Abstract
The ongoing antibiotic-resistance crisis is becoming a global problem affecting public health. Urgent efforts are required to design novel therapeutics against pathogenic bacterial species. Brucella melitensis is an etiological agent of brucellosis, which mostly affects sheep and goats but several cases have also been reported in cattle, water buffalo, yaks and dogs. Infected animals also represent the major source of infection for humans. Development of safer and effective vaccines for brucellosis remains a priority to support disease control and eradication in animals and to prevent infection to humans. In this research study, we designed an in-silico multi-epitopes vaccine for B. melitensis using computational approaches. The pathogen core proteome was screened for good vaccine candidates using subtractive proteomics, reverse vaccinology and immunoinformatic tools. In total, 10 proteins: catalase; siderophore ABC transporter substrate-binding protein; pyridoxamine 5′-phosphate oxidase; superoxide dismutase; peptidylprolyl isomerase; superoxide dismutase family protein; septation protein A; hypothetical protein; binding-protein-dependent transport systems inner membrane component; and 4-hydroxy-2-oxoheptanedioate aldolase were selected for epitopes prediction. To induce cellular and antibody base immune responses, the vaccine must comprise both B and T-cells epitopes. The epitopes were next screened for antigenicity, allergic nature and water solubility and the probable antigenic, non-allergic, water-soluble and non-toxic nine epitopes were shortlisted for multi-epitopes vaccine construction. The designed vaccine construct comprises 274 amino acid long sequences having a molecular weight of 28.14 kDa and instability index of 27.62. The vaccine construct was further assessed for binding efficacy with immune cell receptors. Docking results revealed that the designed vaccine had good binding potency with selected immune cell receptors. Furthermore, vaccine-MHC-I, vaccine-MHC-II and vaccine-TLR-4 complexes were opted based on a least-binding energy score of −5.48 kcal/mol, 0.64 kcal/mol and −2.69 kcal/mol. Those selected were then energy refined and subjected to simulation studies to understand dynamic movements of the docked complexes. The docking results were further validated through MMPBSA and MMGBSA analyses. The MMPBSA calculated −235.18 kcal/mol, −206.79 kcal/mol, and −215.73 kcal/mol net binding free energy, while MMGBSA estimated −259.48 kcal/mol, −206.79 kcal/mol and −215.73 kcal/mol for TLR-4, MHC-I and MHC-II complexes, respectively. These findings were validated by water-swap and entropy calculations. Overall, the designed vaccine construct can evoke proper immune responses and the construct could be helpful for experimental researchers in formulation of a protective vaccine against the targeted pathogen for both animal and human use.
1. Introduction
Antibiotic resistance by bacteria significantly contributes to human morbidity and mortality. This is due to the irrational use of antibiotics in humans, animals, the environment and agricultural fields [1]. Antibiotic resistance is a bacterial evolution process to adjust to changing environmental milieu [2]. Novel approaches are needed to combat this alarming global health concern [3]. The use of bacterial genomic information in vaccine design is a promising approach at the present time. This technique is referred to as reverse vaccinology, which has remarkably changed the field of vaccine design [4]. Vaccines can be used to provoke host immune responses against infectious pathogens. Antigens can be either biological or synthetic in nature [5].
In the beginning of 15th century, smallpox disease resulted in high mortality and morbidity rates. Both Turks and Chinese were trying to induce immunity against smallpox by using powder smallpox lesions [6]. Pasteur’s vaccinology concept was used by Salk and Sabin to design an effective poliovirus vaccine. Reverse vaccinology is a novel tool for vaccine production, which compared to traditional vaccines is a cheap process and can be done in a short time [7]. Identification of new vaccine targets through genomics and computational techniques has not only speed up the vaccine development process but also delivers new antigens not disclosed through experimental methods [8]. For finding putative surface-associated proteins, reverse vaccinology is used without any culturing of microorganisms [9]. Using the same process, meningococcal serogroup B (4DMenB) vaccine was developed [10]. Compared to simple reverse vaccinology, pan-genomic reverse vaccinology is generally more effective as it screens highly conserved targets [11]. As an example, for Streptococcus agalactiae four different protective antigens were unveiled by pan-genomic reverse vaccinology [12,13,14,15].
In this study, comparative genomics and reverse vaccinology methods were used for identification of protective vaccine antigens against Brucella melitensis, which is a Gram-negative coccobacillus bacterium from the Brucellaceae family [16]. B. melitensis is an etiological agent of brucellosis, which mostly affects sheep and goats but several cases have also been reported in cattle, water buffalo, yaks and dogs [17]. The pathogen causes brucellosis in goats and Malta fever in humans [17,18]. Infected animals represent the major source of infection for humans, through direct exposure or through consumption of contaminated and unpasteurized dairy products. In endemic areas, vaccination of susceptible animals would reduce disease prevalence also limiting the risk of disease transmission to humans [19]. Previous efforts on B. melitensis vaccination are dominated by work on the Rev1 vaccine which is effective against sheep and goat brucellosis [20]. The vaccine comprises smooth lipopolysaccharide with O-polysaccharide able to elicit strong antibody responses [21]. Rev1 is a live attenuated vaccine and despite its efficacy, several drawbacks remain due to its residual pathogenicity. The Rev1 vaccine may also result in abortion in pregnant animals [22,23]. Previous attempts to produce safer and effective recombinant brucella vaccines included two antigens (periplasmic bp26 and chaperone trigger factor proteins); however, both candidates were not able to induce protective and accurate immune responses [22,24]. Despite these efforts, no licensed vaccine is available to prevent brucellosis infection. In addition, drug-resistant strains of B. melitensis are making the situation worse. Thus, considering this, herein we applied an integrated approach comprising comparative genomics, subtractive proteomics, reverse vaccinology, immunoinformatic, and biophysics techniques to identify protective antigens from B. melitensis completely sequenced genomes and designed a multi-epitopes vaccine [25,26,27,28,29,30,31,32,33]. The designed vaccine construct then was examined for interactions with host immune receptors to check whether the vaccine is able to be presented to the host immunity system. The findings of this study may help in formulating vaccines design against B. melitensis.
2. Materials and Methods
The study framework used for designing a multi-epitopes vaccine against B. melitensis is illustrated in Figure 1.
Figure 1.
Outline of methodology that was used for designing a multi-epitopes vaccine against B. melitensis.
2.1. Proteomes Retrieval of B. melitensis
The complete proteomic data of fully sequenced B. melitensis strains were extracted from the national center for biotechnological information (NCBI) genome database [34]. The proteomic data were retrieved in FASTA format.
2.2. BPGA Analysis
For identification of suitable vaccine candidates, all the proteomes were screened with the bacterial pan genomic analysis (BPGA) tool [35]. During this analysis, different sets of proteins were identified such as core, accessory and unique. The core proteins were picked and used in downward analysis as they have broad spectrum applicability [36,37,38,39].
2.3. CD-Hit Analysis
In the pre-screening phase, the core sequences were subjected to redundancy checks to remove duplicate copies of the proteins [40]. The non-redundant proteins are single presentations in the proteome and do not require extra computational cost. In the CD-hit (cluster data with high identity with tolerance) analysis, core proteins were analyzed for the presence of redundant proteins [41,42,43]. Non-redundant proteins were selected as the best candidates for the vaccine design [44,45]. The core proteins were clustered at a threshold of 0.5 which means that the sequences that were 50% similar were clustered together.
2.4. Subcellular Localization Phase
In the field of vaccine design, surface proteins elicit robust immune response [46,47,48]. The non-redundant proteome was examined though PSORTb 3.0, which is a bacterial protein subcellular localization prediction program [49].
2.5. Homology Check
In the homology check, the shortlisted proteins were blast through the BLASTp (basic local alignment search tool) against the human proteomes for sequence similarity [50]. Those proteins with E values < 1.0−4, bit scores <100 and <30% sequence identity were selected [36,37]. Homologous proteins provoke autoimmune reactions [51]. A similar homology check was also performed against intestinal probiotic bacteria Lactobacillus rhamnosus (taxid: 47715), L. casei (taxid: 1582) and L. johnsonii (taxid: 33959) to avoid their function inhibition [52].
2.6. Vaccine Candidate’s Prioritization Phase
In the vaccine candidate prioritization phase, the main focus was on prioritization of potential vaccine candidates for vaccine design.
2.7. Virulent Protein Analysis
Virulent proteins are good vaccine candidates as they have the ability to stimulate immune responses of the host [53]. The proteins were blast using BLASTp against the core virulent factor database (VFDB) [54]. The proteins selected as the best choices for vaccine design exhibited a sequence identity ≥30% and a bit score >100. Those proteins that were below the set parameters were discarded [55].
2.8. Physiochemical Analysis
The filtered proteins were analyzed for physiochemical properties including, molecular weight, atomic composition, instability index, theoretical PI, amino acid composition, aliphatic index, grand average of hydropath city (GRAVY) and estimated half-life [56]. These properties were analyzed using the ProtParam-2017 tool [57]. Those proteins having a molecular weight less than 110 kDa and thermostability index less than 40 were considered to be good candidates for vaccine design as they can be easily purified [58].
2.9. Transmembrane Helices
The proteins that have less transmembrane helicase were considered as good candidates for vaccine design [31,33]. For the analysis of transmembrane helicase, HMMTOP 2.0 [59] and TMHMM 2.0 [60] softwares were used. The threshold values were set at 0 and 1. Proteins that were exposed and had values of 0 and 1 were selected [32].
2.10. Antigenicity Prediction
For stimulation of the host immune system, foreign antigens should be capable of binding with the host immune cells; that capability of antigens is known as antigenicity [61]. For the detection of antigenicity of shortlisted proteins, the online server VaxiJen 2.0 was used [62]. The threshold value for the antigenicity of proteins was set at 0.4. Those proteins that had antigenicity scores >0.5 were considered as good choices for vaccine design [47].
2.11. Adhesion Probability Analysis
For stimulation of the host immune system, vaccine antigens should be able to attach to the host cells and start the infection process to be recognized as antigens by the host immune system [55]. After attachment, adaptive immunity is developed which includes T cell receptors and antibodies [63]. For the analysis of the adhesion probability of proteins, the online server Vaxign was used [64]. The threshold for the selection of a good protein was set at >0.5 [36].
2.12. Allergenicity of the Proteins
Those proteins that were allergens to the host were removed and only non-allergenic proteins were selected. The allergenicity of the proteins was determined through the online webserver AllerTOP 2.0 [65].
2.13. Epitopes Prediction
The epitope is the main part of the antigen to which the host immune cells bind. In prediction of epitopes, the B cell and T cell epitopes were analyzed via the immune epitope database (IEDB) tool [66]. The threshold for the IEDB server was set at 0.5. In the epitope-prediction phase, the B cell epitopes were first predicted and then T cell epitopes. The T cell epitopes were analyzed for the prediction of potential binding alleles for the major histocompatibility complex (MHC) class I and MHC class II. Prediction of binding of epitopes with the DRB*10101 alleles was analyzed using the MHcPred tool [67]. The threshold value set was IC50 values ≤100 nM [68]. Those exceeding this threshold value were discarded.
2.14. Physiochemical Analysis of the Predicated Epitopes
The selected epitopes were then subjected to physiochemical properties analysis. The antigenicity and allergenicity of the epitopes were analyzed using VaxiJen 2.0 [62] and AllerTop 2.0 [65], respectively. The virulence property was checked via Virulentpred [69]. The antigenic and non-allergen epitopes were further analyzed using ToxinPred for toxicity [70]. All those epitopes having good water solubility were considered as good candidates. The ProteinSol tool (https://protein-sol.manchester.ac.uk/; accessed on 16 October 2022) was used for the prediction of water solubility.
2.15. Multi-Epitopes Peptide Designing
Peptide vaccines have weak ability to provoke host immune responses. The weak immunogenicity of the peptide vaccine can be handled with the technique of multi-epitopes peptide design [71,72]. In multi-epitopes peptide-design phase, immunodominant epitopes were linked to each other [73]. All the selected screened epitopes were linked with the Gly-Pro-Gly-Pro-Gly (GPGPG) linkers to build an immunopotent multi-epitopes peptide vaccine. Furthermore, the vaccine was linked with the highly immunopotent cholera toxin B adjuvant (CTB) for the enhancement of the vaccine immunogenicity [74,75]. The adjuvant binds to the monoganglioside GM1 receptor and is capable of stimulating cytokines, interferone, cellular and humoral immunity [75]. The adjuvant is used in vaccine design against cancer, tuberculosis and influenza [76].
2.16. Physiochemical Properties Analysis
The final vaccine construct was then subjected to various physiochemical properties. The online server ExPASy Protparam was used for the evaluation of molecular weight, number of amino acids, instability index, Grand average of hydropathicity (GRAVY), theoretical PI value and aliphatic index [57].
2.17. Structural Prediction of Multi-Epitope Peptide
The final vaccine construct was then run on the 3Dpro tool of the SCRATCH protein predictor for construction of the tertiary structure [77]. A good vaccine consists of a smaller number of loops, is small in size and simple in structure. For the removal of loops, the final vaccine construct was then run on the Galaxy Loop tool of the GalaxyWeb online server [78].
2.18. Galaxy Refinement
For the final refinement of the loop-modelled vaccine construct, the GalaxyRefine tool was used for the construction of side chains and removal of steric clashes [79]. The fully refined form of the vaccine construct was considered to be good candidate for vaccine design.
2.19. Disulfide Engineering
The stability of a vaccine is very important; thus, for better stability the bonding of the inner and outer chains was checked in silico. At the disulfide engineering phase, the Design 2.0 webserver was used for the introduction of disulfide bonds in the vaccine construct [80].
2.20. In Silico Codon Optimization and Coding
The sequence of the multi-epitope vaccine construct was translated into a DNA sequence and then cloned into the expression vector to be expressed in Escherichia coli. The conversion of the vaccine into DNA was performed via the Java Codon Adaptation Tool (JCat) [81]. The expression level of the cloned sequence was evaluated through the “GC’ concentration and the “Codon Adaptation Index score (CAI)”. Preferably, the CAI score should be 1 and the GC content should be 30–70% [82].
2.21. Docking and Refinement
The binding affinity of the multi-epitope vaccine construct with the immune cell receptors was analyzed through blind molecular docking [83]. In the PatchDock server [84], the TLR-4, MHC-I and MHC-Ⅱ receptors were selected. The tertiary structures of these receptors were collected from the Protein Data Bank (PDB) by using the codes 4G8A, 1I1Y and 1KG0, respectively. The output solutions were generated by the PatchDock server and were then subjected to further refinement via the online webserver fast interaction refinement in molecular docking (FireDock) [85]. After the refinements, those complexes that exhibited the least global energy were ranked at the top and were selected for further analysis. The intermolecular interactions of complexes were analyzed via UCSF Chimera software [86].
2.22. Molecular Dynamics Stimulation (MDS) Assay
The dynamic behavior of the vaccine complexes was analyzed by using a molecular dynamics stimulation approach. Based on the lowest global energy value, the complexes were chosen for the MDS [87]. The AMBER20 stimulation software was used for simulation on a timescale of 200 ns [88]. For the completion of system setup phase, preprocessing and final production phase, the AMBER SANDER module was used [89]. The intermolecular interactions in the MDS were defined by FF14SB force field [90]. SHAKE algorithm was used to constrain hydrogen bonds [91]. The pressure equilibrium of the system was maintained by NPT ensemble. For the evaluation of trajectories, the CPPTRAJ module was used [92]. To investigate structure stability of complexes, root mean square deviation (RMSD) and root mean square fluctuation (RMSF) plots were produced in XMGRCE [93].
2.23. Free Energy of Immune Receptors and Vaccine Design
The tool MMPBSA.py provided in AMBER20 was used for the determination of binding free energies of the docked complexes [94,95,96]. In binding free energy analysis, a total of 100 frames were selected from the trajectories. Vaccine immune receptor complexes are stable having lower free binding energies [48].
2.24. WaterSwap Validation and Entropy Analysis
The MMPBSA often ignore the water molecule contribution in bridging the vaccine and receptors residues. Therefore, water-swap analysis was conducted with default settings to reconfirm stable intermolecular interactions [97]. Also, entropy energy calculations were done on 5 frames using AMBER normal mode analysis [98].
3. Results
3.1. Retrieval of Complete Proteome, Bacterial Pan-Genome Analysis and Subtractive Proteomics Filters
The study was commenced with the retrieval of 95 fully sequenced strains of B. melitensis. The extracted strains were subjected to bacterial pa-genome analysis steps for retrieval of core sequences. In total, 238,450 core sequences were predicted. The core proteins were good targets for broad spectrum vaccine development. The core sequences were further considered for redundancy analysis. The webserver revealed that the 238,450 core sequences consisted of 2551 non-redundant proteins and 235,899 redundant proteins. The redundant proteins were discarded, and the non-redundant proteins were further considered for surface-localization analysis. The non-redundant proteins are single presentations in the proteomes and require less computational expense to process them [40]. The analysis revealed that non-redundant proteins consisted of 26 outer-membrane, 9 extracellular and 71 periplasmic membrane proteins. Next, VFDB analysis was performed, which predicted that the surface localized proteins consisted of 12 virulent proteins. Virulent proteins are good vaccine targets as they have antigenic epitopes capable of stimulating immune responses [46]. In 12 virulent proteins, 3 proteins were predicted to have more than 1 transmembrane helix. Overall, 12 proteins were predicted to be probably antigenic, have good water solubility and be non-toxic. Moreover, no physiochemically unstable, host or normal flora similar proteins were found. The category and number of proteins are presented in Figure 2. The size of each proteome is presented in Figure 3.
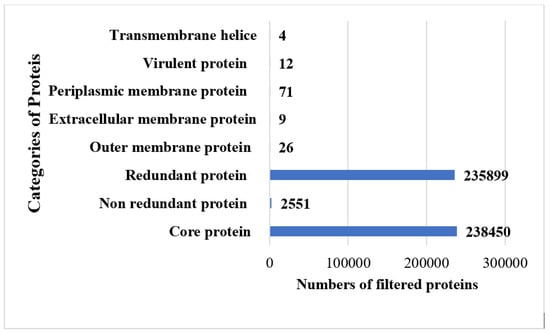
Figure 2.
Shortlisted proteins in each step of subtractive proteomics filter.
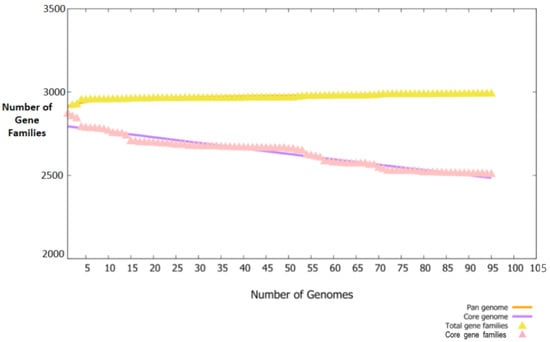
Figure 3.
Pan-core plot. The number of fully sequenced proteomes used are plotted on the X-axis while gene families are plotted against each proteome on the Y-axis.
3.2. Epitopes Prediction
B and B-cell derived T-cell epitopes were predicted from the shortlisted 11 proteins given in Table 1. The predicted B-cell epitopes were further utilized for T-cell epitopes. The predicted MHC-I and MHC-II epitopes are tabulated in Table S1 with least percentile scores. The common and lowest percentile score epitopes were opted for downward analyses.

Table 1.
Predicted B-cells epitopes. The epitopes vary in length and have higher scores than the threshold.
3.3. Selection of Epitopes for Multi-Epitopes Vaccine Construction
Final set epitopes were selected for vaccine construct by applying several filters. Only antigenic, allergenic, water solubility and non-toxic B and B-cell derived T-cell epitopes were considered. Table 2 shows selected epitopes utilized in multi-epitopes vaccine construction. The schematic diagram of vaccine construct comprising selected epitopes is presented in Figure 4.
Table 2.
Selected epitopes for vaccine construct.
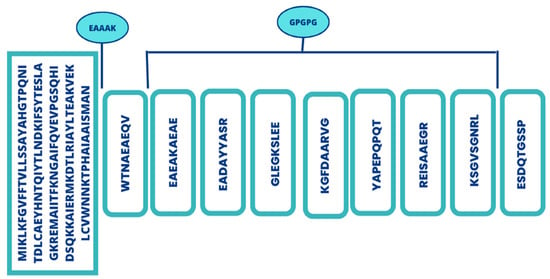
Figure 4.
Schematic representation of multi-epitopes vaccine.
3.4. Structure Prediction and Disulfide Engineering
The vaccine 3D structure was predicted using a scratch predictor. The structure of the vaccine comprises cholera toxin B subunit as an adjuvant molecule, EAAAK and GPGPG linkers along with selected epitopes as presented in Figure 5. The EAAAK and GPGPG linkers are rigid in nature and keep the epitopes separated which will allow the epitopes to be recognized by the host immune system for efficient recognition and processing [38]. The adjuvant cholera toxin B subunit is considered a powerful adjuvant as it generates specific immunity and mucosal antibody responses. The adjuvant binds to GM1 ganglioside, which is present on antigen-presenting cells, lymphocytes and epithelia cells. The adjuvant conjugation to antigens can result in activation of dendritic cells, decrease in antigen dose, and enhanced B and T-cell responses [75,76,99]. Furthermore, to retain structural stability of the vaccine, disulfide bonds were established between weak energy pairs. The enzymatic sensitive residue bonds were supplemented by cysteine bonds. The pairs of amino-acid residues that were disulfide engineered are tabulated in Table 3 and presented in Figure 6 by yellow sticks.
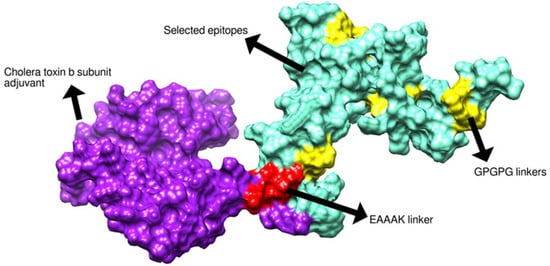
Figure 5.
Three-dimensional structure of designed vaccine.
Table 3.
Pairs of amino-acid residues Chi-3 values and energy.
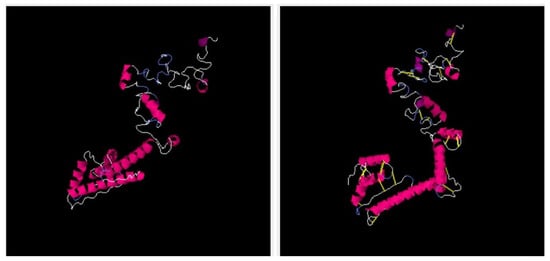
Figure 6.
Left side picture represents wild type structure while right side picture represents mutated structure.
3.5. Loops Refinement
In the loop-refinement phase, the structure of the designed vaccine construct was examined for loops refinement. A total of 10 refine models were generated and model 1 was selected for docking analysis. The top 10 refined models are tabulated in Table 4. The refined model has a good MolProbity score of 1.475, improved clash score of 2.3 and high percentage of residues in Rama-favored regions (92.6).
Table 4.
Model, RMSD, MolProbity, clash score, poor rotamers, Rama-favored and GALAXY energy of refined complexes.
3.6. Codon Optimization Phase
The sequence of the multi-epitopes vaccine construct was translated into a DNA sequence and then cloned into the expression system of Escherichia coli. This was carried out to get higher expression of the cloned vaccine as E. coli is a good expression system. The CAI score of the vaccine was 0.95 and the GC content was 49.56%. Both these values indicate good expression of the vaccine construct might be expected. Cloning of the vaccine construct is mentioned in Figure 7.
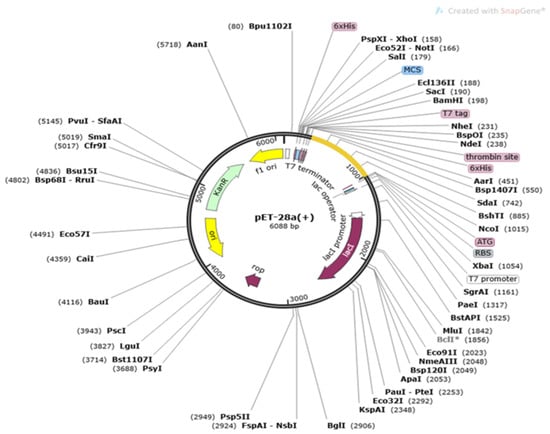
Figure 7.
Cloning of multi-epitopes vaccine constructs (yellow color) into pET28a (+) vector.
3.7. Docking and Refinement
Molecular docking analysis was carried out in order to analyze the binding mode of the vaccine construct with immune cell receptors (MHC-I, MHC-II and TLR-4). The patch dock web server generated the top 20 docked complexes as tabulated in Tables S2–S4. The docked complexes were further refined using the fire docked webserver. The server generated the top 10 refined docked complexes as mentioned in Table 5, Table 6 and Table 7. In the case of the vaccine-MHC-I complex, solution 1 was selected as it has the lowest global energy score of −5.48 kcal/mol. The major energy contribution was seen from attractive van der Waals energy. For vaccine-MHC-II, solution 9 was opted as it has lowest global energy of 0.64 kcal/mol. Similarly, for the vaccine-TLR-4 complex, solution 7 was opted for with a net global energy score of −2.69 kcal/mol. In all the selected complexes, it was found that the vaccine docked with receptors in a stable conformation and the vaccine antigens were exposed to and recognized by the host immune system. The intermolecular docked complexes are presented in Figure 8.
Table 5.
Top 10 refined complexes of vaccine and MHC-I molecule. vdW (van der Waals energy), ACE (atomic contact energy), and HB (hydrogen bond energy).
Table 6.
Top 10 refined complexes of vaccine and MHC-II molecule. vdW (van der Waals energy), ACE (atomic contact energy), and HB (hydrogen bond energy).
Table 7.
Top 10 refined complexes of vaccine and TLR-4 molecule. vdW (van der Waals energy), ACE (atomic contact energy), and HB (hydrogen bond energy).
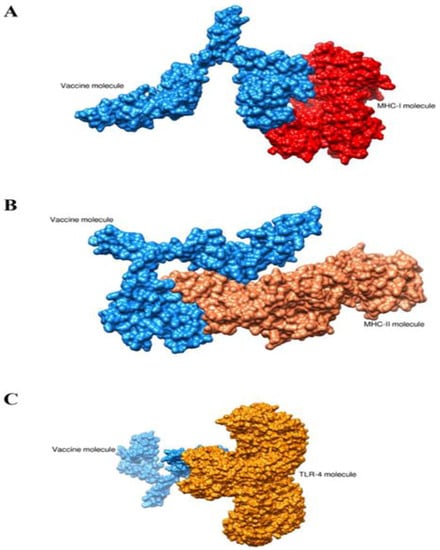
Figure 8.
Intermolecular docked conformation of complexes. (A) vaccine-MHC-I complex, (B) vaccine-MHC-II complex and (C) vaccine-TLR-4 complex.
3.8. MDS Analysis
MDS analysis was carried out to check the binding stability and dynamics of docked complexes. The docked complexes were investigated at 200 ns periods of time. The trajectories of the MDS consisted of RMSD and RMSF analyses [100,101]. As compared to vaccine-MHC-II and TLR-4, vaccine and MHC-I molecules showed stability as the graph is constant through the simulation time. The vaccine-MHC-II complex showed minor changes which may be due to loops present in the structure but towards the simulation end it became stable. Overall, the RMSD graph shows that there is proper conformational stability between vaccine and receptors molecules. The RMSD graph is presented in Figure 9A. Similarly, residue base fluctuations were assessed through RMSF. In RMSF analysis, it was observed that the vaccine-MHC-II complex has proper stability as mentioned in Figure 9B.
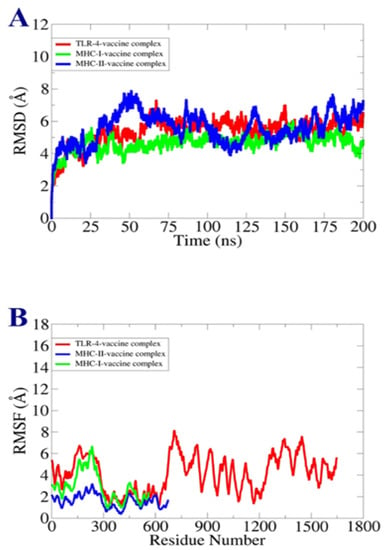
Figure 9.
Simulation trajectories (A) RMSD (B) RMSF.
3.9. Binding Free Energies Calculation
The docking results were further validated using binding free energies calculations. The MMGB/PB/SA analysis was adopted for energy estimation. In MMGBSA analysis, −259.48 kcal/mol, −206.79 kcal/mol, −215.73 kcal/mol delta energy were estimated for vaccine-TLR-4, vaccine-MHC-I and vaccine-MHC-II, respectively. Similarly, in MM-PBSA analysis, for vaccine-TLR-4, vaccine-MHC-I, and vaccine-MHC-II, a net energy of −235.18 kcal/mol, −206.79 kcal/mol and −215.73 kcal/mol was calculated. Details of overall binding energies prediction are tabulated in Table 8.
Table 8.
Binding free energies calculation. The units are kcal/mol.
3.10. WaterSwap and Binding Entropy Calculation
To revalidate the findings and provide more confidence regarding the vaccine’s stable interactions with the receptors, water-swap calculations were performed. Three algorithms were used in the water-swap method i.e. thermodynamic integration, free energy perturbation and Bennetts. The water-swap calculations found highly stable energies for all three complexes [97,102,103]. For the TLR-4-vaccine complex, the water-swap estimations were: thermodynamic integration (−46.5 kcal/mol); free energy perturbation (−47.52 kcal/mol); and Bennetts (−46.97 kcal/mol). The thermodynamic integration, free energy perturbation and Bennetts values for the MHC-I-vaccine complex were −48.5 kcal/mol, −47.0 kcal/mol and −47.41 kcal/mol, respectively. Likewise, for the MHC-II-vaccine complex, the values were −42.1 kcal/mol (thermodynamic integration), −43.6 kcal/mol (free energy perturbation) and −41.6 kcal/mol (Bennetts). All these values indicate good systems convergence and strong intermolecular affinity. Additionally, entropy energy indicates values of 45 kcal/mol for the TLR-4-vaccine complex, 51.87 kcal/mol for the MHC-I-vaccine complex, and 50.2 kcal/mol for the MHC-II-vaccine complex. These findings suggest that the vaccine has less physical freedom and docked well with the receptor for efficient immuen system recognition and processing.
4. Discussion
In this study, nine proteins were prioritized as potential subunit vaccine candidate targets in the complete proteome of B. melitensis based on comprehensive investigation of comparative proteomics, subtractive proteomics, reverse vaccinology, immunoinformatic, and biophysics approaches. Furthermore, antigenic, non-allergic, non-toxic, and water-soluble epitopes were successfully predicted in the mentioned vaccine proteins. A multi-epitopes vaccine construct was built which revealed stable binding conformation and dynamics with different immune receptors such as TLR-4, MHC-I and MHC-II.
The emergence of bacterial resistance to antibiotics is a serious threat to public health [104]. The resistance to antibiotics is alarming as efficacy of commercially available antibiotics is becoming less effective. Additionally, development of new antibiotics is of less interest due to several regulatory challenges faced by pharma companies [105]. This problem can be addressed by a vaccination process [15]. However not all vaccines are effective and helpful in prevention of infectious disease but somehow can reduce the level of infections [106,107]. The conventional Pasture vaccinology suffers from several limitations i.e., time consuming, development of unwanted immune responses, high cost, less efficacy and specificity, not applicable to non-cultivated microbes and less stability [108]. A huge amount of genomic data available in databases could help in identification of good vaccine candidates [12,13,14]. The use of reverse vaccinology, bioinformatics and immmunoinformatics approaches in recent times is an alternative way of designing vaccines against different pathogens [11,15,109]. We used pan-genome analysis, subtractive proteomics analysis immunoinformatics, and molecular docking and simulation methods for the designing of multi-epitopes-based vaccines against B. melitensis. The multi-epitopes vaccines consist of overlapping epitopes and are considered an ideal approach for prevention and treatment of infectious diseases [107]. The success of these vaccines has been elaborated by the EMD640744 vaccine that is currently under phase I clinical trials for advanced solid tumors [107]. These vaccines can generate humoral, cytotoxic and helper T-cell immune responses. They contain epitopes which can be recognized by multiple clones of TCRs and stimulate cellular and humoral immunity simultaneously [107]. They also have the ability to generate enhanced immunogenicity that is long term [107]. Additionally, they are free from unwanted antigens that lead to pathologically adverse reactions [71]. Through subtractive proteomics filters, 10 proteins were selected for B–cell peptides prediction. These proteins were catalase, siderophore ABC transporter substrate-binding protein, pyridoxamine 5′-phosphate oxidase, superoxide dismutase, peptidylprolyl isomerase, superoxide dismutase family protein, septation protein A, hypothetical protein, binding-protein-dependent transport systems inner membrane component, 4-hydroxy-2-oxoheptanedioate aldolase. The shortlisted proteins were utilized for B-cell and T-cell epitopes in the prediction and prioritization phases. With the help of several immunoinformatics approaches, WTNAEAEQV, EAEAKAEAE, EADAYYASR, GLEGKSLEE, KGFDAARVG, YAPEPQPQT, REISAAEGR, KSGVSGNRL, and ESDQTGSSP were prioritized as appropriate epitopes for multi-epitopes vaccine design. The epitopes were tested for antigenicity, allergenicity, toxicity, water solubility, and adhesion probability. Further, non-toxic, probable antigenic, good water-soluble and physiochemically stable epitopes were used in chimeric vaccine construction. The designed vaccine was then utilized for interaction studies with immune cell receptors as it was important to unveil for successful vaccine development. This was achieved by molecular docking that predicted the vaccine candidates’ stable interactions with MHC-I, MHC-II and TLR-4 and thus could evoke humoral and cellular immunity. Intermolecular docked stability of vaccine with immune cell receptors in dynamic environments is important for long-term antigen presenting and processing. The dynamic movement of the docked complexes revealed the vaccine candidates’ proper binding that can generate long-term immunity against the targeted pathogen.
Brucellosis is estimated to cause 500,000 thousand cases each year. Among the Brucella genera, B. melitensis is the most pathogenic species and shows broad resistance to a spectrum of antibiotics especially rifampicin [110]. Therefore, efforts are needed for development of a safe and effective vaccine. In the recent past, several computational efforts have revealed potential vaccine candidates against different bacterial pathogens. A previous study conducted by Ismail, Ahmad and Azam, 2020, successfully predicted an in silico multi-epitopes vaccine against bacterial members of Enterobacteriaceae [38]. In another work, three distinct types of surface peptides were investigated that can effectively provoke the immune response (AtfC), (PMI2533) and (PMI1466) against Proteus mirabilis [111]. Furthermore, computer-aided vaccine-design studies against Pseudomonas aeruginosa [46], Providencia rettgeri [112], Streptococcus pneumoniae [113], and Klebsiella pneumoniae [114] have been successfully carried out in the recent past. The reverse vaccinology approach have also been applied to B. melitensis. However, those studies were not as comprehensive as that conducted herein. In one study, Omp10, Omp25, Omp31 and BtpB were used for epitopes prediction. The designed multi-epitopes vaccine comprised 806 amino acids that were used in different biophysics approaches [115]. In another work, Omp22, Omp28 and Omp19 were utilized for vaccine design [116].
In silico vaccine design is rapidly emerging due to the wide range of genomic data available and development of new bioinformatics tools. These analyses are highly effective and provide new pathways for the synthesis of novel vaccines against resistant pathogens [9,117,118,119]. However, our study has a few limitations. The order of the epitopes in the designed vaccine must be evaluated for optimal biological potency [38]. The choice of delivery route and delivery system are also a challenge [38]. Detailed experimental testing is required to validate the immune potency of the designed vaccine against B. melitensis.
5. Conclusions
In this computer-aided vaccine-design work, 10 proteins; catalase, siderophore ABC transporter substrate-binding protein, pyridoxamine 5′-phosphate oxidase, superoxide dismutase, peptidylprolyl isomerase, superoxide dismutase family protein, septation protein A, hypothetical protein, binding-protein-dependent transport systems inner membrane component and 4-hydroxy-2-oxoheptanedioate aldolase were identified as promising vaccine targets selected for epitopes prediction against B. melitensis. The proteins were forecasted to harbor antigenic epitopes that were capable of eliciting strong humoral, cellular and helper immunological responses. Most of the mentioned targets were not predicted before and are novel in this respect. The designed chimeric multi-epitopes vaccine showed a robust interactions network with different immune receptors ensuring that the vaccine is capable of eliciting a variety of immunological reactions. The vaccine type is safe from allergic and reactogenic responses and accurate/specific in generating immunological response and memory. Further, the multi-epitopes vaccine will be easy to design and could provide better immunogenicity and antigenicity compared to single peptide vaccines. The vaccine construct may also provide experimentalists a ready framework to test vaccine epitopes in vivo and in vitro biological models and thus may speed up the development of a safe, effective and broad spectrum vaccine against B. melitensis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/vaccines11020263/s1, Table S1. Predicted T-cells epitopes. Table S2. Top 20 docked complexes of vaccine and MHC-I molecules. Table S3. Top 20 docked complexes of vaccine and MHC-II molecule. Table S4. Top 20 docked complexes of vaccine and TLR-4 molecule.
Author Contributions
Conceptualization, S.A. and M.K.; methodology, M.M. and S.K.; software, M.M.; validation, A.U., M.u.H. and S.S.; formal analysis, A.I.A.-H.; investigation, M.M. and S.A.A.; resources, M.I.; data curation, M.M.; writing—original draft preparation, M.M. and M.H.; writing—review and editing, S.A.; visualization, M.K.; supervision, S.A.; project administration, S.A.; funding acquisition, S.A. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are thankful to the Department of Health and Biological Sciences, Abasyn University, Peshawar, Pakistan for financial support.
Institutional Review Board Statement
Not relevant to this study.
Informed Consent Statement
Not relevant to this study.
Data Availability Statement
All the data is available in this study.
Conflicts of Interest
The authors confirmed that there is no conflict of interest associated with the study and data reported in it.
References
- Caniça, M.; Manageiro, V.; Abriouel, H.; Moran-Gilad, J.; Franz, C.M.A.P. Antibiotic Resistance in Foodborne Bacteria. Trends Food Sci. Technol. 2019, 84, 41–44. [Google Scholar] [CrossRef]
- MacLean, R.C.; San Millan, A. The Evolution of Antibiotic Resistance. Science 2019, 365, 1082–1083. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 2: Management Strategies and New Agents. Pharm. Ther. 2015, 40, 344. [Google Scholar]
- Masignani, V.; Pizza, M.; Moxon, E.R. The Development of a Vaccine against Meningococcus B Using Reverse Vaccinology. Front. Immunol. 2019, 10, 751. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Clem, A.S. Fundamentals of Vaccine Immunology. J. Glob. Infect Dis. 2011, 3, 73. [Google Scholar] [CrossRef]
- Bidmos, F.A.; Siris, S.; Gladstone, C.A.; Langford, P.R. Bacterial Vaccine Antigen Discovery in the Reverse Vaccinology 2.0 Era: Progress and Challenges. Front. Immunol. 2018, 9, 2315. [Google Scholar] [CrossRef]
- Ahmad, S.; Shahid, F.; Tahir ul Qamar, M.; Abbasi, S.W.; Sajjad, W.; Ismail, S.; Alrumaihi, F.; Allemailem, K.S.; Almatroudi, A.; Ullah Saeed, H.F. Immuno-Informatics Analysis of Pakistan-Based HCV Subtype-3a for Chimeric Polypeptide Vaccine Design. Vaccines 2021, 9, 293. [Google Scholar] [CrossRef] [PubMed]
- Dalsass, M.; Brozzi, A.; Medini, D.; Rappuoli, R. Comparison of Open-Source Reverse Vaccinology Programs for Bacterial Vaccine Antigen Discovery. Front. Immunol. 2019, 10, 113. [Google Scholar] [CrossRef]
- Abdullah, M.; Kadivella, M.; Sharma, R.; Faisal, S.M.; Azam, S. Designing of Multiepitope-Based Vaccine against Leptospirosis Using Immuno-Informatics Approaches. bioRxiv 2021. [Google Scholar] [CrossRef]
- Santos, A.; Ali, A.; Barbosa, E.; Silva, A.; Miyoshi, A.; Barh, D.; Azevedo, V. The Reverse Vaccinology-A Contextual Overview. IIOABJ 2011, 2, 8–15. [Google Scholar]
- Rappuoli, R. Reverse Vaccinology. Curr. Opin. Microbiol. 2000, 3, 445–450. [Google Scholar] [CrossRef]
- Adu-Bobie, J.; Capecchi, B.; Serruto, D.; Rappuoli, R.; Pizza, M. Two Years into Reverse Vaccinology. Vaccine 2003, 21, 605–610. [Google Scholar] [CrossRef]
- Mora, M.; Veggi, D.; Santini, L.; Pizza, M.; Rappuoli, R. Reverse Vaccinology. Drug Discov. Today 2003, 8, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Delany, I.; Rappuoli, R.; Seib, K.L. Vaccines, Reverse Vaccinology, and Bacterial Pathogenesis. Cold Spring Harb. Perspect. Med. 2013, 3, a012476. [Google Scholar] [CrossRef]
- Mazlan, M.; Khairani-Bejo, S.; Hamzah, H.; Nasruddin, N.S.; Salleh, A.; Zamri-Saad, M. Pathological Changes, Distribution and Detection of Brucella Melitensis in Foetuses of Experimentally-Infected Does. Vet. Q. 2021, 41, 36–49. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health Brucellosis. (Brucella Abortus, B. Melitensis and B. Suis) (Infection with B. Abortus, B. Melitensis and B. Suis). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. 2016. Available online: https://www.woah.org/fileadmin/Home/fr/Health_standards/tahm/3.01.04_BRUCELLOSIS.pdf (accessed on 20 December 2022).
- Mantur, B.; Amarnath, S.; Shinde, R. Review of Clinical and Laboratory Features of Human Brucellosis. Indian J. Med. Microbiol. 2007, 25, 188–202. [Google Scholar] [CrossRef]
- de Figueiredo, P.; Ficht, T.A.; Rice-Ficht, A.; Rossetti, C.A.; Adams, L.G. Pathogenesis and Immunobiology of Brucellosis: Review of Brucella--Host Interactions. Am. J. Pathol. 2015, 185, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Ponsart, C.; Riou, M.; Locatelli, Y.; Jacques, I.; Fadeau, A.; Jay, M.; Simon, R.; Perrot, L.; Freddi, L.; Breton, S.; et al. Brucella Melitensis Rev.1 Vaccination Generates a Higher Shedding Risk of the Vaccine Strain in Alpine Ibex (Capra Ibex) Compared to the Domestic Goat (Capra Hircus). Vet. Res. 2019, 50, 1–13. [Google Scholar] [CrossRef]
- González, D.; Grilló, M.J.; de Miguel, M.J.; Ali, T.; Arce-Gorvel, V.; Delrue, R.M.; Conde-Álvarez, R.; Muñoz, P.; López-Goñi, I.; Iriarte, M.; et al. Brucellosis Vaccines: Assessment of Brucella Melitensis Lipopolysaccharide Rough Mutants Defective in Core and O-Polysaccharide Synthesis and Export. PLoS ONE 2008, 3, e2760. [Google Scholar] [CrossRef] [PubMed]
- Costa Oliveira, S.; Costa Macedo, G.; Augusto de Almeida, L.; Souza de Oliveira, F.; Onate, A.; Cassataro, J.; Hernan Giambartolomei, G. Recent Advances in Understanding Immunity Against Brucellosis: Application for Vaccine Development. Open Vet. Sci. J. 2014, 4. [Google Scholar] [CrossRef]
- Dorneles, E.M.S.; Sriranganathan, N.; Lage, A.P. Recent Advances in Brucella Abortus Vaccines. Vet. Res. 2015, 46, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Hudson, M.; Walters, N.; Bargatze, R.F.; Pascual, D.W. Selection of Protective Epitopes for Brucella Melitensis by DNA Vaccination. Infect. Immun. 2005, 73, 7297–7303. [Google Scholar] [CrossRef]
- Mugunthan, S.P.; Harish, M.C. Multi-Epitope-Based Vaccine Designed by Targeting Cytoadherence Proteins of Mycoplasma Gallisepticum. ACS Omega 2021, 6, 13742–13755. [Google Scholar] [CrossRef] [PubMed]
- Adeoti, O.M. Prediction of Multi-Epitopic Domains of a Putative Oral Vaccine against Hepatitis C Virus. Int. J. Immunol. Microbiol. 2021, 1, 16–22. [Google Scholar] [CrossRef]
- Ud-Din, M.; Albutti, A.; Ullah, A.; Ismail, S.; Ahmad, S.; Naz, A.; Khurram, M.; Haq, M.; Afsheen, Z.; Bakri, Y.; et al. Vaccinomics to Design a Multi-Epitopes Vaccine for Acinetobacter Baumannii. Int. J. Environ. Res. Public Health 2022, 19, 5568. [Google Scholar] [CrossRef]
- Fatima, I.; Ahmad, S.; Abbasi, S.W.; Ashfaq, U.A.; Shahid, F.; ul Qamar, M.T.; Rehman, A.; Allemailem, K.S. Designing of a Multi-Epitopes-Based Peptide Vaccine against Rift Valley Fever Virus and Its Validation through Integrated Computational Approaches. Comput. Biol. Med. 2022, 141, 105151. [Google Scholar] [CrossRef]
- Ismail, S.; Abbasi, S.W.; Yousaf, M.; Ahmad, S.; Muhammad, K.; Waheed, Y. Design of a Multi-Epitopes Vaccine against Hantaviruses: An Immunoinformatics and Molecular Modelling Approach. Vaccines 2022, 10, 378. [Google Scholar] [CrossRef]
- Omoniyi, A.A.; Adebisi, S.S.; Musa, S.A.; Nzalak, J.O.; Danborno, B.; Bauchi, Z.M.; Badmus, I.T.; Olatomide, O.D.; Oladimeji, O.J.; Nyengaard, J.R. Immunoinformatics Analysis and In-Silico Design of Multi-Epitopes Vaccine Against Lassa Virus. 2021. Available online: https://assets.researchsquare.com/files/rs-355782/v1/9bf6a115-7a8a-4638-a451-8d0a7330b307.pdf?c=1637244600 (accessed on 22 December 2022).
- Alharbi, M.; Alshammari, A.; Alasmari, A.F.; Alharbi, S.M.; ul Qamar, M.; Ullah, A.; Ahmad, S.; Irfan, M.; Khalil, A.A.K. Designing of a Recombinant Multi-Epitopes Based Vaccine against Enterococcus Mundtii Using Bioinformatics and Immunoinformatics Approaches. Int. J. Environ. Res. Public Health 2022, 19, 3729. [Google Scholar] [CrossRef]
- Rida, T.; Ahmad, S.; Ullah, A.; Ismail, S.; Tahir ul Qamar, M.; Afsheen, Z.; Khurram, M.; Saqib Ishaq, M.; Alkhathami, A.G.; Alatawi, E.A. Pan-Genome Analysis of Oral Bacterial Pathogens to Predict a Potential Novel Multi-Epitopes Vaccine Candidate. Int. J. Environ. Res. Public Health 2022, 19, 8408. [Google Scholar] [CrossRef]
- Ullah, A.; Ahmad, S.; Ismail, S.; Afsheen, Z.; Khurram, M.; Tahir ul Qamar, M.; AlSuhaymi, N.; Alsugoor, M.H.; Allemailem, K.S. Towards A Novel Multi-Epitopes Chimeric Vaccine for Simulating Strong Immune Responses and Protection against Morganella Morganii. Int. J. Environ. Res. Public Health 2021, 18, 10961. [Google Scholar] [CrossRef] [PubMed]
- Coordinators, N.R. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2017, 45, D12. [Google Scholar]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA-an Ultra-Fast Pan-Genome Analysis Pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef]
- Sajjad, R.; Ahmad, S.; Azam, S.S. In Silico Screening of Antigenic B-Cell Derived T-Cell Epitopes and Designing of a Multi-Epitope Peptide Vaccine for Acinetobacter Nosocomialis. J. Mol. Graph. Model 2020, 94, 107477. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Azam, S.S. A Novel Approach of Virulome Based Reverse Vaccinology for Exploring and Validating Peptide-Based Vaccine Candidates against the Most Troublesome Nosocomial Pathogen: Acinetobacter Baumannii. J. Mol. Graph. Model 2018, 83, 1–11. [Google Scholar] [CrossRef]
- Ismail, S.; Ahmad, S.; Azam, S.S. Vaccinomics to Design a Novel Single Chimeric Subunit Vaccine for Broad-Spectrum Immunological Applications Targeting Nosocomial Enterobacteriaceae Pathogens. Eur. J. Pharm. Sci. 2020, 146, 105258. [Google Scholar] [CrossRef] [PubMed]
- Ismail, S.; Ahmad, S.; Azam, S.S. Immunoinformatics Characterization of SARS-CoV-2 Spike Glycoprotein for Prioritization of Epitope Based Multivalent Peptide Vaccine. J. Mol. Liq. 2020, 314, 113612. [Google Scholar] [CrossRef] [PubMed]
- Sanober, G.; Ahmad, S.; Azam, S.S. Identification of Plausible Drug Targets by Investigating the Druggable Genome of MDR Staphylococcus Epidermidis. Gene Rep. 2017, 7, 147–153. [Google Scholar] [CrossRef]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A Web Server for Clustering and Comparing Biological Sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Abbas, G.; Zafar, I.; Ahmad, S.; Azam, S.S. Immunoinformatics Design of a Novel Multi-Epitope Peptide Vaccine to Combat Multi-Drug Resistant Infections Caused by Vibrio Vulnificus. Eur. J. Pharm. Sci. 2020, 142, 105160. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Raza, S.; Uddin, R.; Azam, S.S. Comparative Subtractive Proteomics Based Ranking for Antibiotic Targets against the Dirtiest Superbug: Acinetobacter Baumannii. J. Mol. Graph. Model 2018, 82, 74–92. [Google Scholar] [CrossRef]
- Rashid, M.I.; Naz, A.; Ali, A.; Andleeb, S. Prediction of Vaccine Candidates against Pseudomonas Aeruginosa: An Integrated Genomics and Proteomics Approach. Genomics 2017, 109, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Naz, K.; Naz, A.; Ashraf, S.T.; Rizwan, M.; Ahmad, J.; Baumbach, J.; Ali, A. PanRV: Pangenome-Reverse Vaccinology Approach for Identifications of Potential Vaccine Candidates in Microbial Pangenome. BMC Bioinform. 2019, 20, 123. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Naz, A.; Ahmad, J.; Naz, K.; Obaid, A.; Parveen, T.; Ahsan, M.; Ali, A. VacSol: A High Throughput in Silico Pipeline to Predict Potential Therapeutic Targets in Prokaryotic Pathogens Using Subtractive Reverse Vaccinology. BMC Bioinform. 2017, 18, 106. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved Protein Subcellular Localization Prediction with Refined Localization Subcategories and Predictive Capabilities for All Prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef]
- Blast, N. Basic Local Alignment Search Tool. National Library of Medicine: National Center for Biotechnology Information. 2015. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 20 December 2022).
- Naz, K.; Ullah, N.; Naz, A.; Irum, S.; Dar, H.A.; Zaheer, T.; Shahid, F.; Ali, A. The Epidemiological and Pangenome Landscape of Staphylococcus Aureus and Identification of Conserved Novel Candidate Vaccine Antigens. Curr. Proteom. 2022, 19, 114–126. [Google Scholar] [CrossRef]
- Ahmad, S.; Ranaghan, K.E.; Azam, S.S. Combating Tigecycline Resistant Acinetobacter Baumannii: A Leap Forward towards Multi-Epitope Based Vaccine Discovery. Eur. J. Pharm. Sci. 2019, 132, 1–17. [Google Scholar] [CrossRef]
- Ma, J.; Qiu, J.; Wang, S.; Ji, Q.; Xu, D.; Wang, H.; Wu, Z.; Liu, Q. A Novel Design of Multi-Epitope Vaccine Against Helicobacter Pylori by Immunoinformatics Approach. Int. J. Pept. Res. Ther. 2021, 27, 1027–1042. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef]
- Baseer, S.; Ahmad, S.; Ranaghan, K.E.; Azam, S.S. Towards a Peptide-Based Vaccine against Shigella Sonnei: A Subtractive Reverse Vaccinology Based Approach. Biologicals 2017, 50, 87–99. [Google Scholar] [CrossRef]
- Asad, Y.; Ahmad, S.; Rungrotmongkol, T.; Ranaghan, K.E.; Azam, S.S. Immuno-Informatics Driven Proteome-Wide Investigation Revealed Novel Peptide-Based Vaccine Targets against Emerging Multiple Drug Resistant Providencia Stuartii. J. Mol. Graph. Model 2018, 80, 238–250. [Google Scholar] [CrossRef]
- ProtParam, E. ExPASy-ProtParam Tool; SIB: Lausanne, Switzerland, 2017. [Google Scholar]
- Albekairi, T.H.; Alshammari, A.; Alharbi, M.; Alshammary, A.F.; Tahir ul Qamar, M.; Ullah, A.; Irfan, M.; Ahmad, S. Designing of a Novel Multi-Antigenic Epitope-Based Vaccine against E. Hormaechei: An Intergraded Reverse Vaccinology and Immunoinformatics Approach. Vaccines 2022, 10, 665. [Google Scholar] [CrossRef] [PubMed]
- Tusnady, G.E.; Simon, I. The HMMTOP Transmembrane Topology Prediction Server. Bioinformatics 2001, 17, 849–850. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, P.; Luo, J.; Jiang, Y. Secreted Protein Prediction System Combining CJ-SPHMM, TMHMM, and PSORT. Mamm. Genome 2003, 14, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.; Negre, N.N.; White, K.P.; Li, C.; Shah, P.K. Determinants of Antigenicity and Specificity in Immune Response for Protein Sequences. BMC Bioinform. 2011, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A Server for Prediction of Protective Antigens, Tumour Antigens and Subunit Vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, F.A.; Oettgen, H.C. Adaptive Immunity. J. Allergy Clin. Immunol. 2010, 125, S33–S40. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xiang, Z.; Mobley, H.L.T. Vaxign: The First Web-Based Vaccine Design Program for Reverse Vaccinology and Applications for Vaccine Development. Biomed. Res. Int. 2010, 2010, 297505. [Google Scholar] [CrossRef]
- Dimitrov, I.; Flower, D.R.; Doytchinova, I. AllerTOP-a Server for in Silico Prediction of Allergens. BMC Bioinform. 2013, 14, S4. [Google Scholar] [CrossRef] [PubMed]
- Dhanda, S.K.; Mahajan, S.; Paul, S.; Yan, Z.; Kim, H.; Jespersen, M.C.; Jurtz, V.; Andreatta, M.; Greenbaum, J.A.; Marcatili, P. IEDB-AR: Immune Epitope Database—Analysis Resource in 2019. Nucleic Acids Res. 2019, 47, W502–W506. [Google Scholar] [CrossRef]
- Guan, P.; Doytchinova, I.A.; Zygouri, C.; Flower, D.R. MHCPred: A Server for Quantitative Prediction of Peptide--MHC Binding. Nucleic Acids Res. 2003, 31, 3621–3624. [Google Scholar] [CrossRef] [PubMed]
- Naz, A.; Awan, F.M.; Obaid, A.; Muhammad, S.A.; Paracha, R.Z.; Ahmad, J.; Ali, A. Identification of Putative Vaccine Candidates against Helicobacter Pylori Exploiting Exoproteome and Secretome: A Reverse Vaccinology Based Approach. Infect. Genet. Evol. 2015, 32, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Gupta, D. VirulentPred: A SVM Based Prediction Method for Virulent Proteins in Bacterial Pathogens. BMC Bioinform. 2008, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Kapoor, P.; Chaudhary, K.; Gautam, A.; Kumar, R.; Raghava, G.P.S. Peptide Toxicity Prediction. In Computational Peptidology; Springer: Cham, Switzerland, 2015; pp. 143–157. [Google Scholar]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2019, 120, 3210–3229. [Google Scholar] [CrossRef]
- Reche, P.A.; Fernandez-Caldas, E.; Flower, D.R.; Fridkis-Hareli, M.; Hoshino, Y. Peptide-Based Immunotherapeutics and Vaccines. J. Immunol. Res. 2014, 2014, 256784. [Google Scholar] [CrossRef]
- Alshammari, A.; Alharbi, M.; Alghamdi, A.; Alharbi, S.A.; Ashfaq, U.A.; Tahir ul Qamar, M.; Ullah, A.; Irfan, M.; Khan, A.; Ahmad, S. Computer-Aided Multi-Epitope Vaccine Design against Enterobacter Xiangfangensis. Int. J. Environ. Res. Public Health 2022, 19, 7723. [Google Scholar] [CrossRef] [PubMed]
- Baldauf, K.J.; Royal, J.M.; Hamorsky, K.T.; Matoba, N. Cholera Toxin B: One Subunit with Many Pharmaceutical Applications. Toxins 2015, 7, 974–996. [Google Scholar] [CrossRef]
- Stratmann, T. Cholera Toxin Subunit B as Adjuvant----an Accelerator in Protective Immunity and a Break in Autoimmunity. Vaccines 2015, 3, 579–596. [Google Scholar] [CrossRef]
- Antonio-Herrera, L.; Badillo-Godinez, O.; Medina-Contreras, O.; Tepale-Segura, A.; García-Lozano, A.; Gutierrez-Xicotencatl, L.; Soldevila, G.; Esquivel-Guadarrama, F.R.; Idoyaga, J.; Bonifaz, L.C. The Nontoxic Cholera B Subunit Is a Potent Adjuvant for Intradermal DC-Targeted Vaccination. Front. Immunol. 2018, 9, 2212. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Randall, A.Z.; Sweredoski, M.J.; Baldi, P. SCRATCH: A Protein Structure and Structural Feature Prediction Server. Nucleic Acids Res. 2005, 33, W72–W76. [Google Scholar] [CrossRef]
- Ko, J.; Park, H.; Heo, L.; Seok, C. GalaxyWEB Server for Protein Structure Prediction and Refinement. Nucleic Acids Res. 2012, 40, W294–W297. [Google Scholar] [CrossRef]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein Structure Refinement Driven by Side-Chain Repacking. Nucleic Acids Res. 2013, 41, W384–W388. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.B.; Dombkowski, A.A. Disulfide by Design 2.0: A Web-Based Tool for Disulfide Engineering in Proteins. BMC Bioinform. 2013, 14, 346. [Google Scholar] [CrossRef] [PubMed]
- Grote, A.; Hiller, K.; Scheer, M.; Münch, R.; Nörtemann, B.; Hempel, D.C.; Jahn, D. JCat: A Novel Tool to Adapt Codon Usage of a Target Gene to Its Potential Expression Host. Nucleic Acids Res. 2005, 33, W526–W531. [Google Scholar] [CrossRef]
- Ahmad, S.; Navid, A.; Farid, R.; Abbas, G.; Ahmad, F.; Zaman, N.; Parvaiz, N.; Azam, S.S. Design of a Novel Multi Epitope-Based Vaccine for Pandemic Coronavirus Disease (COVID-19) by Vaccinomics and Probable Prevention Strategy against Avenging Zoonotics. Eur. J. Pharm. Sci. 2020, 151, 105387. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Lim-Wilby, M. Molecular Docking. In Molecular Modeling of Proteins; Springer: Cham, Switzerland, 2008; pp. 365–382. [Google Scholar]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for Rigid and Symmetric Docking. Nucleic Acids Res. 2005, 33, W363–W367. [Google Scholar] [CrossRef]
- Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: Fast Interaction Refinement in Molecular Docking. Proteins Struct. Funct. Bioinform. 2007, 69, 139–159. [Google Scholar] [CrossRef]
- Kaliappan, S.; Bombay, I.I.T. UCSF Chimera-Overview. 2018. Available online: http://doer.col.org/handle/123456789/9120 (accessed on 16 October 2022).
- Hansson, T.; Oostenbrink, C.; van Gunsteren, W. Molecular Dynamics Simulations. Curr. Opin. Struct. Biol. 2002, 12, 190–196. [Google Scholar] [CrossRef]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.; Brozell, S.R.; Cerutti, D.; Cheatham, T.; Cruzeiro, V.W.D.; Darden, T.; Duke, R.E.; Giambasu, G.; et al. Amber 2020. Available online: https://ambermd.org/doc12/Amber20.pdf (accessed on 23 December 2022).
- Kerrigan, J.E. AMBER 10.0 Introductory Tutorial; Citeseer: Princeton, NJ, USA, 2009. [Google Scholar]
- Case, D.A.; Babin, V.; Berryman, J.T.; Betz, R.M.; Cai, Q.; Cerutti, D.S.; Cheatham, T.E., III; Darden, T.A.; Duke, R.E.; Gohlke, H.; et al. The FF14SB Force Field. Amber 2014, 14, 29–31. [Google Scholar]
- Kräutler, V.; van Gunsteren, W.F.; Hünenberger, P.H. A Fast SHAKE Algorithm to Solve Distance Constraint Equations for Small Molecules in Molecular Dynamics Simulations. J. Comput. Chem. 2001, 22, 501–508. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Turner, P.J. XMGRACE, Version 5.1. 19. Center for Coastal and Land-Margin Research; Oregon Graduate Institute of Science and Technology: Beaverton, OR, USA, 2005. [Google Scholar]
- Miller, B.R.; McGee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA.Py: An Efficient Program for End-State Free Energy Calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Sahakyan, H. Improving Virtual Screening Results with MM/GBSA and MM/PBSA Rescoring. J. Comput Aided Mol. Des. 2021, 35, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA Methods to Estimate Ligand-Binding Affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Woods, C.J.; Malaisree, M.; Hannongbua, S.; Mulholland, A.J. A Water-Swap Reaction Coordinate for the Calculation of Absolute Protein-Ligand Binding Free Energies. J. Chem. Phys. 2011, 134, 054114. [Google Scholar] [CrossRef] [PubMed]
- Genheden, S.; Kuhn, O.; Mikulskis, P.; Hoffmann, D.; Ryde, U. The Normal-Mode Entropy in the MM/GBSA Method: Effect of System Truncation, Buffer Region, and Dielectric Constant. J. Chem. Inf. Model 2012, 52, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Czerkinsky, C.; Holmgren, J. Mucosally Induced Immunological Tolerance, Regulatory T Cells and the Adjuvant Effect by Cholera Toxin B Subunit. Scand. J. Immunol. 2010, 71, 1–11. [Google Scholar] [CrossRef]
- Sargsyan, K.; Grauffel, C.; Lim, C. How Molecular Size Impacts RMSD Applications in Molecular Dynamics Simulations. J. Chem. Theory Comput. 2017, 13, 1518–1524. [Google Scholar] [CrossRef]
- Ahmad, S.; Raza, S.; Uddin, R.; Azam, S.S. Binding Mode Analysis, Dynamic Simulation and Binding Free Energy Calculations of the MurF Ligase from Acinetobacter Baumannii. J. Mol. Graph. Model. 2017, 77, 72–85. [Google Scholar] [CrossRef]
- Bergström, C.A.S.; Larsson, P. Computational Prediction of Drug Solubility in Water-Based Systems: Qualitative and Quantitative Approaches Used in the Current Drug Discovery and Development Setting. Int. J. Pharm. 2018, 540, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shalom, I.; Lin, C.; Radak, B.; Sherman, W.; Gilson, M. Fast Equilibration of Water between Buried Sites and Bulk by MD with Parallel Monte Carlo Water Moves on GPUs. ChemRxiv 2021. Available online: https://chemrxiv.org/engage/chemrxiv/article-details/6123cb101d1cc2684fcab40a (accessed on 16 October 2022).
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Tacconelli, E.; Magrini, N.; Kahlmeter, G.; Singh, N. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. World Health Organ. 2017, 27. Available online: https://www.thelancet.com/journals/laninf/article/PIIS1473-3099(17)30753-3/fulltext (accessed on 16 October 2022).
- Bijker, M.S.; Melief, C.J.M.; Offringa, R.; van der Burg, S.H. Design and Development of Synthetic Peptide Vaccines: Past, Present and Future. Expert Rev. Vaccines 2007, 6, 591–603. [Google Scholar] [CrossRef]
- Zhang, L. Multi-Epitope Vaccines: A Promising Strategy against Tumors and Viral Infections. Cell. Mol. Immunol. 2018, 15, 182. [Google Scholar] [CrossRef]
- Rappuoli, R. From Pasteur to Genomics: Progress and Challenges in Infectious Diseases. Nat. Med. 2004, 10, 1177. [Google Scholar] [CrossRef]
- del Tordello, E.; Rappuoli, R.; Delany, I. Reverse Vaccinology: Exploiting Genomes for Vaccine Design. In Human Vaccines; Elsevier: Amsterdam, The Netherlands, 2017; pp. 65–86. [Google Scholar]
- Shevtsov, A.; Syzdykov, M.; Kuznetsov, A.; Shustov, A.; Shevtsova, E.; Berdimuratova, K.; Mukanov, K.; Ramankulov, Y. Antimicrobial Susceptibility of Brucella Melitensis in Kazakhstan. Antimicrob. Resist Infect. Control. 2017, 6, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ehsan, N.; Ahmad, S.; Navid, A.; Azam, S.S. Identification of Potential Antibiotic Targets in the Proteome of Multi-Drug Resistant Proteus Mirabilis. Meta Gene 2018, 18, 167–173. [Google Scholar] [CrossRef]
- Gul, S.; Ahmad, S.; Ullah, A.; Ismail, S.; Khurram, M.; Tahir ul Qamar, M.; Hakami, A.R.; Alkhathami, A.G.; Alrumaihi, F.; Allemailem, K.S. Designing a Recombinant Vaccine against Providencia Rettgeri Using Immunoinformatics Approach. Vaccines 2022, 10, 189. [Google Scholar] [CrossRef] [PubMed]
- Munia, M.; Mahmud, S.; Mohasin, M.; Kibria, K.M.K. In Silico Design of an Epitope-Based Vaccine against Choline Binding Protein A of Streptococcus Pneumoniae. Inf. Med. Unlocked 2021, 23, 100546. [Google Scholar] [CrossRef]
- Zargaran, F.N.; Akya, A.; Rezaeian, S.; Ghadiri, K.; Lorestani, R.C.; Madanchi, H.; Safaei, S.; Rostamian, M. B Cell Epitopes of Four Fimbriae Antigens of Klebsiella Pneumoniae: A Comprehensive in Silico Study for Vaccine Development. Int. J. Pept. Res. Ther. 2021, 27, 875–886. [Google Scholar] [CrossRef]
- Li, M.; Zhu, Y.; Niu, C.; Xie, X.; Haimiti, G.; Guo, W.; Yu, M.; Chen, Z.; Ding, J.; Zhang, F. Design of a Multi-Epitope Vaccine Candidate against Brucella Melitensis. Sci. Rep. 2022, 12, 10146. [Google Scholar] [CrossRef]
- Yao, M.; Guo, X.; Wu, X.; Bai, Q.; Sun, M.; Yin, D. Evaluation of the Combined Use of Major Outer Membrane Proteins in the Serodiagnosis of Brucellosis. Infect. Drug Resist. 2022, 15, 4093–4100. [Google Scholar] [CrossRef] [PubMed]
- Kazi, A.; Chuah, C.; Majeed, A.B.A.; Leow, C.H.; Lim, B.H.; Leow, C.Y. Current Progress of Immunoinformatics Approach Harnessed for Cellular-and Antibody-Dependent Vaccine Design. Pathog Glob Health 2018, 112, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Khan, F.; Kumar, A. Exploring Highly Antigenic Protein of Campylobacter Jejuni for Designing Epitope Based Vaccine: Immunoinformatics Approach. Int. J. Pept. Res. Ther. 2019, 25, 1159–1172. [Google Scholar] [CrossRef]
- Sheth, H.B.; Glasier, L.M.; Ellert, N.W.; Cachia, P.; Kohn, W.; Lee, K.K.; Paranchych, W.; Hodges, R.S.; Irvin, R.T. Development of an Anti-Adhesive Vaccine for Pseudomonas Aeruginosa Targeting the C-Terminal Region of the Pilin Structural Protein. Biomed Pept. Proteins Nucleic Acids 1995, 1, 141–148. [Google Scholar] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).