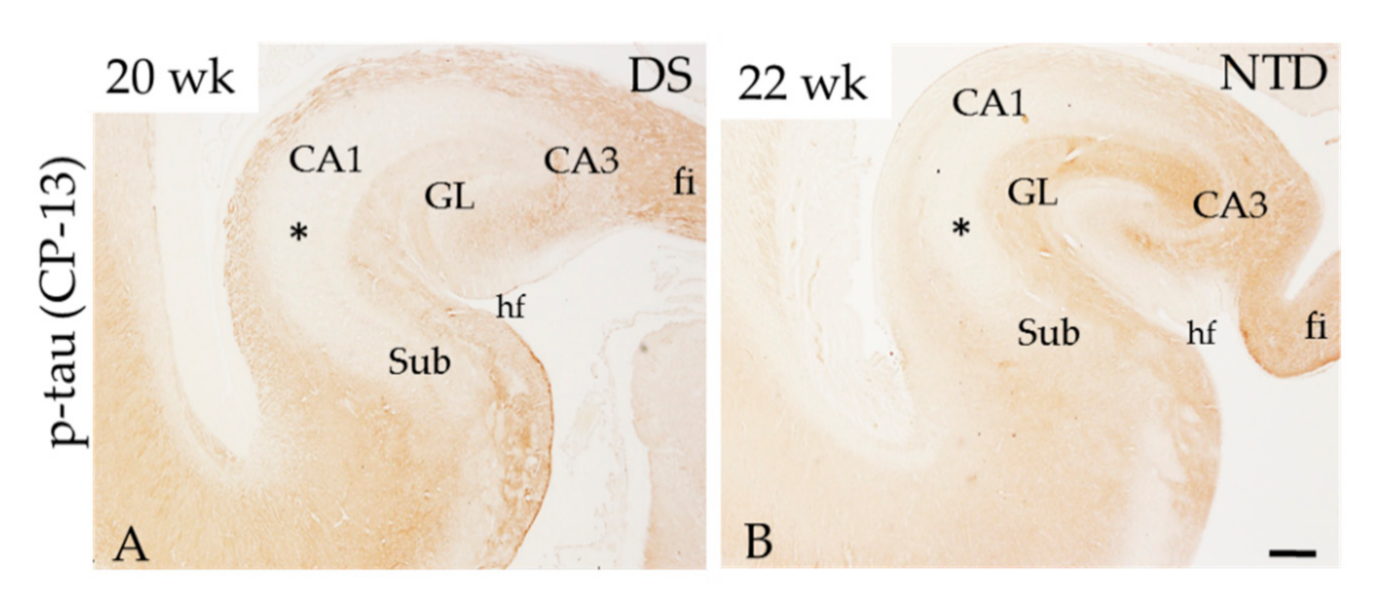
3.3. Hippocampal APP/Aβ, Aβ1–42 and Phosphorylated Tau Immunoreactivity
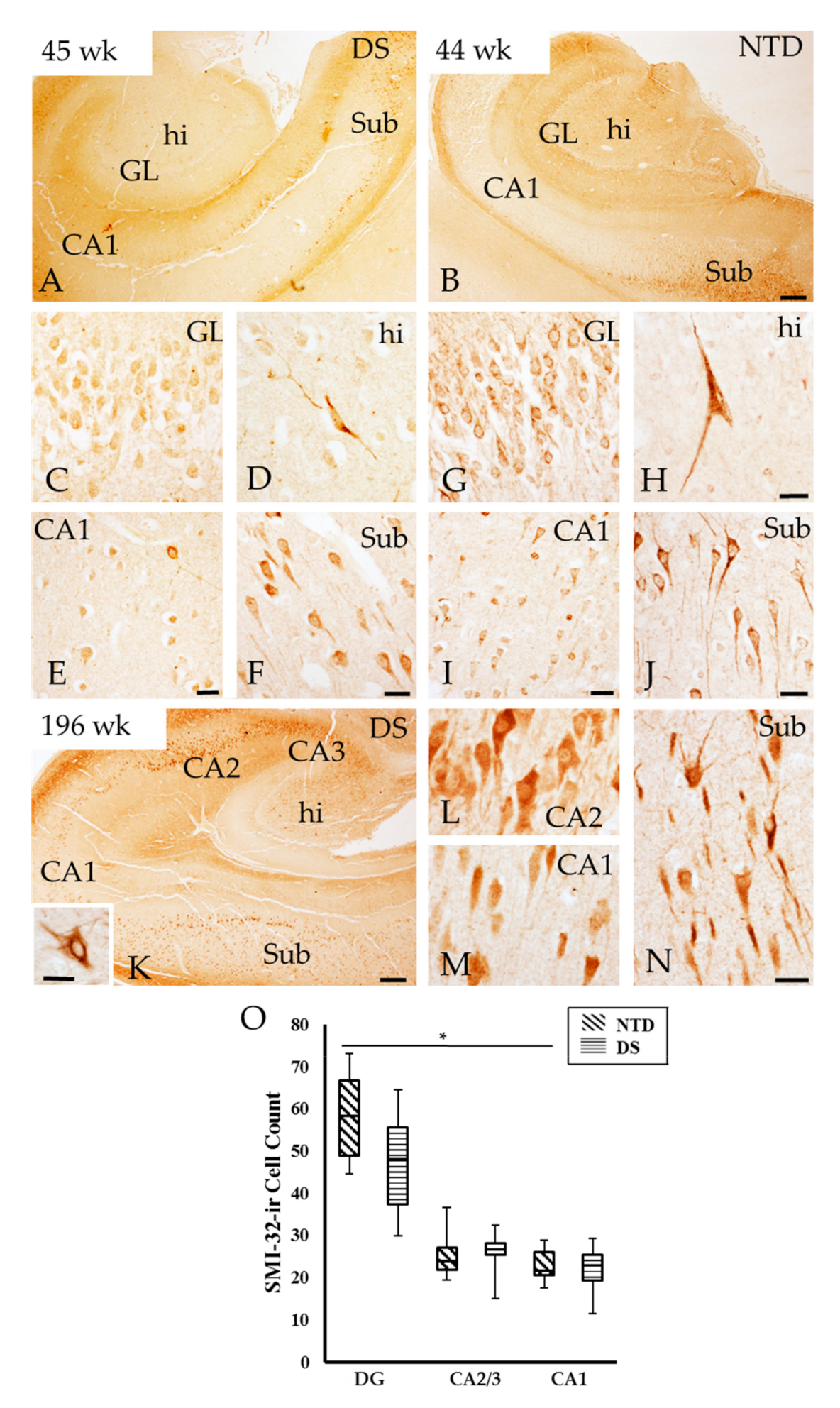
3.4. Neurofilament Positive Profiles in the Postnatal Hippocampal Complex
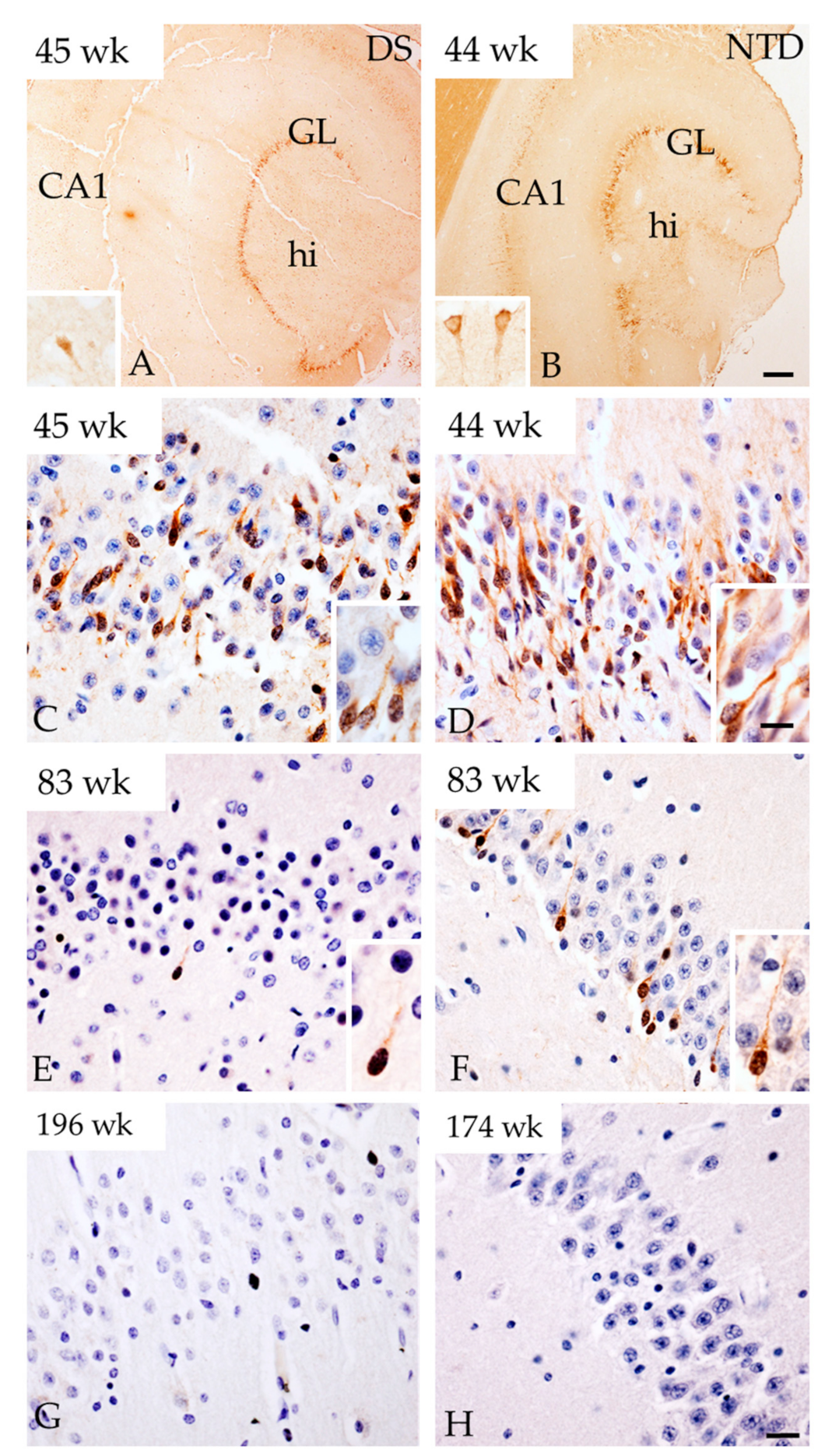
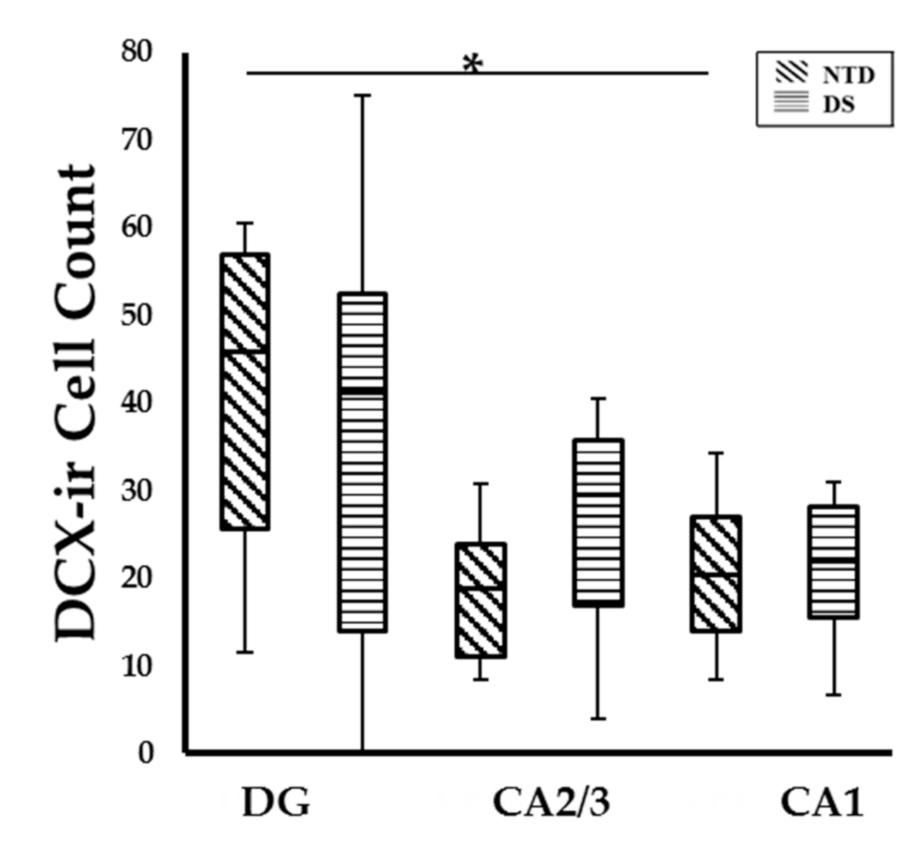
3.5. Postnatal Hippocampal Cellular Proliferation and Neurogenesis
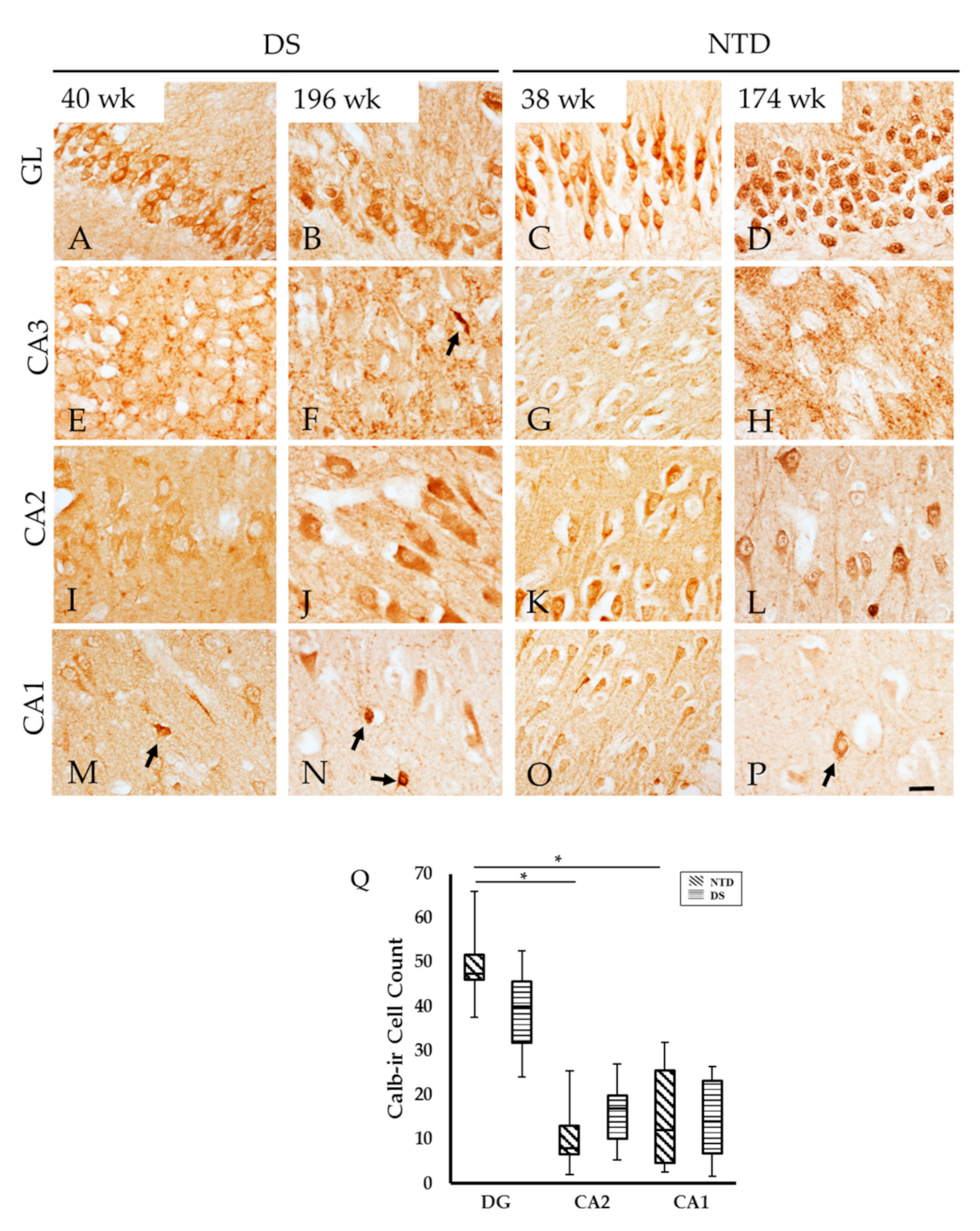
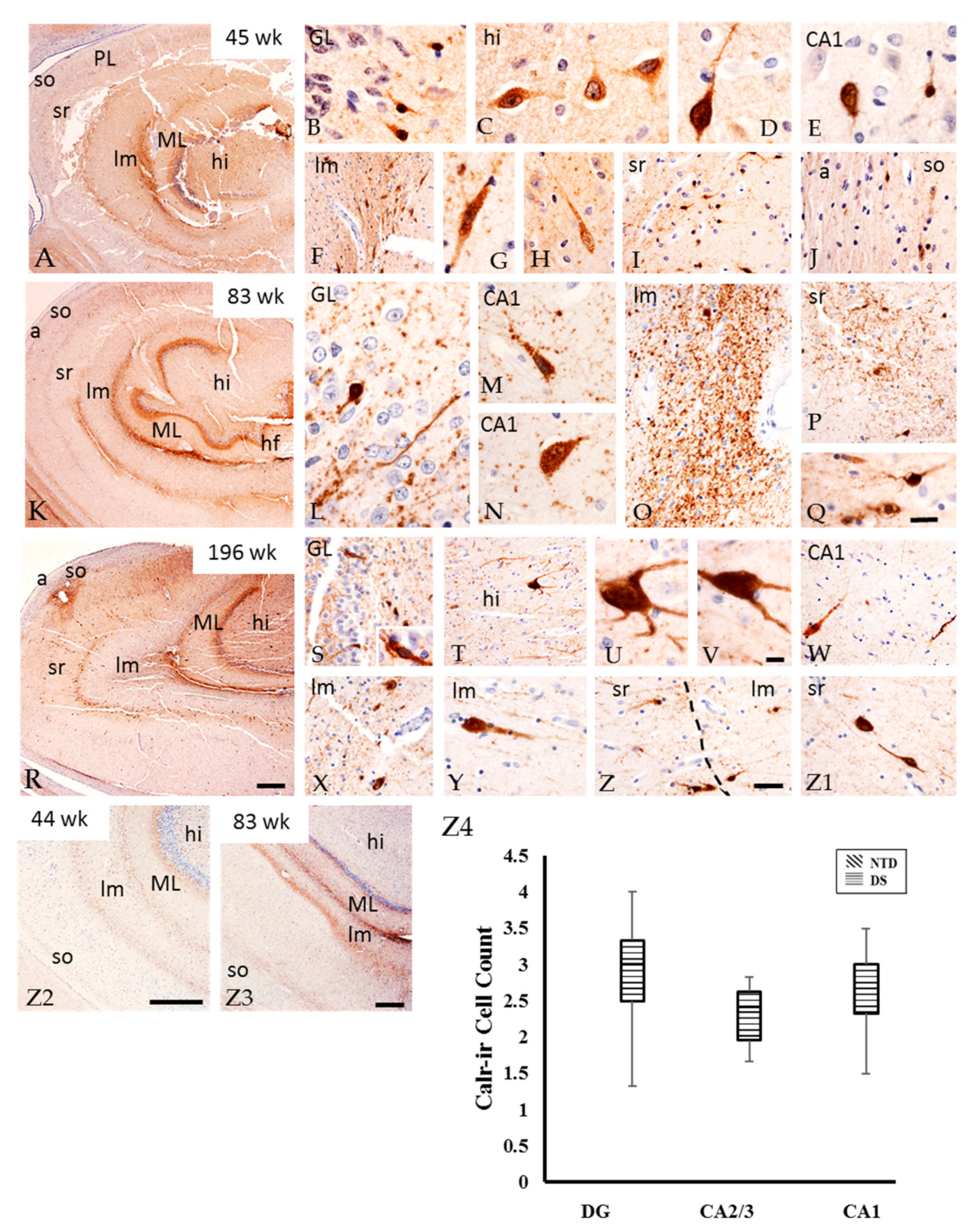
3.6. Postnatal Hippocampal Calcium Binding Protein Immunoreactivity
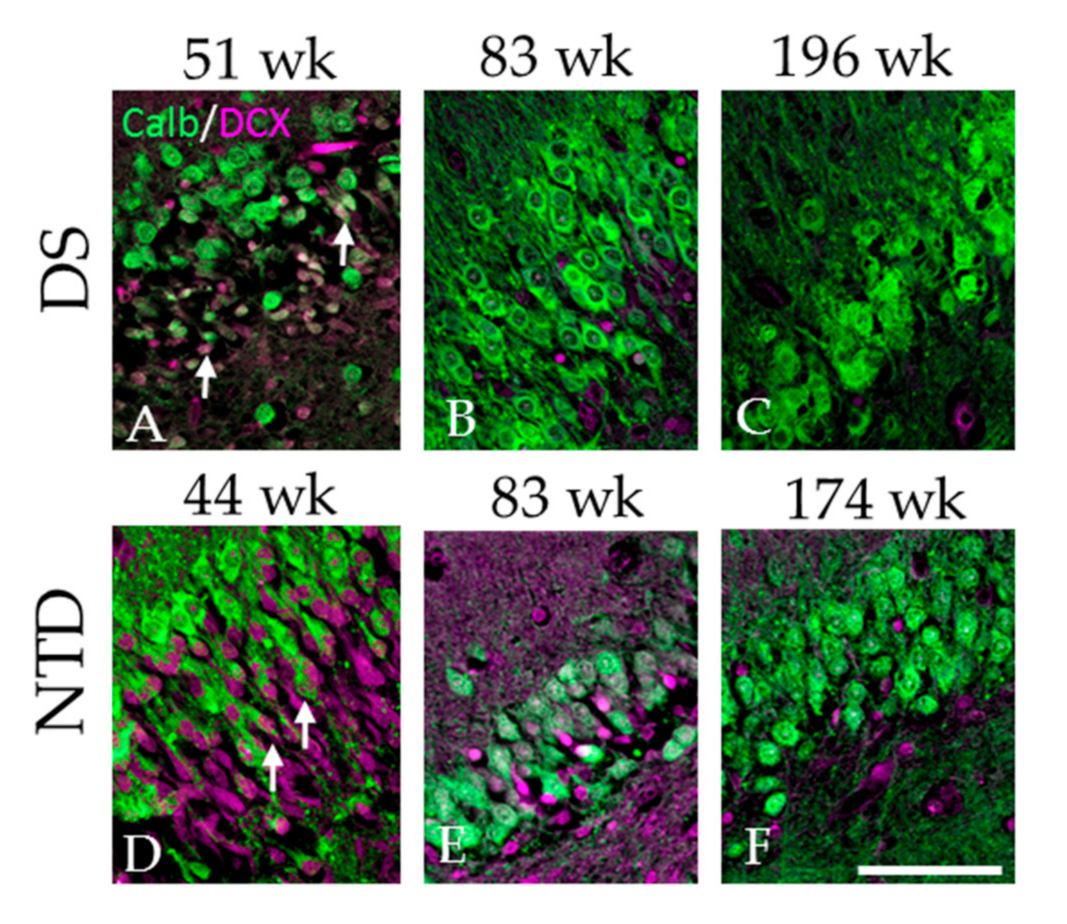
3.7. Colocalization of DCX and Calb in the DG
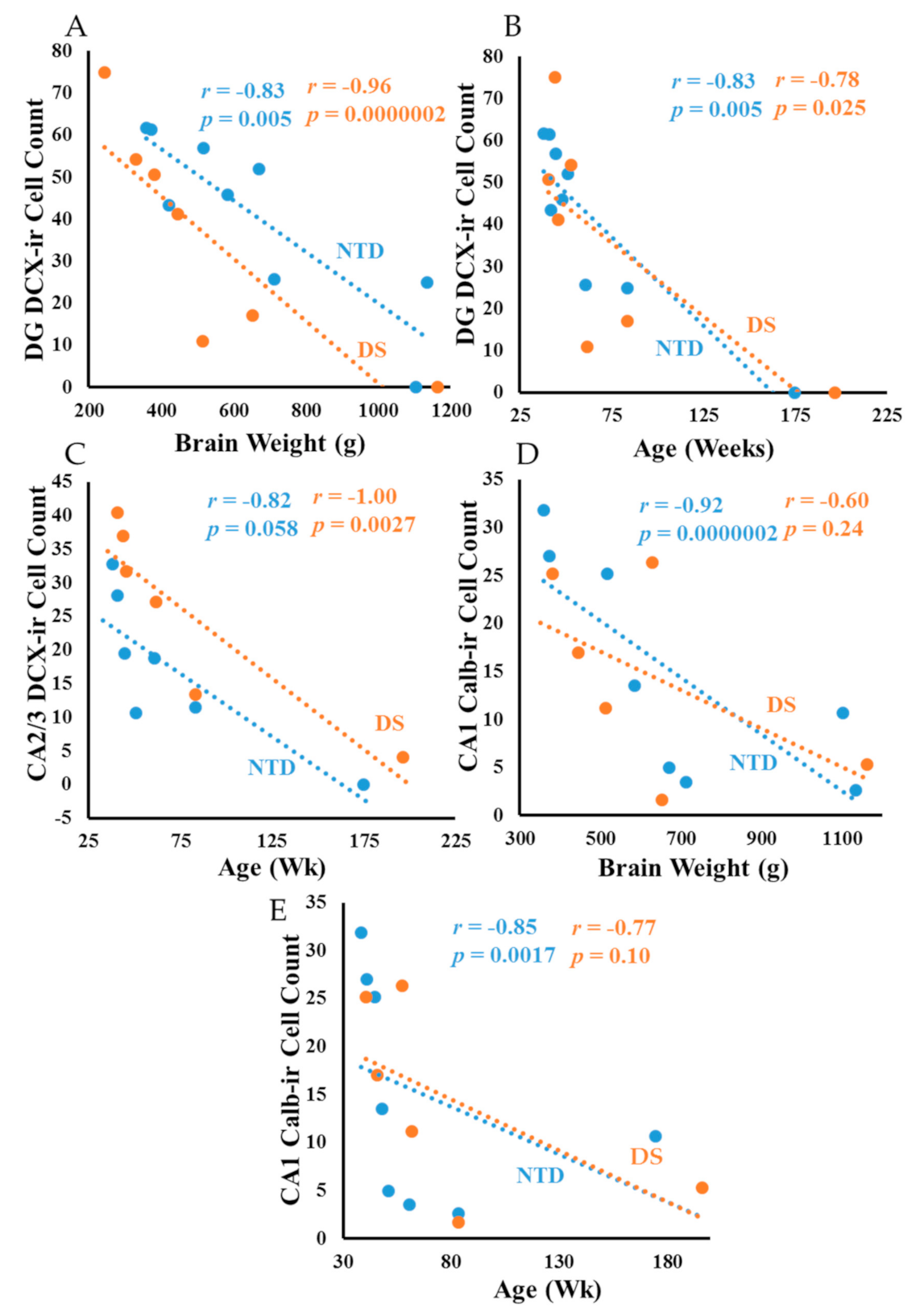
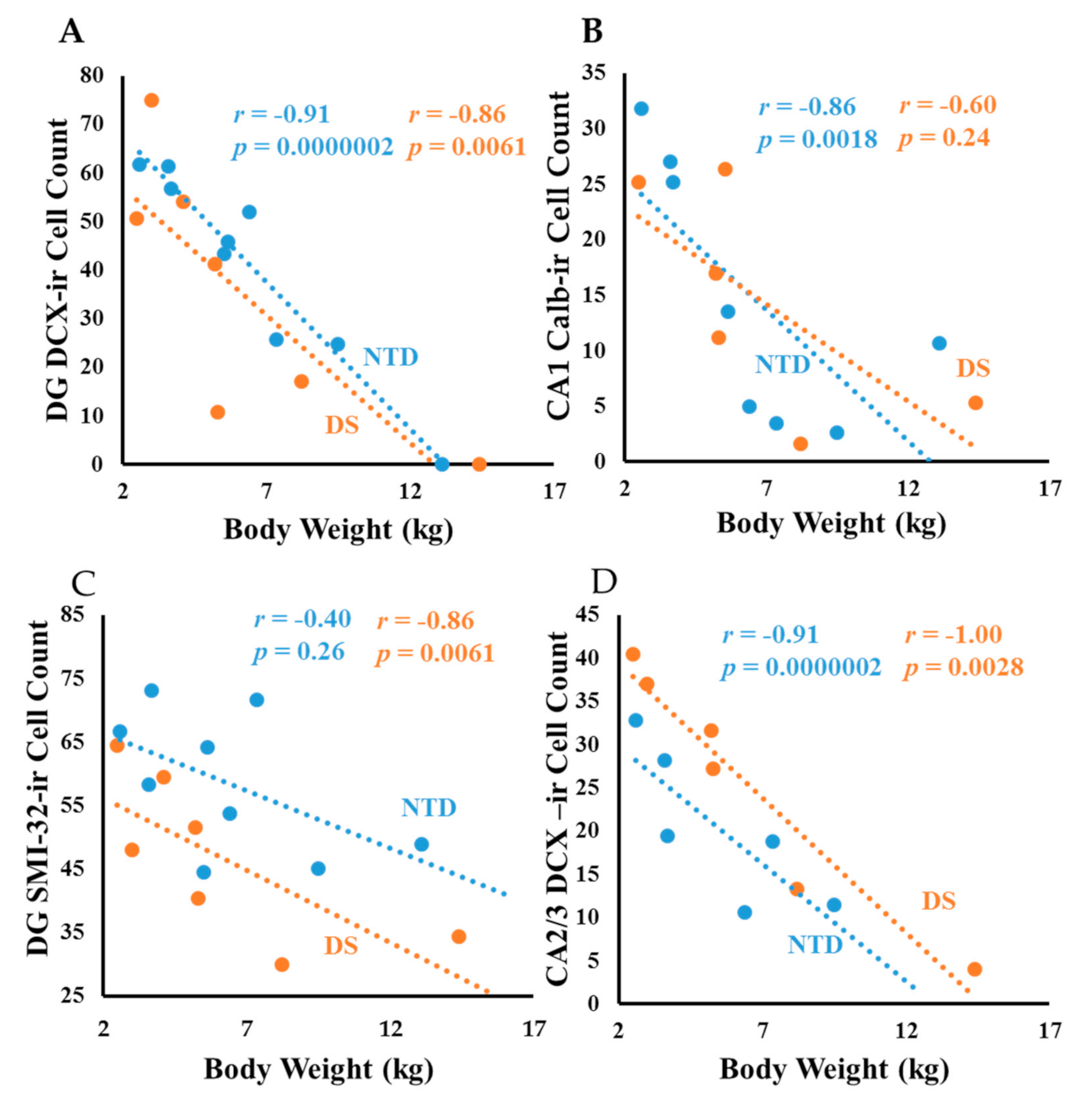
3.8. Correlations
4. Discussion
4.1. Postnatal Hippocampal Neurogenesis in DS
4.2. Postnatal Hippocampal Cytoarchitecture in DS
4.3. Postnatal Hippocampal Pyramidal Cell Maturation in DS
4.4. Postnatal Hippocampal CBP in DS
4.5. Postnatal Hippocampal APP/Aβ1–42 and Tau in DS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bull, M.J. Health supervision for children with Down syndrome. Pediatrics 2011, 128, 393–406. [Google Scholar] [CrossRef]
- Haydar, T.F.; Reeves, R.H. Trisomy 21 and early brain development. Trends Neurosci. 2012, 35, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Coyle, J.T.; Oster-Granite, M.L.; Gearhart, J.D. The neurobiologic consequences of Down syndrome. Brain Res. Bull. 1986, 16, 773–787. [Google Scholar] [CrossRef]
- Lejeune, J.; Gautier, M.; Turpin, R. Mongolism; a chromosomal disease (trisomy). Bull. Acad. Natl. Med. 1959, 143, 256–265. [Google Scholar] [PubMed]
- Mann, D.M.; Yates, P.O.; Marcyniuk, B.; Ravindra, C.R. The topography of plaques and tangles in Down’s syndrome patients of different ages. Neuropathol. Appl. Neurobiol. 1986, 12, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Davidson, Y.S.; Robinson, A.; Prasher, V.P.; Mann, D.M.A. The age of onset and evolution of Braak tangle stage and Thal amyloid pathology of Alzheimer’s disease in individuals with Down syndrome. Acta Neuropathol. Commun. 2018, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.E.; Miguel, J.C.; He, B.; Malek-Ahmadi, M.; Abrahamson, E.E.; Ikonomovic, M.D.; Lott, I.; Doran, E.; Alldred, M.J.; Ginsberg, S.D.; et al. Frontal cortex and striatal cellular and molecular pathobiology in individuals with Down syndrome with and without dementia. Acta Neuropathol. 2019, 137, 413–436. [Google Scholar] [CrossRef] [PubMed]
- Lanfranchi, S.; Jerman, O.; Pont, E.D.; Alberti, A.; Vianello, R. Executive function in adolescents with Down Syndrome. J. Intellect. Disabil. Res. 2010, 54, 308–319. [Google Scholar] [CrossRef]
- Næss, K.A.B.; Lervåg, A.; Lyster, S.A.H.; Hulme, C. Longitudinal relationships between language and verbal short-term memory skills in children with Down syndrome. J. Exp. Child. Psychol. 2015, 135, 43–55. [Google Scholar] [CrossRef]
- Carlesimo, G.A.; Marotta, L.; Vicari, S. Long-term memory in mental retardation: Evidence for a specific impairment in subjects with Down’s syndrome. Neuropsychologia 1997, 35, 71–79. [Google Scholar] [CrossRef]
- Dusek, J.A.; Eichenbaum, H. The hippocampus and memory for orderly stimulus relations. Proc. Natl. Acad. Sci. USA 1997, 94, 7109–7114. [Google Scholar] [CrossRef] [PubMed]
- Fortin, N.J.; Agster, K.L.; Eichenbaum, H.B. Critical role of the hippocampus in memory for sequences of events. Nat. Neurosci. 2002, 5, 458–462. [Google Scholar] [CrossRef]
- Bird, C.M.; Burgess, N. The hippocampus and memory: Insights from spatial processing. Nat. Rev. Neurosci. 2008, 9, 182–194. [Google Scholar] [CrossRef]
- Humphrey, T. The development of the human hippocampal fissure. J. Anat. 1967, 101, 655–676. [Google Scholar]
- Insausti, R.; Amaral, D.G. Hippocampal formation. In Human Nervous System, 2nd ed.; Paxinos, G., Mai, J.K., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 871–914. [Google Scholar]
- Pinter, J.D.; Brown, W.E.; Eliez, S.; Schmitt, J.E.; Capone, G.T.; Reiss, A.L. Amygdala and hippocampal volumes in children with Down syndrome: A high-resolution MRI study. Neurology 2001, 56, 972–974. [Google Scholar] [CrossRef]
- Śmigielska-Kuzia, J.; Boćkowski, L.; Sobaniec, W.; Sendrowski, K.; Olchowik, B.; Cholewa, M.; Lukasiewicz, A.; Łebkowska, U. A volumetric magnetic resonance imaging study of brain structures in children with Down syndrome. Neurol. Neurochir. Pol. 2011, 45, 363–369. [Google Scholar] [CrossRef]
- Carducci, F.; Onorati, P.; Condoluci, C.; Gennaro, G.D.; Quarato, P.P.; Pierallini, A.; Sara, M.; Miano, S.; Cornia, R.; Albertini, G. Whole-brain voxel-based morphometry study of children and adolescents with Down syndrome. Funct. Neurol. 2013, 28, 19–28. [Google Scholar]
- Stagni, F.; Giacomini, A.; Emili, M.; Guidi, S.; Bartesaghi, R. Neurogenesis impairment: An early developmental defect in Down syndrome. Free Radic. Biol. Med. 2018, 114, 15–32. [Google Scholar] [CrossRef]
- Teller, J.K.; Russo, C.; DeBusk, L.M.; Angelini, G.; Zaccheo, D.; Dagna-Bricarelli, F.; Scartezzini, P.; Bertolini, S.; Mann, D.M.; Tabaton, M.; et al. Presence of soluble amyloid beta-peptide precedes amyloid plaque formation in Down’s syndrome. Nat. Med. 1996, 2, 93–95. [Google Scholar] [CrossRef]
- Giacomini, A.; Stagni, F.; Trazzi, S.; Guidi, S.; Emili, M.; Brigham, E.; Ciani, E.; Bartesaghi, R. Inhibition of APP gamma-secretase restores Sonic Hedgehog signaling and neurogenesis in the TS65Dn mouse model of Down syndrome. Neurobiol. Dis. 2015, 82, 385–396. [Google Scholar] [CrossRef]
- Freude, K.K.; Penjwini, M.; Davis, J.L.; LaFerla, F.M.; Blurton-Jones, M. Soluble amyloid precursor protein induces rapid neural differentiation of human embryonic stem cells. J. Biol. Chem. 2011, 286, 24264–24274. [Google Scholar] [CrossRef]
- Nicolas, M.; Hassan, B.A. Amyloid precursor protein and neural development. Development 2014, 141, 2543–2548. [Google Scholar] [CrossRef]
- Milenkovic, I.; Stojanovic, T.; Aronica, E.; Fülöp, L.; Bozsó, Z.; Máté, Z.; Yanagawa, Y.; Adle-Biassette, H.; Lubec, G.; Szabo, G.; et al. GABAA receptor subunit deregulation in the hippocampus of human foetuses with Down syndrome. Brain Struct. Funct. 2018, 223, 1501–1518. [Google Scholar] [CrossRef]
- Simic, G.; Diana, A.; Hof, P.R. Phosphorylation pattern of tau associated with distinct changes of the growth cone cytoskeleton. Prog. Mol. Subcell. Biol. 2003, 32, 33–48. [Google Scholar] [PubMed]
- Milenkovic, I.; Jarc, J.; Dassler, E.; Aronica, E.; Lyer, A.; Adle-Biassette, H.; Scharrer, A.; Reischer, T.; Hainfellner, J.A.; Kovacs, G.G. The physiological phosphorylation of tau is critically changed in fetal brains of individuals with Down syndrome. Neurophathol. Appl. Neurobiol. 2018, 44, 314–327. [Google Scholar] [CrossRef]
- Ábrahám, H.; Vincze, A.; Veszprémi, B.; Kravják, A.; Gömöri, É.; Kovács, G.G.; Seress, L. Impaired myelination of the human hippocampal formation in Down syndrome. Int. J. Dev. Neurosci. 2012, 30, 147–158. [Google Scholar] [CrossRef] [PubMed]
- US Department of Health and Human Services National Institutes of Health. Available online: https://www.nichd.nih.gov/ncmhep/initiatives/know-your-terms/moms (accessed on 4 June 2020).
- Mufson, E.J.; Perez, S.E.; Nadeem, M.; Mahady, L.; Kanaan, N.M.; Abrahamson, E.E.; Ikonomovic, M.D.; Crawford, F.; Alvarez, V.; Stein, T.; et al. Progression of tau pathology within cholinergic nucleus basalis neurons in chronic traumatic encephalopathy: A chronic effects of neurotrauma consortium study. Brain Inj. 2016, 30, 1399–1413. [Google Scholar] [CrossRef]
- Insausti, R.; Cebada-Sancehez, S.; Marcos, P. Postnatal development of the human hippocampal formation. Adv. Anat. Embryol. Cell Biol. 2010, 206, 1–86. [Google Scholar] [PubMed]
- Nkomozepi, P.; Mazengenya, P.; Ihunwo, A.O. Age-related changes in Ki-67 and DCX expression in the BALB/ c mouse (Mus Musculus) brain. Int. J. Dev. Neurosci. 2019, 72, 36–47. [Google Scholar] [CrossRef]
- Fasemore, T.M.; Patzke, N.; Kaswera-Kyamakya, C.; Gilissen, E.; Manger, P.R.; Ihunwo, A.O. The distribution of Ki-67 and doublecortin-immunopositive cells in the brains of three Strepsirrhine primates: Galago demidoff, Perodicticus potto, and Lemur catta. Neuroscience 2018, 372, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]
- Guidi, S.; Bonasoni, P.; Ceccarelli, C.; Santini, D.; Gualtieri, F.; Ciani, E.; Bartesaghi, R. Neurogenesis impairment and increased cell death reduce total neuron number in the hippocampal region of fetuses with Down syndrome. Brain Pathol. 2008, 18, 180–197. [Google Scholar] [CrossRef]
- Sorrells, S.; Paredes, M.; Cebrian-Silla, A.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Scudamore, C.L.; Hodgson, H.K.; Patterson, L.; Macdonald, A.; Brown, F.; Smith, K.C. The effect of post-mortem delay on immunohistochemical labelling-a short review. Comp. Clin. Pathol. 2011, 20, 95–101. [Google Scholar] [CrossRef]
- Gleeson, J.G.; Lin, P.T.; Flanagan, L.A.; Walsh, C.A. Doublecortin is a microtubule-associated protein and is expressed widely by migrating neurons. Neuron 1999, 23, 257–271. [Google Scholar] [CrossRef]
- Mizuguchi, M.; Qin, J.; Yamada, M.; Ikeda, K.; Takashima, S. High expression of doublecortin and KIAA0369 protein in fetal brain suggests their specific role in neuronal migration. Am. J. Pathol. 1999, 155, 1713–1721. [Google Scholar] [CrossRef]
- Ohshima, T. Neuronal migration and protein kinases. Front. Neurosci. 2015, 8, 458. [Google Scholar] [CrossRef]
- Corbo, J.C.; Deuel, T.A.; Long, J.M.; Laporte, P.; Tsai, E.; Wynshaw-Boris, A.; Walsh, C.A. Doublecortin is required in mice for lamination of the hippocampus but not the neocortex. J. Neurosci. 2002, 22, 7548–7557. [Google Scholar] [CrossRef] [PubMed]
- Kadar, A.; Wittmann, G.; Liposits, Z.; Fekete, C. Improved method for combination of immunocytochemistry and nissl staining. J. Neurosci. Methods 2009, 184, 115–118. [Google Scholar] [CrossRef]
- Kato, H.; Nishitoh, H. Stress responses from the endoplasmic reticulum in cancer. Front. Oncol. 2015, 5, 93. [Google Scholar] [CrossRef]
- Yuan, A.; Rao, M.V.; Veeranna; Nixon, R.A. Neurofilaments and neurofilament proteins in health and disease. Cold Spring Harb. Perspect. Biol. 2017, 9, a018309. [Google Scholar] [CrossRef]
- Ouda, L.; Druga, R.; Syka, J. Distribution of SMI-32-immunoreactive neurons in the central auditory system of the rat. Brain Struct. Funct. 2011, 217, 19–36. [Google Scholar] [CrossRef]
- Li, Y.; Jung, P.; Brown, A. Axonal transport of neurofilaments: A single population of intermittently moving polymers. J. Neurosci. 2012, 32, 746–758. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lavenex, P.; Lavenex, P.B.; Bennett, J.L.; Amaral, D.G. Postmortem changes in the neuroanatomical characteristics of the primate brain: Hippocampal formation. J. Comp. Neurol. 2009, 512, 27–51. [Google Scholar] [CrossRef]
- Torack, R.M.; Miller, J.W. Denervation induced abnormal phosphorylation in hippocampal neurons. Brain Res. 1995, 669, 135–139. [Google Scholar] [CrossRef]
- Hamlet, E.D.; Boger, H.A.; Ledreux, A.; Kelley, C.M.; Mufson, E.J.; Falangola, M.F.; Guilfoyle, D.N.; Nixon, R.A.; Patterson, D.; Duval, N.; et al. Cognitive impairment, neuroimaging, and Alzheimer neuropathology in mouse models of Down syndrome. Curr. Alzheimer Res. 2016, 13, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Seress, L.; Mrzljak, L. Postnatal development of mossy cells in the human dentate gyrus: A light microscopic golgi study. Hippocampus 1993, 2, 127–141. [Google Scholar] [CrossRef]
- Fairless, R.; Williams, S.K.; Diem, R. Calcium-binding proteins as determinants of central nervous system neuronal vulnerability to disease. Int. J. Mol. Sci. 2019, 20, 2146. [Google Scholar] [CrossRef]
- Yáñez, M.; Gil-Longo, J.; Campos-Toimil, M. Calcium Binding Proteins. Adv. Exp. Med. Biol. 2012, 740, 461–482. [Google Scholar]
- Wu, D.; Bacaj, T.; Morishita, W.; Goswami, D.; Arendt, K.L.; Xu, W.; Chen, L.; Malenka, R.C.; Südhof, T.C. Postsynaptic synaptotagmins mediate AMPA receptor exocytosis during LTP. Nature 2017, 544, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Rusakov, D.A. Ca2+-dependent mechanisms of presynaptic control at central synapses. Neuroscientist 2006, 12, 317–326. [Google Scholar] [CrossRef]
- Caillard, O.; Moreno, H.; Schwaller, B.; Llano, I.; Celio, M.R.; Marty, A. Role of the calcium-binding protein parvalbumin in short-term synaptic plasticity. Proc. Natl. Acad. Sci. USA 2000, 97, 13372–13377. [Google Scholar] [CrossRef]
- De Lecea, L.; Del Rio, J.A.; Soriano, E. Developmental expression of parvalbumin mRNA in the cerebral cortex and hippocampus of the rat. Brain Res. Mol. Brain Res. 1995, 32, 1–13. [Google Scholar] [CrossRef]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Deng, X.; Gu, L.; Sui, N.; Guo, J.; Liang, J. Parvalbumin interneuron in the ventral hippocampus functions as a discriminator in social memory. Proc. Natl. Acad. Sci. USA 2019, 116, 16583–16592. [Google Scholar] [CrossRef]
- Pavlova, M.A.; Galli, J.; Pagani, F.; Micheletti, S.; Guerreschi, M.; Sokolov, A.N.; Fallgatter, A.J.; Fazzi, E.M. Social cognition in Down syndrome: Face tuning in face-like non-face images. Front. Psychol. 2018, 9, 2583. [Google Scholar] [CrossRef]
- Camp, A.J.; Wijesinghe, R. Calretinin: Modulator of neuronal excitability. Int. J. Biochem. Cell Biol. 2009, 41, 2118–2121. [Google Scholar] [CrossRef] [PubMed]
- Guidi, S.; Giacomini, A.; Stagni, F.; Emili, M.; Uguagliati, B.; Bonasoni, M.P.; Bartesaghi, R. Abnormal development of the inferior temporal region in fetuses with Down syndrome. Brain Pathol. 2018, 28, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Zorilla de San Martin, J.; Trigo, F.F.; Kawaguchi, S.Y. Axonal GABAA receptors depolarize presynaptic terminals and facilitate transmitter release in cerebellar purkinje cells. J. Physiol. 2017, 595, 7477–7493. [Google Scholar] [CrossRef]
- Contestabile, A.; Magara, S.; Cancedda, L. The GABAergic hypothesis for cognition disabilities in Down syndrome. Front. Cell Neurosci. 2017, 11, 54. [Google Scholar] [CrossRef] [PubMed]
- Wong, O.G.; Cheung, C.L.; Ip, P.P.; Ngan, H.Y.; Cheung, A.N. Amyloid precursor protein overexpression in Down syndrome trophoblast reduces cell invasiveness and interferes with syncytialization. Am. J. Pathol. 2018, 188, 2307–2317. [Google Scholar] [CrossRef]
- Gyure, K.A.; Durham, R.; Stewart, W.F.; Smialek, J.E.; Troncoso, J.C. Intraneuronal abeta-amyloid precedes development of amyloid plaques in Down syndrome. Arch. Pathol. Lab. Med. 2001, 125, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Nalivaeva, N.N.; Turner, A.J. The amyloid precursor protein: A biochemical enigma in brain development, function and disease. FEBS Lett. 2013, 587, 2046–2054. [Google Scholar] [CrossRef]
- Korenberg, J.R.; Pulst, S.M.; Neve, R.L.; West, R. The Alzheimer amyloid precursor protein maps to human chromosome 21 bands q21.105–q21.05. Genomics 1989, 5, 124–127. [Google Scholar] [CrossRef]
- Wang, H.Y.; Stucky, A.; Liu, J.; Shen, C.; Trocme-Thibierge, C.; Morain, P. Dissociating beta-amyloid from alpha 7 nicotinic acetylcholine receptor by a novel therapeutic agent, S 24795, normalizes alpha 7 nicotinic acetylcholine and NMDA receptor function in Alzheimer’s disease brain. J. Neurosci. 2009, 29, 10961–10973. [Google Scholar] [CrossRef] [PubMed]
- Manczak, M.; Reddy, P.H. Abnormal interaction between the mitochondrial fission protein Drp1 and hyperphosphorylated tau in Alzheimers disease neurons: Implications for mitochondrial dysfunction and neuronal damage. Hum. Mol. Genet. 2012, 21, 2538–2547. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Pathways towards and away from Alzheimers disease. Nature 2004, 430, 631–639. [Google Scholar] [CrossRef]
- Mufson, E.J.; Benzing, W.C.; Kordower, J.H. Dissociation of galaninergic and neurotrophic plasticity in Down syndrome and Alzheimer disease. Prog. Clin. Biol. Res. 1995, 393, 105–122. [Google Scholar] [PubMed]
- Andreadis, A.; Brown, W.M.; Kosik, K.S. Structure and novel exons of the human tau gene. Biochemistry 1992, 31, 10626–10633. [Google Scholar] [CrossRef]
- Jovanov-Milošević, N.; Petrović, D.; Sedmak, G.; Vukšić, M.; Hof, P.R.; Simić, G. Human fetal tau protein isoform: Possibilities for Alzheimer’s disease treatment. Int. J. Biochem. Cell Biol. 2012, 44, 1290–1294. [Google Scholar] [CrossRef] [PubMed]
- Weingarten, M.D.; Lockwood, A.H.; Hwo, S.Y.; Kirschner, M.W. A protein factor essential for microtubule assembly. Proc. Natl. Acad. Sci. USA 1975, 72, 1858–1862. [Google Scholar] [CrossRef]
- Lemoine, L.; Ledreux, A.; Mufson, E.J.; Perez, S.E.; Simic, G.; Doran, E.; Lott, I.; Carroll, S.; Bharani, K.; Thomas, S.; et al. Regional binding of tau and amyloid PET tracers in Down syndrome autopsy brain tissue. Mol. Neurodegener. 2020, 15, 68. [Google Scholar] [CrossRef] [PubMed]
- Coppelletta, J.M.; Wolbach, S.B. Body length and organ weights of infants and children. Am. J. Pathol. 1933, 9, 55. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C.D. | Sex | Age at Birth (wk) | Age at Death (wk) | Post-natal Life (wk) | Brain Weight (g) | Body Weight (kg) | Height (cm) | PMI (h) | Tissue Source | Cause of Death/Comorbidity | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | DS | M | 31.7 | 40.7 | 9 | 381.0 | 2.5 | 49 | 58 | LCH | congenital heart disease |
| 2 | DS | F | 40.0 | 44.0 | 4 | 243.0 | 3 | 49.3 | 20 | LCH | congenital heart disease |
| 3 | DS | F | 37.0 | 45.7 | 8.5 | 445.0 | 5.2 | 55 | 16 | LCH | congenital heart disease |
| 4 | DS | M | 38.0 | 53.0 | 15 | 330.0 | 4.1 | 49 | 21 | LCH | lung disease |
| 5 | DS | M | 40.0 | 57.4 | 17.2 | 630.0 | 5.6 | 48 | 14 | PCH | congenital heart disease |
| 6 | DS | F | 40.0 | 61.7 | 21.7 | 514.0 | 5.3 | 60 | 34 | LCH | acute pneumonia |
| 7 | DS | M | 40.0 | 83.5 | 23.5 | 652.0 | 8.2 | 84.3 | 10 | PCH | congenital heart disease |
| 8 | DS | M | 40.0 | 196.4 | 156.4 | 1163.0 | 14.4 | 97 | 22 | LCH | lung damage |
| 9 | NTD | F | 38.1 | 38.1 | 0 | 360.0 | 2.6 | 48.5 | -- | PCH | pulmonary hemorrhage |
| 10 | NTD | F | 37.0 | 41.0 | 4 | 373.3 | 3.6 | 55.5 | 6 | PCH | congenital heart disease |
| 11 | NTD | F | 39.0 | 42.0 | 3 | 420.9 | 5.52 | 39 | 16 | PCH | congenital heart disease |
| 12 | NTD | M | 40.0 | 44.7 | 4.7 | 518.0 | 3.7 | 54.5 | 26 | PCH | congenital heart disease |
| 13 | NTD | M | 39.0 | 48.0 | 54 | 584.2 | 5.7 | 55.8 | 26 | PCH | congenital heart disease |
| 14 | NTD | M | 40.0 | 51.0 | 11 | 670.0 | 6.4 | 63.5 | 17 | PCH | septicemia |
| 15 | NTD | F | 27.0 | 60.8 | 33.7 | 713.5 | 7.4 | 70.2 | -- | PCH | systemic inflammatory response syndrome |
| 16 | NTD | M | 40.0 | 83.5 | 23.54 | 1134.9 | 9.5 | 77.8 | 29 | PCH | acute pneumonia |
| 17 | NTD | M | 40.0 | 174.7 | 135 | 1103.0 | 13.1 | 90.1 | 18 | PCH | lymphoma |
| Description | Dilution | Company | |
|---|---|---|---|
| SMI-32 | Mouse monoclonal purified anti-neurofilament H, non-phosphorylated antibody | 1:500 | Biolegend, San Diego, CA, USA |
| SMI-34 | Mouse monoclonal purified anti-neurofilament H, phosphorylated antibody | 1:200 | Biolegend, San Diego, CA, USA |
| DCX | Neuronal migration protein Doublecortin (E-6) | 1:250 | Santa Cruz Biotechnology, Dallas, TX, USA |
| Ki-67 | Monoclonal mouse anti-human Ki-67 antigen clone MIB-1 | 1:200 | Dako, Denmark A/S, Glostrup, Denmark |
| Calb | Rabbit polyclonal anti-calbindin D-28K | 1:500 | Swant, Marly, Switzerland |
| Parv | Mouse monoclonal anti-parvalbumin | 1:500 | Millipore, Billerica, CA, USA |
| Calr | Rabbit anti-calretinin | 1:500 | Millipore, Billerica, CA, USA |
| APP/Aβ (6E10) | Mouse monoclonal against human β-Amyloid (Aβ, 1–16 aa) and APP | 1:300 | BioLegend, San Diego, CA, USA |
| Aβ1–42 | Rabbit polyclonal anti-Aβ1–42 | 1:100 | Millipore, Billerica, CA, USA |
| CP-13 | Phospho-tau (Ser202) mouse monoclonal antibody | 1:100 | gift from Peter Davies |
| PHF-1 | Phospho-tau (Ser396/Ser404) mouse monoclonal antibody | 1:100 | gift from Peter Davies |
| AT8 | Phospho-tau (Ser202, Thr205) mouse monoclonal antibody | 1:100 | Invitrogen, Carlsbad, CA, USA |
| NTD (n = 9) | DS (n = 8) | p-Value | |
|---|---|---|---|
| Age (wk) | 64.87 ± 14.49 * Min. 50.38; Max. 79.36 | 72.80 ± 18.29 * Min. 54.51; Max. 91.09 | p > 0.05 a |
| Brain Weight (g) | 653.08 ± 97.15 * Min. 555.93; Max. 750.23 | 544.75 ± 101.40 * Min. 443.35; Max. 646.15 | p > 0.05 a |
| Body Weight (kg) | 6.38 ± 3.29 * Min. 2.59; Max. 13.10 | 6.036 ± 3.81 * Min. 2.50; Max. 14.40 | p > 0.05 a |
| Height (cm) | 61.66 ± 15.64 * Min. 39; Max. 90.10 | 61.45 ± 18.78 * Min. 48; Max. 97 | p > 0.05 a |
| PMI (h) | 19.71 ± 2.99 * Min. 16.72; Max. 22.70 | 24.37 ± 5.41 * Min. 18.96; Max. 29.780 | p > 0.05 a |
| Gender (%) | Male (55.55%); Female (44.44%) | Male (62.50%); Female (37.50%) | p > 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, D.G.; Utagawa, E.C.; Arva, N.C.; Schafernak, K.T.; Mufson, E.J.; Perez, S.E. Postnatal Cytoarchitecture and Neurochemical Hippocampal Dysfunction in Down Syndrome. J. Clin. Med. 2021, 10, 3414. https://doi.org/10.3390/jcm10153414
Moreno DG, Utagawa EC, Arva NC, Schafernak KT, Mufson EJ, Perez SE. Postnatal Cytoarchitecture and Neurochemical Hippocampal Dysfunction in Down Syndrome. Journal of Clinical Medicine. 2021; 10(15):3414. https://doi.org/10.3390/jcm10153414
Chicago/Turabian StyleMoreno, David G., Emma C. Utagawa, Nicoleta C. Arva, Kristian T. Schafernak, Elliott J. Mufson, and Sylvia E. Perez. 2021. "Postnatal Cytoarchitecture and Neurochemical Hippocampal Dysfunction in Down Syndrome" Journal of Clinical Medicine 10, no. 15: 3414. https://doi.org/10.3390/jcm10153414
APA StyleMoreno, D. G., Utagawa, E. C., Arva, N. C., Schafernak, K. T., Mufson, E. J., & Perez, S. E. (2021). Postnatal Cytoarchitecture and Neurochemical Hippocampal Dysfunction in Down Syndrome. Journal of Clinical Medicine, 10(15), 3414. https://doi.org/10.3390/jcm10153414