Development of an Effective and Stable Genotype-Matched Live Attenuated Newcastle Disease Virus Vaccine Based on a Novel Naturally Recombinant Malaysian Isolate Using Reverse Genetics

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Plasmids
2.2. Construction of Helper Plasmids
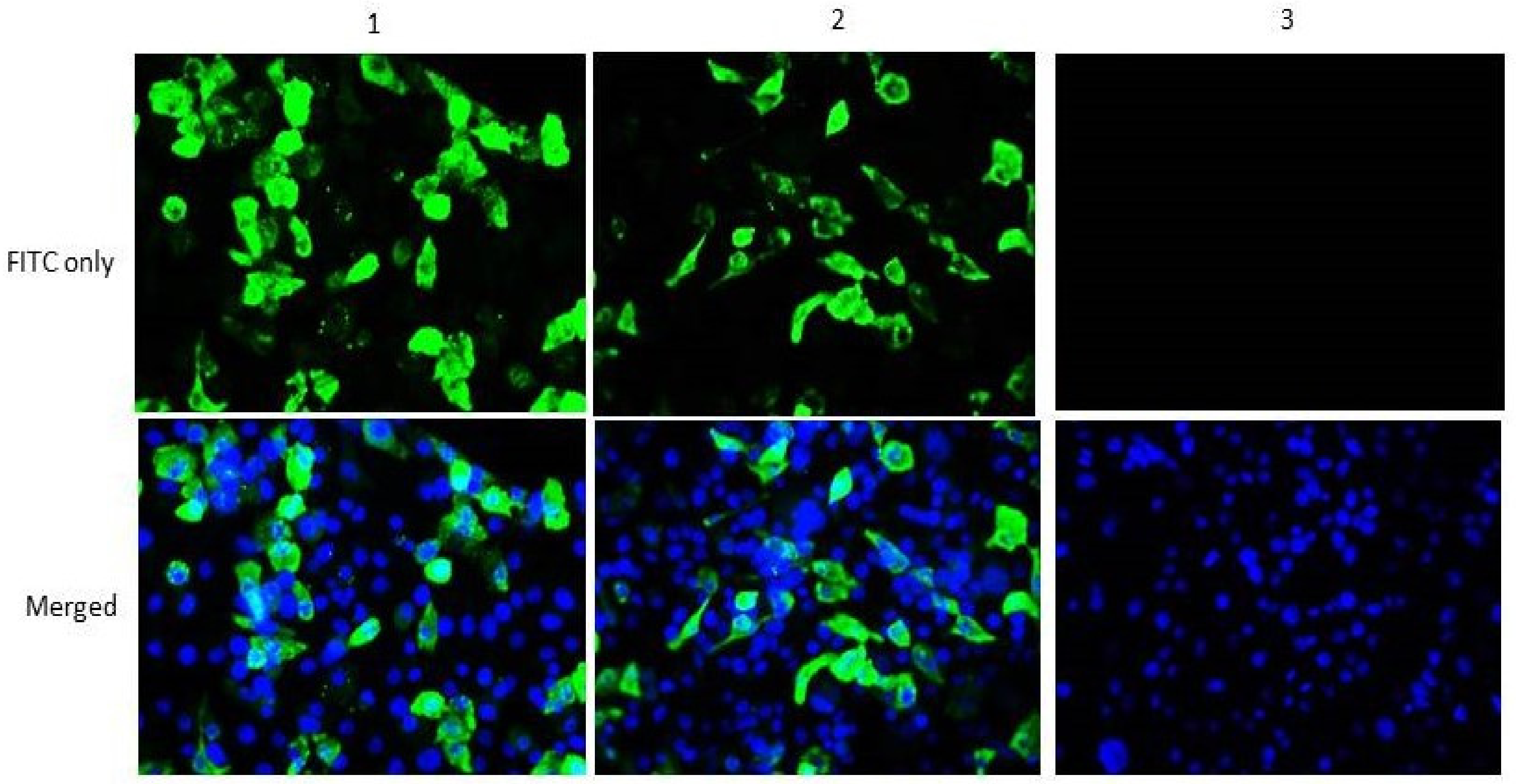
2.3. In Vitro Expression of Helper Plasmids
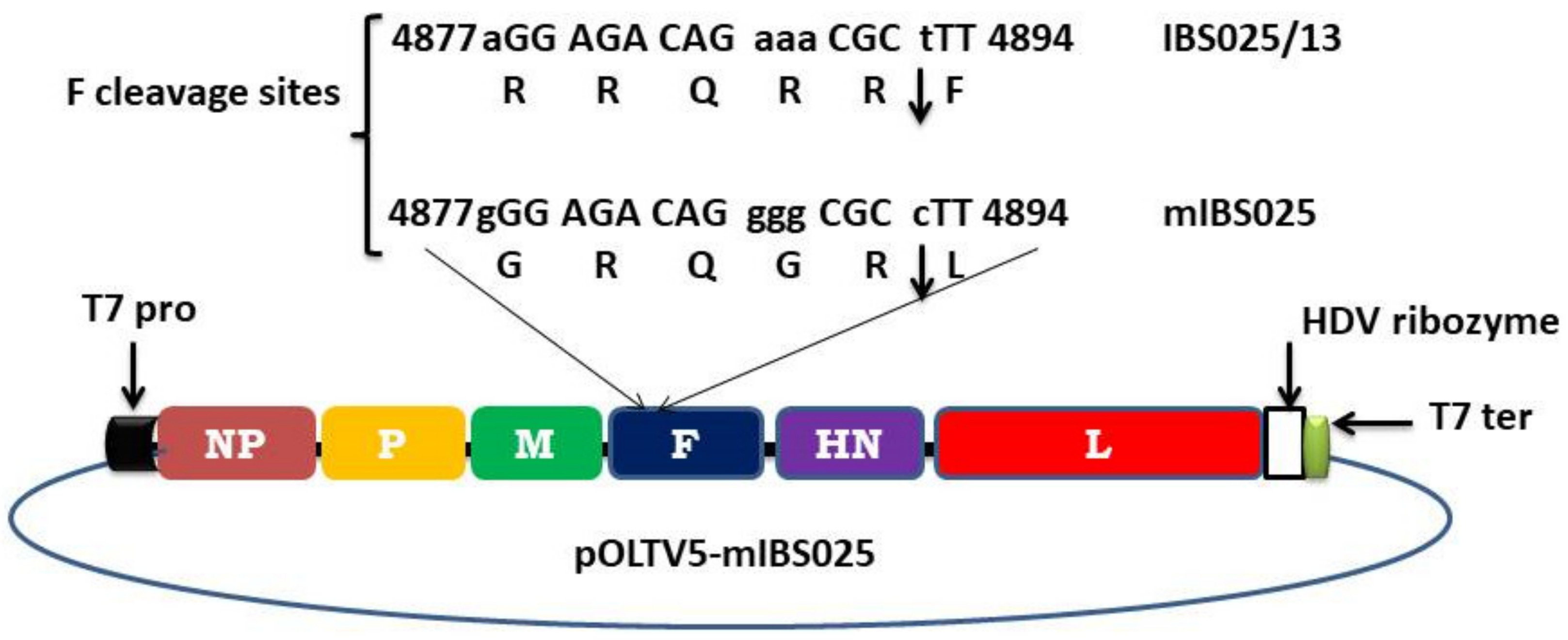
2.4. Construction of Full Length Antigenomic Plasmid
2.5. Recovery of the Recombinant NDV mIBS025
2.6. Ethical Clearance
2.7. Pathogenicity of the Rescued Virus
2.8. Determination of F Protein Cleavage Site and Phenotype Stability
2.9. Vaccine Efficacy Trial
2.10. Serological Testing
2.11. Chicken Trial
2.12. Virus Shedding Determination
2.13. Data Analysis
3. Results
3.1. Construction of Functional Helper Plasmids
3.2. Rescue of Recombinant NDVmIBS025
3.3. Pathogenicity of the Recombinant Virus
3.4. F Protein Cleavage Site and Phenotypic Stability of the Recombinant Virus
3.5. Immunogenicity of NDV Strain mIBS025 and LaSota Vaccines
3.6. Protective Efficacy of NDV mIBS025 and LaSota
3.7. Post Challenge Cloacal and Oropharyngeal Virus Shedding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alexander, D.J.; Aldous, E.W.; Fuller, C.M. The long view: A selective review of 40 years of Newcastle disease research. Avian Pathol. 2012, 41, 329–335. [Google Scholar]
- Ecco, R.; Susta, L.; Afonso, C.L.; Miller, P.J.; Brown, C. Neurological lesions in chickens experimentally infected with virulent Newcastle disease virus isolates. Avian Pathol. 2011, 40, 145–152. [Google Scholar]
- Susta, L.; Jones, M.E.B.; Cattoli, G.; Cardenas-Garcia, S.; Miller, P.J.; Brown, C.C.; Afonso, C.L. Pathologic Characterization of Genotypes XIV and XVII Newcastle Disease Viruses and Efficacy of Classical Vaccination on Specific Pathogen-Free Birds. Vet. Pathol. 2015, 52, 120–131. [Google Scholar]
- Cattoli, G.; Fusaro, A.; Monne, I.; Molia, S.; Le Menach, A.; Maregeya, B.; Nchare, A.; Bangana, I.; Maina, A.G.; N’Goran Koffi, J.N.; et al. Emergence of a new genetic lineage of Newcastle disease virus in West and Central Africa-Implications for diagnosis and control. Vet. Microbiol. 2010, 142, 168–176. [Google Scholar]
- Alexander, D.J. Ecology and Epidemiology of Newcastle Disease. In Avian Influenza and Newcastle Disease: A Field and Laboratory Manual; Springer: New York, NY, USA, 2009; pp. 19–26. [Google Scholar]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar]
- Yusoff, K.; Tan, W.S. Newcastle disease virus: Macromolecules and opportunities. Avian Pathol. 2001, 30, 439–455. [Google Scholar]
- Murulitharan, K.; Yusoff, K.; Omar, A.R.; Molouki, A. Characterization of Malaysian velogenic NDV strain AF2240-I genomic sequence: A comparative study. Virus Genes 2013, 46, 431–440. [Google Scholar]
- Karsunke, J.; Heiden, S.; Murr, M.; Karger, A.; Franzke, K.; Mettenleiter, T.C.; Römer-Oberdörfer, A. W protein expression by Newcastle disease virus. Virus Res. 2019, 263, 207–216. [Google Scholar]
- Hao, H.; Chen, S.; Liu, P.; Ren, S.; Gao, X.; Wang, Y.; Wang, X.; Zhang, S.; Yang, Z. Genetic variation in V gene of class II Newcastle disease virus. Infect. Genet. Evol. 2016, 37, 14–20. [Google Scholar]
- De Leeuw, O.S.; Koch, G.; Hartog, L.; Ravenshorst, N.; Peeters, B.P.H. Virulence of Newcastle disease virus is determined by the cleavage site of the fusion protein and by both the stem region and globular head of the haemagglutinin-neuraminidase protein. J. Gen. Virol. 2005, 86, 1759–1769. [Google Scholar]
- Panda, A.; Huang, Z.; Elankumaran, S.; Rockemann, D.D.; Samal, S.K. Role of fusion protein cleavage site in the virulence of Newcastle disease virus. Microb. Pathog. 2004, 36, 1–10. [Google Scholar]
- OIE. Newcastle disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Mammals, Birds and Bees; Biological Standards Commission: Paris, France, 2012; Volume 1, pp. 555–574. ISBN 9789290447184. [Google Scholar]
- Dortmans, J.C.; Koch, G.; Rottier, P.J.; Peeters, B.P. Virulence of newcastle disease virus: What is known so far? Vet. Res. 2011, 42, 122. [Google Scholar]
- Vijayarani, K.; Muthusamy, S.; Tirumurugaan, K.G.; Sakthivelan, S.M.; Kumanan, K. Pathotyping of a Newcastle disease virus isolated from peacock (Pavo cristatus). Trop. Anim. Health Prod. 2010, 42, 415–419. [Google Scholar]
- Bello, M.B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Omar, A.R. Diagnostic and Vaccination Approaches for Newcastle Disease Virus in Poultry: The Current and Emerging Perspectives. BioMed Res. Int. 2018, 2018, 7278459. [Google Scholar]
- Aldous, E.W.; Mynn, J.K.; Banks, J.; Alexander, D.J. A molecular epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) isolates by phylogenetic analysis of a partial nucleotide sequence of the fusion protein gene. Avian Pathol. 2003, 32, 237–255. [Google Scholar]
- Ballagi-Pordány, A.; Wehmann, E.; Herczeg, J.; Belák, S.; Lomniczi, B. Identification and grouping of Newcastle disease virus strains by restriction site analysis of a region from the F gene. Arch. Virol. 1996, 141, 243–261. [Google Scholar]
- Czeglédi, A.; Ujvári, D.; Somogyi, E.; Wehmann, E.; Werner, O.; Lomniczi, B. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications. Virus Res. 2006, 120, 36–48. [Google Scholar]
- Kim, L.M.; King, D.J.; Suarez, D.L.; Wong, C.W.; Afonso, C.L. Characterization of class I newcastle disease virus isolates from Hong Kong live bird markets and detection using real-time reverse transcription-PCR. J. Clin. Microbiol. 2007, 45, 1310–1314. [Google Scholar]
- Diel, D.G.; da Silva, L.H.A.; Liu, H.; Wang, Z.; Miller, P.J.; Afonso, C.L. Genetic diversity of avian paramyxovirus type 1: Proposal for a unified nomenclature and classification system of Newcastle disease virus genotypes. Infect. Genet. Evol. 2012, 12, 1770–1779. [Google Scholar]
- Snoeck, C.J.; Owoade, A.A.; Couacy-Hymann, E.; Alkali, B.R.; Okwen, M.P.; Adeyanju, A.T.; Komoyo, G.F.; Nakouné, E.; Le Faou, A.; Muller, C.P. High Genetic Diversity of Newcastle Disease Virus in Poultry in West and Central Africa: Cocirculation of Genotype XIV and Newly Defined Genotypes XVII and XVIII. J. Clin. Microbiol. 2013, 51, 2250–2260. [Google Scholar]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet. Evol. 2010, 10, 26–35. [Google Scholar]
- Bello, M.B.; Yusoff, K.M.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Jibril, A.H.; Tambuwal, F.M.; Omar, A.R. Genotype Diversity of Newcastle Disease Virus in Nigeria: Disease Control Challenges and Future Outlook. Adv. Virol. 2018, 2018, 6097291. [Google Scholar]
- Dimitrov, K.M.; Ramey, A.M.; Qiu, X.; Bahl, J.; Afonso, C.L. Infection, Genetics and Evolution Temporal, geographic, and host distribution of avian paramyxovirus 1 ( Newcastle disease virus ). MEEGID 2016, 39, 22–34. [Google Scholar]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar]
- Tan, S.W.; Ideris, A.; Omar, A.R.; Yusoff, K.; Hair-Bejo, M. Sequence and phylogenetic analysis of Newcastle disease virus genotypes isolated in Malaysia between 2004 and 2005. Arch. Virol. 2010, 155, 63–70. [Google Scholar]
- Kwon, H.-J.; Cho, S.-H.; Ahn, Y.-J.; Seo, S.-H.; Choi, K.-S.; Kim, S.-J. Molecular epidemiology of Newcastle disease in Republic of Korea. Vet. Microbiol. 2003, 95, 39–48. [Google Scholar]
- Aljumaili, O.A.; Yeap, S.K.; Omar, A.R.; Aini, I. Isolation and Characterization of Genotype VII Newcastle Disease Virus from NDV Vaccinated Farms in Malaysia. Pertanika J. Trop. Agric. Sci. 2017, 40, 677–690. [Google Scholar]
- Miller, P.J.; Haddas, R.; Simanov, L.; Lublin, A.; Rehmani, S.F.; Wajid, A.; Bibi, T.; Khan, T.A.; Yaqub, T.; Setiyaningsih, S.; et al. Identification of new sub-genotypes of virulent Newcastle disease virus with potential panzootic features. Infect. Genet. Evol. 2015, 29, 216–229. [Google Scholar]
- Roohani, K.; Tan, S.W.; Yeap, S.K.; Ideris, A.; Bejo, M.H.; Omar, A.R. Characterisation of genotype VII Newcastle disease virus (NDV) isolated from NDV vaccinated chickens, and the efficacy of LaSota and recombinant genotype VII vaccines against challenge with velogenic NDV. J. Vet. Sci. 2015, 16, 447–457. [Google Scholar]
- Zhu, J.; Hu, S.; Xu, H.; Liu, J.; Zhao, Z.; Wang, X.; Liu, X. Characterization of virulent Newcastle disease viruses from vaccinated chicken flocks in Eastern China. BMC Vet. Res. 2016, 12, 113. [Google Scholar]
- Jeon, W.J.; Lee, E.K.; Lee, Y.J.; Jeong, O.M.; Kim, Y.J.; Kwon, J.H.; Choi, K.S. Protective efficacy of commercial inactivated Newcastle disease virus vaccines in chickens against a recent Korean epizootic strain. J. Vet. Sci. 2008, 9, 295–300. [Google Scholar]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar]
- Satharasinghe, D.A.; Murulitharan, K.; Tan, S.W.; Yeap, S.K.; Munir, M.; Ideris, A.; Omar, A.R. Detection of Inter-Lineage Natural Recombination in Avian Paramyxovirus Serotype 1 Using Simplified Deep Sequencing Platform. Front. Microbiol. 2016, 7, 1907. [Google Scholar]
- Oyebanji, V.O.; Emikpe, B.O.; Omolade, A.O.; Odeniyi, M.O.; Salami, A.; Osowole, O.I.; Kasali, O.B.; Akinboade, O.A. Evaluation of immune response in challenged chickens vaccinated with Newcastle disease vaccine using gums from Cedrela odorata and Khaya senegalensis as delivery agents. J. Immunoass. Immunochem. 2017, 38, 378–388. [Google Scholar]
- Rasoli, M.; Yeap, S.K.; Tan, S.W.; Moeini, H.; Ideris, A.; Bejo, M.H.; Alitheen, N.B.M.; Kaiser, P.; Omar, A.R. Alteration in lymphocyte responses, cytokine and chemokine profiles in chickens infected with genotype VII and VIII velogenic Newcastle disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 11–21. [Google Scholar]
- Winterfield, R.W.; Dhillon, A.S.; Alby, L.J. Vaccination of chickens against Newcastle disease with live and inactivated Newcastle disease virus. Poult. Sci. 1980, 59, 240–246. [Google Scholar]
- Feizi, A.; Nazeri, M. Comparative Study of Antibody Titers Obtained from Avinew, Lasota, and Clone30 Vaccines in Broiler Chicks with Hi Test. Aust. J. Basic Appl. Sci. 2011, 5, 554–558. [Google Scholar]
- Choi, K.S.; Kye, S.J.; Kim, J.Y.; Lee, H.S. Genetic and antigenic variation of shedding viruses from vaccinated chickens after challenge with virulent Newcastle disease virus. Avian Dis. 2013, 57, 303–306. [Google Scholar]
- Xiao, S.; Nayak, B.; Samuel, A.; Paldurai, A.; Kanabagattebasavarajappa, M.; Prajitno, T.Y.; Bharoto, E.E.; Collins, P.L.; Samal, S.K. Generation by Reverse Genetics of an Effective, Stable, Live-Attenuated Newcastle Disease Virus Vaccine Based on a Currently Circulating, Highly Virulent Indonesian Strain. PLoS ONE 2012, 7, e52751. [Google Scholar]
- Sabra, M.; Dimitrov, K.M.; Goraichuk, I.V.; Wajid, A.; Sharma, P.; Williams-Coplin, D.; Basharat, A.; Rehmani, S.F.; Muzyka, D.V.; Miller, P.J.; et al. Phylogenetic assessment reveals continuous evolution and circulation of pigeon-derived virulent avian avulaviruses 1 in Eastern Europe, Asia, and Africa. BMC Vet. Res. 2017, 13, 291. [Google Scholar]
- Kim, S.-H.; Wanasen, N.; Paldurai, A.; Xiao, S.; Collins, P.L.; Samal, S.K. Newcastle disease virus fusion protein is the major contributor to protective immunity of genotype-matched vaccine. PLoS ONE 2013, 8, e74022. [Google Scholar]
- Bello, M.B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Jibril, A.H.; Peeters, B.P.H.; Omar, A.R. Exploring the Prospects of Engineered Newcastle Disease Virus in Modern Vaccinology. Viruses 2020, 12, 451. [Google Scholar]
- Hu, H.; Roth, J.P.; Estevez, C.N.; Zsak, L.; Liu, B.; Yu, Q. Generation and evaluation of a recombinant Newcastle disease virus expressing the glycoprotein (G) of avian metapneumovirus subgroup C as a bivalent vaccine in turkeys. Vaccine 2011, 29, 8624–8633. [Google Scholar]
- Mohamed Amin, Z.; Che Ani, M.A.; Tan, S.W.; Yeap, S.K.; Alitheen, N.B.; Syed Najmuddin, S.U.F.; Kalyanasundram, J.; Chan, S.C.; Veerakumarasivam, A.; Chia, S.L.; et al. Evaluation of a Recombinant Newcastle Disease Virus Expressing Human IL12 against Human Breast Cancer. Sci. Rep. 2019, 9, 13999. [Google Scholar]
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector Newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar]
- Senne, D.A.; King, D.J.; Kapczynski, D.R. Control of Newcastle disease by vaccination. Dev. Biol. 2004, 119, 165–170. [Google Scholar]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar]
- Han, Q.; Gao, X.; Wu, P.; Xiao, S.; Wang, X.; Liu, P.; Tong, L.; Hao, H.; Zhang, S.; Dang, R.; et al. Re-evaluation the immune efficacy of Newcastle disease virus vaccine in commercial laying chickens. Res. Vet. Sci. 2017, 111, 63–66. [Google Scholar]
- Ecco, R.; Brown, C.; Susta, L.; Cagle, C.; Cornax, I.; Pantin-Jackwood, M.; Miller, P.J.; Afonso, C.L. In vivo transcriptional cytokine responses and association with clinical and pathological outcomes in chickens infected with different Newcastle disease virus isolates using formalin-fixed paraffin-embedded samples. Vet. Immunol. Immunopathol. 2011, 141, 221–229. [Google Scholar]
- Kapczynski, D.R.; King, D.J. Protection of chickens against overt clinical disease and determination of viral shedding following vaccination with commercially available Newcastle disease virus vaccines upon challenge with highly virulent virus from the California 2002 exotic Newcastl. Vaccine 2005, 23, 3424–3433. [Google Scholar]
- Miller, P.J.; Afonso, C.L.; El Attrache, J.; Dorsey, K.M.; Courtney, S.C.; Guo, Z.; Kapczynski, D.R. Effects of Newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Dev. Comp. Immunol. 2013, 41, 505–513. [Google Scholar]
- Miller, P.J.; Kim, L.M.; Ip, H.S.; Afonso, C.L. Evolutionary dynamics of Newcastle disease virus. Virology 2009, 391, 64–72. [Google Scholar]

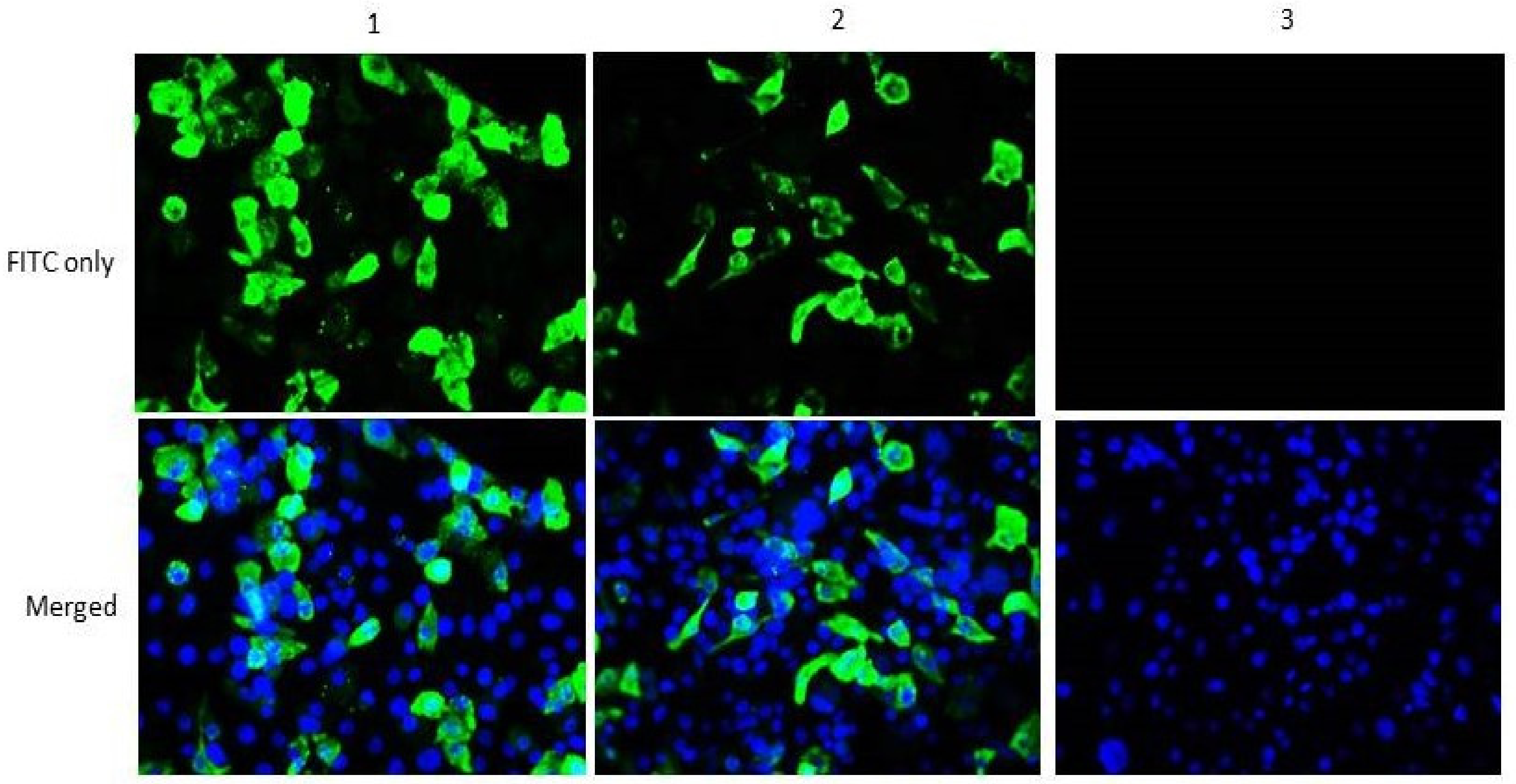{kind=link}
{kind=link}
{kind=link}
| Strain | Accession Number | Genotype | Nucleotide Divergence (%) | Amino Acid Divergence (%) | Year of Isolation | Country of Origin |
|---|---|---|---|---|---|---|
| IBS002 | KR074404.1 | VII | 18.71 | 10.27 | 2011 | Malaysia |
| IBS005 | KR074405.1 | VII | 18.71 | 10.27 | 2011 | Malaysia |
| MB076 | KR074406.1 | VII | 17.07 | 10.18 | 2005 | Malaysia |
| MB128 | KR074407.1 | VII | 17.07 | 10.18 | 2004 | Malaysia |
| IBS025/13 | KT355595.1 | VII | 18.50 | 11.01 | 2013 | Malaysia |
| NCXCP | MG869266.1 | XII | 19.42 | 12.62 | 2011 | Vietnam |
| NDV15A1 | MG869268.1 | XII | 19.10 | 10.71 | 2015 | Vietnam |
| NDVLC15 | MG869269.1 | XII | 18.83 | 11.35 | 2015 | Vietnam |
| NDVQG | MG869270.1 | XII | 18.28 | 11.35 | 2008 | Vietnam |
| NCXKH | MG869271.1 | XII | 18.60 | 11.93 | 2011 | Vietnam |
| Ban 010 | HQ697254.1 | VII | 18.47 | 11.06 | 2010 | Indonesia |
| Suk 019 | HQ697255.1 | VII | 18.45 | 10.46 | 2010 | Indonesia |
| Muk 003 | HQ697256.1 | VII | 18.31 | 10.46 | 2009 | Indonesia |
| Sragen 014 | HQ697258.1 | VII | 18.39 | 11.04 | 2010 | Indonesia |
| Kud 018 | HQ697260.1 | VII | 18.39 | 11.04 | 2010 | Indonesia |
| Bali 020 | HQ697261.1 | VII | 18.40 | 10.73 | 2010 | Indonesia |
| Cockatoo 87 | KF767104.1 | VII | 17.05 | 11.14 | 1988 | Indonesia |
| Lory 88 | KF767105.1 | VII | 17.45 | 10.81 | 1988 | Indonesia |
| C300 | KF767106.1 | VII | 17.26 | 10.62 | 1976 | Indonesia |
| Belitung | MK069428.1 | VII | 19.18 | 11.23 | 2015 | Indonesia |
| Kulonprogo | MK069429.1 | VII | 19.33 | 11.60 | 2017 | Indonesia |
| Virus | Pathogenicity | Virus Titration | |||
|---|---|---|---|---|---|
| ICPI | MDT (Hours) | F Cleavage Site | EID50 | HA Titre | |
| IBS025/13 | 1.86 | 58.4 | 112RRQKRF117 | 9.2 log10 | 10 log2 |
| mIBS025 | 0.00 | 150.4 | 112GRQGRL117 | 8.9 log10 | 8 log2 |
| Stability in SPF Embryonated Eggs | Stability in SPF 1-Day Old Chicks | ||||||
|---|---|---|---|---|---|---|---|
| Passage | ICPI | HA Titre | F Cleavage Site | Passage | RT-PCR | Clinical Signs | F Cleavage Site |
| P1 | 0.00 | 8 log2 | 112GRQGRL117 | P1 | + | NIL | 112GRQGRL117 |
| P3 | 0.13 | 9 log2 | 112GRQGRL117 | P5 | + | NIL | 112GRQGRL117 |
| P5 | 0.00 | 8 log2 | 112GRQGRL117 | P7 | + | NIL | 112GRQGRL117 |
| P10 | - | 9 log2 | 112GRQGRL117 | P10 | + | NIL | 112GRQGRL117 |
| P15 | - | 8 log2 | 112GRQGRL117 | P12 | + | NIL | 112GRQGRL117 |
| Days Post Challenge | PBS Control | LaSota Vaccine | mIBS025 Vaccine |
|---|---|---|---|
| 1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| 2 | 0.43 ± 0.53 | 0.14 ± 0.38 | 0.29 ± 0.49 |
| 3 | 1.14 ± 0.38 | 0.57 ± 0.53 | 0.43 ± 0.53 |
| 4 | 2.5 ± 0.54 | 0.29 ± 0.48 * | 0.43 ± 0.53 * |
| 5 | 2.67 ± 0.57 | 0.0 ± 0.0 * | 0.0 ± 0.0 * |
| 6–14 | Nil | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Time Points | LaSota Vaccine | mIBS025 Vaccine | Control | ||||
|---|---|---|---|---|---|---|---|
| Number of Positive/Total | Virus Copy Number Mean (log10) ± SD | Number of Positive/Total | Virus Copy Number Mean (log 10) ± SD | Number of Positive/Total | Virus Copy Number Mean (Log 10) ± SD | ||
| 3 dpc | 10/10 | 7.38 ± 0.36 * | 10/10 | 6.17 ± 0.40 * | 10/10 | 10.09 ± 0.09 | |
| 5 dpc | 10/10 | 6.81 ± 0.56 * | 9/10 | 5.82 ± 0.21 *,a | 5/5 | 9.81 ± 0.62 | |
| 7 dpc | 8/10 | 5.53 ± 0.88 | 7/10 | 4.59 ± 0.43 | NS | - | |
| 10 dpc | 5/10 | 4.96 ± 0.61 | 2/10 | 2.23 ± 0.03 a | NS | - | |
| 12 dpc | 4/10 | 4.14 ± 0.62 | 0/10 | ND | NS | - | |
| Time Points | LaSota Vaccine | mIBS025 Vaccine | Control | |||
|---|---|---|---|---|---|---|
| Number of Positive/Total | Virus Copy Number Mean (log 10) ± SD | Number of Positive/Total | Virus Copy Number Mean (log 10) ± SD | Number of Positive/Total | Virus Copy Number Mean (log 10) ± SD | |
| 3 dpc | 10/10 | 6.58 ± 0.48 * | 10/10 | 6.06 ± 0.87 * | 10/10 | 9.04 ± 0.96 |
| 5 dpc | 10/10 | 7.02 ± 0.56 | 8/10 | 4.67 ± 0.85 a,* | 5/5 | 9.11 ± 0.05 |
| 7 dpc | 7/10 | 6.19 ± 0.50 * | 6/10 | 4.07 ± 0.53 a | NS | - |
| 10 dpc | 5/10 | 4.92 ± 1.26 | 2/10 | 2.14 ± 0.25 a | NS | - |
| 12 dpc | 4/10 | 3.55 ± 0.49 | 0/10 | ND | NS | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello, M.B.; Mahamud, S.N.A.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Omar, A.R. Development of an Effective and Stable Genotype-Matched Live Attenuated Newcastle Disease Virus Vaccine Based on a Novel Naturally Recombinant Malaysian Isolate Using Reverse Genetics. Vaccines 2020, 8, 270. https://doi.org/10.3390/vaccines8020270
Bello MB, Mahamud SNA, Yusoff K, Ideris A, Hair-Bejo M, Peeters BPH, Omar AR. Development of an Effective and Stable Genotype-Matched Live Attenuated Newcastle Disease Virus Vaccine Based on a Novel Naturally Recombinant Malaysian Isolate Using Reverse Genetics. Vaccines. 2020; 8(2):270. https://doi.org/10.3390/vaccines8020270
Chicago/Turabian StyleBello, Muhammad Bashir, Siti Nor Azizah Mahamud, Khatijah Yusoff, Aini Ideris, Mohd Hair-Bejo, Ben P. H. Peeters, and Abdul Rahman Omar. 2020. "Development of an Effective and Stable Genotype-Matched Live Attenuated Newcastle Disease Virus Vaccine Based on a Novel Naturally Recombinant Malaysian Isolate Using Reverse Genetics" Vaccines 8, no. 2: 270. https://doi.org/10.3390/vaccines8020270
APA StyleBello, M. B., Mahamud, S. N. A., Yusoff, K., Ideris, A., Hair-Bejo, M., Peeters, B. P. H., & Omar, A. R. (2020). Development of an Effective and Stable Genotype-Matched Live Attenuated Newcastle Disease Virus Vaccine Based on a Novel Naturally Recombinant Malaysian Isolate Using Reverse Genetics. Vaccines, 8(2), 270. https://doi.org/10.3390/vaccines8020270