
Intranasal Vaccination Affords Localization and Persistence of Antigen-Specific CD8+ T Lymphocytes in the Female Reproductive Tract

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents
2.3. Adoptive Transfer of OT-I Cells and In Vivo Blocking of CXCR3
2.4. Immunizations
2.5. Analyses of the Phenotype of Antigen Specific T Lymphocytes in FRT
2.6. Statistical Analysis
3. Results and Discussion
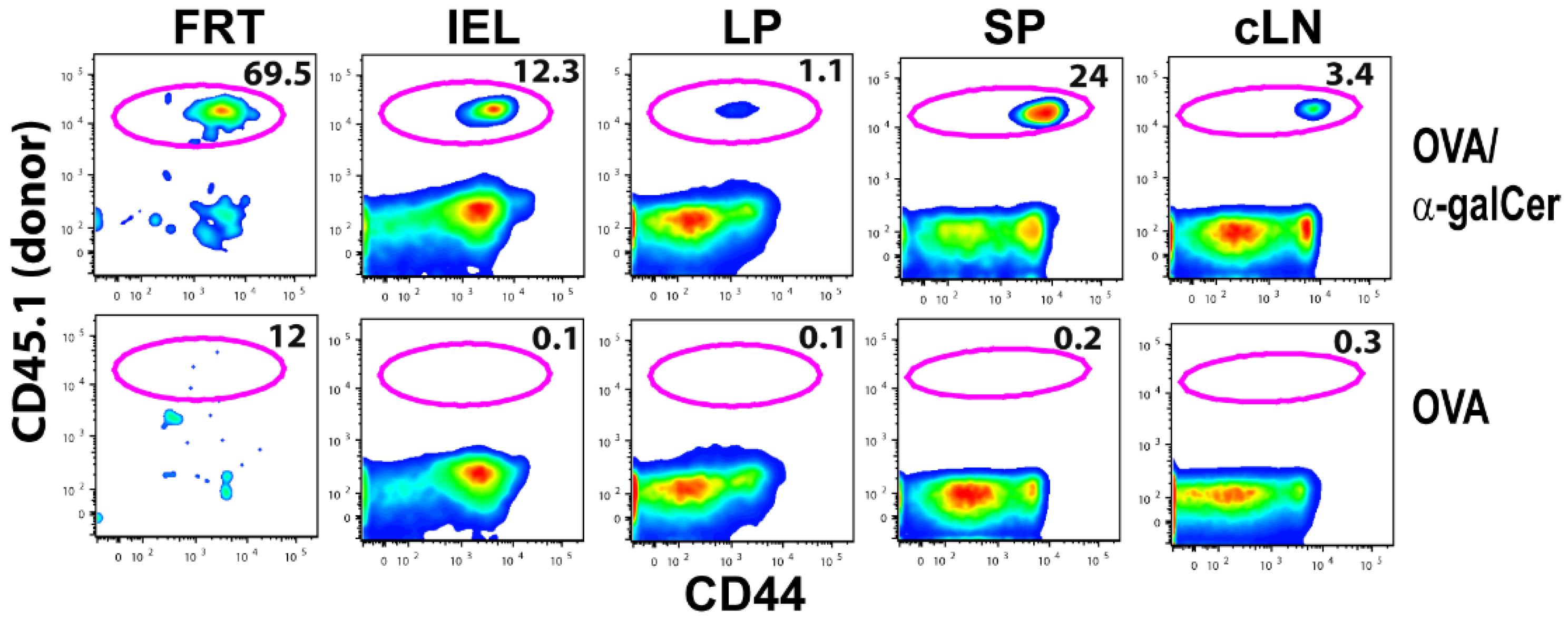
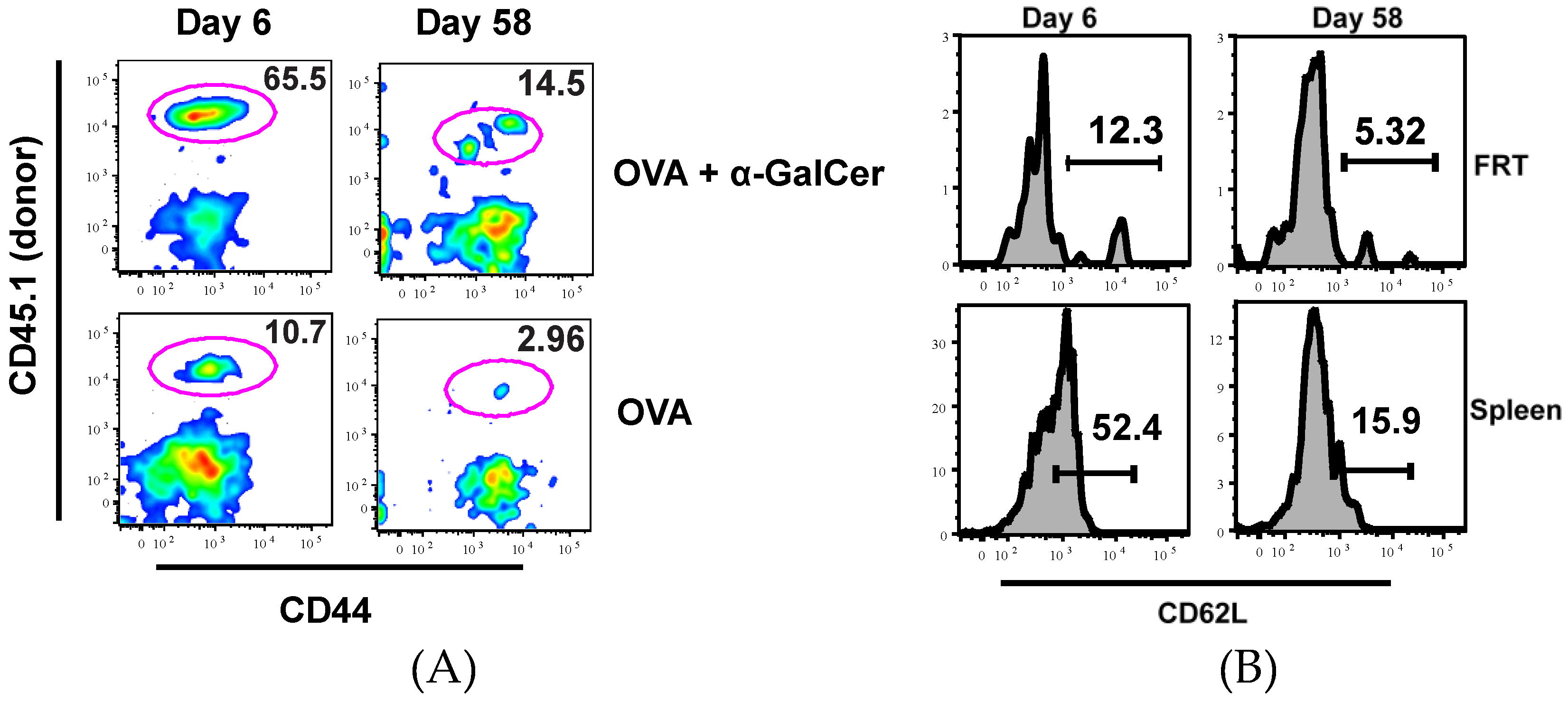
3.1. Intranasal Immunization Preferentially Induces Antigen-Specific CD8+ T Lymphocytes in the FRT
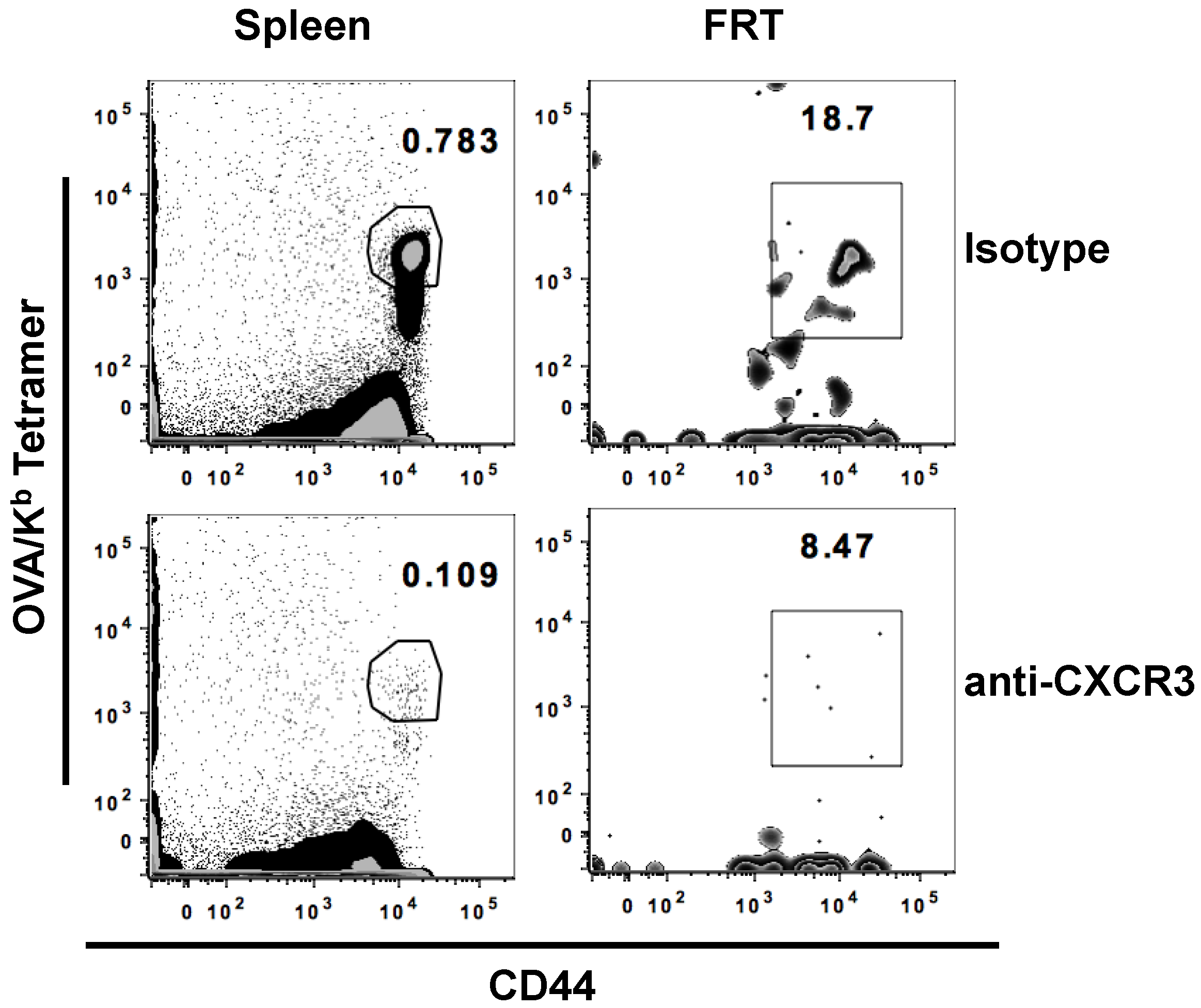
3.2. Potential Role of CXCR3 Signaling for Intranasal Induction of Antigen-Specific CD8+ T Cells Migration into FRT
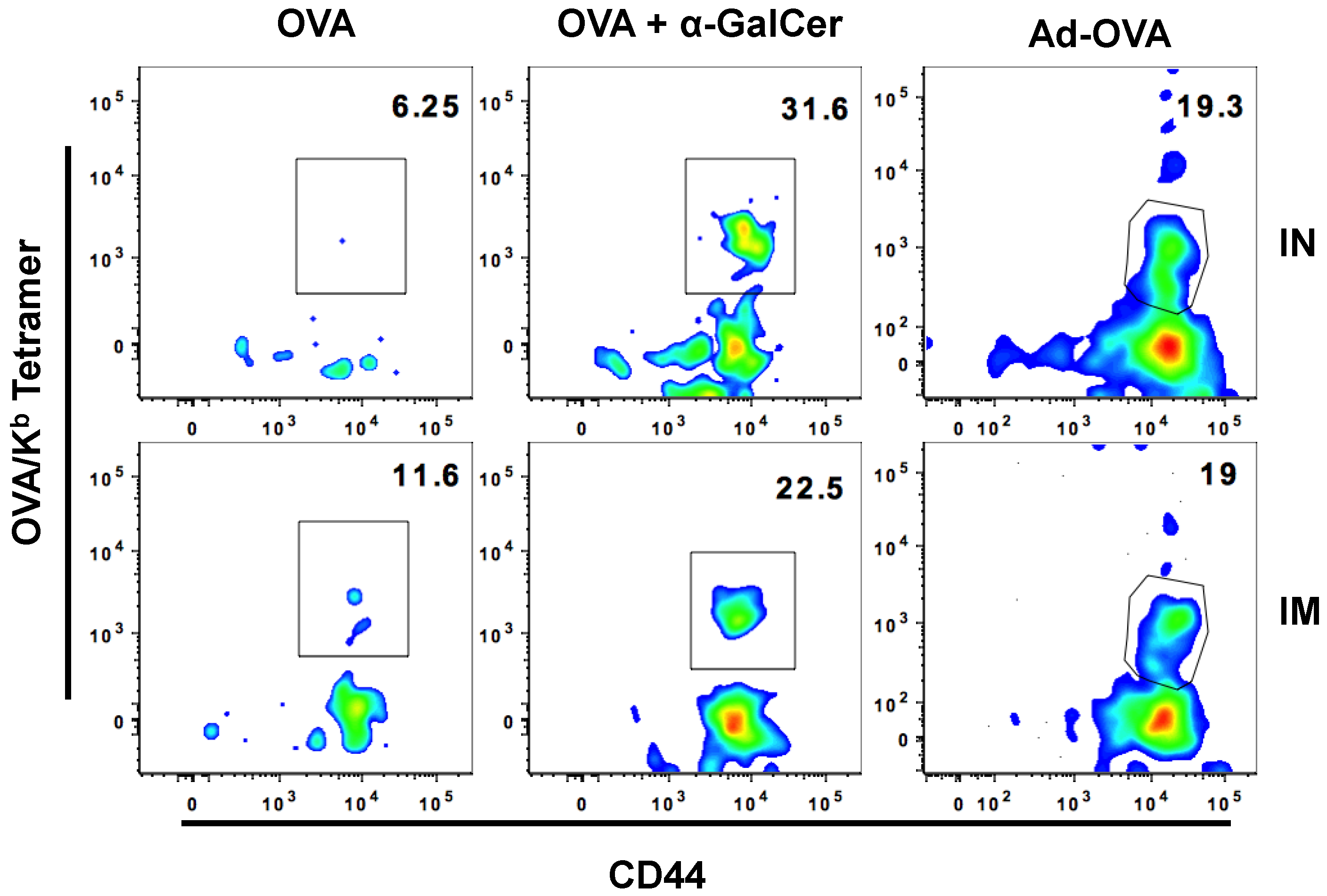
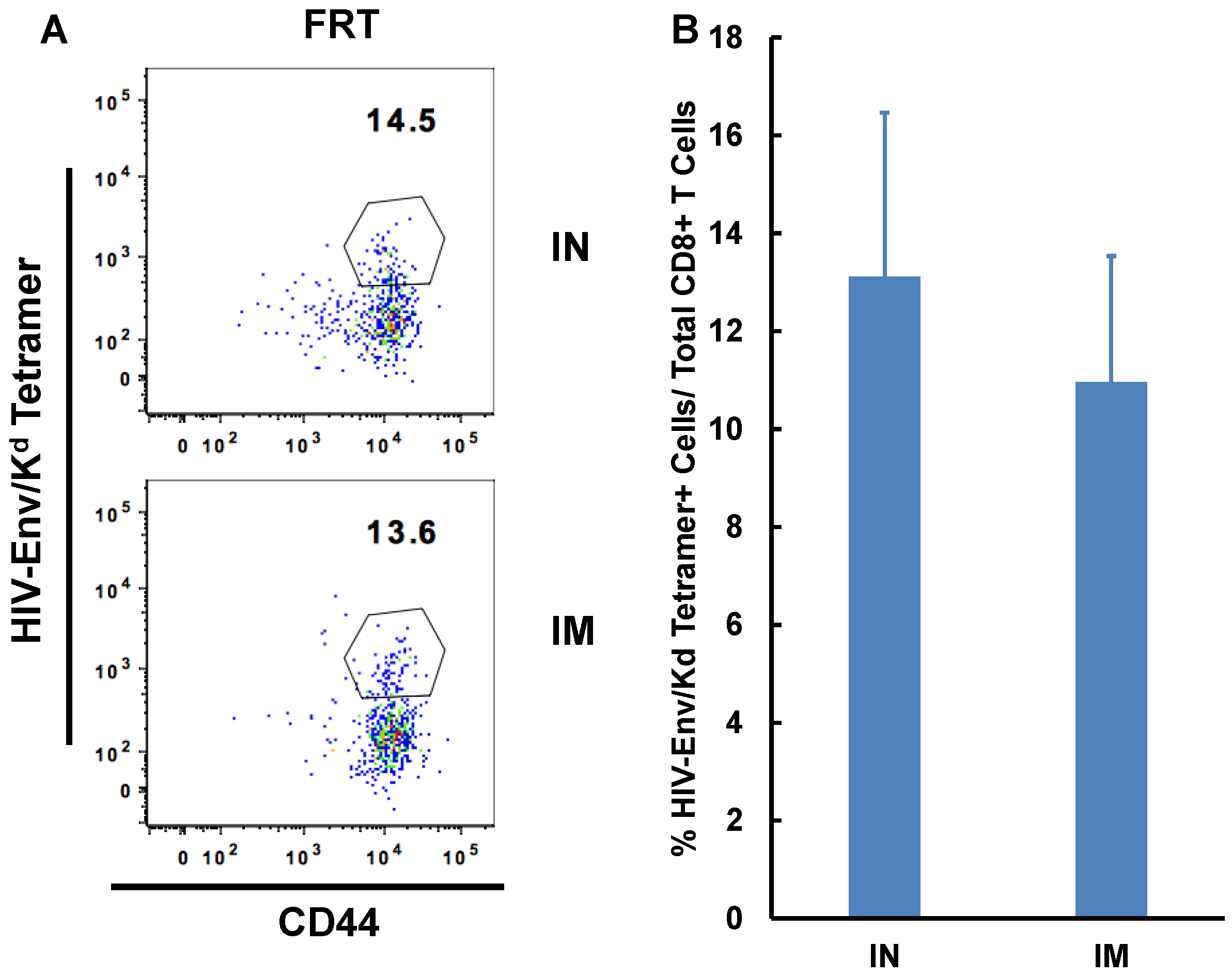
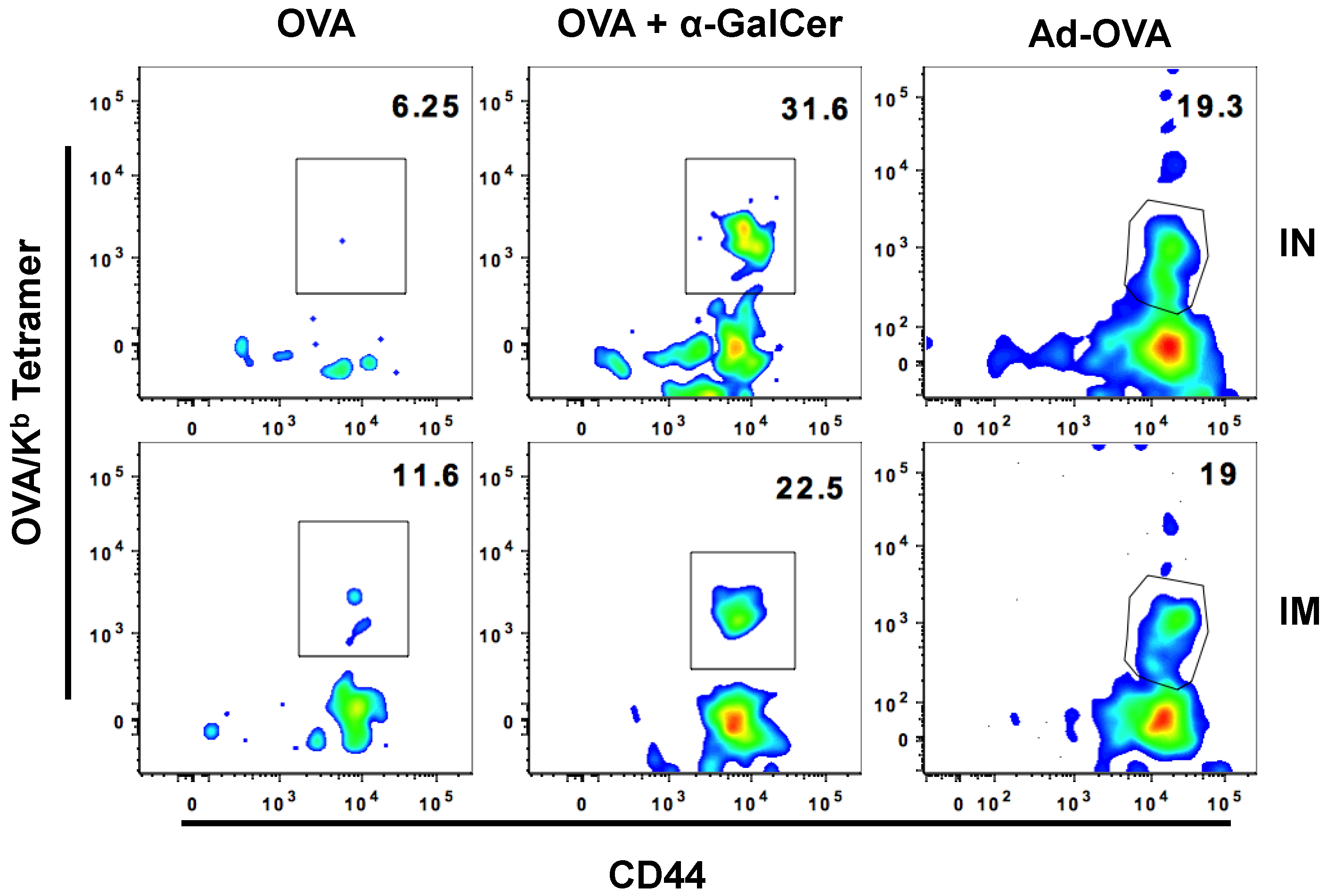
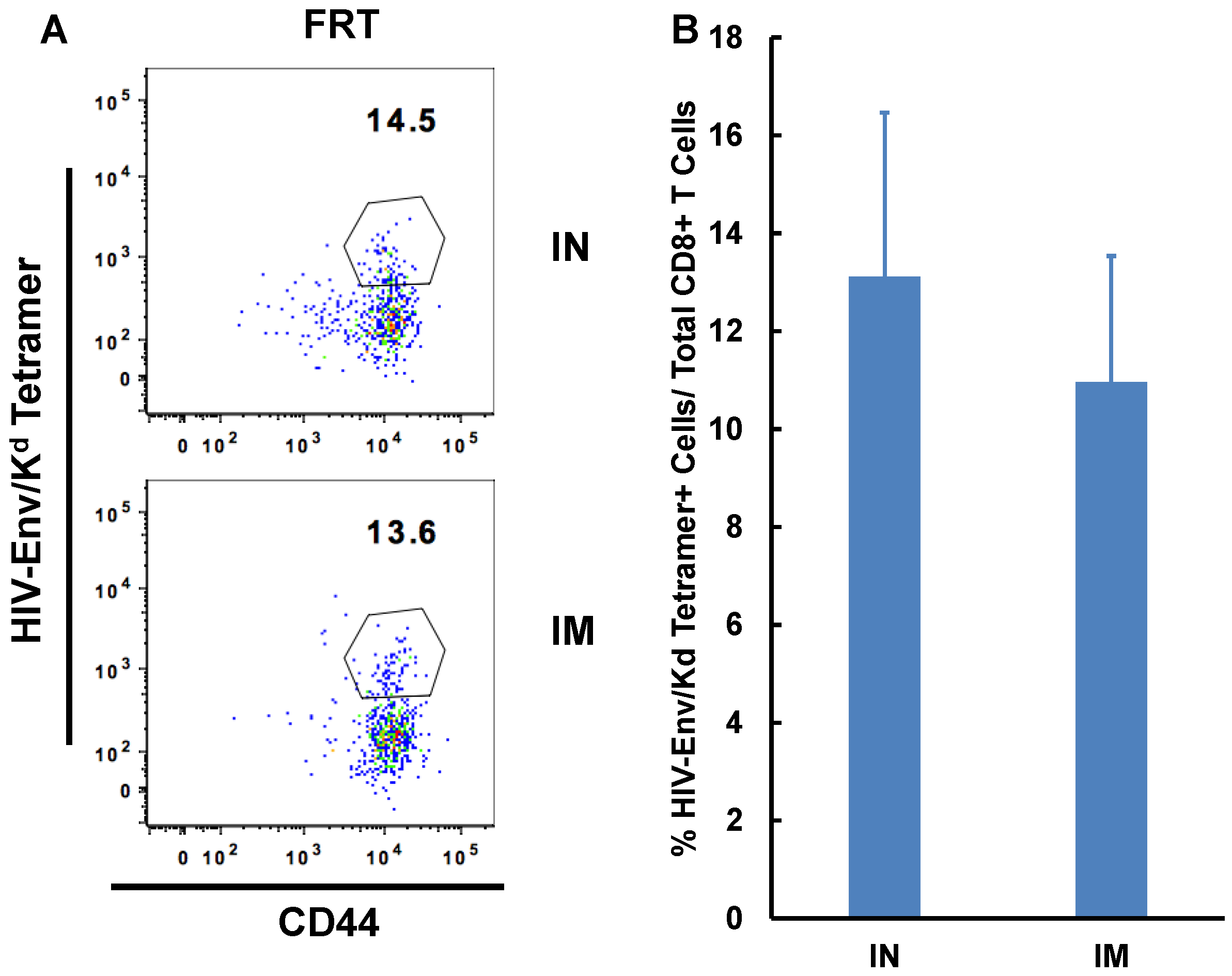
3.3. IN Route is Effective to Deliver Both Protein and DNA/Virus-Vector Based Vaccines for Efficient Generation and Localization of Antigen-Specific CD8+ T Cell Responses in the Female Reproductive Tract
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| FRT | Female Reproductive Tract |
| IN | Intranasal |
| IM | Intramuscular |
| OVA | Ovalbumin |
| ENV | HIV envelope |
| Ad | Adenovirus vector |
| αGalCer | Alpha galactosylceramide |
References
- Holmgren, J.; Czerkinsky, C. Mucosal immunity and vaccines. Nat. Med. 2005, 11, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Neutra, M.R.; Kozlowski, P.A. Mucosal vaccines: The promise and the challenge. Nat. Rev. Immunol. 2006, 6, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Belyakov, I.M.; Ahlers, J.D. What role does the route of immunization play in the generation of protective immunity against mucosal pathogens? J. Immunol. 2009, 183, 6883–6892. [Google Scholar] [CrossRef] [PubMed]
- Belyakov, I.M.; Ahlers, J.D. Mucosal immunity and HIV-1 infection: Applications for mucosal aids vaccine development. Curr. Top. Microbiol. Immunol. 2012, 354, 157–179. [Google Scholar] [PubMed]
- Vasilevsky, S.; Greub, G.; Nardelli-Haefliger, D.; Baud, D. Genital chlamydia trachomatis: Understanding the roles of innate and adaptive immunity in vaccine research. Clin. Microbiol. Rev. 2014, 27, 346–370. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Suwanto, A.; Okabe, M.; Sato, S.; Nochi, T.; Imai, T.; Koyanagi, N.; Kunisawa, J.; Kawaguchi, Y.; Kiyono, H. Vaginal memory T cells induced by intranasal vaccination are critical for protective T cell recruitment and prevention of genital HSV-2 disease. J. Virol. 2014, 88, 13699–13708. [Google Scholar] [CrossRef] [PubMed]
- Schenkel, J.M.; Fraser, K.A.; Vezys, V.; Masopust, D. Sensing and alarm function of resident memory CD8(+) T cells. Nat. Immunol. 2013, 14, 509–513. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.R.; Bienenstock, J. Evidence for a common mucosal immunologic system. I. Migration of b immunoblasts into intestinal, respiratory, and genital tissues. J. Immunol. 1979, 122, 1892–1898. [Google Scholar] [PubMed]
- Czerkinsky, C.; Holmgren, J. Topical immunization strategies. Mucosal. Immunol. 2010, 3, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Igietseme, J.U.; Uriri, I.M.; Kumar, S.N.; Ananaba, G.A.; Ojior, O.O.; Momodu, I.A.; Candal, D.H.; Black, C.M. Route of infection that induces a high intensity of gamma interferon-secreting T cells in the genital tract produces optimal protection against chlamydia trachomatis infection in mice. Infect. Immun. 1998, 66, 4030–4035. [Google Scholar] [PubMed]
- Johansson, E.L.; Wassen, L.; Holmgren, J.; Jertborn, M.; Rudin, A. Nasal and vaginal vaccinations have differential effects on antibody responses in vaginal and cervical secretions in humans. Infect. Immun. 2001, 69, 7481–7486. [Google Scholar] [CrossRef] [PubMed]
- De Bernardis, F.; Boccanera, M.; Adriani, D.; Girolamo, A.; Cassone, A. Intravaginal and intranasal immunizations are equally effective in inducing vaginal antibodies and conferring protection against vaginal candidiasis. Infect. Immun. 2002, 70, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.B.; Butcher, E.C. Homing of naive and memory t lymphocyte subsets to peyer’s patches, lymph nodes, and spleen. J. Immunol. 1997, 159, 1746–1752. [Google Scholar] [PubMed]
- Lefrancois, L.; Parker, C.M.; Olson, S.; Muller, W.; Wagner, N.; Schon, M.P.; Puddington, L. The role of beta7 integrins in CD8 T cell trafficking during an antiviral immune response. J. Exp. Med. 1999, 189, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Johansson-Lindbom, B.; Svensson, M.; Wurbel, M.A.; Malissen, B.; Marquez, G.; Agace, W. Selective generation of gut tropic t cells in gut-associated lymphoid tissue (galt): Requirement for galt dendritic cells and adjuvant. J. Exp. Med. 2003, 198, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Anjuere, F.; Bekri, S.; Bihl, F.; Braud, V.M.; Cuburu, N.; Czerkinsky, C.; Hervouet, C.; Luci, C. B cell and T cell immunity in the female genital tract: Potential of distinct mucosal routes of vaccination and role of tissue-associated dendritic cells and natural killer cells. Clin. Microbiol. Infect. 2012, 18, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Courtney, A.N.; Nehete, P.N.; Nehete, B.P.; Thapa, P.; Zhou, D.; Sastry, K.J. Alpha-galactosylceramide is an effective mucosal adjuvant for repeated intranasal or oral delivery of hiv peptide antigens. Vaccine 2009, 27, 3335–3341. [Google Scholar] [CrossRef] [PubMed]
- Courtney, A.N.; Thapa, P.; Singh, S.; Wishahy, A.M.; Zhou, D.; Sastry, J. Intranasal but not intravenous delivery of the adjuvant alpha-galactosylceramide permits repeated stimulation of natural killer T cells in the lung. Eur. J Immunol. 2011, 41, 3312–3322. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Yang, G.; Byrareddy, S.N.; Barry, M.A.; Sastry, K.J. Natural killer T cell and TLR9 agonists as mucosal adjuvants for sublingual vaccination with clade c HIV-1 envelope protein. Vaccine 2014, 32, 6934–6940. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Yang, G.; Schluns, K.S.; Anthony, S.M.; Sastry, K.J. Sublingual vaccination induces mucosal and systemic adaptive immunity for protection against lung tumor challenge. PLoS ONE 2014, 9, e90001. [Google Scholar] [CrossRef] [PubMed]
- Fraillery, D.; Baud, D.; Pang, S.Y.; Schiller, J.; Bobst, M.; Zosso, N.; Ponci, F.; Nardelli-Haefliger, D. Salmonella enterica serovar Typhi Ty21a expressing human papillomavirus type 16 lL as a potential live vaccine against cervical cancer and typhoid fever. Clin. Vaccine Immunol. 2007, 14, 1285–1295. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Weaver, E.A.; Nehete, P.N.; Nehete, B.P.; Buchl, S.J.; Palmer, D.; Montefiori, D.C.; Ng, P.; Sastry, K.J.; Barry, M.A. Protection against mucosal SHIV challenge by peptide and helper-dependent adenovirus vaccines. Viruses 2009, 1, 920–938. [Google Scholar] [CrossRef] [PubMed]
- Lefrancois, L.; Lycke, N. Isolation of mouse small intestinal intraepithelial lymphocytes, peyer’s patch, and lamina propria cells. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]
- Singh, S.; Nehete, P.; Hanley, P.; Nehete, B.; Yang, G.; He, H.; Anthony, S.M.; Schluns, K.S.; Sastry, K.J. Procedures for mucosal immunization and analyses of cellular immune response to candidate hiv vaccines in murine and nonhuman primate models. Methods Mol. Biol. 2014, 1184, 417–455. [Google Scholar] [PubMed]
- Shin, H.; Iwasaki, A. A vaccine strategy that protects against genital herpes by establishing local memory T cells. Nature 2012, 491, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Casey, K.A.; Fraser, K.A.; Schenkel, J.M.; Moran, A.; Abt, M.C.; Beura, L.K.; Lucas, P.J.; Artis, D.; Wherry, E.J.; Hogquist, K.; et al. Antigen-independent differentiation and maintenance of effector-like resident memory T cells in tissues. J. Immunol. 2012, 188, 4866–4875. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.R.; Ko, H.J.; Kim, E.D.; Chang, S.Y.; Seo, S.U.; Cuburu, N.; Ryu, S.; Kim, S.; Kweon, M.N. Mucosa-associated epithelial chemokine/CCL28 expression in the uterus attracts CCR10+ IGA plasma cells following mucosal vaccination via estrogen control. J. Immunol. 2011, 187, 3044–3052. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, Y.; Lu, B.; Gerard, C.; Iwasaki, A. CD8(+) T lymphocyte mobilization to virus-infected tissue requires CD4(+) T-cell help. Nature 2009, 462, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Thapa, M.; Carr, D.J. CXCR3 deficiency increases susceptibility to genital herpes simplex virus type 2 infection: Uncoupling of CD8+ T-cell effector function but not migration. J. Virol. 2009, 83, 9486–9501. [Google Scholar] [CrossRef] [PubMed]
- Slutter, B.; Pewe, L.L.; Kaech, S.M.; Harty, J.T. Lung airway-surveilling CXCR3(hi) memory CD8(+) T cells are critical for protection against influenza a virus. Immunity 2013, 39, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Deruaz, M.; Luster, A.D. Chemokine-mediated immune responses in the female genital tract mucosa. Immunol. Cell Biol. 2015, 93, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Ogra, P.L.; Faden, H.; Welliver, R.C. Vaccination strategies for mucosal immune responses. Clin. Microbiol. Rev. 2001, 14, 430–445. [Google Scholar] [CrossRef] [PubMed]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, D.R.; Liu, J.; Carville, A.; Mansfield, K.G.; Havenga, M.J.; Goudsmit, J.; Barouch, D.H. Trafficking of antigen-specific CD8+ T lymphocytes to mucosal surfaces following intramuscular vaccination. J. Immunol. 2008, 181, 4188–4198. [Google Scholar] [CrossRef] [PubMed]
- Bagarazzi, M.L.; Boyer, J.D.; Javadian, M.A.; Chattergoon, M.A.; Shah, A.R.; Cohen, A.D.; Bennett, M.K.; Ciccarelli, R.B.; Ugen, K.E.; Weiner, D.B. Systemic and mucosal immunity is elicited after both intramuscular and intravaginal delivery of human immunodeficiency virus type 1 DNA plasmid vaccines to pregnant chimpanzees. J. Infect. Dis. 1999, 180, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.W.; Cun, A.S.; Harris-McCoy, K.; Ertl, H.C. Intramuscular rather than oral administration of replication-defective adenoviral vaccine vector induces specific CD8+ T cell responses in the gut. Vaccine 2007, 25, 2187–2193. [Google Scholar] [CrossRef] [PubMed]
- von Andrian, U.H.; Mackay, C.R. T-cell function and migration. Two sides of the same coin. N. Engl. J. Med. 2000, 343, 1020–1034. [Google Scholar] [PubMed]
- Johansson-Lindbom, B.; Agace, W.W. Generation of gut-homing T cells and their localization to the small intestinal mucosa. Immunol. Rev. 2007, 215, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Masopust, D.; Choo, D.; Vezys, V.; Wherry, E.J.; Duraiswamy, J.; Akondy, R.; Wang, J.; Casey, K.A.; Barber, D.L.; Kawamura, K.S.; et al. Dynamic T cell migration program provides resident memory within intestinal epithelium. J. Exp. Med. 2010, 207, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Stagg, A.J.; Kamm, M.A.; Knight, S.C. Intestinal dendritic cells increase T cell expression of alpha4beta7 integrin. Eur. J. Immunol. 2002, 32, 1445–1454. [Google Scholar] [CrossRef]
- Mora, J.R.; Bono, M.R.; Manjunath, N.; Weninger, W.; Cavanagh, L.L.; Rosemblatt, M.; Von Andrian, U.H. Selective imprinting of gut-homing T cells by peyer’s patch dendritic cells. Nature 2003, 424, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Wuest, T.R.; Carr, D.J. Dysregulation of CXCR3 signaling due to cxcl10 deficiency impairs the antiviral response to herpes simplex virus 1 infection. J. Immunol. 2008, 181, 7985–7993. [Google Scholar] [CrossRef] [PubMed]
- Shane, H.L.; Klonowski, K.D. Every breath you take: The impact of environment on resident memory CD8 T cells in the lung. Front. Immunol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Ariotti, S.; Beltman, J.B.; Borsje, R.; Hoekstra, M.E.; Halford, W.P.; Haanen, J.B.; de Boer, R.J.; Schumacher, T.N. Subtle CXCR3-dependent chemotaxis of ctls within infected tissue allows efficient target localization. J. Immunol. 2015, 195, 5285–5295. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood | Spleen | |||
|---|---|---|---|---|
| OVA | OVA + αGalCer | OVA | OVA + αGalCer | |
| Day 6 | 0.66 (± 0.41) | 11.02(± 5.72) | 0.26(± 0.03) | 3.09 (± 1.39) |
| Day 58 | 0.03(± 0.01) | 0.75(± 0.69) | 0.001(± 0.002) | 0.79(± 0.36) |
| OVA | OVA + αGalCer | |
|---|---|---|
| Day 6 | 4(± 3) | 104(± 59) |
| Day 58 | 7(± 5) | 42(± 14) |
| Isotype | a-CXCR3 | |
|---|---|---|
| Spleen | 0.71 (±0.53) | 0.14(±0.13) * |
| DLN | 0.15(±0.02) | 0.06(±0.01) * |
| FRT | 15.52 (±3.22) | 10.97(±2.86) * |
| IN | IM | |
|---|---|---|
| OVA | 0 | 5.412(± 6.01) |
| OVA + αGalCer | 16.45(± 9.98) | 19.34(± 17.01) |
| Ad-OVA | 30.66 (± 8.93) | 33.95(± 8.54) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Schluns, K.S.; Yang, G.; Anthony, S.M.; Barry, M.A.; Sastry, K.J. Intranasal Vaccination Affords Localization and Persistence of Antigen-Specific CD8+ T Lymphocytes in the Female Reproductive Tract. Vaccines 2016, 4, 7. https://doi.org/10.3390/vaccines4010007
Singh S, Schluns KS, Yang G, Anthony SM, Barry MA, Sastry KJ. Intranasal Vaccination Affords Localization and Persistence of Antigen-Specific CD8+ T Lymphocytes in the Female Reproductive Tract. Vaccines. 2016; 4(1):7. https://doi.org/10.3390/vaccines4010007
Chicago/Turabian StyleSingh, Shailbala, Kimberly S. Schluns, Guojun Yang, Scott M. Anthony, Michael A. Barry, and K. Jagannadha Sastry. 2016. "Intranasal Vaccination Affords Localization and Persistence of Antigen-Specific CD8+ T Lymphocytes in the Female Reproductive Tract" Vaccines 4, no. 1: 7. https://doi.org/10.3390/vaccines4010007
APA StyleSingh, S., Schluns, K. S., Yang, G., Anthony, S. M., Barry, M. A., & Sastry, K. J. (2016). Intranasal Vaccination Affords Localization and Persistence of Antigen-Specific CD8+ T Lymphocytes in the Female Reproductive Tract. Vaccines, 4(1), 7. https://doi.org/10.3390/vaccines4010007