
Pooled Analysis of the Effect of Pre-Existing Ad5 Neutralizing Antibodies on the Immunogenicity of Adenovirus Type 5 Vector-Based COVID-19 Vaccine from Eight Clinical Trials

Abstract
1. Introduction
2. Methods
2.1. Study Design and Data Source
2.2. Antibody Detection
2.3. Statistical Analysis
3. Results
3.1. Study Population
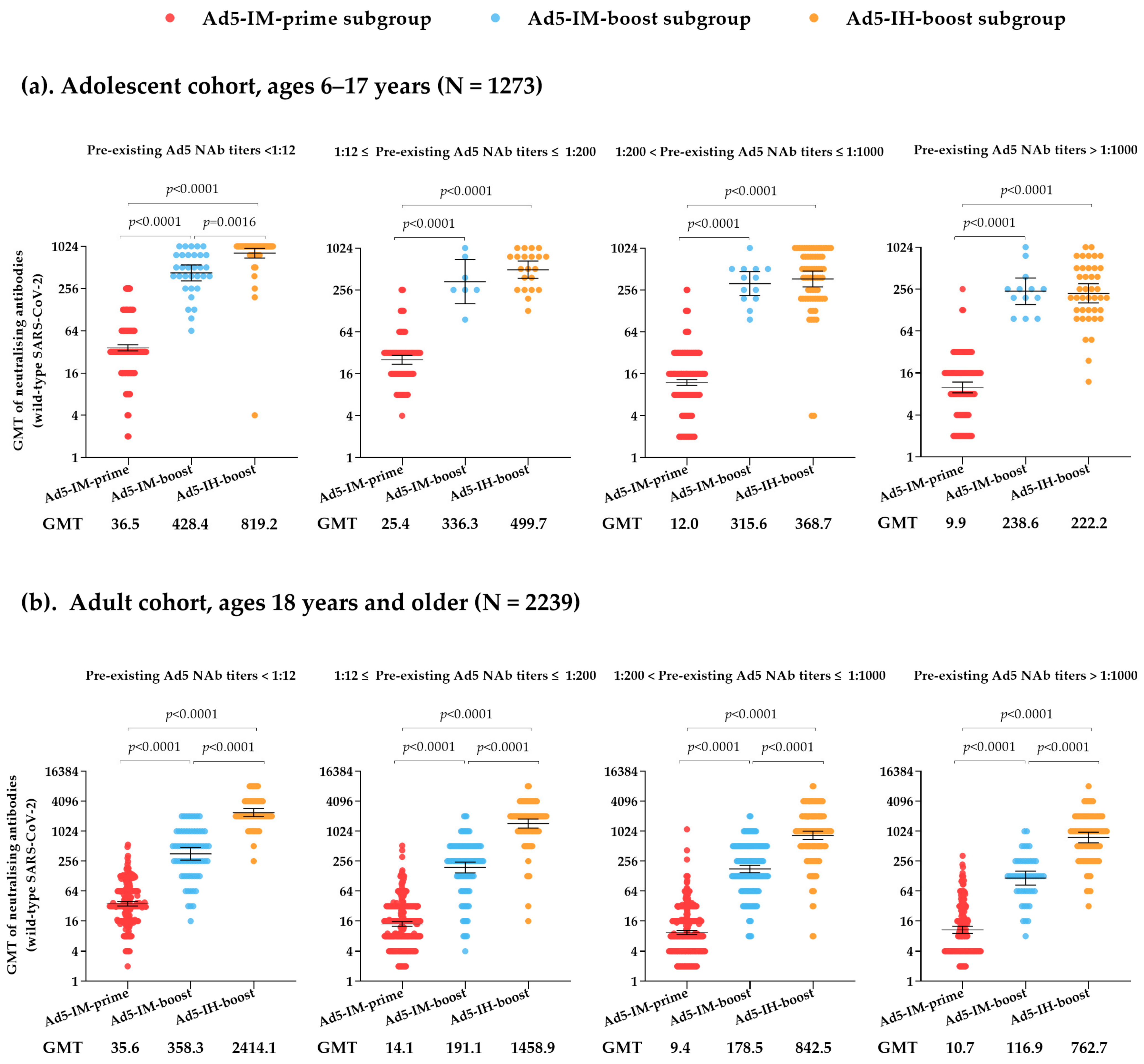
3.2. Impact of Pre-Existing Ad5 NAbs on Live SARS-CoV-2 NAbs After Receiving Ad5-nCoV as a Primary Immunization Strategy
3.3. Impact of Pre-Existing Ad5 NAbs on Live SARS-CoV-2 NAbs After Receiving Ad5-nCoV as a Heterologous Boosting Immunization Strategy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russell, W.C. Adenoviruses: Update on structure and function. J. Gen. Virol. 2009, 90, 1–20. [Google Scholar] [CrossRef]
- Tatsis, N.; Ertl, H.C. Adenoviruses as vaccine vectors. Mol. Ther. 2004, 10, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Bangari, D.S.; Mittal, S.K. Development of nonhuman adenoviruses as vaccine vectors. Vaccine 2006, 24, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Li, C.; Hu, P.; Wang, Q.; Zheng, X.; Zhao, Y.; Shi, Y.; Yang, S.; Yi, C.; Feng, Y.; et al. An adenovirus serotype 2-vectored ebolavirus vaccine generates robust antibody and cell-mediated immune responses in mice and rhesus macaques. Emerg. Microbes Infect. 2018, 7, 101. [Google Scholar] [CrossRef]
- Tiemessen, M.M.; Solforosi, L.; Dekking, L.; Czapska-Casey, D.; Serroyen, J.; Sullivan, N.J.; Volkmann, A.; Pau, M.G.; Callendret, B.; Schuitemaker, H.; et al. Protection against Marburg Virus and Sudan Virus in NHP by an Adenovector-Based Trivalent Vaccine Regimen Is Correlated to Humoral Immune Response Levels. Vaccines 2022, 10, 1263. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, N.J.; Sanchez, A.; Rollin, P.E.; Yang, Z.Y.; Nabel, G.J. Development of a preventive vaccine for Ebola virus infection in primates. Nature 2000, 408, 605–609. [Google Scholar] [CrossRef]
- Afrough, S.; Rhodes, S.; Evans, T.; White, R.; Benest, J. Immunologic Dose-Response to Adenovirus-Vectored Vaccines in Animals and Humans: A Systematic Review of Dose-Response Studies of Replication Incompetent Adenoviral Vaccine Vectors when Given via an Intramuscular or Subcutaneous Route. Vaccines 2020, 8, 131. [Google Scholar] [CrossRef]
- Zhu, F.C.; Wurie, A.H.; Hou, L.H.; Liang, Q.; Li, Y.H.; Russell, J.B.; Wu, S.P.; Li, J.X.; Hu, Y.M.; Guo, Q.; et al. Safety and immunogenicity of a recombinant adenovirus type-5 vector-based Ebola vaccine in healthy adults in Sierra Leone: A single-centre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 389, 621–628. [Google Scholar] [CrossRef]
- Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatulin, A.I.; Dzharullaeva, A.S.; Tukhvatulina, N.M.; Shcheblyakov, D.V.; Shmarov, M.M.; Tokarskaya, E.A.; Simakova, Y.V.; Egorova, D.A.; et al. Safety and immunogenicity of GamEvac-Combi, a heterologous VSV- and Ad5-vectored Ebola vaccine: An open phase I/II trial in healthy adults in Russia. Hum. Vaccines Immunother. 2017, 13, 613–620. [Google Scholar] [CrossRef]
- Guo, Q.; Chan, J.F.; Poon, V.K.; Wu, S.; Chan, C.C.; Hou, L.; Yip, C.C.; Ren, C.; Cai, J.P.; Zhao, M.; et al. Immunization With a Novel Human Type 5 Adenovirus-Vectored Vaccine Expressing the Premembrane and Envelope Proteins of Zika Virus Provides Consistent and Sterilizing Protection in Multiple Immunocompetent and Immunocompromised Animal Models. J. Infect. Dis. 2018, 218, 365–377. [Google Scholar] [CrossRef]
- Baden, L.R.; Stieh, D.J.; Sarnecki, M.; Walsh, S.R.; Tomaras, G.D.; Kublin, J.G.; McElrath, M.J.; Alter, G.; Ferrari, G.; Montefiori, D.; et al. Safety and immunogenicity of two heterologous HIV vaccine regimens in healthy, HIV-uninfected adults (TRAVERSE): A randomised, parallel-group, placebo-controlled, double-blind, phase 1/2a study. Lancet HIV 2020, 7, e688–e698. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.X.; Zou, X.H.; Jiang, S.Y.; Lu, N.N.; Han, M.; Zhao, J.H.; Guo, X.J.; Zhao, S.C.; Lu, Z.Z. Prevalence of serum neutralizing antibodies to adenovirus type 5 (Ad5) and 41 (Ad41) in children is associated with age and sanitary conditions. Vaccine 2016, 34, 5579–5586. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zhou, Y.; Wu, H.; Wang, Z.; Zhan, Y.; Feng, X.; Geng, R.; Wu, Y.; Kong, W.; Yu, X. Seroprevalence of neutralizing antibodies to human adenovirus type 5 in healthy adults in China. J. Med. Virol. 2012, 84, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Huang, W.; Zhou, X.; Zhao, Q.; Wang, Q.; Jia, B. Seroprevalence of neutralizing antibodies to human adenoviruses type-5 and type-26 and chimpanzee adenovirus type-68 in healthy Chinese adults. J. Med. Virol. 2013, 85, 1077–1084. [Google Scholar] [CrossRef]
- Mennechet, F.J.D.; Paris, O.; Ouoba, A.R.; Salazar Arenas, S.; Sirima, S.B.; Takoudjou Dzomo, G.R.; Diarra, A.; Traore, I.T.; Kania, D.; Eichholz, K.; et al. A review of 65 years of human adenovirus seroprevalence. Expert. Rev. Vaccines 2019, 18, 597–613. [Google Scholar] [CrossRef]
- Sakurai, F.; Tachibana, M.; Mizuguchi, H. Adenovirus vector-based vaccine for infectious diseases. Drug Metab. Pharmacokinet. 2022, 42, 100432. [Google Scholar] [CrossRef]
- Tang, R.; Zheng, H.; Wang, B.S.; Gou, J.B.; Guo, X.L.; Chen, X.Q.; Chen, Y.; Wu, S.P.; Zhong, J.; Pan, H.X.; et al. Safety and immunogenicity of aerosolised Ad5-nCoV, intramuscular Ad5-nCoV, or inactivated COVID-19 vaccine CoronaVac given as the second booster following three doses of CoronaVac: A multicentre, open-label, phase 4, randomised trial. Lancet Respir. Med. 2023, 11, 613–623. [Google Scholar] [CrossRef]
- Wu, Y.F.; Wei, M.W.; Wang, R.J.; Guo, X.L.; Pan, H.X.; Gao, Y.C.; Li, X.L.; Wang, X.; Ma, X.M.; Wan, P.; et al. Immunogenicity consistency and safety with different production scales of recombinant adenovirus type-5 vectored COVID-19 vaccine in healthy adults: A randomized, double-blinded, immunobridging trial. Expert Rev. Vaccines 2023, 22, 662–670. [Google Scholar] [CrossRef]
- Jin, P.F.; Guo, X.L.; Gou, J.B.; Hou, L.H.; Song, Z.Z.; Zhu, T.; Pan, H.X.; Zhu, J.H.; Shi, F.J.; Du, P.; et al. Immunogenicity and safety of heterologous immunisation with Ad5-nCOV in healthy adults aged 60 years and older primed with an inactivated SARS-CoV-2 vaccine (CoronaVac): A phase 4, randomised, observer-blind, non-inferiority trial. Lancet Reg. Health West. Pac. 2023, 38, 100829. [Google Scholar] [CrossRef]
- Li, J.X.; Wu, S.P.; Guo, X.L.; Tang, R.; Huang, B.Y.; Chen, X.Q.; Chen, Y.; Hou, L.H.; Liu, J.X.; Zhong, J.; et al. Safety and immunogenicity of heterologous boost immunisation with an orally administered aerosolised Ad5-nCoV after two-dose priming with an inactivated SARS-CoV-2 vaccine in Chinese adults: A randomised, open-label, single-centre trial. Lancet Respir. Med. 2022, 10, 739–748. [Google Scholar] [CrossRef]
- Huang, T.; Zhang, S.; Dai, D.F.; Wang, B.S.; Zhuang, L.; Huang, H.T.; Wang, Z.F.; Zhao, J.S.; Li, Q.P.; Wu, S.P.; et al. Safety and immunogenicity of heterologous boosting with orally aerosolised or intramuscular Ad5-nCoV vaccine and homologous boosting with inactivated vaccines (BBIBP-CorV or CoronaVac) in children and adolescents: A randomised, open-label, parallel-controlled, non-inferiority, single-centre study. Lancet Respir. Med. 2023, 11, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Sprangers, M.C.; Lakhai, W.; Koudstaal, W.; Verhoeven, M.; Koel, B.F.; Vogels, R.; Goudsmit, J.; Havenga, M.J.; Kostense, S. Quantifying adenovirus-neutralizing antibodies by luciferase transgene detection: Addressing preexisting immunity to vaccine and gene therapy vectors. J. Clin. Microbiol. 2003, 41, 5046–5052. [Google Scholar] [CrossRef] [PubMed]
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-existing immunity against Ad vectors: Humoral, cellular, and innate response, what’s important? Hum. Vaccines Immunother. 2014, 10, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Singh, J.; Shaman, H.; Singh, B.; Anantharaj, A.; Sharanabasava, P.; Pandey, R.; Lodha, R.; Pandey, A.K.; Medigeshi, G.R. Pre-existing antibody levels negatively correlate with antibody titers after a single dose of BBV152 vaccination. Nat. Commun. 2022, 13, 3451. [Google Scholar] [CrossRef]
- Venkatraman, N.; Ndiaye, B.P.; Bowyer, G.; Wade, D.; Sridhar, S.; Wright, D.; Powlson, J.; Ndiaye, I.; Dieye, S.; Thompson, C.; et al. Safety and Immunogenicity of a Heterologous Prime-Boost Ebola Virus Vaccine Regimen in Healthy Adults in the United Kingdom and Senegal. J. Infect. Dis. 2019, 219, 1187–1197. [Google Scholar] [CrossRef]
- Shukarev, G.; Callendret, B.; Luhn, K.; Douoguih, M.; EBOVAC1 consortium. A two-dose heterologous prime-boost vaccine regimen eliciting sustained immune responses to Ebola Zaire could support a preventive strategy for future outbreaks. Hum. Vaccines Immunother. 2017, 13, 266–270. [Google Scholar] [CrossRef]
- Voysey, M.; Costa Clemens, S.A.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: A pooled analysis of four randomised trials. Lancet 2021, 397, 881–891. [Google Scholar] [CrossRef]
- Jeyanathan, M.; Fritz, D.K.; Afkhami, S.; Aguirre, E.; Howie, K.J.; Zganiacz, A.; Dvorkin-Gheva, A.; Thompson, M.R.; Silver, R.F.; Cusack, R.P.; et al. Aerosol delivery, but not intramuscular injection, of adenovirus-vectored tuberculosis vaccine induces respiratory-mucosal immunity in humans. JCI Insight 2022, 7, e155655. [Google Scholar] [CrossRef]
- Afkhami, S.; D’Agostino, M.R.; Zhang, A.; Stacey, H.D.; Marzok, A.; Kang, A.; Singh, R.; Bavananthasivam, J.; Ye, G.; Luo, X.; et al. Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell 2022, 185, 896–915.e19. [Google Scholar] [CrossRef]
- Lavelle, E.C.; Ward, R.W. Mucosal vaccines—Fortifying the frontiers. Nat. Rev. Immunol. 2022, 22, 236–250. [Google Scholar] [CrossRef]
- Xu, F.; Wu, S.; Yi, L.; Peng, S.; Wang, F.; Si, W.; Hou, L.; Zhu, T. Safety, mucosal and systemic immunopotency of an aerosolized adenovirus-vectored vaccine against SARS-CoV-2 in rhesus macaques. Emerg. Microbes Infect. 2022, 11, 438–441. [Google Scholar] [CrossRef]
- Jeyanathan, V.; Afkhami, S.; D’Agostino, M.R.; Zganiacz, A.; Feng, X.; Miller, M.S.; Jeyanathan, M.; Thompson, M.R.; Xing, Z. Differential Biodistribution of Adenoviral-Vectored Vaccine Following Intranasal and Endotracheal Deliveries Leads to Different Immune Outcomes. Front. Immunol. 2022, 13, 860399. [Google Scholar] [CrossRef]

{kind=link}
| Immunization Schedule [Dosage] | 6–17 Years a | 18 Years or Older a | |
|---|---|---|---|
| Ad5-IM-prime | Ad5-IM [3.0 × 1010 vp] | NCT04916886 | / |
| Ad5-IM [5.0 × 1010 vp] | / | NCT04313127 NCT04341389 NCT04916886 | |
| Ad5-IM [1.0 × 1011 vp] | / | NCT04313127 NCT04341389 | |
| Ad5-IM [1.5 × 1011 vp] | / | NCT04313127 | |
| Ad5-IM-boost | ICV × 2 + Ad5-IM [3.0 × 1010 vp] | NCT05330871 | / |
| ICV × 1 + Ad5-IM [5.0 × 1010 vp] | / | NCT04892459 NCT04952727 | |
| ICV × 2 + Ad5-IM [5.0 × 1010 vp] | / | NCT04892459 NCT04952727 | |
| ICV × 3 + Ad5-IM [5.0 × 1010 vp] | / | NCT05303584 | |
| Ad5-IH-boost | ICV × 2 + Ad5-IH [1.0 × 1010 vp] | NCT05330871 | NCT05043259 |
| ICV × 2 + Ad5-IH [2.0 × 1010 vp] | / | NCT05043259 | |
| ICV × 3 + Ad5-IH [1.0 × 1010 vp] | / | NCT05303584 | |
| Ad5-IM-Prime | Ad5-IM-Boost | Ad5-IH-Boost | p Value | |
|---|---|---|---|---|
| Adolescent cohort, ages 6–17 years (N = 1273) | ||||
| Participants, n | 1001 | 65 | 207 | / |
| Median age (IQR) | 12.0 (10.0, 14.0) | 13.0 (9.0, 14.0) | 12.0 (10.0, 14.0) | 0.6141 |
| Sex (%) | ||||
| Male | 542 (54.1%) | 31 (47.7%) | 103 (49.8%) | 0.3446 |
| Female | 459 (45.9%) | 34 (52.3%) | 104 (50.2%) | |
| Pre-existing Ad5 neutralising antibodies | ||||
| Geometric mean titers (95% CI) | 118.3 (101.7, 137.6) | 73.0 (37.1, 143.7) | 116.4 (81.5, 166.0) | 0.3146 |
| Participants with titers < 1:12 (%) | 336 (33.6%) | 32 (49.2%) | 76 (36.7%) | 0.0008 |
| 1:12 ≤ Participants with titers ≤ 1:200 (%) | 114 (11.4%) | 7 (10.8%) | 21 (10.1%) | |
| 1:200 < Participants with titers ≤ 1:1000 (%) | 425 (42.4%) | 13 (20.0%) | 69 (33.3%) | |
| Participants with titers > 1:1000 (%) | 126 (12.6%) | 13 (20.0%) | 41 (19.8%) | |
| Adult cohort, ages 18 years and older (N = 2239) | ||||
| Participants, n | 1450 | 396 | 393 | / |
| Median age (IQR) | 42.0 (33.0, 50.0) | 53.0 (44.0, 65.0) | 43.0 (35.0, 50.0) | <0.0001 |
| Sex (%) | ||||
| Male | 749 (51.7%) | 215 (54.3%) | 171 (43.5%) | 0.0048 |
| Female | 701 (48.3%) | 181 (45.7%) | 222 (56.5%) | |
| Pre-existing Ad5 neutralising antibodies | ||||
| Geometric mean titers (95% CI) | 146.5 (131.5, 163.3) | 165.7 (134.8, 203.8) | 236.8 (189.9, 295.1) | 0.0004 |
| Participants with titers < 1:12 (%) | 297 (20.5%) | 66 (16.7%) | 59 (15.0%) | 0.0026 |
| 1:12 ≤ Participants with titers ≤ 1:200 (%) | 364 (25.1%) | 109 (27.5%) | 94 (23.9%) | |
| 1:200 < Participants with titers ≤ 1:1000 (%) | 593 (40.9%) | 175 (44.2%) | 160 (40.7%) | |
| Participants with titers > 1:1000 (%) | 200 (13.8%) | 46 (11.6%) | 80 (20.4%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Li, Y.; Li, X.; Wang, F.; Qi, R.; Zhu, T.; Li, J. Pooled Analysis of the Effect of Pre-Existing Ad5 Neutralizing Antibodies on the Immunogenicity of Adenovirus Type 5 Vector-Based COVID-19 Vaccine from Eight Clinical Trials. Vaccines 2025, 13, 333. https://doi.org/10.3390/vaccines13030333
Liu W, Li Y, Li X, Wang F, Qi R, Zhu T, Li J. Pooled Analysis of the Effect of Pre-Existing Ad5 Neutralizing Antibodies on the Immunogenicity of Adenovirus Type 5 Vector-Based COVID-19 Vaccine from Eight Clinical Trials. Vaccines. 2025; 13(3):333. https://doi.org/10.3390/vaccines13030333
Chicago/Turabian StyleLiu, Wenqing, Yuqing Li, Xiaolong Li, Feiyu Wang, Runjie Qi, Tao Zhu, and Jingxin Li. 2025. "Pooled Analysis of the Effect of Pre-Existing Ad5 Neutralizing Antibodies on the Immunogenicity of Adenovirus Type 5 Vector-Based COVID-19 Vaccine from Eight Clinical Trials" Vaccines 13, no. 3: 333. https://doi.org/10.3390/vaccines13030333
APA StyleLiu, W., Li, Y., Li, X., Wang, F., Qi, R., Zhu, T., & Li, J. (2025). Pooled Analysis of the Effect of Pre-Existing Ad5 Neutralizing Antibodies on the Immunogenicity of Adenovirus Type 5 Vector-Based COVID-19 Vaccine from Eight Clinical Trials. Vaccines, 13(3), 333. https://doi.org/10.3390/vaccines13030333