
Sph2(176–191) and Sph2(446–459): Identification of B-Cell Linear Epitopes in Sphingomyelinase 2 (Sph2), Naturally Recognized by Patients Infected by Pathogenic Leptospires

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Studied Proteins
2.2. Epitope Prediction
2.3. Antigenicity Analysis
2.4. Conservation Analysis
2.5. Peptide Synthesis
2.6. Studied Population
2.7. Evaluation of Natural Immunogenicity of Predicted Epitopes
2.8. Statistical Analysis of Data
3. Results
3.1. Predicted Epitopes
3.2. Antigenicity Assessment
3.3. Conservation Analysis
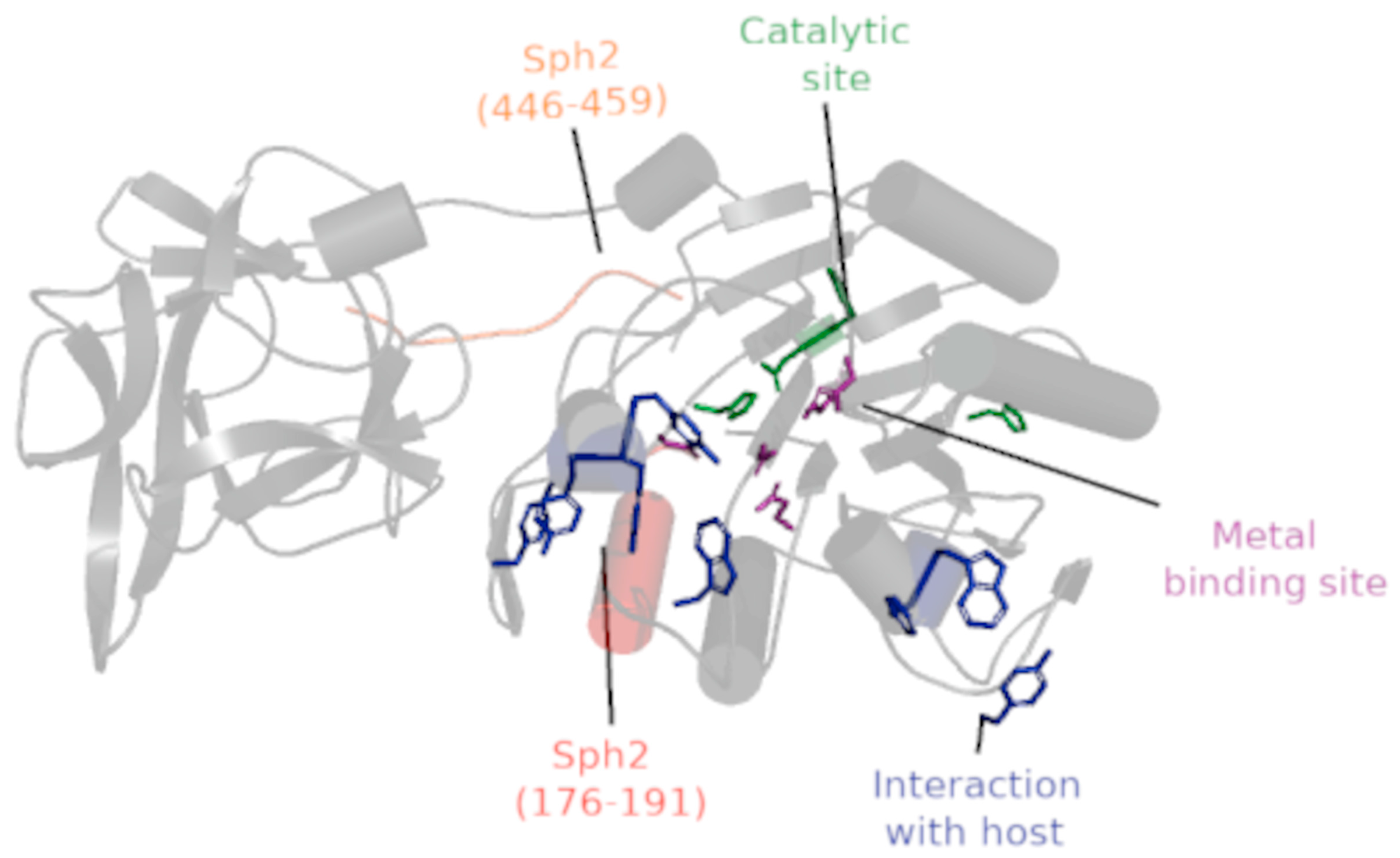
3.4. Epitopes Location and Sph2 Active Sites
3.5. Studied Population
3.6. Naturally Acquired Antibodies against Sph2(176-191) and Sph2(446-459)
3.7. Associations between Humoral Response and Clinical Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adler, B.; de la Pena Moctezuma, A. Leptospira and leptospirosis. Vet. Microbiol. 2010, 140, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef]
- Bharti, A.R.; Nally, J.E.; Ricaldi, J.N.; Matthias, M.A.; Diaz, M.M.; Lovett, M.A.; Levett, P.N.; Gilman, R.H.; Willig, M.R.; Gotuzzo, E.; et al. Leptospirosis: A zoonotic disease of global importance. Lancet Infect. Dis. 2003, 3, 757–771. [Google Scholar] [CrossRef]
- Saúde, M.D. Leptospirose: Diagnóstico e Manejo Clínico. 2014. Available online: https://bvsms.saude.gov.br/bvs/publicacoes/leptospirose-diagnostico-manejo-clinico2.pdf (accessed on 1 February 2023).
- Schneider, C. Leptospirosis in the Americas—Surveillance and Drivers; PAHO Health Emergencies Department GLEAN Meeting; Pan American Health Organization: Washington, DC, USA, 2016. [Google Scholar]
- Plank, R.; Dean, D. Overview of the epidemiology, microbiology, and pathogenesis of Leptospira spp. in humans. Microbes Infect. 2000, 2, 1265–1276. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.I.; Goarant, C.; Picardeau, M. Leptospira: The dawn of the molecular genetics era for an emerging zoonotic pathogen. Nat. Rev. Microbiol. 2009, 7, 736–747. [Google Scholar] [CrossRef] [PubMed]
- De Brito, T.; Silva, A.; Abreu, P.A.E. Pathology and pathogenesis of human leptospirosis: A commented review. Rev. Do Inst. De Med. Trop. De Sao Paulo 2018, 60, e23. [Google Scholar] [CrossRef] [PubMed]
- Dolhnikoff, M.; Mauad, T.; Bethlem, E.P.; Carvalho, C.R. Pathology and pathophysiology of pulmonary manifestations in leptospirosis. Braz. J. Infect. Dis. Off. Publ. Braz. Soc. Infect. Dis. 2007, 11, 142–148. [Google Scholar] [CrossRef]
- Vincent, A.T.; Schiettekatte, O.; Goarant, C.; Neela, V.K.; Bernet, E.; Thibeaux, R.; Ismail, N.; Mohd Khalid, M.K.N.; Amran, F.; Masuzawa, T.; et al. Revisiting the taxonomy and evolution of pathogenicity of the genus Leptospira through the prism of genomics. PLoS Negl. Trop. Dis. 2019, 13, e0007270. [Google Scholar] [CrossRef]
- Dellagostin, O.A.; Grassmann, A.A.; Hartwig, D.D.; Felix, S.R.; da Silva, E.F.; McBride, A.J. Recombinant vaccines against leptospirosis. Hum. Vaccines 2011, 7, 1215–1224. [Google Scholar] [CrossRef]
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef]
- Narayanavari, S.A.; Sritharan, M.; Haake, D.A.; Matsunaga, J. Multiple leptospiral sphingomyelinases (or are there?). Microbiology 2012, 158, 1137–1146. [Google Scholar] [CrossRef]
- Guegan, R.; Camadro, J.M.; Saint Girons, I.; Picardeau, M. Leptospira spp. possess a complete haem biosynthetic pathway and are able to use exogenous haem sources. Mol. Microbiol. 2003, 49, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Bramley, A.J.; Patel, A.H.; O’Reilly, M.; Foster, R.; Foster, T.J. Roles of alpha-toxin and beta-toxin in virulence of Staphylococcus aureus for the mouse mammary gland. Infect. Immun. 1989, 57, 2489–2494. [Google Scholar] [CrossRef]
- Gonzalez-Zorn, B.; Dominguez-Bernal, G.; Suarez, M.; Ripio, M.T.; Vega, Y.; Novella, S.; Vazquez-Boland, J.A. The smcL gene of Listeria ivanovii encodes a sphingomyelinase C that mediates bacterial escape from the phagocytic vacuole. Mol. Microbiol. 1999, 33, 510–523. [Google Scholar] [CrossRef]
- Picardeau, M.; Bulach, D.M.; Bouchier, C.; Zuerner, R.L.; Zidane, N.; Wilson, P.J.; Creno, S.; Kuczek, E.S.; Bommezzadri, S.; Davis, J.C.; et al. Genome sequence of the saprophyte Leptospira biflexa provides insights into the evolution of Leptospira and the pathogenesis of leptospirosis. PLoS ONE 2008, 3, e1607. [Google Scholar] [CrossRef] [PubMed]
- Narayanavari, S.A.; Kishore, N.M.; Sritharan, M. Structural analysis of the Leptospiral sphingomyelinases: In silico and experimental evaluation of Sph2 as an Mg-dependent sphingomyelinase. J. Mol. Microbiol. Biotechnol. 2012, 22, 24–34. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Geng, Y.; Yang, J.W.; Guo, X.K.; Zhao, G.P. Cytotoxic activity and probable apoptotic effect of Sph2, a sphigomyelinase hemolysin from Leptospira interrogans strain Lai. BMB Rep. 2008, 41, 119–125. [Google Scholar] [CrossRef]
- Carvalho, E.; Barbosa, A.S.; Gomez, R.M.; Oliveira, M.L.; Romero, E.C.; Goncales, A.P.; Morais, Z.M.; Vasconcellos, S.A.; Ho, P.L. Evaluation of the expression and protective potential of Leptospiral sphingomyelinases. Curr. Microbiol. 2010, 60, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Artiushin, S.; Timoney, J.F.; Nally, J.; Verma, A. Host-inducible immunogenic sphingomyelinase-like protein, Lk73.5, of Leptospira interrogans. Infect. Immun. 2004, 72, 742–749. [Google Scholar] [CrossRef]
- Cia, G.; Pucci, F.; Rooman, M. Critical review of conformational B-cell epitope prediction methods. Brief. Bioinform. 2023, 24, bbac567. [Google Scholar] [CrossRef]
- Caoili, S.E.C. Comprehending B-Cell Epitope Prediction to Develop Vaccines and Immunodiagnostics. Front. Immunol. 2022, 13, 908459. [Google Scholar] [CrossRef]
- Fontes, S.D.S.; Maia, F.M.; Ataides, L.S.; Conte, F.P.; Lima-Junior, J.D.C.; Rozental, T.; da Silva Assis, M.R.; Junior, A.A.P.; Fernandes, J.; de Lemos, E.R.S.; et al. Identification of Immunogenic Linear B-Cell Epitopes in C. burnetii Outer Membrane Proteins Using Immunoinformatics Approaches Reveals Potential Targets of Persistent Infections. Pathogens 2021, 10, 1250. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, R.C.; Fernandes, J.; de Sampaio Lemos, E.R.; de Paiva Conte, F.; Rodrigues-da-Silva, R.N. The Serological Cross-Detection of Bat-Borne Hantaviruses: A Valid Strategy or Taking Chances? Viruses 2021, 13, 1188. [Google Scholar] [CrossRef]
- Conte, F.P.; Tinoco, B.C.; Santos Chaves, T.; Oliveira, R.C.; Figueira Mansur, J.; Mohana-Borges, R.; Lemos, E.R.S.; Neves, P.; Rodrigues-da-Silva, R.N. Identification and validation of specific B-cell epitopes of hantaviruses associated to hemorrhagic fever and renal syndrome. PLoS Negl. Trop. Dis. 2019, 13, e0007915. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-da-Silva, R.N.; Correa-Moreira, D.; Soares, I.F.; de-Luca, P.M.; Totino, P.R.R.; Morgado, F.N.; Oliveira Henriques, M.D.G.; Peixoto Candea, A.L.; Singh, B.; Galinski, M.R.; et al. Immunogenicity of synthetic peptide constructs based on PvMSP9(E795-A808), a linear B-cell epitope of the P. vivax Merozoite Surface Protein-9. Vaccine 2019, 37, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-da-Silva, R.N.; Martins da Silva, J.H.; Singh, B.; Jiang, J.; Meyer, E.V.; Santos, F.; Banic, D.M.; Moreno, A.; Galinski, M.R.; Oliveira-Ferreira, J.; et al. In silico Identification and Validation of a Linear and Naturally Immunogenic B-Cell Epitope of the Plasmodium vivax Malaria Vaccine Candidate Merozoite Surface Protein-9. PLoS ONE 2016, 11, e0146951. [Google Scholar] [CrossRef]
- Janin, J.; Wodak, S. Conformation of amino acid side-chains in proteins. J. Mol. Biol. 1978, 125, 357–386. [Google Scholar] [CrossRef]
- Ponnuswamy, P.K.; Prabhakaran, M.; Manavalan, P. Hydrophobic packing and spatial arrangement of amino acid residues in globular proteins. Biochim. Et Biophys. Acta 1980, 623, 301–316. [Google Scholar] [CrossRef]
- Parker, J.M.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar] [CrossRef]
- Larsen, J.E.P.; Lund, O.; Nielsen, M. Improved method for predicting linear B-cell epitopes. Immunome Res. 2006, 2, 2. [Google Scholar] [CrossRef][Green Version]
- Emini, E.A.; Hughes, J.V.; Perlow, D.; Boger, J. Induction of hepatitis A virus-neutralizing antibody by a virus-specific synthetic peptide. J. Virol. 1985, 55, 836–839. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins Struct. Funct. Bioinform. 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Ponomarenko, J.; Bui, H.-H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. BcePred: Prediction of continuous B-cell epitopes in antigenic sequences using physico-chemical properties. In Proceedings of the International Conference on Artificial Immune Systems, Catania, Italy, 13–16 September 2004; pp. 197–204. [Google Scholar]
- Singh, H.; Ansari, H.R.; Raghava, G.P. Improved method for linear B-cell epitope prediction using antigen’s primary sequence. PLoS ONE 2013, 8, e62216. [Google Scholar] [CrossRef] [PubMed]
- Sweredoski, M.J.; Baldi, P. COBEpro: A novel system for predicting continuous B-cell epitopes. Protein Eng. Des. Sel. PEDS 2009, 22, 113–120. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Luu, T.; Pham, S.; Deshpande, S. Automated multiple peptide synthesis: Improvements in obtaining quality peptides. Int. J. Pept. Protein Res. 1996, 47, 91–97. [Google Scholar] [CrossRef]
- Gausepohl, H.; Boulin, C.; Kraft, M.; Frank, R.W. Automated multiple peptide synthesis. Pept. Res. 1992, 5, 315–320. [Google Scholar]
- Goni, F.M.; Alonso, A. Sphingomyelinases: Enzymology and membrane activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef]
- Segers, R.P.; van Gestel, J.A.; van Eys, G.J.; van der Zeijst, B.A.; Gaastra, W. Presence of putative sphingomyelinase genes among members of the family Leptospiraceae. Infect. Immun. 1992, 60, 1707–1710. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Kohn, B.; Plog, S.; Weingart, C.; Nockler, K.; Mayer-Scholl, A.; Gruber, A.D. An emerging pulmonary haemorrhagic syndrome in dogs: Similar to the human leptospiral pulmonary haemorrhagic syndrome? Vet. Med. Int. 2010, 2010, 928541. [Google Scholar] [CrossRef]
- Thompson, J.C.; Manktelow, B.W. Pathogenesis of renal lesions in haemoglobinaemic and non-haemoglobinaemic leptospirosis. J. Comp. Pathol. 1989, 101, 201–214. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, S.; Park, S.C.; Kim, M.J. Cytotoxic activities of Leptospira interrogans hemolysin SphH as a pore-forming protein on mammalian cells. Infect. Immun. 2002, 70, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.P.; Figueiredo, F.B.; Morgado, F.N.; Renzetti, A.; de Souza, S.M.M.; Pereira, S.A.; Rodrigues-Da-Silva, R.N.; Lima-Junior, J.D.C.; De Luca, P.M. Leishmania infantum Virulence Factor A2 Protein: Linear B-Cell Epitope Mapping and Identification of Three Main Linear B-Cell Epitopes in Vaccinated and Naturally Infected Dogs. Front. Immunol. 2018, 9, 1690. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-da-Silva, R.N.; Soares, I.F.; Lopez-Camacho, C.; Martins da Silva, J.H.; Perce-da-Silva, D.S.; Teva, A.; Ramos Franco, A.M.; Pinheiro, F.G.; Chaves, L.B.; Pratt-Riccio, L.R.; et al. Plasmodium vivax Cell-Traversal Protein for Ookinetes and Sporozoites: Naturally Acquired Humoral Immune Response and B-Cell Epitope Mapping in Brazilian Amazon Inhabitants. Front. Immunol. 2017, 8, 77. [Google Scholar] [CrossRef]
- Barazzone, G.C.; Teixeira, A.F.; Azevedo, B.O.P.; Damiano, D.K.; Oliveira, M.P.; Nascimento, A.; Lopes, A.P.Y. Revisiting the Development of Vaccines Against Pathogenic Leptospira: Innovative Approaches, Present Challenges, and Future Perspectives. Front. Immunol. 2021, 12, 760291. [Google Scholar] [CrossRef]
- Matsuo, Y.; Yamada, A.; Tsukamoto, K.; Tamura, H.; Ikezawa, H.; Nakamura, H.; Nishikawa, K. A distant evolutionary relationship between bacterial sphingomyelinase and mammalian DNase I. Protein Sci. A Publ. Protein Soc. 1996, 5, 2459–2467. [Google Scholar] [CrossRef]
- Oda, M.; Hashimoto, M.; Takahashi, M.; Ohmae, Y.; Seike, S.; Kato, R.; Fujita, A.; Tsuge, H.; Nagahama, M.; Ochi, S.; et al. Role of sphingomyelinase in infectious diseases caused by Bacillus cereus. PLoS ONE 2012, 7, e38054. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Zorn, B.; Dominguez-Bernal, G.; Suarez, M.; Ripio, M.T.; Vega, Y.; Novella, S.; Rodriguez, A.; Chico, I.; Tierrez, A.; Vazquez-Boland, J.A. SmcL, a novel membrane-damaging virulence factor in Listeria. Int. J. Med Microbiol. IJMM 2000, 290, 369–374. [Google Scholar] [CrossRef]
- Browne, E.S.; Callefe, J.L.R.; Jesus, E.R.S.; Zeppelini, C.G.; Cremonese, C.; Costa, F. A Systematic Review of the geographic distribution of pathogenic Leptospira serovars in the Americas, 1930-2017. An. Da Acad. Bras. De Cienc. 2022, 94, e20201026. [Google Scholar] [CrossRef]
- Lo, M.; Cordwell, S.J.; Bulach, D.M.; Adler, B. Comparative transcriptional and translational analysis of leptospiral outer membrane protein expression in response to temperature. PLoS Negl. Trop. Dis. 2009, 3, e560. [Google Scholar] [CrossRef]
- Xu, Y.; Ye, Q. Human leptospirosis vaccines in China. Hum. Vaccines Immunother. 2018, 14, 984–993. [Google Scholar] [CrossRef]
- Lin, X.; Xiao, G.; Luo, D.; Kong, L.; Chen, X.; Sun, D.; Yan, J. Chimeric epitope vaccine against Leptospira interrogans infection and induced specific immunity in guinea pigs. BMC Microbiol. 2016, 16, 241. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Teixeira, A.F.; Filho, A.F.S.; Souza, G.O.; Vasconcellos, S.A.; Heinemann, M.B.; Romero, E.C.; Nascimento, A. Immune response and protective profile elicited by a multi-epitope chimeric protein derived from Leptospira interrogans. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2017, 57, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Validi, M.; Karkhah, A.; Prajapati, V.K.; Nouri, H.R. Immuno-informatics based approaches to design a novel multi epitope-based vaccine for immune response reinforcement against Leptospirosis. Mol. Immunol. 2018, 104, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Garba, B.; Bahaman, A.R.; Zakaria, Z.; Bejo, S.K.; Mutalib, A.R.; Bande, F.; Suleiman, N. Antigenic potential of a recombinant polyvalent DNA vaccine against pathogenic leptospiral infection. Microb. Pathog. 2018, 124, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Lata, S.; Shankar, U.N.; Akif, M. Immunoinformatics-Based Designing of a Multi-Epitope Chimeric Vaccine From Multi-Domain Outer Surface Antigens of Leptospira. Front. Immunol. 2021, 12, 735373. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein | Species | Serogroup | Serovar | Uniprot ID | Length (a.a.) | Mass (Da) |
|---|---|---|---|---|---|---|
| Sph2 | L. interrogans | Icterohaemorrhagiae | Lai | P59116 | 623 | 71030 |
| Australis | Lora | M3E462 | 507 | 58766 | ||
| Bataviae | Bataviae | M6TDN9 | 423 | 48137 | ||
| Pyrogenes | Zanoni | M6I333 | 507 | 58766 | ||
| Pyrogenes | Pyrogenes | M7AA65 | 567 | 64539 | ||
| L. alexanderi | Manhao | Manhao 3 | V6I1S1 | 458 | 52886 | |
| L. alstonii | Ranarum | Pingchang | T0FYI7 | 543 | 61077 | |
| Undesignated | Sichuan | M6D0Q2 | 543 | 61077 | ||
| L. borgpetersenii | Pomona | Pomona | M6W693 | 566 | 64409 | |
| Sejroe | Hardjo-bovis | M6BNC3 | 556 | 63269 | ||
| L. noguchii | Panama | Panama | T0FTK3 | 616 | 70064 | |
| Autumnalis | Autumnalis | M6UEI2 | 591 | 67772 | ||
| L. kirschneri | Autumnalis | Bulgarica | M6F3J0 | 559 | 64827 | |
| L. santarosai | Javanica | Arenal | M6JW85 | 622 | 70525 | |
| L. weilii | Ranarum | Ranarum | N1WDQ5 | 664 | 74211 | |
| Tarassovi | Topaz | M3FRC5 | 758 | 81501 | ||
| SphH | L. interrogans | Icterohaemorrhagiae | Lai | O34095 | 554 | 64433 |
| SpH1 | L. interrogans | Icterohaemorrhagiae | Lai | P59115 | 597 | 68192 |
| Sph3 | L. interrogans | Icterohaemorrhagiae | Lai | A0A0E2DC81 | 596 | 68095 |
| Sph4 | L. interrogans | Icterohaemorrhagiae | Lai | A0A0E2DCF7 | 623 | 70995 |
| smcL | L. ivanovii | Q9RLV9 | 335 | 38455 | ||
| BCSmase | B. cereus | P09599 | 333 | 36949 | ||
| Sph | S. aureus | A0A7U4AUV1 | 330 | 37238 | ||
| SphH | Pseudomonas sp. | Q93HR5 | 516 | 58114 |
| Sph2(176-191) | Sph2(446-459) | |
|---|---|---|
| Predicted Sequence | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| Bepipred | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| ESA | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| ABCpred | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| ElliPro | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| BCePred-ERS | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| BCePRed-HS | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| BCePRed-PS | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| Lbtope | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| COBEpro | GHDERAKRISKSDYVK | TPTKSGHKKKYDQV |
| Gene | Specie | Serogroup | Serovar | Sph2 | Sph2(176-191) | Sph2(446-459) | ||
|---|---|---|---|---|---|---|---|---|
| Identity | Identity | GHDERAKRISKSDYVK | Identity | TPTKSGHKKKYDQV | ||||
| Sph2 | L. interrogans | Australis | Lora | 55.0% | 43.8% | .QEK..RLLVD.K... | 71.4% | ........R....I |
| Bataviae | Bataviae | 90.0% | 100.0% | ................ | 85.7% | ...Q.A.RR..... | ||
| Pyrogenes | Zanoni | 55.0% | 43.8% | .QEK..RLLVD.K... | 71.4% | ...Q.A.RR..... | ||
| Pyrogenes | Pyrogenes | 67.1% | 56.3% | .QE...Q..AS.S.I. | 92.9% | ........R..... | ||
| L. alexanderi | Manhao | Manhao 3 | 73.2% | 50.0% | .QN...Q..VS.N.IQ | 78.6% | ......R.R...R. | |
| L. alstonii | Ranarum | Pingchang | 69.7% | 43.8% | AQN...EL.AS..HI. | 85.7% | .......RR..... | |
| Undesignated | Sichuan | 69.7% | 43.8% | AQN...EL.AS..HI. | 85.7% | .......RR..... | ||
| L. borgpetersenii | Pomona | Pomona | 70.6% | 50.0% | .QN...Q..VS.N.IQ | 78.6% | ......R.R...R. | |
| Sejroe | Hardjo-bovis | 70.6% | 50.0% | .QN...Q..VS.N.IQ | 78.6% | ......R.R...R. | ||
| L. noguchii | Panama | Panama | 89.6% | 100.0% | ................ | 85.7% | ...E....R..... | |
| Autumnalis | Autumnalis | 89.6% | 100.0% | ................ | 85.7% | ........R....I | ||
| L. kirschneri | Autumnalis | Bulgarica | 47.2% | 31.3% | .QE...NLLLN.QHIQ. | 78.6% | ...F.T......R. | |
| L. santarosai | Javanica | Arenal | 69.3% | 50.0% | .QN...E..AS.N.IR | 85.7% | ........R...R. | |
| L. weilii | Ranarum | Ranarum | 64.1% | 43.8% | .QND..E..ASAN.I. | 92.9% | ........R..... | |
| Tarassovi | Topaz | 57.6% | 56.3% | .QK...EQ.AN...I. | 85.7% | .......RR..... | ||
| SphH | L. interrogans | Icterohaemorrhagiae | Lai | 53.0% | 43.8% | .QEK..RLLVD.K... | 71.4% | ...Q.A.RR..... |
| Sph1 | L. interrogans | Icterohaemorrhagiae | Lai | 66.4% | 56.3% | .QE...Q..AS.S.I. | 92.9% | ........R..... |
| Sph3 | L. interrogans | Icterohaemorrhagiae | Lai | 66.4% | 56.3% | .QE...Q..AS.S.I. | 92.9% | ........R..... |
| Sph4 | L. interrogans | Icterohaemorrhagiae | Lai | 99.3% | 100.0% | ................ | 100.0% | .............. |
| smcL | L. ivanovii | - | - | 46.0% | 43.8% | .QMH..DL.AQA..M. | 28.6% | ..VIGRS.HGW.KT. |
| Sph | B. cereus | - | - | 49.0% | 43.8% | .QSQ..DL.GAA..I. | 35.7% | .V.SWLK.YT..DY |
| Sph | S. aureus | - | - | 48.7% | 43.8% | .QYK..DL.GQ.S.I. | 35.7% | KK...NSL..VATL |
| SphH | Pseudomonas sp. | - | - | 42.0% | 37.5% | AL.IPSP.W.A..G.N | 28.6% | DNDQGQCL.DG... |
| Specie (Serovar)-Protein | Catalytic Site | Amino Acids in the Central Metal-Binding Site | Surface-Exposed Amino Acids Involved with the Host Membrane Interaction | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. interrogans (Lai)-Sph2 | H293 | D393 | Y394 | H433 | N161 | E200 | D341 | N343 | D432 | W172 | Y242 | W274 | F275 | Y382 | Y383 | Y384 | Y425 |
| L. interrogans (Australis)-Sph2 | H230 | T328 | W398 | - | S98 | G137 | D278 | N280 | - | W109 | S179 | Q211 | Y212 | Y319 | Y320 | Y321 | Y337 |
| L. interrogans (Bataviae)-Sph2 | H236 | D336 | Y337 | H376 | N104 | E143 | D284 | N286 | D375 | W115 | Y185 | W217 | F218 | Y325 | Y326 | Y327 | Y368 |
| L. interrogans (Lora)-Sph2 | H175 | E274 | Y274 | R314 | N40 | G79 | S221 | N223 | D313 | S51 | F121 | Y156 | Y157 | Y262 | T263 | S264 | Y306 |
| L. interrogans (Zanoni)-Sph2 | H175 | E274 | Y274 | R314 | N40 | G79 | S221 | N223 | D313 | S51 | F121 | Y156 | Y157 | Y262 | T263 | S264 | Y306 |
| L. interrogans (Pyrogenes)-Sph2 | H232 | D332 | Y332 | R373 | S100 | G139 | D280 | N282 | D372 | W111 | S181 | Q213 | Y214 | Y321 | Y322 | Y323 | Y365 |
| L. alexanderi (Manhao3)-Sph2 | H126 | D226 | Y226 | H266 | - | E33 | D174 | N176 | D265 | W5 | D75 | F107 | F108 | F215 | Y216 | Y217 | Y258 |
| L. alstonii (Pingchang)-Sph2 | H211 | D311 | Y311 | H351 | N79 | E118 | D259 | N261 | D350 | W90 | L160 | Y192 | F193 | F300 | Y301 | Y302 | Y343 |
| L. alstonii (Sichuan)-Sph2 | H211 | D311 | Y311 | H351 | N79 | E118 | D259 | N261 | D350 | W90 | L160 | 192Y | F193 | F300 | Y301 | Y302 | Y343 |
| L. borgpetersenii (Pomona)-Sph2 | H234 | D334 | Y334 | H374 | N102 | E141 | D282 | N284 | D373 | W113 | D183 | V215 | F216 | F325 | Y326 | Y327 | Y366 |
| L. borgpetersenii (Hardjo-bovis)-Sph2 | H224 | D324 | Y324 | H364 | N92 | E131 | D272 | N274 | D363 | W103 | D173 | V205 | F206 | F313 | Y314 | Y315 | Y356 |
| L. noguchii (Panama)-Sph2 | H286 | D386 | Y386 | H426 | N154 | E193 | D334 | N336 | D425 | W165 | Y235 | W267 | F268 | Y375 | Y376 | Y377 | Y418 |
| L. noguchii (Autumnalis)-Sph2 | H261 | D361 | Y361 | H401 | N129 | E168 | D309 | N311 | D400 | W140 | Y210 | W242 | F243 | Y350 | Y351 | Y352 | Y393 |
| L. kirschneri (Bulgarica)-Sph2 | H224 | N329 | Y330 | Y369 | N88 | G127 | N274 | G276 | D368 | T99 | T170 | Y203 | F204 | Y315 | L316 | Q317 | Y361 |
| L. santarosai (Arenal)-Sph2 | H290 | D390 | Y390 | H430 | N158 | E197 | D338 | N340 | D429 | W169 | E239 | F271 | F272 | F379 | Y380 | Y381 | Y422 |
| L. weilii (Ranarum)-Sph2 | H331 | D431 | Y431 | Y472 | N199 | R238 | N379 | N381 | D471 | W210 | S280 | Q312 | Y313 | F420 | K421 | Y422 | Y464 |
| L. weilii (Topaz)-Sph2 | H421 | N521 | Y522 | S562 | N289 | G328 | D469 | N471 | D561 | W300 | S370 | S402 | S403 | L510 | K511 | Y512 | H554 |
| L. interrogans (Lai)-SphH | H222 | E321 | Y322 | R361 | N87 | G126 | S268 | N270 | D360 | S98 | F168 | Y203 | Y204 | Y309 | T310 | S311 | Y353 |
| L. interrogans(Lai)-Sph1 | H262 | D362 | Y363 | R403 | S130 | G169 | D310 | N312 | D402 | W141 | S211 | Q243 | Y244 | Y351 | Y352 | Y353 | Y395 |
| L. interrogans (Lai)-Sph4 | H261 | D361 | Y362 | R402 | S129 | G168 | D309 | N311 | D401 | W140 | S210 | Q242 | Y243 | Y350 | Y351 | Y352 | Y394 |
| L. interrogans (Lai)-Sph4 | H293 | D393 | Y394 | H433 | N161 | E200 | D341 | N343 | D432 | W172 | Y242 | W274 | F275 | Y382 | Y383 | Y384 | Y425 |
| L. ivanovii-smcL | H180 | D282 | Y283 | H325 | N51 | E88 | D229 | N231 | D324 | - | F130 | R161 | L162 | E271 | S272 | Y273 | Y317 |
| B. cereus-Sph | H173 | D280 | Y281 | H323 | N43 | E80 | D222 | N224 | D322 | - | S123 | N154 | L155 | Y269 | N270 | F271 | Y315 |
| S. aureus-Sph | H178 | D279 | Y280 | H322 | N49 | E86 | D227 | N229 | D321 | - | T128 | N159 | D160 | Y268 | N269 | Y270 | Y314 |
| Pseudomonas sp.-SphH | H166 | D277 | Y278 | H320 | N37 | E75 | D214 | N216 | D319 | F48 | N117 | R147 | L148 | Y256 | Q257 | Y258 | W312 |
| Epidemiological and Clinical Data | RL (n = 51) | NRL (n = 36) | |
|---|---|---|---|
| n (%) | |||
| Male | 46 (90.2%) | 28 (77.8%) | |
| Female | 5 (9.8%) | 8 (22.2%) | |
| Deaths | 6 (11.8%) | 4 (11.1%) | |
| Median (Interquartile Range) | |||
| Age | 37 (22–49) | 36.5 (17–45) | |
| Days of Symptoms | 9 (6–15) | 7 (5–16) | |
| Symptoms | n (%) | p-value | |
| Fever | 44 (86.3%) | 29 (80.6%) | |
| Myalgia | 43 (84.3%) | 24 (66.7%) | |
| Headache | 34 (66.7%) | 21 (58.3%) | |
| Jaundice | 35 (68.6%) | 15 (41.7%) | 0.0159 |
| Calf Pain | 31 (60.8%) | 15 (41.7%) | 0.0257 |
| Renal Insufficiency | 20 (39.2%) | 5 (13.9%) | 0.0463 |
| Pulmonary Hemorrhage | 7 (13.7%) | 0 (0%) | 0.0382 |
| Hemorrhagic Signs | 8 (15.7%) | 2 (5.6%) | |
| Prostration | 27 (52.9%) | 18 (50.0%) | |
| Vomit | 24 (47.1%) | 18 (50.0%) | |
| Diarrhea | 20 (39.2%) | 11 (30.6%) | |
| Respiratory Changes | 18 (35.3%) | 10 (27.8%) | |
| Conjunctival Congestion | 10 (19.6%) | 8 (22.2%) | |
| Abdominal Pain | 4 (7.8%) | 5 (13.9%) | |
| Diagnosed Leptospira spp. Serovars | n (%) | ||
| Tarassovi | 16 (31.4%) | ||
| Copenhageni | 12 (23.5%) | ||
| Grippotyphosa | 2 (3.9%) | ||
| Canicola | 2 (3.9%) | ||
| Icterohaemorrhagiae | 1 (2%) | ||
| Wolffi | 1 (2%) | ||
| Pomona | 1 (2%) | ||
| Tarassovi/Copenhageni | 5 (9.8%) | ||
| Copenhageni/Icterohaemorrhagiae | 5 (9.8%) | ||
| Tarassovi/Icterohaemorrhagiae | 1 (2%) | ||
| Copenhageni/Hebdomadis | 1 (2%) | ||
| Tarassovi/Grippotyphosa | 1 (2%) | ||
| Wolffi/Sejroe | 1 (2%) | ||
| Australis/Hebdomadis/Autumnalis | 1 (2%) | ||
| Copenhageni/Canicola/Icterohaemorrhagiae/Tarassovi | 1 (2%) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ataides, L.S.; de Moraes Maia, F.; Conte, F.P.; Isaac, L.; Barbosa, A.S.; da Costa Lima-Junior, J.; Avelar, K.E.S.; Rodrigues-da-Silva, R.N. Sph2(176–191) and Sph2(446–459): Identification of B-Cell Linear Epitopes in Sphingomyelinase 2 (Sph2), Naturally Recognized by Patients Infected by Pathogenic Leptospires. Vaccines 2023, 11, 359. https://doi.org/10.3390/vaccines11020359
Ataides LS, de Moraes Maia F, Conte FP, Isaac L, Barbosa AS, da Costa Lima-Junior J, Avelar KES, Rodrigues-da-Silva RN. Sph2(176–191) and Sph2(446–459): Identification of B-Cell Linear Epitopes in Sphingomyelinase 2 (Sph2), Naturally Recognized by Patients Infected by Pathogenic Leptospires. Vaccines. 2023; 11(2):359. https://doi.org/10.3390/vaccines11020359
Chicago/Turabian StyleAtaides, Laura Sant’Anna, Fernanda de Moraes Maia, Fernando Paiva Conte, Lourdes Isaac, Angela Silva Barbosa, Josué da Costa Lima-Junior, Kátia Eliane Santos Avelar, and Rodrigo Nunes Rodrigues-da-Silva. 2023. "Sph2(176–191) and Sph2(446–459): Identification of B-Cell Linear Epitopes in Sphingomyelinase 2 (Sph2), Naturally Recognized by Patients Infected by Pathogenic Leptospires" Vaccines 11, no. 2: 359. https://doi.org/10.3390/vaccines11020359
APA StyleAtaides, L. S., de Moraes Maia, F., Conte, F. P., Isaac, L., Barbosa, A. S., da Costa Lima-Junior, J., Avelar, K. E. S., & Rodrigues-da-Silva, R. N. (2023). Sph2(176–191) and Sph2(446–459): Identification of B-Cell Linear Epitopes in Sphingomyelinase 2 (Sph2), Naturally Recognized by Patients Infected by Pathogenic Leptospires. Vaccines, 11(2), 359. https://doi.org/10.3390/vaccines11020359