
Bacterium-like Particles from Corynebacterium pseudodiphtheriticum as Mucosal Adjuvant for the Development of Pneumococcal Vaccines

 , , , ,
, , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. C. pseudodiphteriticum Culture and Obtention of BLPs
2.2. Animals, Immunizations, and Pneumococcal Infections
2.3. Antibody Quantification
2.4. Infection Challenge Experiments and Pneumococcal Cell Counts
2.5. Lung Tissue Injury Markers
2.6. Cytokine Concentrations
2.7. Statistical Analysis
3. Results
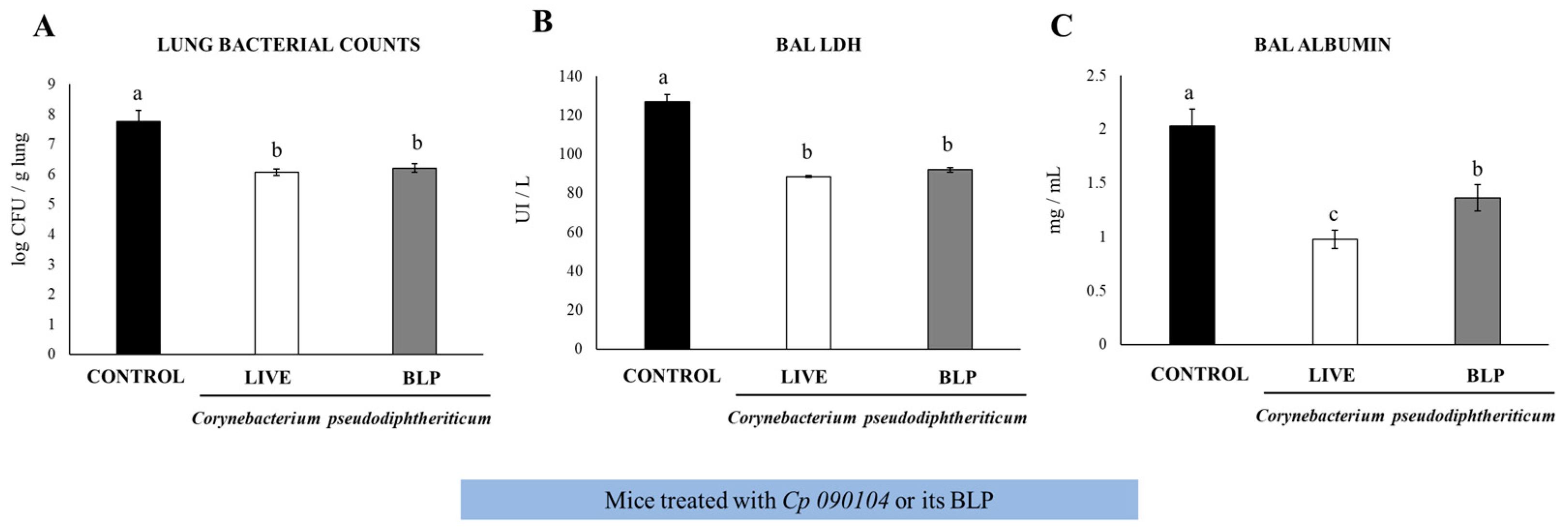
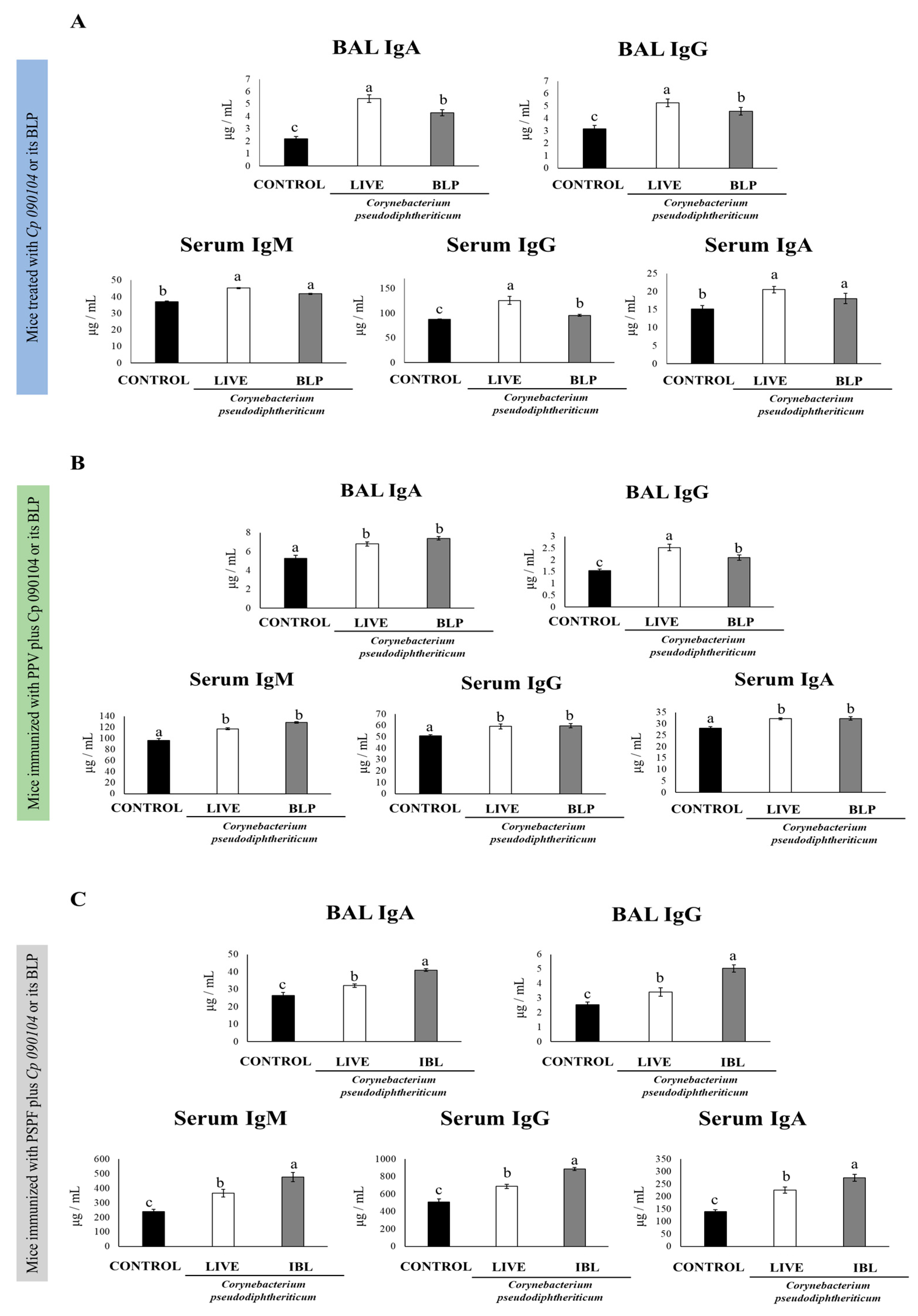
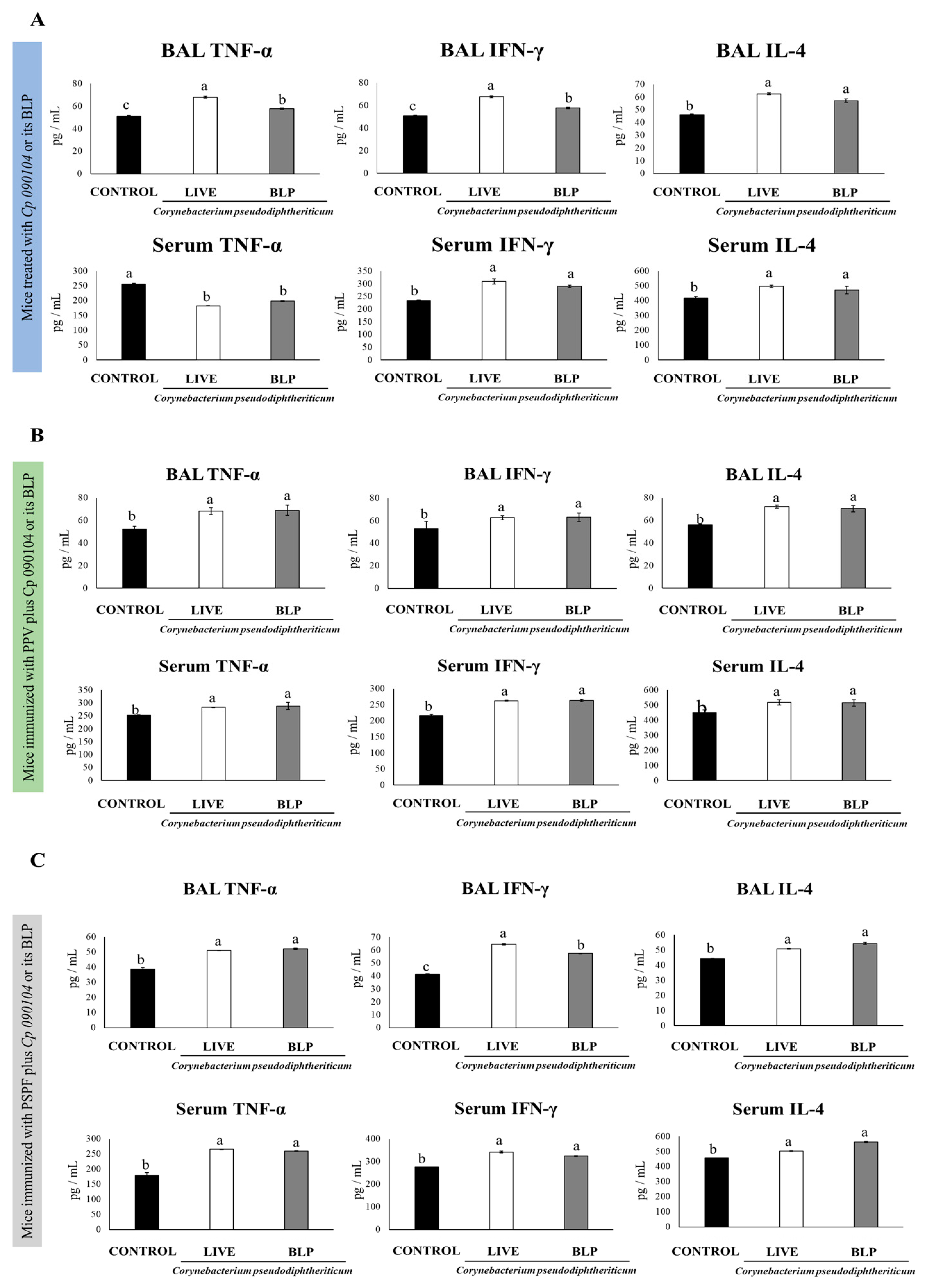
3.1. Cp 090104 and CP-Derived BLPs Improve the Adaptive Immune Response against S. pneumoniae in Infant Mice
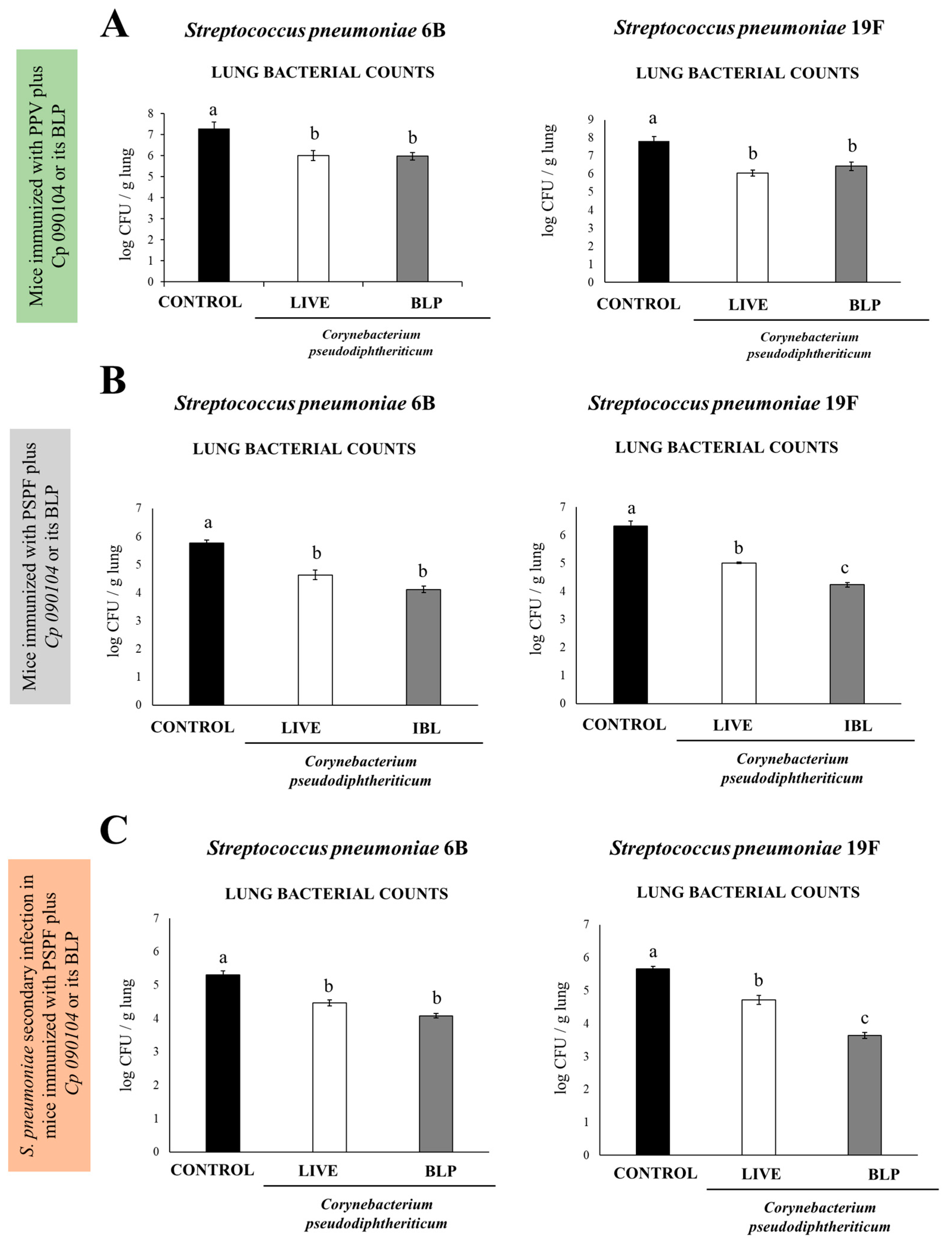
3.2. Cp 090104 and CP-Derived BLPs Improve the Adaptive Immune Response Induced by a Commercial Pneumococcal Polysaccharide Vaccine
3.3. C. pseudodiphteriticum 090104 and CP-Derived BLPs Improve the Adaptive Immune Response Induced by the Experimental Chimeric Recombinant Protein Vaccine PSPF
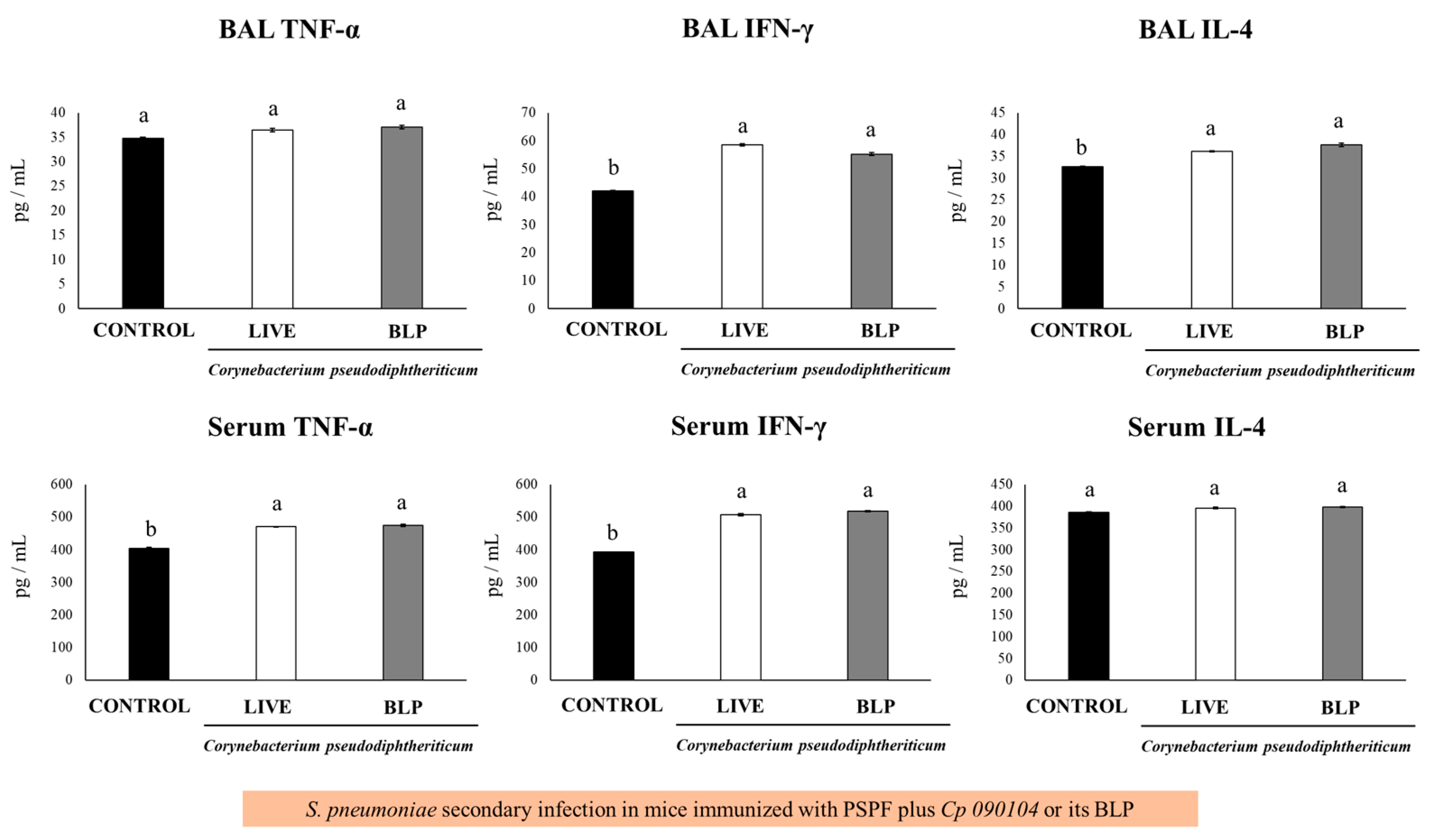
3.4. Immunization with PSPF and C. pseudodiphteriticum 090104 or CP-Derived BLPs Improves Resistance to Secondary Pneumococcal Infection Induced after Viral Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrejko, K.; Ratnasiri, B.; Hausdorff, W.P.; Laxminarayan, R.; Lewnard, J.A. Antimicrobial Resistance in Paediatric Streptococcus pneumoniae Isolates amid Global Implementation of Pneumococcal Conjugate Vaccines: A Systematic Review and Meta-Regression Analysis. Lancet Microbe 2021, 2, e450–e460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, X.; Guo, M.; Chen, X.; Li, B.; Yu, J.; Gu, T.; Kong, W.; Wu, Y. Combined Prime-Boost Immunization with Systemic and Mucosal Pneumococcal Vaccines Based on Pneumococcal Surface Protein A to Enhance Protection against Lethal Pneumococcal Infections. Immunol. Res. 2019, 67, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Goulart, C.; Rodriguez, D.; Kanno, A.I.; Silva, J.L.S.C.; Leite, L.C.C. Early Pneumococcal Clearance in Mice Induced by Systemic Immunization with Recombinant BCG PspA-PdT Prime and Protein Boost Correlates with Cellular and Humoral Immune Response in Bronchoalveolar Fluids (BALF). Vaccine X 2020, 4, 100049. [Google Scholar] [CrossRef] [PubMed]
- Principi, N.; Esposito, S. Development of Pneumococcal Vaccines over the Last 10 Years. Expert Opin. Biol. Ther. 2018, 18, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Laiño, J.; Villena, J.; Suvorov, A.; Zelaya, H.; Ortiz Moyano, R.; Salva, S.; Alvarez, S. Nasal Immunization with Recombinant Chimeric Pneumococcal Protein and Cell Wall from Immunobiotic Bacteria Improve Resistance of Infant Mice to Streptococcus pneumoniae Infection. PLoS ONE 2018, 13, e0206661. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Guo, J.; Wang, D.; Yu, J.; Gu, T.; Jiang, C.; Kong, W.; Wu, Y. Broad protective immune responses elicited by bacterium-like particle-based intranasal pneumococcal particle vaccine displaying PspA2 and PspA4 fragments. Hum. Vaccin. Immunother. 2019, 15, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Uematsu, S. Development of Prime–Boost-Type next-Generation Mucosal Vaccines. Int. Immunol. 2020, 32, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Goulart, C.; Rodriguez, D.; Kanno, A.I.; Converso, T.R.; Lu, Y.-J.; Malley, R.; Leite, L.C.C. A Combination of Recombinant Mycobacterium Bovis BCG Strains Expressing Pneumococcal Proteins Induces Cellular and Humoral Immune Responses and Protects against Pneumococcal Colonization and Sepsis. Clin. Vaccine Immunol. 2017, 24, e00133-17. [Google Scholar] [CrossRef]
- de Steenhuijsen Piters, W.A.A.; Binkowska, J.; Bogaert, D. Early Life Microbiota and Respiratory Tract Infections. Cell Host Microbe 2020, 28, 223–232. [Google Scholar] [CrossRef]
- Kanmani, P.; Clua, P.; Vizoso-Pinto, M.G.; Rodriguez, C.; Alvarez, S.; Melnikov, V.; Takahashi, H.; Kitazawa, H.; Villena, J. Respiratory Commensal Bacteria Corynebacterium pseudodiphtheriticum Improves Resistance of Infant Mice to Respiratory Syncytial Virus and Streptococcus pneumoniae Superinfection. Front. Microbiol. 2017, 8, 1613. [Google Scholar] [CrossRef]
- Ortiz Moyano, R.; Raya Tonetti, F.; Tomokiyo, M.; Kanmani, P.; Vizoso-Pinto, M.G.; Kim, H.; Quilodrán-Vega, S.; Melnikov, V.; Alvarez, S.; Takahashi, H.; et al. The Ability of Respiratory Commensal Bacteria to Beneficially Modulate the Lung Innate Immune Response Is a Strain Dependent Characteristic. Microorganisms 2020, 8, 727. [Google Scholar] [CrossRef]
- Dentice Maidana, S.; Ortiz Moyano, R.; Vargas, J.M.; Fukuyama, K.; Kurata, S.; Melnikov, V.; Jure, M.Á.; Kitazawa, H.; Villena, J. Respiratory Commensal Bacteria Increase Protection against Hypermucoviscous Carbapenem-Resistant Klebsiella Pneumoniae ST25 Infection. Pathogens 2022, 11, 1063. [Google Scholar] [CrossRef]
- Ortiz Moyano, R.; Raya Tonetti, F.; Fukuyama, K.; Elean, M.; Tomokiyo, M.; Suda, Y.; Melnikov, V.; Kitazawa, H.; Villena, J. The Respiratory Commensal Bacterium Corynebacterium pseudodiphtheriticum as a Mucosal Adjuvant for Nasal Vaccines. Vaccines 2023, 11, 611. [Google Scholar] [CrossRef]
- Twigg, H.L. Humoral Immune Defense (Antibodies): Recent Advances. Proc. Am. Thorac. Soc. 2005, 2, 417–421. [Google Scholar] [CrossRef]
- Saeland, E.; Vidarsson, G.; Leusen, J.H.W.; van Garderen, E.; Nahm, M.H.; Vile-Weekhout, H.; Walraven, V.; Stemerding, A.M.; Verbeek, J.S.; Rijkers, G.T.; et al. Central Role of Complement in Passive Protection by Human IgG1 and IgG2 Anti-Pneumococcal Antibodies in Mice1. J. Immunol. 2003, 170, 6158–6164. [Google Scholar] [CrossRef]
- Gingerich, A.D.; Royer, F.; McCormick, A.L.; Scasny, A.; Vidal, J.E.; Mousa, J.J. Synergistic Protection against Secondary Pneumococcal Infection by Human Monoclonal Antibodies Targeting Distinct Epitopes. J. Immunol. 2023, 210, 50–60. [Google Scholar] [CrossRef]
- Janoff, E.N.; Fasching, C.; Orenstein, J.M.; Rubins, J.B.; Opstad, N.L.; Dalmasso, A.P. Killing of Streptococcus pneumoniae by Capsular Polysaccharide–Specific Polymeric IgA, Complement, and Phagocytes. J. Clin. Investg. 1999, 104, 1139–1147. [Google Scholar] [CrossRef]
- Fukuyama, Y.; King, J.D.; Kataoka, K.; Kobayashi, R.; Gilbert, R.S.; Oishi, K.; Hollingshead, S.K.; Briles, D.E.; Fujihashi, K. Secretory-IgA Antibodies Play an Important Role in the Immunity to Streptococcus pneumoniae. J. Immunol. 2010, 185, 1755–1762. [Google Scholar] [CrossRef]
- Sun, K.; Johansen, F.-E.; Eckmann, L.; Metzger, D.W. An Important Role for Polymeric Ig Receptor-Mediated Transport of IgA in Protection against Streptococcus pneumoniae Nasopharyngeal Carriage1. J. Immunol. 2004, 173, 4576–4581. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, T.; Akahori, Y.; Toyama, M.; Miyamura, N.; Ishii, K.; Saijo, S.; Iwakura, Y.; Kinjo, Y.; Miyazaki, Y.; Oishi, K.; et al. Dectin-2-Dependent NKT Cell Activation and Serotype-Specific Antibody Production in Mice Immunized with Pneumococcal Polysaccharide Vaccine. PLoS ONE 2013, 8, e78611. [Google Scholar] [CrossRef] [PubMed]
- La Pine, T.R.; Joyner, J.L.; Augustine, N.H.; Kwak, S.D.; Hill, H.R. Defective production of IL-18 and IL-12 by cord blood mononuclear cells influences the T helper-1 interferon gamma response to group B Streptococci. Pediatr. Res. 2003, 54, 276–281. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hu, Y.; Li, L.; Xu, W.; Wu, K.; Xiao, J.; Peng, Y.; Liu, Y.; Yin, Y.; Zhang, X. IL-4 Plays an Essential Role in DnaJ-ΔA146Ply-Mediated Immunoprotection against Streptococcus pneumoniae in Mice. Mol. Immunol. 2022, 143, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Van Der Poll, T.; Keogh, C.V.; Buurman, W.A.; Lowry, S.F. Passive Immunization against Tumor Necrosis Factor-Alpha Impairs Host Defense during Pneumococcal Pneumonia in Mice. Am. J. Respir. Crit. Care Med. 1997, 155, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Calbo, E.; Alsina, M.; Rodríguez-Carballeira, M.; Lite, J.; Garau, J. The Impact of Time on the Systemic Inflammatory Response in Pneumococcal Pneumonia. Eur. Respir. J. 2010, 35, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Puchta, A.; Naidoo, A.; Verschoor, C.P.; Loukov, D.; Thevaranjan, N.; Mandur, T.S.; Nguyen, P.; Jordana, M.; Loeb, M.; Xing, Z.; et al. TNF Drives Monocyte Dysfunction with Age and Results in Impaired Anti-Pneumococcal Immunity. PLOS Pathog. 2016, 12, e1005368. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, Y.; Ouellet, N.; Deslauriers, A.-M.; Simard, M.; Olivier, M.; Bergeron, M.G. Cytokine Kinetics and Other Host Factors in Response to Pneumococcal Pulmonary Infection in Mice. Infect. Immun. 1998, 66, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.B.; Malamba, R.; Mthunthama, N.; Jarman, E.R.; Jambo, K.; Jere, K.; Zijlstra, E.E.; Molyneux, M.E.; Dennis, J.; French, N. Inhaled Delivery of 23-Valent Pneumococcal Polysaccharide Vaccine Does Not Result in Enhanced Pulmonary Mucosal Immunoglobulin Responses. Vaccine 2008, 26, 5400–5406. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, N.; Zhang, F.; Zhou, H.; Ma, W.; Mao, H.; Wang, M.; Ke, Z.; Wang, J.; Qi, L. Mechanisms of Immune-Related Long Non-Coding RNAs in Spleens of Mice Vaccinated with 23-Valent Pneumococcal Polysaccharide Vaccine (PPV23). Vaccines 2023, 11, 529. [Google Scholar] [CrossRef] [PubMed]
- Puech, V.; Chami, M.; Lemassu, A.; Lanéelle, M.-A.; Schiffler, B.; Gounon, P.; Bayan, N.; Benz, R.; Daffé, M. Structure of the Cell Envelope of Corynebacteria: Importance of the Non-Covalently Bound Lipids in the Formation of the Cell Wall Permeability Barrier and Fracture Plane. Microbiology 2001, 147, 1365–1382. [Google Scholar] [CrossRef]
- Mokrozub, V.V.; Lazarenko, L.M.; Sichel, L.M.; Babenko, L.P.; Lytvyn, P.M.; Demchenko, O.M.; Melnichenko, Y.O.; Boyko, N.V.; Biavati, B.; DiGioia, D.; et al. The Role of Beneficial Bacteria Wall Elasticity in Regulating Innate Immune Response. EPMA J. 2015, 6, 13. [Google Scholar] [CrossRef]
- Wolf, A.J.; Underhill, D.M. Peptidoglycan Recognition by the Innate Immune System. Nat. Rev. Immunol. 2018, 18, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.T.; Kim, S.Y.; Kim, M.S.; Lee, S.E.; Rhee, J.H. Intranasal immunization with recombinant PspA fused with a flagellin enhances cross-protective immunity against Streptococcus pneumoniae infection in mice. Vaccine 2011, 29, 5731–5739. [Google Scholar] [CrossRef]
- Cuadros, C.; Lopez-Hernandez, F.J.; Dominguez, A.L.; McClelland, M.; Lustgarten, J. Flagellin fusion proteins as adjuvants or vaccines induce specific immune responses. Infect. Immun. 2004, 72, 2810–2816. [Google Scholar] [CrossRef]
- Liu, L.; Johnson, H.L.; Cousens, S.; Perin, J.; Scott, S.; Lawn, J.E.; Rudan, I.; Campbell, H.; Cibulskis, R.; Li, M.; et al. Global, Regional, and National Causes of Child Mortality: An Updated Systematic Analysis for 2010 with Time Trends since 2000. Lancet 2012, 379, 2151–2161. [Google Scholar] [CrossRef]
- Bosch, A.A.T.M.; Biesbroek, G.; Trzcinski, K.; Sanders, E.A.M.; Bogaert, D. Viral and Bacterial Interactions in the Upper Respiratory Tract. PLOS Pathog. 2013, 9, e1003057. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Price, L.B.; Hungate, B.A.; Abraham, A.G.; Larsen, L.A.; Christensen, K.; Stegger, M.; Skov, R.; Andersen, P.S. Staphylococcus Aureus and the Ecology of the Nasal Microbiome. Sci. Adv. 2015, 1, e1400216. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, D.M.; Givon-Lavi, N.; Shemer-Avni, Y.; Bar-Ziv, J.; Alonso, W.J.; Greenberg, D.; Dagan, R. Influence of Pneumococcal Vaccines and Respiratory Syncytial Virus on Alveolar Pneumonia, Israel. Emerg. Infect. Dis. 2013, 19, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Cebey-López, M.; Pardo-Seco, J.; Gómez-Carballa, A.; Martinón-Torres, N.; Martinón-Sánchez, J.M.; Justicia-Grande, A.; Rivero-Calle, I.; Pinnock, E.; Salas, A.; Fink, C.; et al. Bacteremia in Children Hospitalized with Respiratory Syncytial Virus Infection. PLoS ONE 2016, 11, e0146599. [Google Scholar] [CrossRef] [PubMed]
- Hament, J.-M.; Aerts, P.C.; Fleer, A.; van Dijk, H.; Harmsen, T.; Kimpen, J.L.L.; Wolfs, T.F.W. Direct Binding of Respiratory Syncytial Virus to Pneumococci: A Phenomenon That Enhances Both Pneumococcal Adherence to Human Epithelial Cells and Pneumococcal Invasiveness in a Murine Model. Pediatr. Res. 2005, 58, 1198–1203. [Google Scholar] [CrossRef]
- Smith, C.M.; Sandrini, S.; Datta, S.; Freestone, P.; Shafeeq, S.; Radhakrishnan, P.; Williams, G.; Glenn, S.M.; Kuipers, O.P.; Hirst, R.A.; et al. Respiratory Syncytial Virus Increases the Virulence of Streptococcus pneumoniae by Binding to Penicillin Binding Protein 1a. A New Paradigm in Respiratory Infection. Am. J. Respir. Crit. Care Med. 2014, 190, 196–207. [Google Scholar] [CrossRef]
- Madhi, S.A.; Klugman, K.P.; The Vaccine Trialist Group. A Role for Streptococcus pneumoniae in Virus-Associated Pneumonia. Nat. Med. 2004, 10, 811–813. [Google Scholar] [CrossRef]
- Khan, M.N.; Xu, Q.; Pichichero, M.E. Protection against Streptococcus pneumoniae Invasive Pathogenesis by a Protein-Based Vaccine Is Achieved by Suppression of Nasopharyngeal Bacterial Density during Influenza A Virus Coinfection. Infect. Immun. 2017, 85, e00530-16. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz Moyano, R.; Raya Tonetti, F.; Elean, M.; Imamura, Y.; Fukuyama, K.; Suda, Y.; Melnikov, V.; Suvorov, A.; Vizoso-Pinto, M.G.; Kitazawa, H.; et al. Bacterium-like Particles from Corynebacterium pseudodiphtheriticum as Mucosal Adjuvant for the Development of Pneumococcal Vaccines. Vaccines 2024, 12, 412. https://doi.org/10.3390/vaccines12040412
Ortiz Moyano R, Raya Tonetti F, Elean M, Imamura Y, Fukuyama K, Suda Y, Melnikov V, Suvorov A, Vizoso-Pinto MG, Kitazawa H, et al. Bacterium-like Particles from Corynebacterium pseudodiphtheriticum as Mucosal Adjuvant for the Development of Pneumococcal Vaccines. Vaccines. 2024; 12(4):412. https://doi.org/10.3390/vaccines12040412
Chicago/Turabian StyleOrtiz Moyano, Ramiro, Fernanda Raya Tonetti, Mariano Elean, Yoshiya Imamura, Kohtaro Fukuyama, Yoshihito Suda, Vyacheslav Melnikov, Alexander Suvorov, María Guadalupe Vizoso-Pinto, Haruki Kitazawa, and et al. 2024. "Bacterium-like Particles from Corynebacterium pseudodiphtheriticum as Mucosal Adjuvant for the Development of Pneumococcal Vaccines" Vaccines 12, no. 4: 412. https://doi.org/10.3390/vaccines12040412
APA StyleOrtiz Moyano, R., Raya Tonetti, F., Elean, M., Imamura, Y., Fukuyama, K., Suda, Y., Melnikov, V., Suvorov, A., Vizoso-Pinto, M. G., Kitazawa, H., & Villena, J. (2024). Bacterium-like Particles from Corynebacterium pseudodiphtheriticum as Mucosal Adjuvant for the Development of Pneumococcal Vaccines. Vaccines, 12(4), 412. https://doi.org/10.3390/vaccines12040412