
Comparison of the Immune Effects of an mRNA Vaccine and a Subunit Vaccine against Herpes Zoster Administered by Different Injection Methods


Abstract
1. Introduction
2. Materials and Methods
2.1. Vaccine Preparation
2.2. Mouse Studies
2.3. gE-Specific IgG Titer Detection
2.4. Cytokine Analysis
2.5. Enzyme-Linked Immunospot (ELISPOT) Assay
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
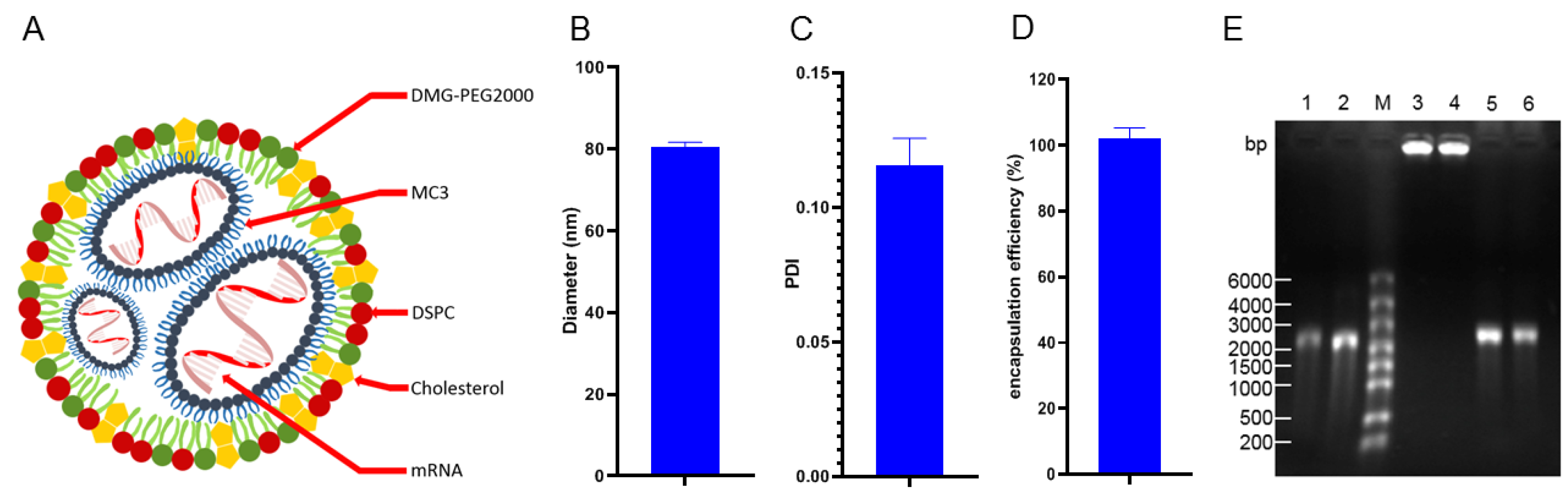
3.1. Almost All gE mRNA Was Coated by LNPs to Form Particles with a Uniform Diameter
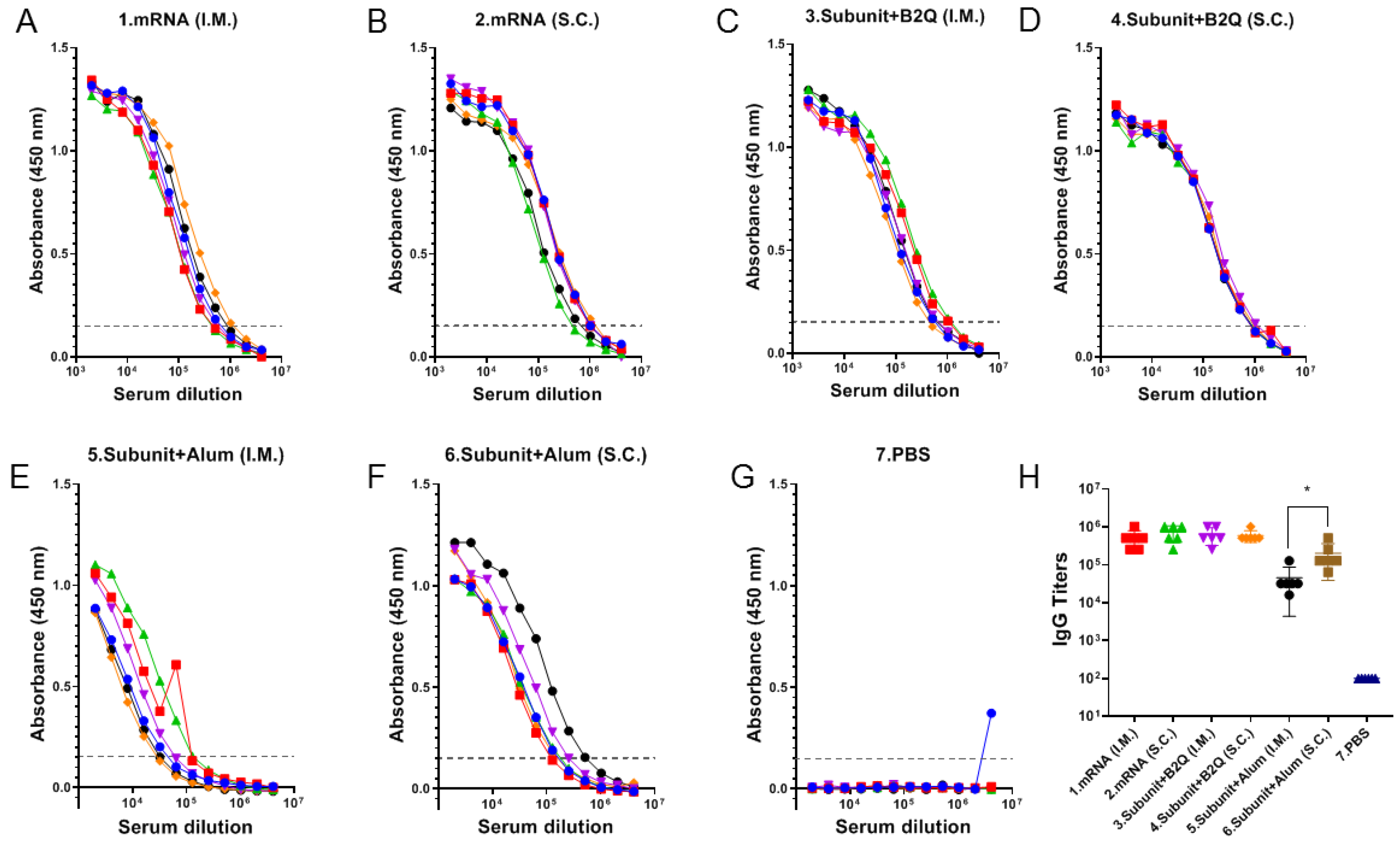
3.2. The Effect of the Injection Route on Humoral Immune Response Intensity Was Related to Vaccine Type and Adjuvant Composition
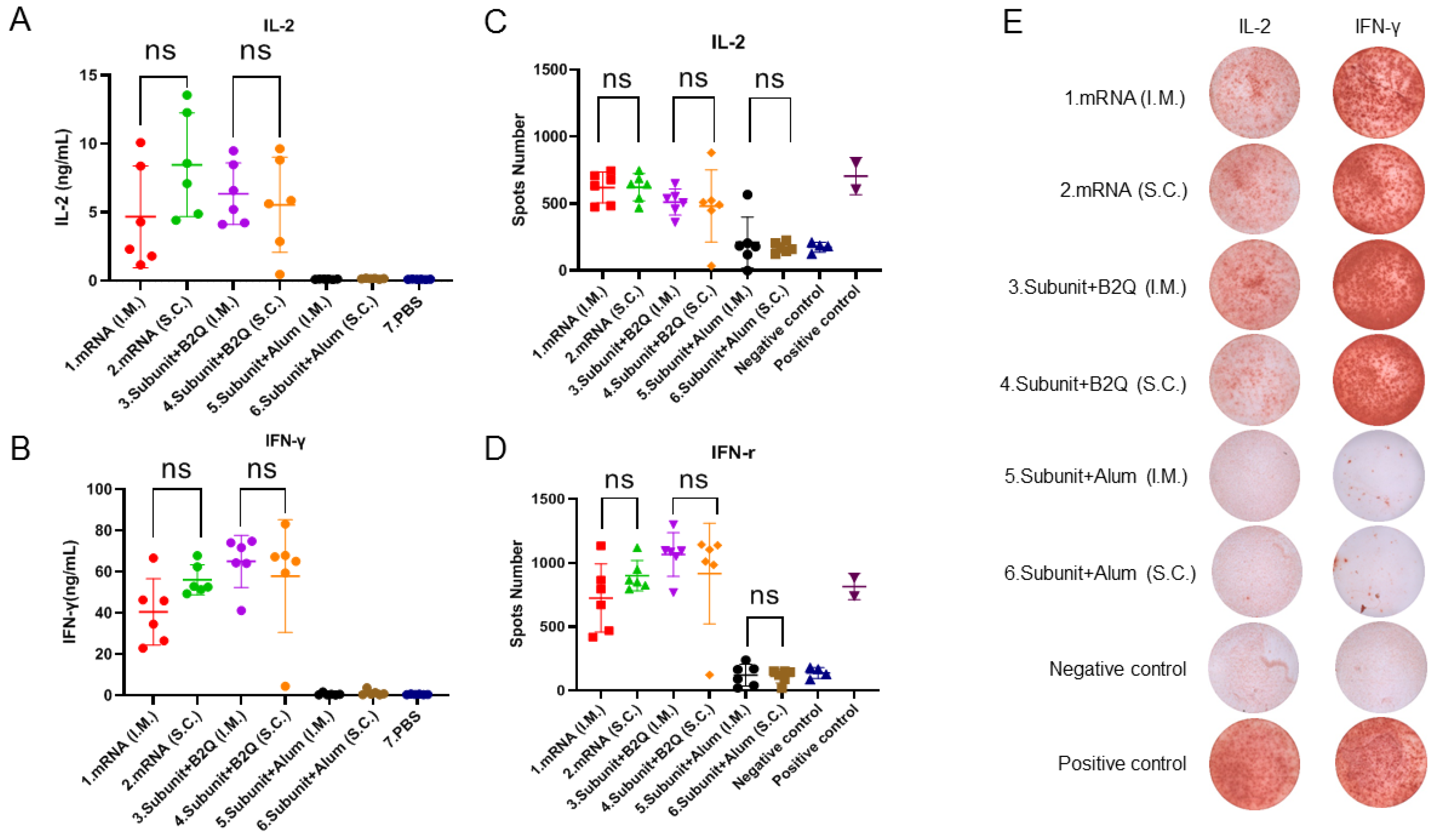
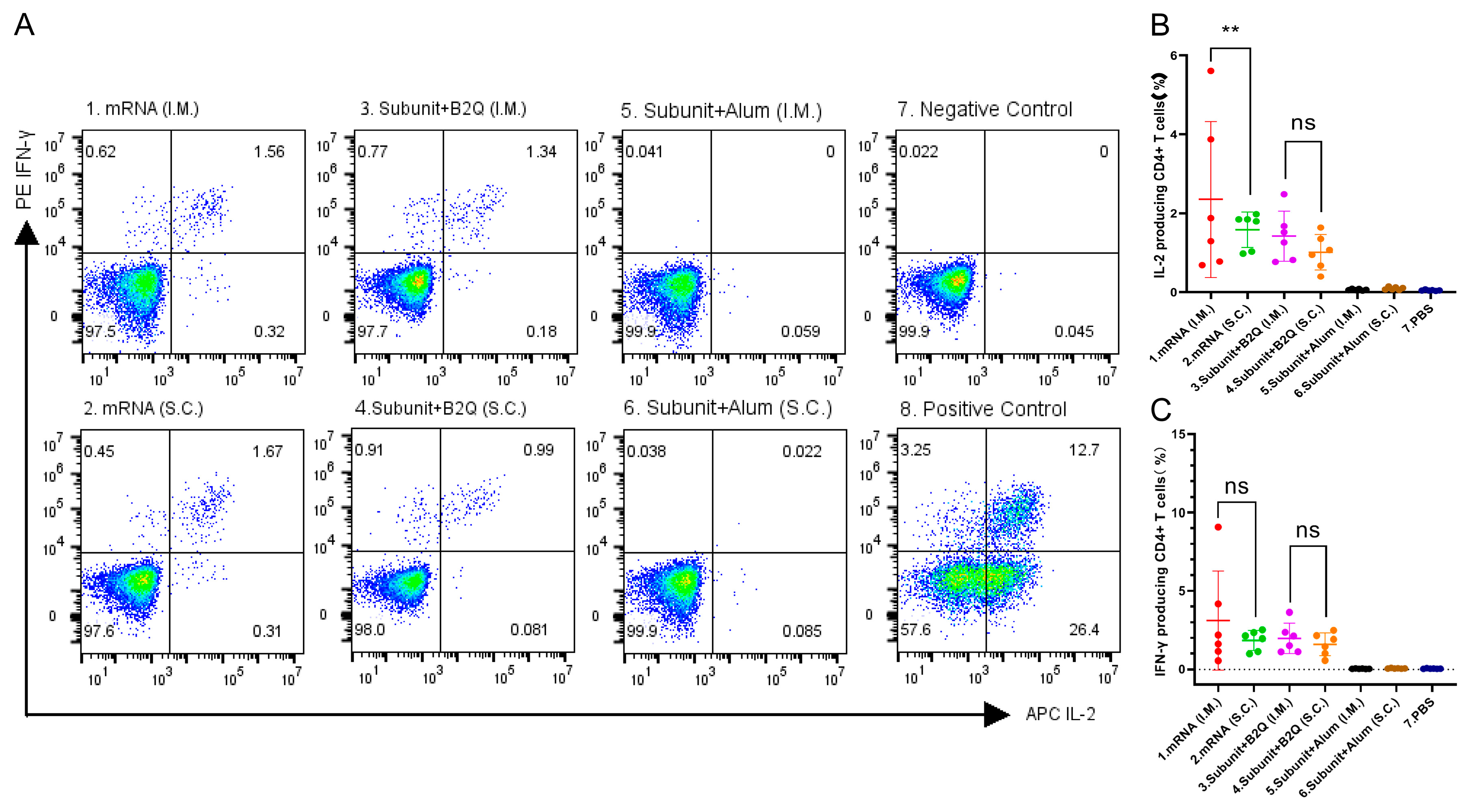
3.3. Cellular Immune Responses Induced by Intramuscular or Subcutaneous Injections of the mRNA or Subunit Vaccines Were Not Significantly Different
3.4. Different Types of Vaccines with the Same Injection Methods Can Induce an Immune Response of Comparable Intensity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zerboni, L.; Sen, N.; Oliver, S.L.; Arvin, A.M. Molecular mechanisms of varicella zoster virus pathogenesis. Nat. Rev. Microbiol. 2014, 12, 197–210. [Google Scholar] [CrossRef]
- Gao, Z.; Yang, F.; Qi, F.; Li, X.; Li, S. Evaluating the impact of universal varicella vaccination among preschool-aged children in Qingdao, China: An interrupted time-series analysis. Hum. Vaccin. Immunother. 2022, 18, 2094641. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.E. Selective decline in cellular immune response to varicella-zoster in the elderly. Neurology 1980, 30, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Bresnitz, E.; Popmihajlov, Z.; Weinberg, A.; Liaw, K.L.; Willis, E.; Curtis, J.R. Studies with herpes zoster vaccines in immune compromised patients. Expert. Rev. Vaccines 2017, 16, 1217–1230. [Google Scholar] [CrossRef] [PubMed]
- Gilden, D.H.; Vafai, A.; Shtram, Y.; Becker, Y.; Devlin, M.; Wellish, M. Varicella-zoster virus DNA in human sensory ganglia. Nature 1983, 306, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Esiri, M.M.; Tomlinson, A.H. Herpes Zoster. Demonstration of virus in trigeminal nerve and ganglion by immunofluorescence and electron microscopy. J. Neurol. Sci. 1972, 15, 35–48. [Google Scholar] [CrossRef]
- Weller, T.H. Varicella and herpes zoster. Changing concepts of the natural history, control, and importance of a not-so-benign virus. N. Engl. J. Med. 1983, 309, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, K.G. Antiviral therapy. Varicella-zoster virus infections, herpes labialis and mucocutaneous herpes, and cytomegalovirus infections. Lancet 1984, 2, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.; Oxman, M.N.; et al. Varicella zoster virus infection. Nat. Rev. Dis. Primers. 2015, 1, 15016. [Google Scholar] [CrossRef] [PubMed]
- Oxman, M.N.; Levin, M.J.; Johnson, G.R.; Schmader, K.E.; Straus, S.E.; Gelb, L.D.; Arbeit, R.D.; Simberkoff, M.S.; Gershon, A.A.; Davis, L.E.; et al. A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N. Engl. J. Med. 2005, 352, 2271–2284. [Google Scholar] [PubMed]
- Cunningham, A.L.; Lal, H.; Kovac, M.; Chlibek, R.; Hwang, S.J.; Diez-Domingo, J.; Godeaux, O.; Levin, M.J.; McElhaney, J.E.; Puig-Barbera, J.; et al. Efficacy of the Herpes Zoster Subunit Vaccine in Adults 70 Years of Age or Older. N. Engl. J. Med. 2016, 375, 1019–1032. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Steinberg, S.P. Antibody responses to varicella-zoster virus and the role of antibody in host defense. Am. J. Med. Sci. 1981, 282, 12–17. [Google Scholar] [CrossRef]
- Asada, H. VZV-specific cell-mediated immunity, but not humoral immunity, correlates inversely with the incidence of herpes zoster and the severity of skin symptoms and zoster-associated pain: The SHEZ study. Vaccine 2019, 37, 6776–6781. [Google Scholar] [CrossRef] [PubMed]
- Mo, C.; Schneeberger, E.E.; Arvin, A.M. Glycoprotein E of varicella-zoster virus enhances cell-cell contact in polarized epithelial cells. J. Virol. 2000, 74, 11377–11387. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.H.; Zhang, Q.B.; Liu, B.; Piao, X.H.; Yan, Y.L.; Hu, X.G.; Zhou, K.; Zhang, Y.T.; Feng, N.P. Enhanced immunization via dissolving microneedle array-based delivery system incorporating subunit vaccine and saponin adjuvant. Int. J. Nanomed. 2017, 12, 4763–4772. [Google Scholar] [CrossRef]
- Li, C.; Lee, A.; Grigoryan, L.; Arunachalam, P.S.; Scott, M.K.D.; Trisal, M.; Wimmers, F.; Sanyal, M.; Weidenbacher, P.A.; Feng, Y.; et al. Mechanisms of innate and adaptive immunity to the Pfizer-BioNTech BNT162b2 vaccine. Nat. Immunol. 2022, 23, 543–555. [Google Scholar] [CrossRef]
- Krieg, A.M.; Yi, A.K.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995, 374, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Wang, Y.; Luan, N.; Liu, C. Immunogenicity of Varicella-Zoster Virus Glycoprotein E Formulated with Lipid Nanoparticles and Nucleic Immunostimulators in Mice. Vaccines (Basel) 2021, 9, 310. [Google Scholar] [CrossRef]
- Guimaraes, P.P.G.; Zhang, R.; Spektor, R.; Spektor, R.; Tan, M.; Chung, A.; Billingsley, M.M.; El-Mayta, R.; Riley, R.S.; Wang, L.; et al. Ionizable lipid nanoparticles encapsulating barcoded mRNA for accelerated in vivo delivery screening. J. Control Release 2019, 316, 404–417. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, J.; Cao, H.; Liu, C. Immune Responses to Varicella-Zoster Virus Glycoprotein E Formulated with Poly(Lactic-co-Glycolic Acid) Nanoparticles and Nucleic Acid Adjuvants in Mice. Virol. Sin. 2021, 36, 122–132. [Google Scholar] [CrossRef]
- Chen, D.; Yang, H.; Cao, Y.; Cheng, W.; Duan, T.; Fan, C.; Fan, S.; Feng, L.; Gao, Y.; He, F.; et al. Expert consensus for managing pregnant women and neonates born to mothers with suspected or confirmed novel coronavirus (COVID-19) infection. Int. J. Gynaecol. Obstet. 2020, 149, 130–136. [Google Scholar] [PubMed]
- Luan, N.; Cao, H.; Wang, Y.; Lin, K.; Liu, C. Ionizable Lipid Nanoparticles Enhanced the Synergistic Adjuvant Effect of CpG ODNs and QS21 in a Varicella Zoster Virus Glycoprotein E Subunit Vaccine. Pharmaceutics 2022, 14, 973. [Google Scholar] [CrossRef] [PubMed]
- Schnyder, J.L.; Garcia Garrido, H.M.; De Pijper, C.A.; Daams, J.G.; Stijnis, C.; Goorhuis, A.; Grobusch, M.P. Comparison of equivalent fractional vaccine doses delivered by intradermal and intramuscular or subcutaneous routes: A systematic review. Travel Med. Infect. Dis. 2021, 41, 102007. [Google Scholar] [CrossRef] [PubMed]
- Welsby, I.; Detienne, S.; N’Kuli, F.; Thomas, S.; Wouters, S.; Bechtold, V.; De Wit, D.; Gineste, R.; Reinheckel, T.; Elouahabi, A.; et al. Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21. Front. Immunol. 2016, 7, 663. [Google Scholar]
- Chu, R.S.; Targoni, O.S.; Krieg, A.M.; Lehmann, P.V.; Harding, C.V. CpG oligodeoxynucleotides act as adjuvants that switch on T helper 1 (Th1) immunity. J. Exp. Med. 1997, 186, 1623–1631. [Google Scholar] [CrossRef]
- Syenina, A.; Gan, E.S.; Toh, J.Z.N.; de Alwis, R.; Lin, L.Z.; Tham, C.Y.L.; Yee, J.X.; Leong, Y.S.; Sam, H.; Cheong, C.; et al. Adverse effects following anti-COVID-19 vaccination with mRNA-based BNT162b2 are alleviated by altering the route of administration and correlate with baseline enrichment of T and NK cell genes. PLoS Biol. 2022, 20, e3001643. [Google Scholar] [CrossRef]
- Beals, C.R.; Railkar, R.A.; Schaeffer, A.K.; Levin, Y.; Kochba, E.; Meyer, B.K.; Evans, R.K.; Sheldon, E.A.; Lasseter, K.; Lang, N.; et al. Immune response and reactogenicity of intradermal administration versus subcutaneous administration of varicella-zoster virus vaccine: An exploratory, randomised, partly blinded trial. Lancet Infect Dis. 2016, 16, 915–922. [Google Scholar] [CrossRef]
- Peyraud, N.; Zehrung, D.; Jarrahian, C.; Frivold, C.; Orubu, T.; Giersing, B. Potential use of microarray patches for vaccine delivery in low- and middle- income countries. Vaccine 2019, 37, 4427–4434. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Group | gE mRNA (μg) | gE Protein (μg) | CpG ODNs (μg) | QS21 (μg) | Alum | Injection | |
|---|---|---|---|---|---|---|---|
| BW006S | 2395S | ||||||
| 1. mRNA (I.M.) | 15 | - | - | - | - | - | I.M. |
| 2. mRNA (S.C.) | 15 | - | - | - | - | - | S.C. |
| 3. Subunit + B2Q (I.M.) | - | 10 | 5 | 5 | 5 | - | I.M. |
| 4. Subunit + B2Q (S.C.) | - | 10 | 5 | 5 | 5 | - | S.C. |
| 5. Subunit + Alum (I.M.) | - | 10 | - | - | - | √ | I.M. |
| 6. Subunit + Alum (S.C.) | - | 10 | - | - | - | √ | S.C. |
| 7. PBS (I.M.) | - | - | - | - | - | - | I.M. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, K.; Cao, H.; Luan, N.; Wang, Y.; Hu, J.; Liu, C. Comparison of the Immune Effects of an mRNA Vaccine and a Subunit Vaccine against Herpes Zoster Administered by Different Injection Methods. Vaccines 2023, 11, 1003. https://doi.org/10.3390/vaccines11051003
Lin K, Cao H, Luan N, Wang Y, Hu J, Liu C. Comparison of the Immune Effects of an mRNA Vaccine and a Subunit Vaccine against Herpes Zoster Administered by Different Injection Methods. Vaccines. 2023; 11(5):1003. https://doi.org/10.3390/vaccines11051003
Chicago/Turabian StyleLin, Kangyang, Han Cao, Ning Luan, Yunfei Wang, Jingping Hu, and Cunbao Liu. 2023. "Comparison of the Immune Effects of an mRNA Vaccine and a Subunit Vaccine against Herpes Zoster Administered by Different Injection Methods" Vaccines 11, no. 5: 1003. https://doi.org/10.3390/vaccines11051003
APA StyleLin, K., Cao, H., Luan, N., Wang, Y., Hu, J., & Liu, C. (2023). Comparison of the Immune Effects of an mRNA Vaccine and a Subunit Vaccine against Herpes Zoster Administered by Different Injection Methods. Vaccines, 11(5), 1003. https://doi.org/10.3390/vaccines11051003