
A Recombinant Chimeric Protein-Based Vaccine Containing T-Cell Epitopes from Amastigote Proteins and Combined with Distinct Adjuvants, Induces Immunogenicity and Protection against Leishmania infantum Infection

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Blood Samples
2.2. Parasite and Mice
2.3. Construction of ChimT Protein
2.4. Purification of Recombinant ChimT Protein
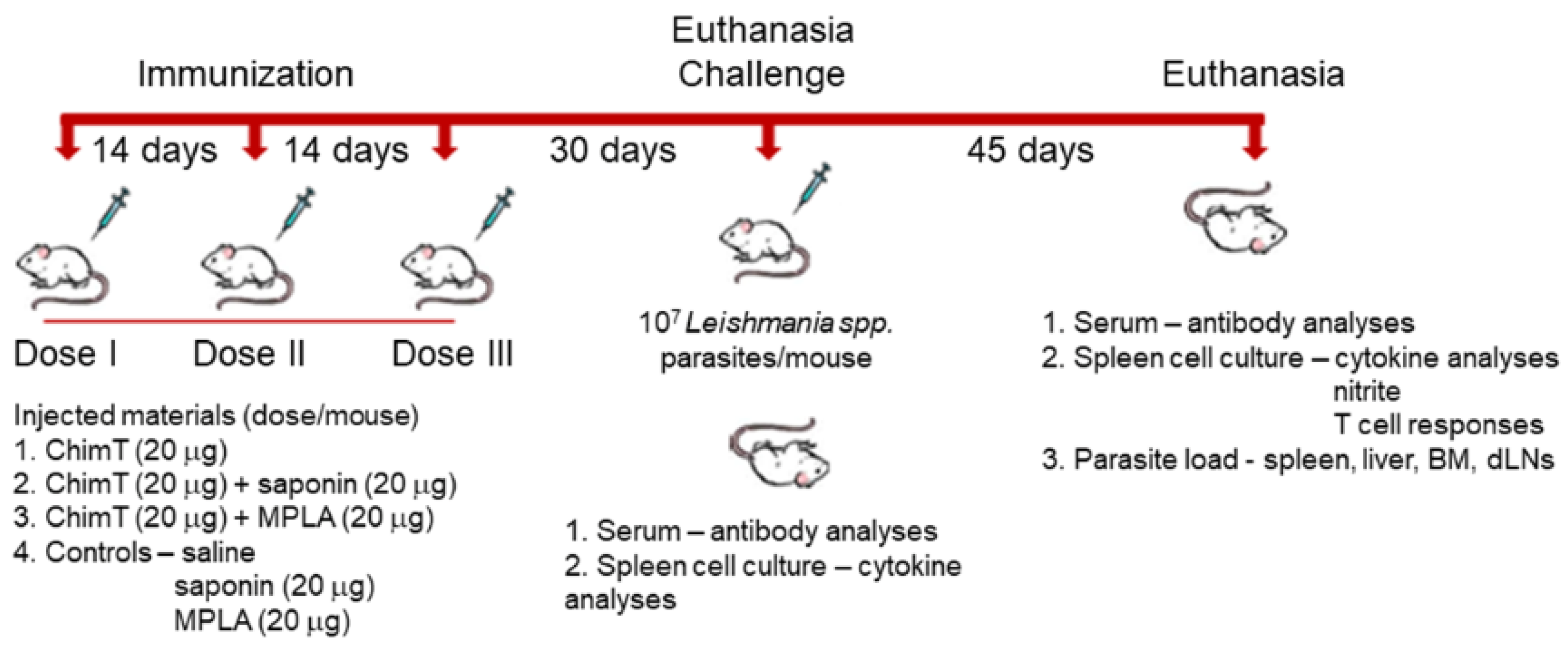
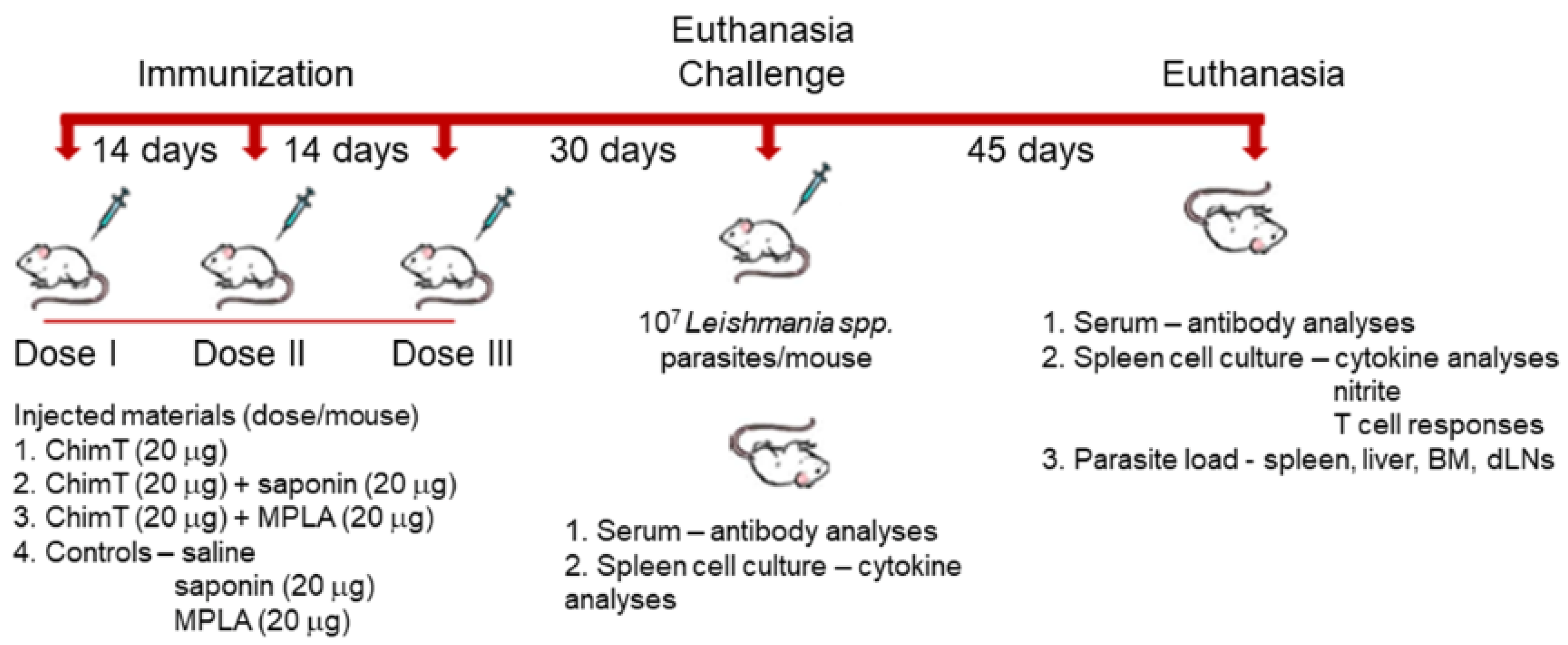
2.5. Mouse Immunization and Experimental Infection
2.6. Cellular Response
2.6.1. Cytokine and Nitrite Production and Evaluation of T-Cell Subtypes Producing IFN-γ
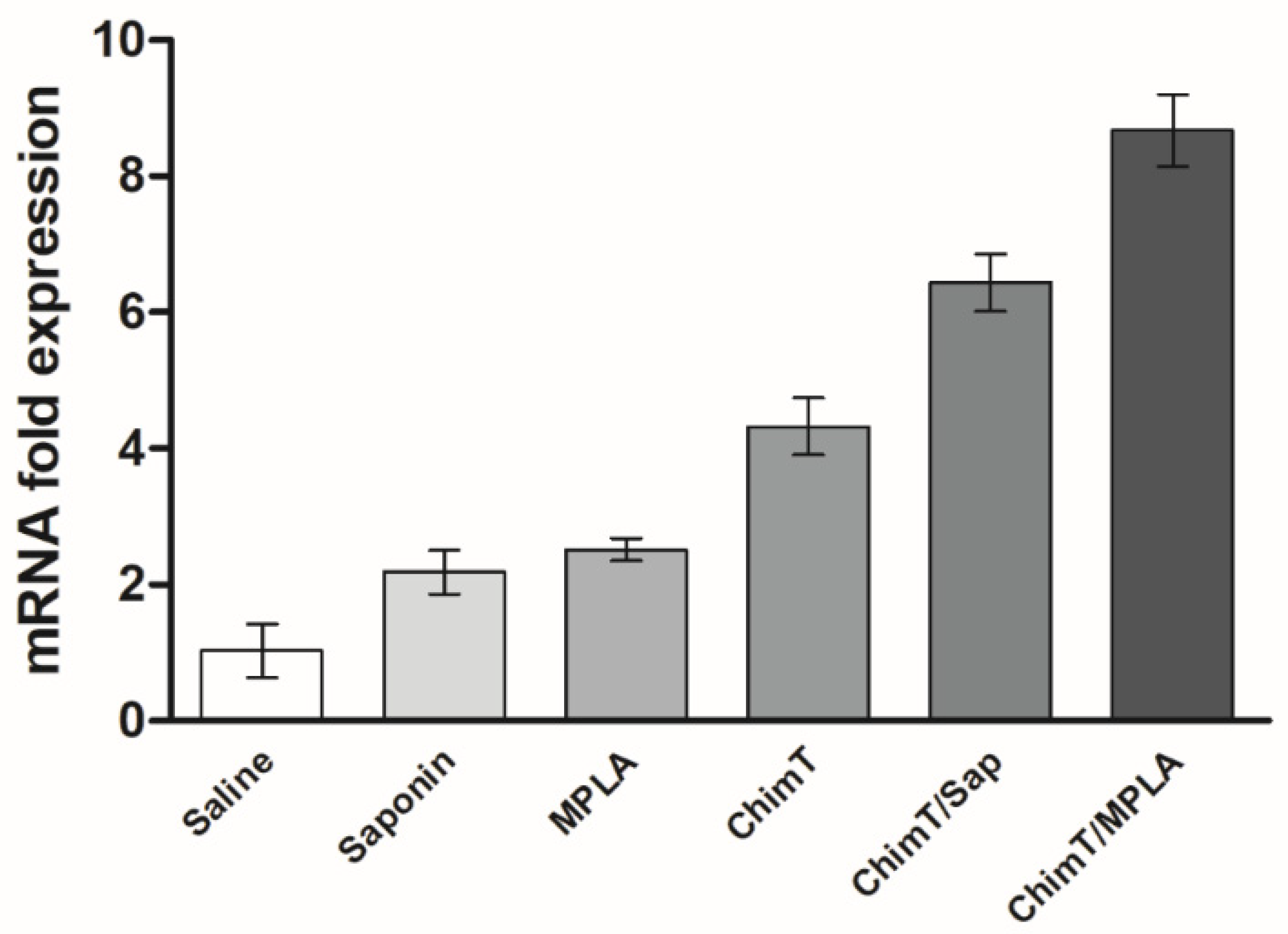
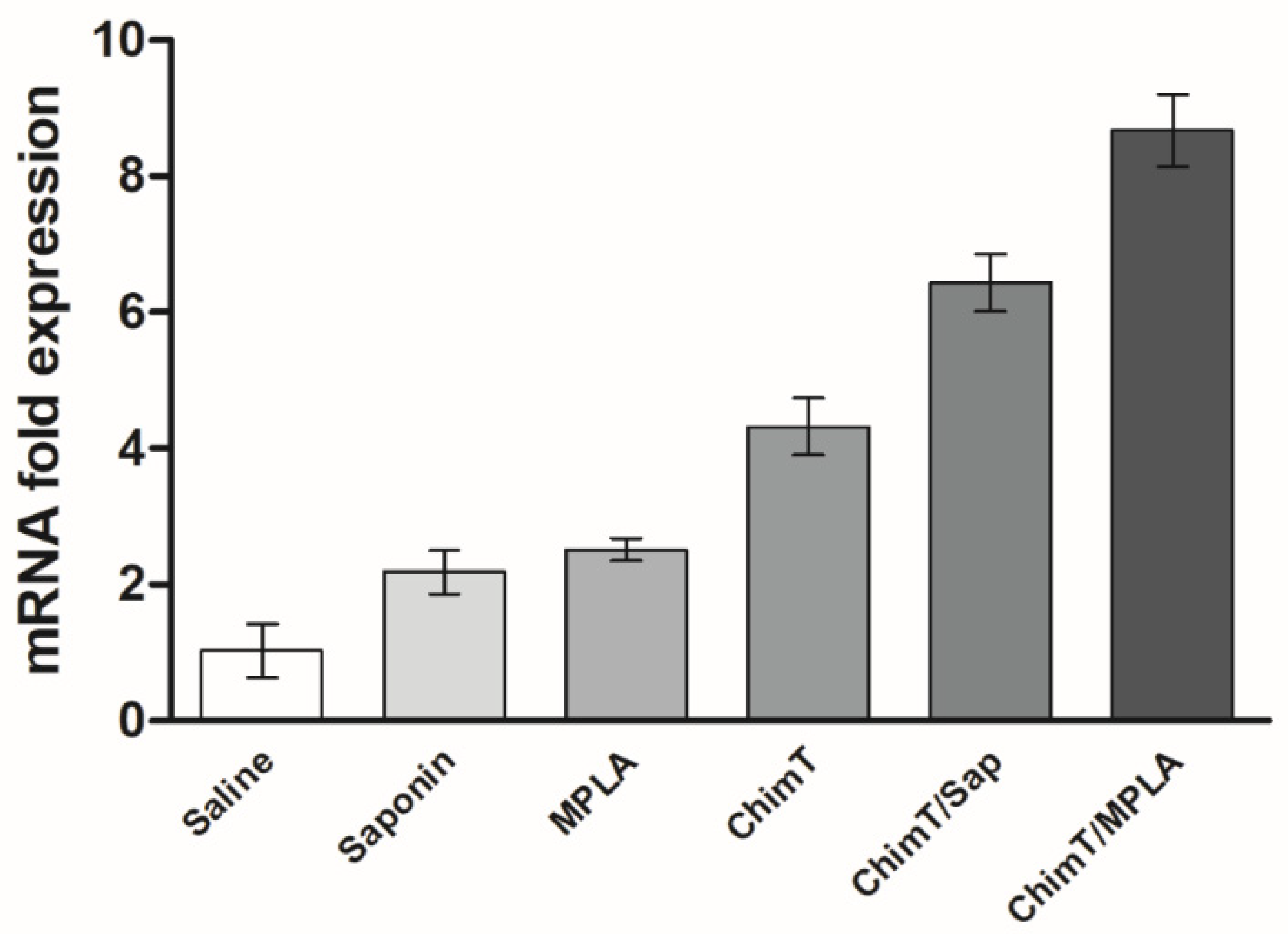
2.6.2. IFN-γ Expression in the Infected and Immunized Mice
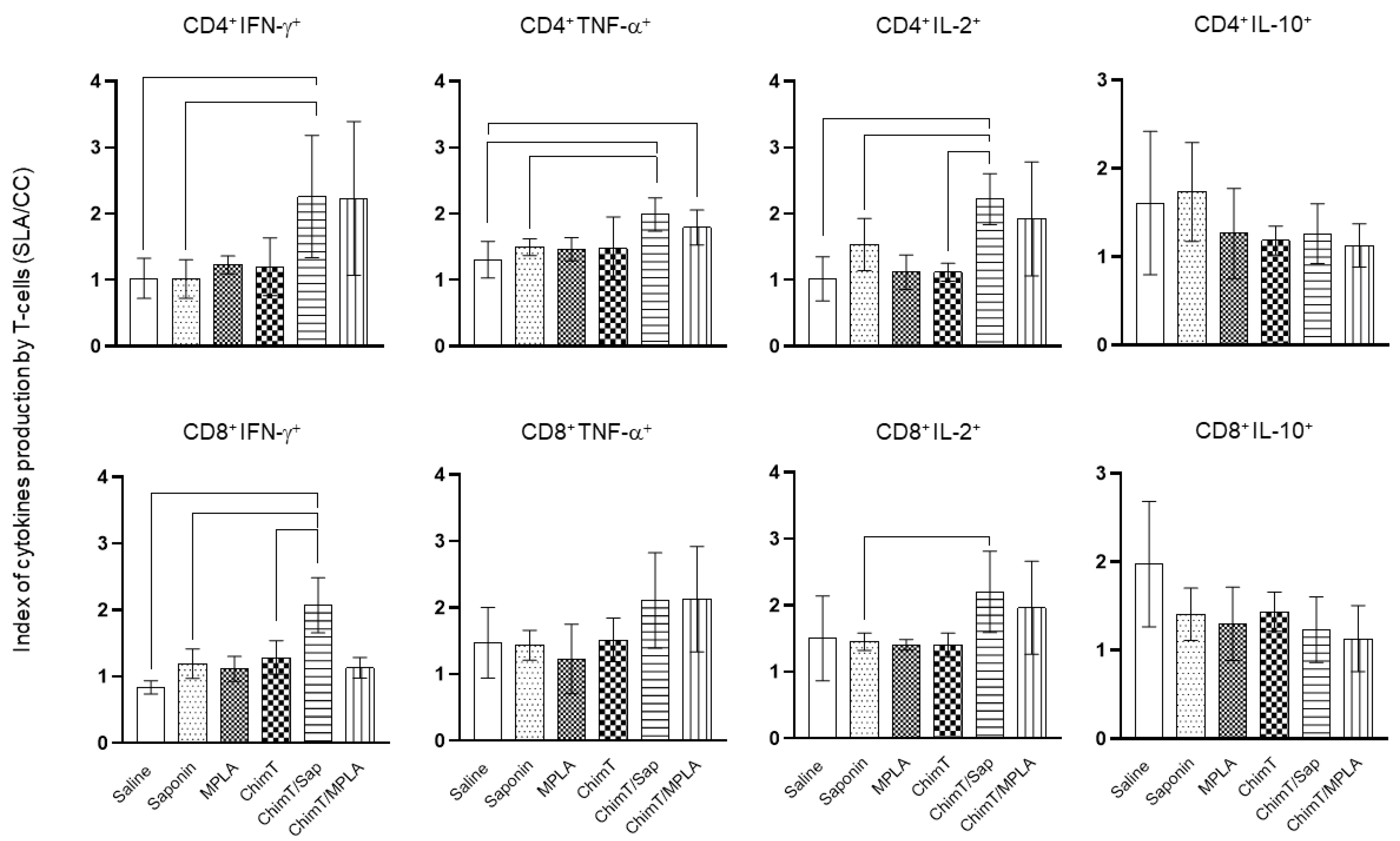
2.6.3. Polyfunctional T-Cell Analyses by Flow Cytometry
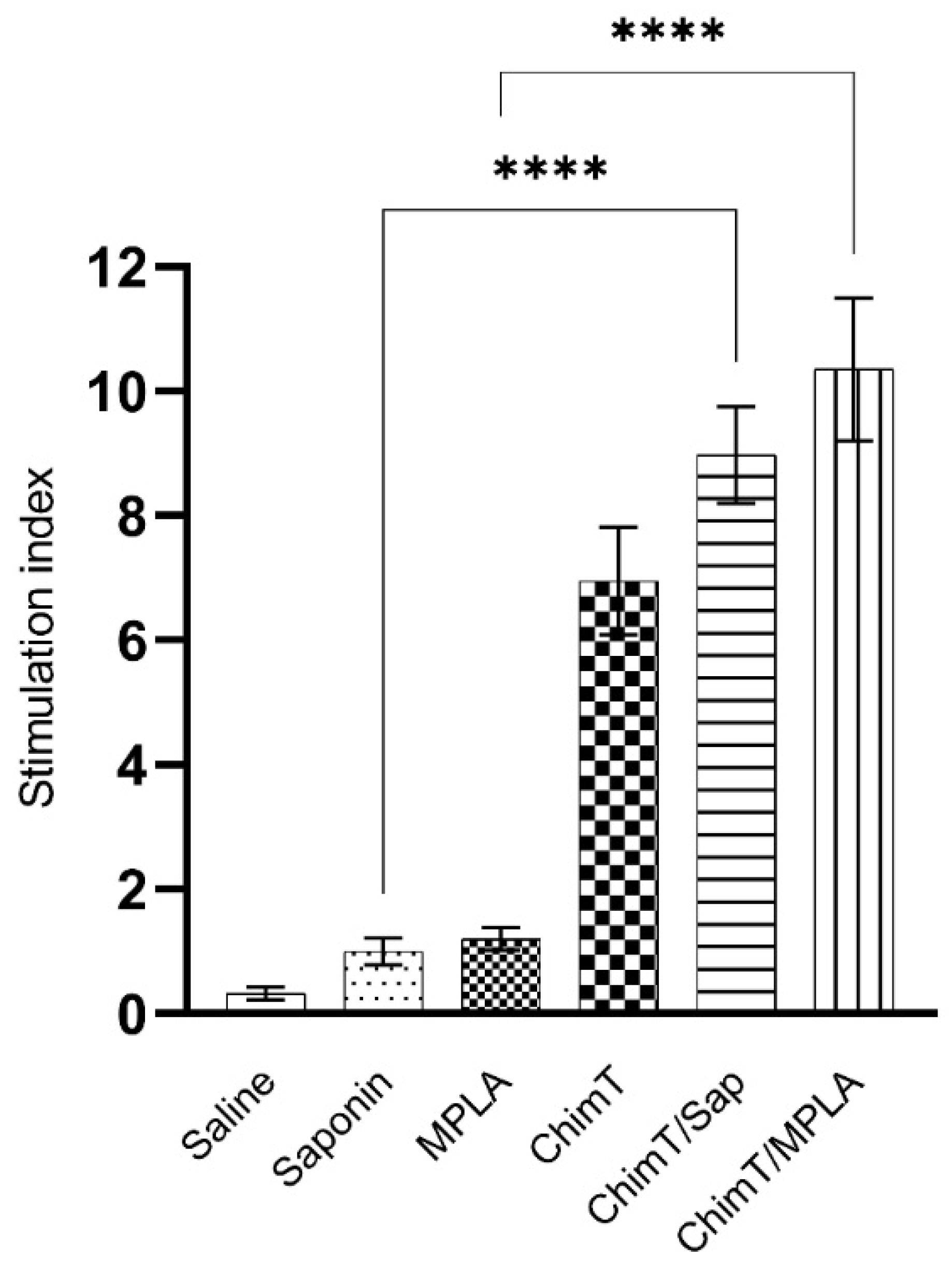
2.6.4. In Vitro Splenocyte Proliferation
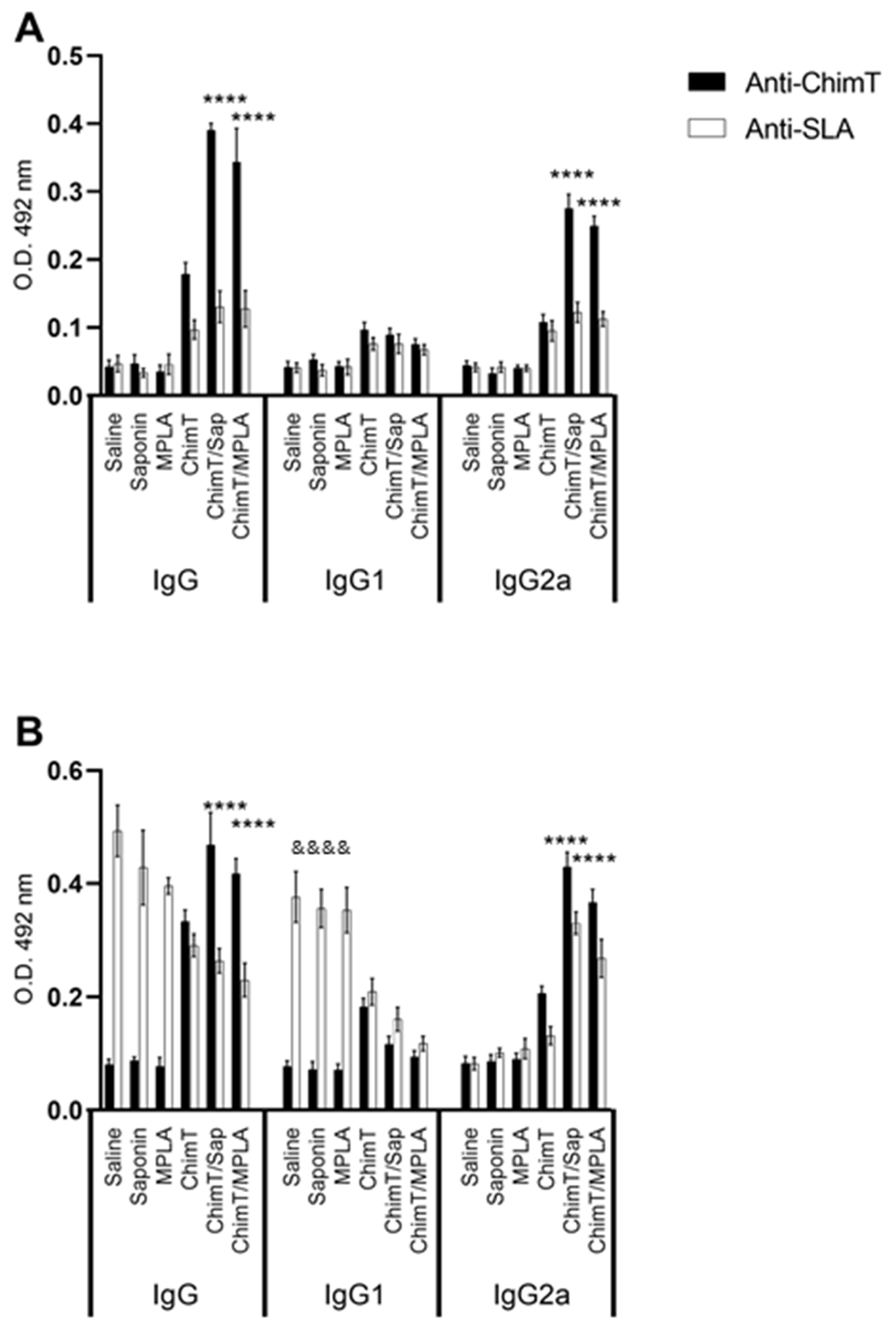
2.7. Analysis of IgG Production and Isotype Profile
2.8. Parasite Load
2.8.1. Limiting Dilution Technique
2.8.2. qPCR Assay
2.9. Immunogenicity Stimulation in Human Cells
2.10. Statistical Analysis
3. Results
3.1. Construction and Characterization of Recombinant Chimeric Protein, ChimT
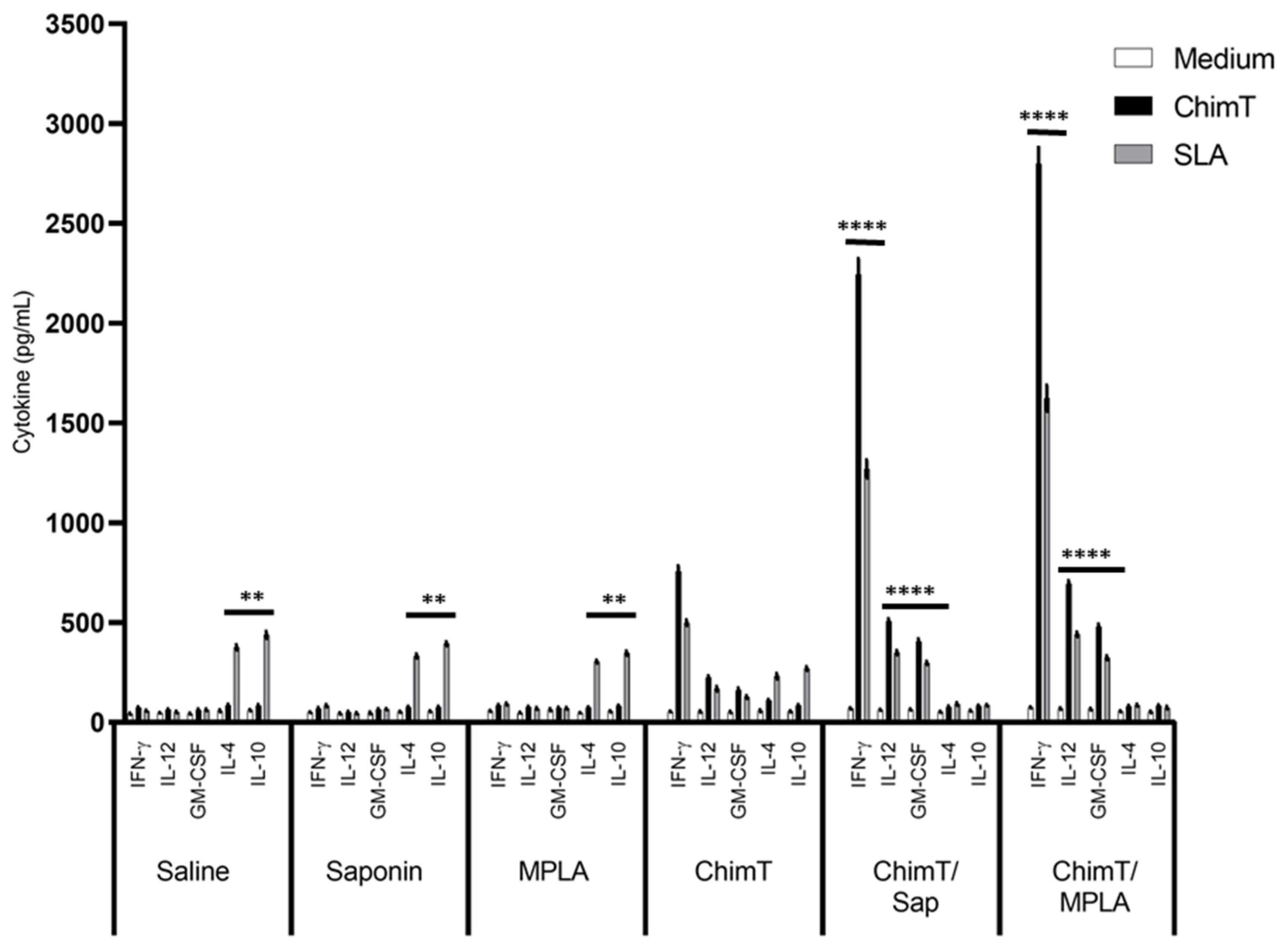
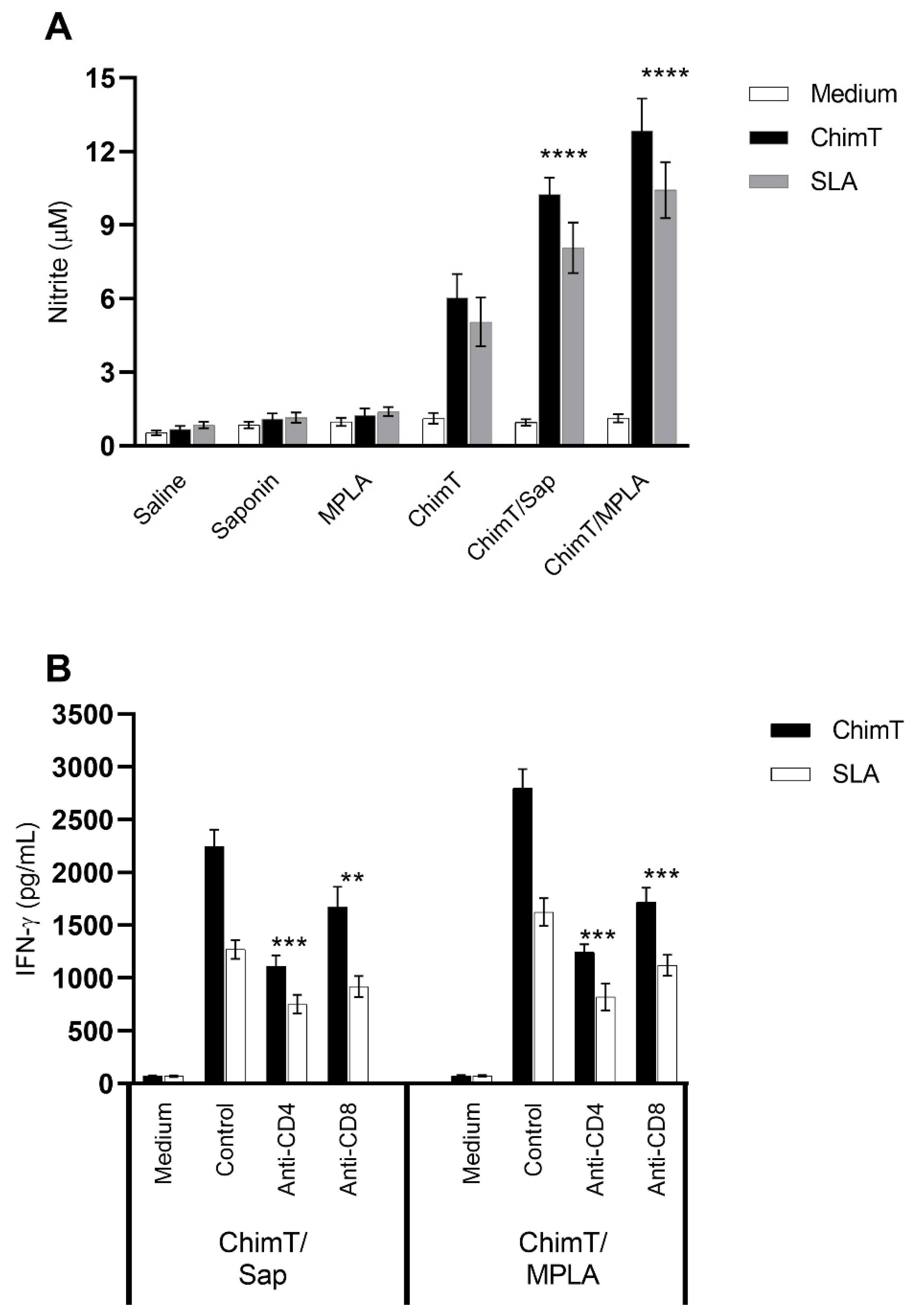
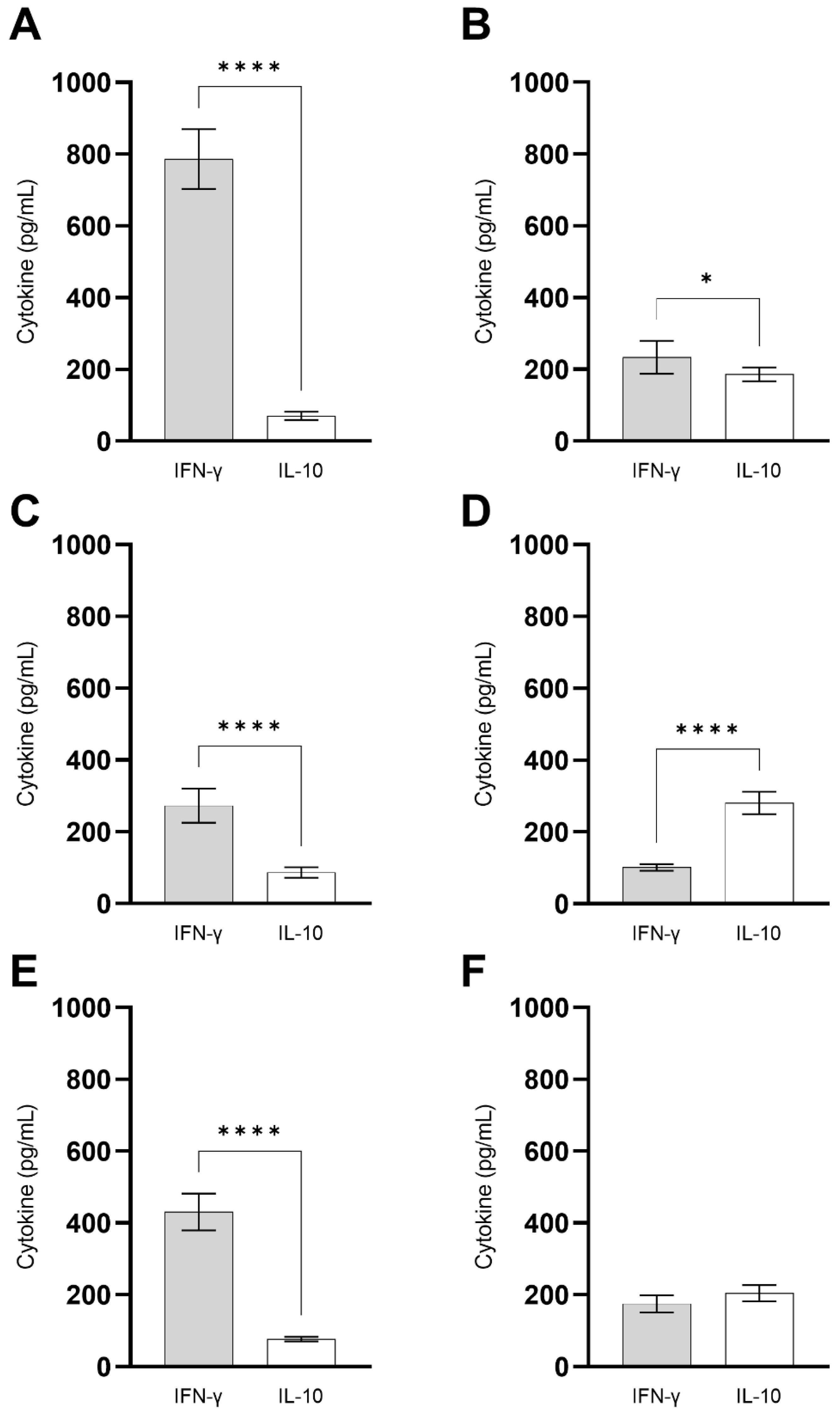
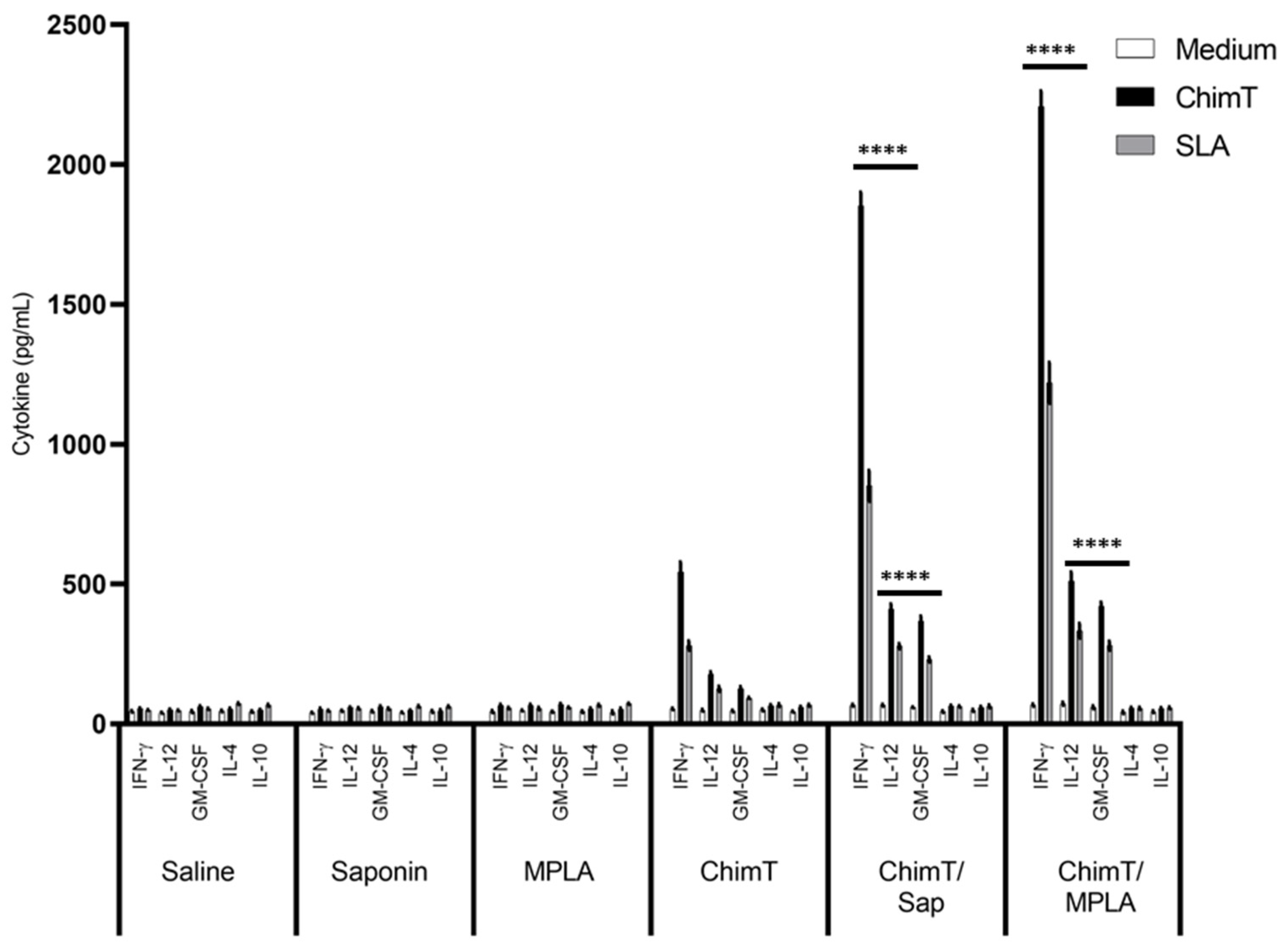
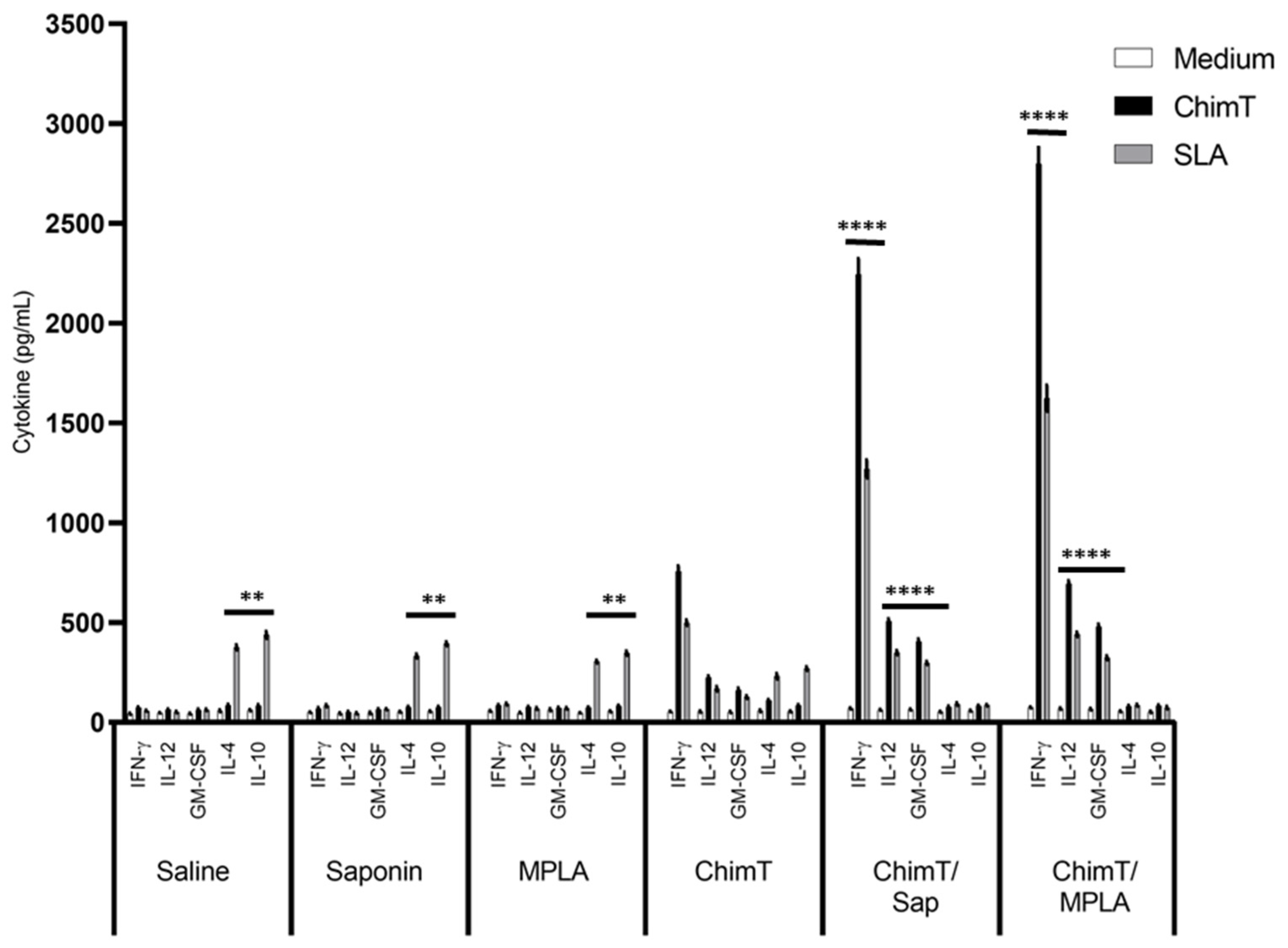
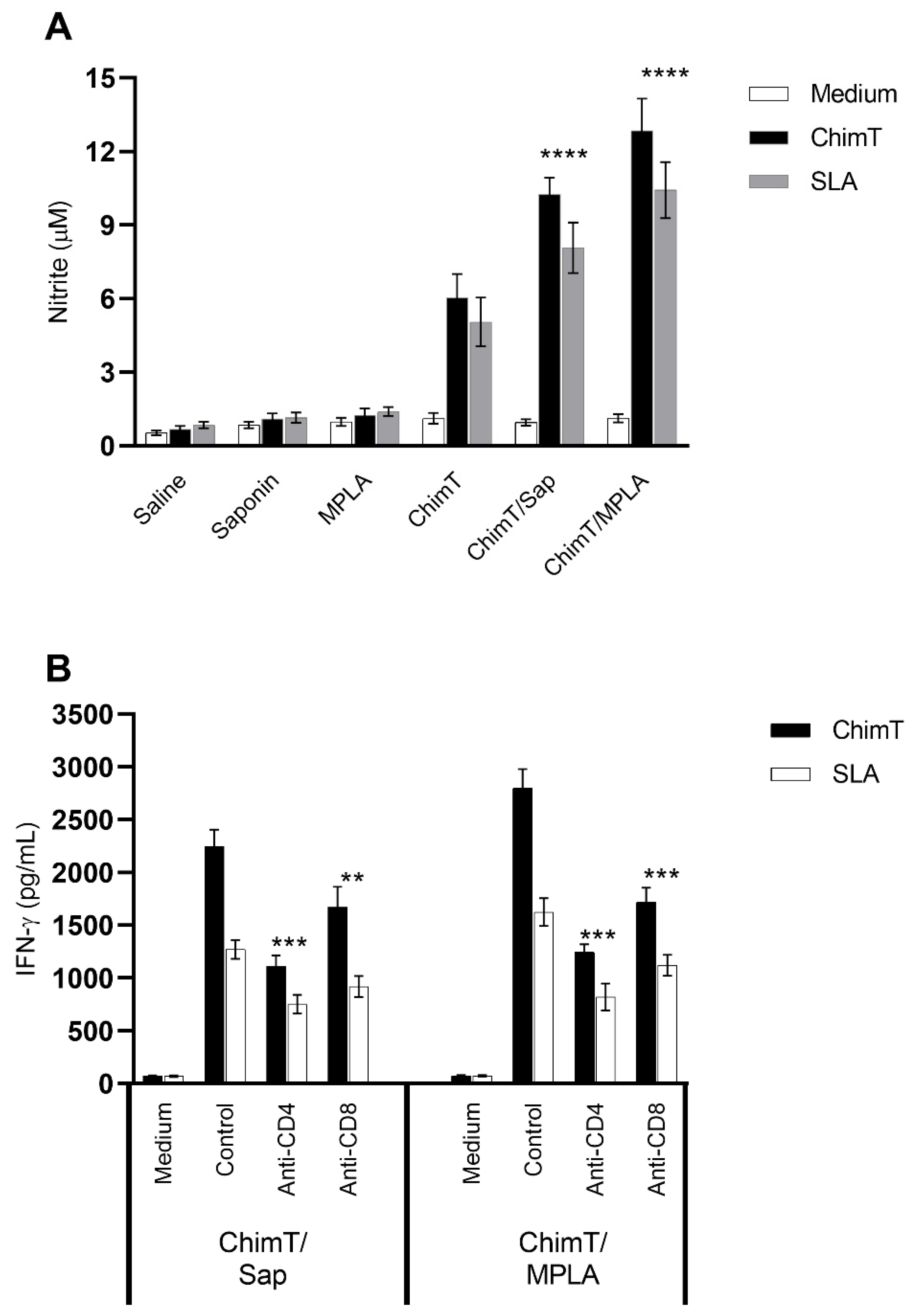
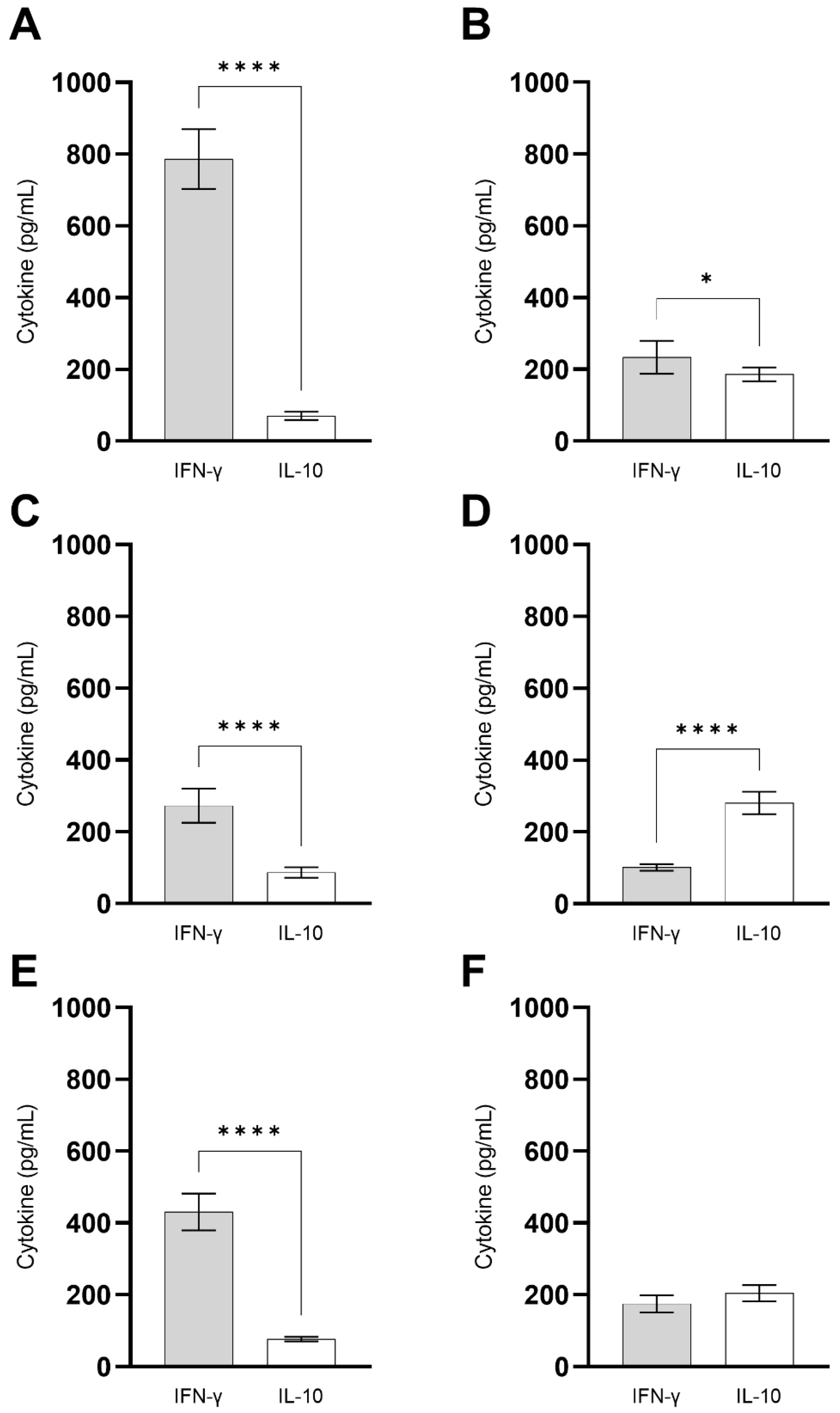
3.2. ChimT Plus Adjuvant Stimulates the Development of a Th1-Type Cellular Response before and after Infection
3.3. ChimT Plus Adjuvant Stimulates Specific IgG2a Isotype Production before and after Parasite Challenge
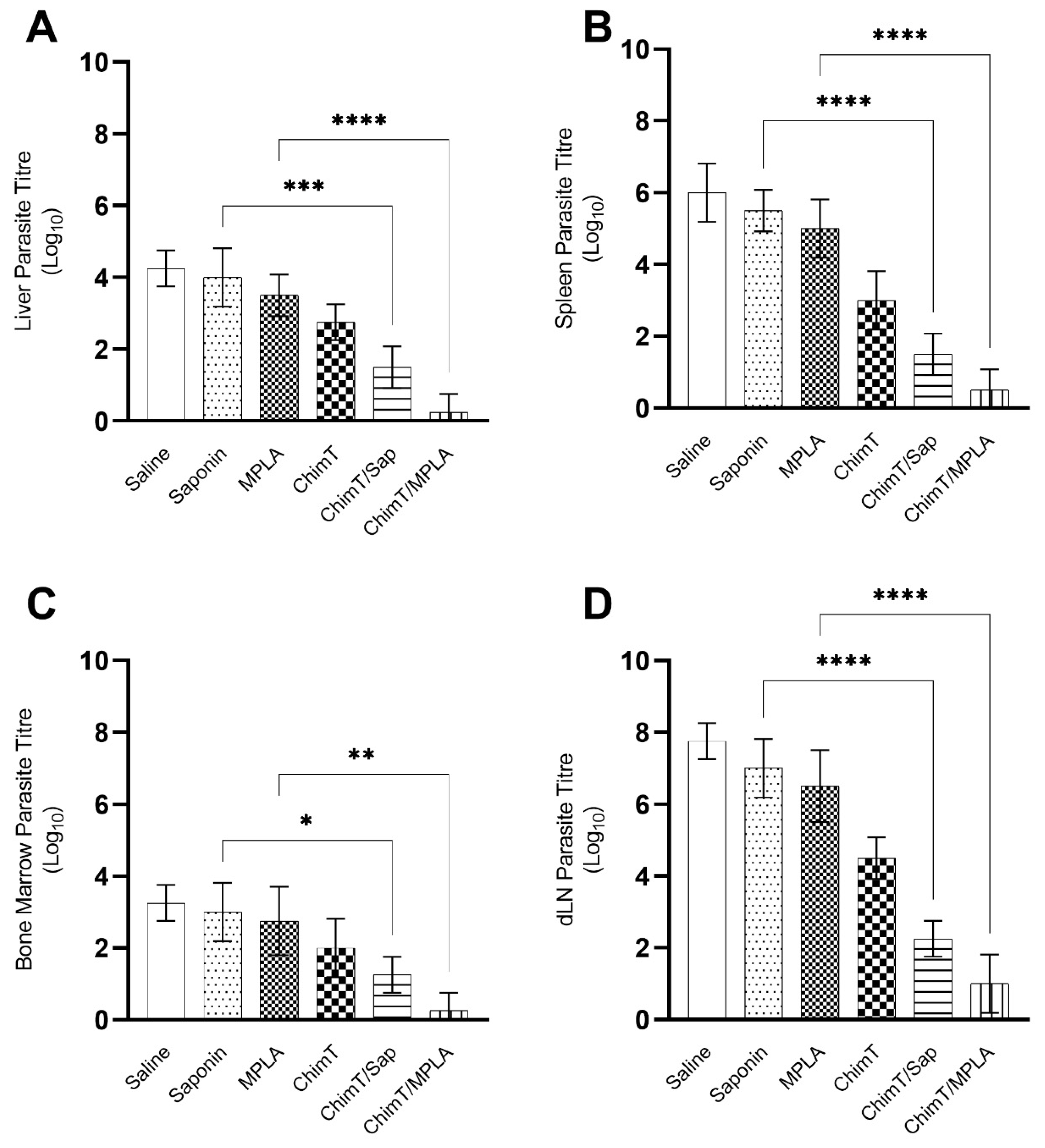
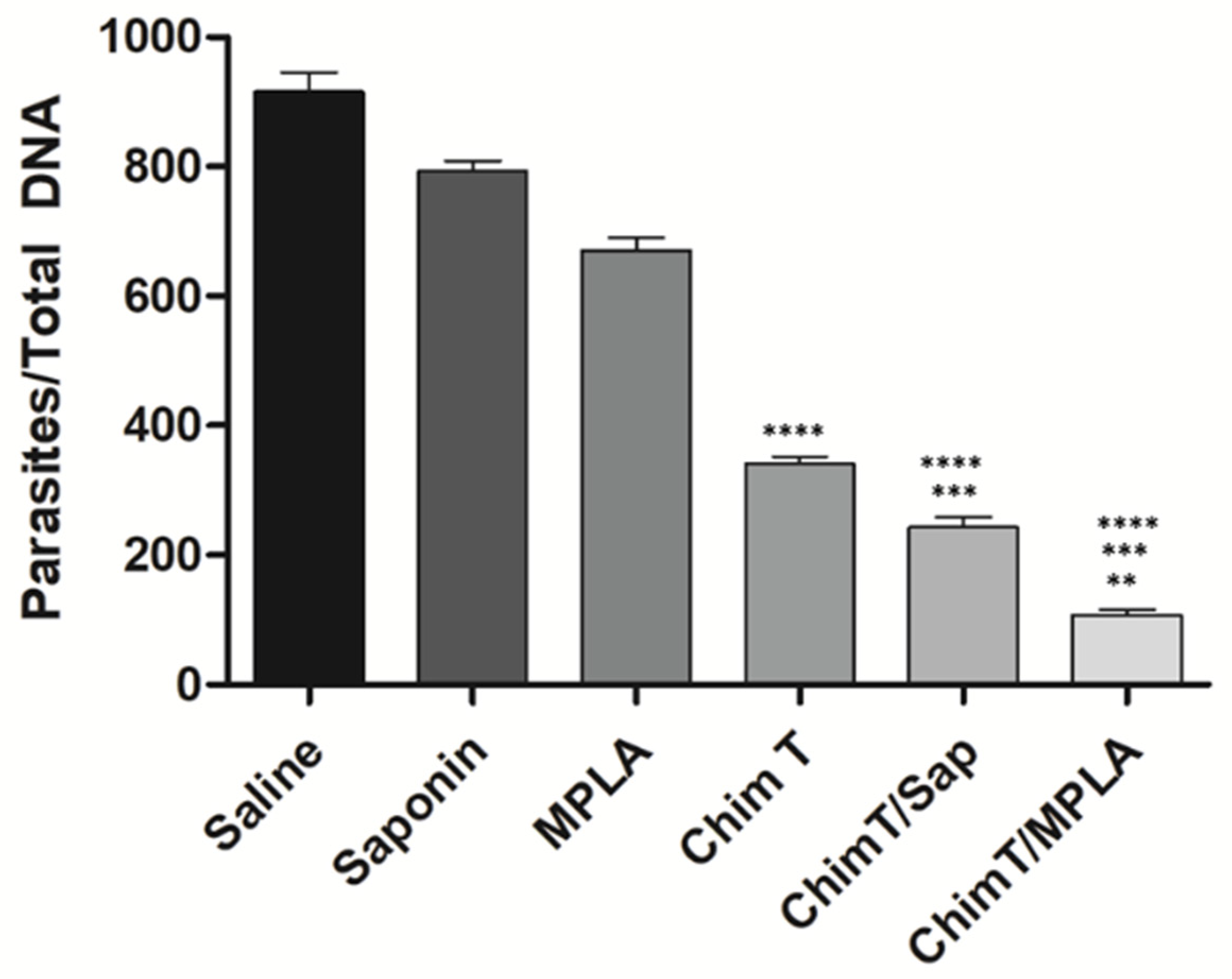
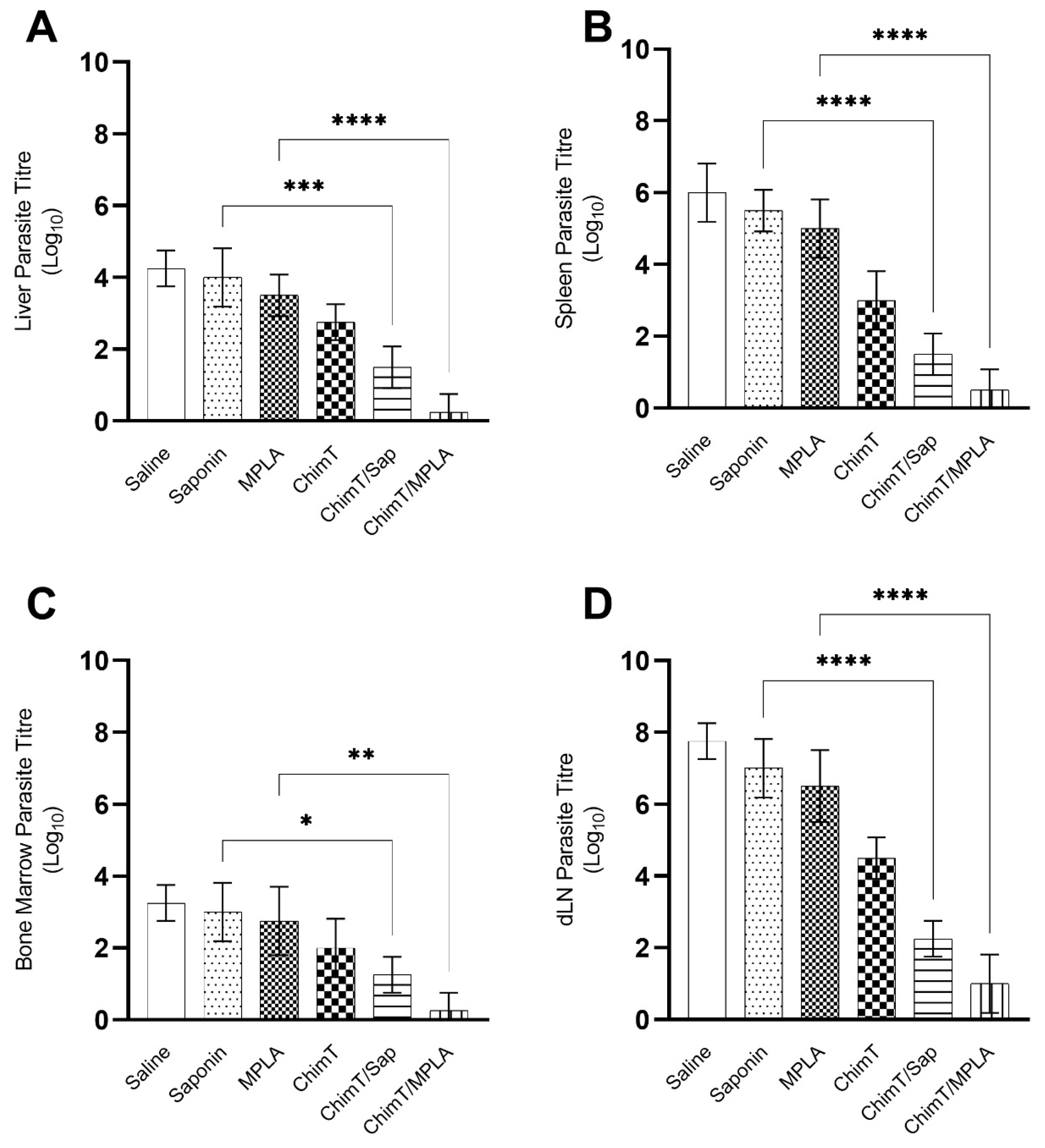
3.4. ChimT/Sap and ChimT/MPLA Protect Mice from Parasite Infection
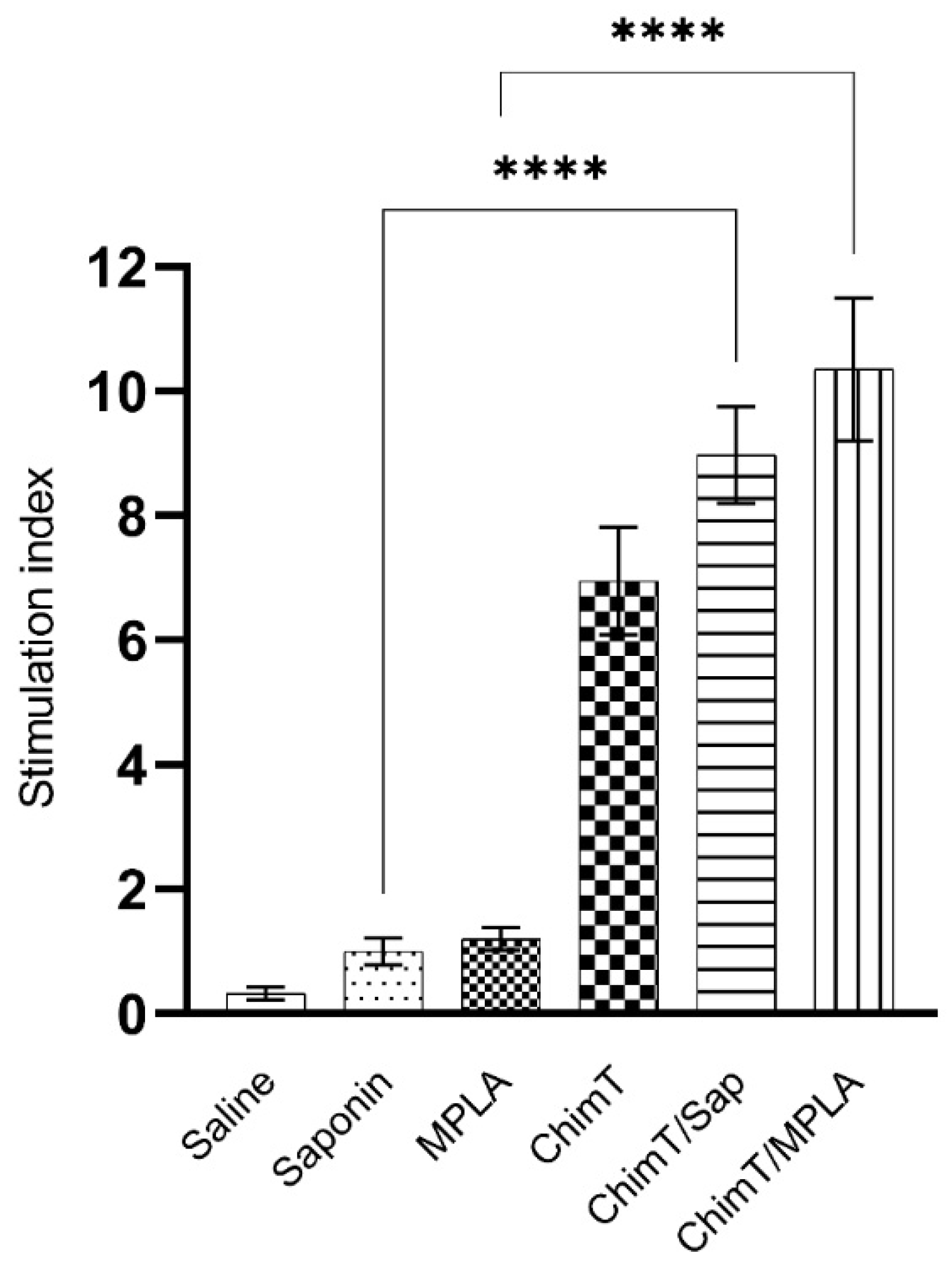
3.5. ChimT Stimulates IFN-γ Production from Human PBMCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Organisation, W.H. Leishmaniasis. Available online: http://www.who.int/topics/leishmaniasis/en/ (accessed on 1 July 2022).
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Res 2017, 6, 750. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Reithinger, R.; Dujardin, J.C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous leishmaniasis. Lancet Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Ready, P.D. Epidemiology of visceral leishmaniasis. Clin. Epidemiol. 2014, 6, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Stockdale, L.; Newton, R. A review of preventative methods against human leishmaniasis infection. PLoS Negl. Trop. Dis. 2013, 7, e2278. [Google Scholar] [CrossRef]
- Savoia, D. Recent updates and perspectives on leishmaniasis. J. Infect. Dev. Ctries 2015, 9, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Montenegro Quinonez, C.A.; Runge-Ranzinger, S.; Rahman, K.M.; Horstick, O. Effectiveness of vector control methods for the control of cutaneous and visceral leishmaniasis: A meta-review. PLoS Negl. Trop. Dis. 2021, 15, e0009309. [Google Scholar] [CrossRef]
- Rostami, M.N.; Khamesipour, A. Potential biomarkers of immune protection in human leishmaniasis. Med. Microbiol. Immunol. 2021, 210, 81–100. [Google Scholar] [CrossRef]
- Dubie, T.; Mohammed, Y. Review on the Role of Host Immune Response in Protection and Immunopathogenesis during Cutaneous Leishmaniasis Infection. J. Immunol. Res. 2020, 2020, 2496713. [Google Scholar] [CrossRef]
- Samant, M.; Sahu, U.; Pandey, S.C.; Khare, P. Role of Cytokines in Experimental and Human Visceral Leishmaniasis. Front. Cell Infect. Microbiol. 2021, 11, 624009. [Google Scholar] [CrossRef]
- Mirzaei, A.; Maleki, M.; Masoumi, E.; Maspi, N. A historical review of the role of cytokines involved in leishmaniasis. Cytokine 2021, 145, 155297. [Google Scholar] [CrossRef] [PubMed]
- Guha, R.; Das, S.; Ghosh, J.; Naskar, K.; Mandala, A.; Sundar, S.; Dujardin, J.C.; Roy, S. Heterologous priming-boosting with DNA and vaccinia virus expressing kinetoplastid membrane protein-11 induces potent cellular immune response and confers protection against infection with antimony resistant and sensitive strains of Leishmania (Leishmania) donovani. Vaccine 2013, 31, 1905–1915. [Google Scholar] [CrossRef] [PubMed]
- Foroughi-Parvar, F.; Hatam, G.R.; Sarkari, B.; Kamali-Sarvestani, E. Leishmania infantum FML pulsed-dendritic cells induce a protective immune response in murine visceral leishmaniasis. Immunotherapy 2015, 7, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Amit, A.; Vijayamahantesh; Dikhit, M.R.; Singh, A.K.; Kumar, V.; Suman, S.S.; Singh, A.; Kumar, A.; Thakur, A.K.; Das, V.R.; et al. Immunization with Leishmania donovani protein disulfide isomerase DNA construct induces Th1 and Th17 dependent immune response and protection against experimental visceral leishmaniasis in Balb/c mice. Mol. Immunol. 2017, 82, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Helou, D.G.; Mauras, A.; Fasquelle, F.; Lanza, J.S.; Loiseau, P.M.; Betbeder, D.; Cojean, S. Intranasal vaccine from whole Leishmania donovani antigens provides protection and induces specific immune response against visceral leishmaniasis. PLoS Negl. Trop. Dis. 2021, 15, e0009627. [Google Scholar] [CrossRef]
- Martinez-Florez, A.; Martori, C.; Monteagudo, P.L.; Rodriguez, F.; Alberola, J.; Rodriguez-Cortes, A. Sirolimus enhances the protection achieved by a DNA vaccine against Leishmania infantum. Parasit Vectors 2020, 13, 294. [Google Scholar] [CrossRef]
- Oliveira-da-Silva, J.A.; Machado, A.S.; Ramos, F.F.; Tavares, G.S.V.; Lage, D.P.; Mendonca, D.V.C.; Pereira, I.A.G.; Santos, T.T.O.; Martins, V.T.; Carvalho, L.M.; et al. A Leishmania amastigote-specific hypothetical protein evaluated as recombinant protein plus Th1 adjuvant or DNA plasmid-based vaccine to protect against visceral leishmaniasis. Cell Immunol. 2020, 356, 104194. [Google Scholar] [CrossRef]
- Ribeiro, P.A.F.; Vale, D.L.; Dias, D.S.; Lage, D.P.; Mendonca, D.V.C.; Ramos, F.F.; Carvalho, L.M.; Carvalho, A.; Steiner, B.T.; Roque, M.C.; et al. Leishmania infantum amastin protein incorporated in distinct adjuvant systems induces protection against visceral leishmaniasis. Cytokine 2020, 129, 155031. [Google Scholar] [CrossRef]
- Yadav, S.; Prakash, J.; Singh, O.P.; Gedda, M.R.; Chauhan, S.B.; Sundar, S.; Dubey, V.K. IFN-gamma(+) CD4(+)T cell-driven prophylactic potential of recombinant LDBPK_252400 hypothetical protein of Leishmania donovani against visceral leishmaniasis. Cell Immunol. 2021, 361, 104272. [Google Scholar] [CrossRef]
- Brito, R.C.F.; Ruiz, J.C.; Cardoso, J.M.O.; Ostolin, T.; Reis, L.E.S.; Mathias, F.A.S.; Aguiar-Soares, R.D.O.; Roatt, B.M.; Correa-Oliveira, R.; Resende, D.M.; et al. Chimeric Vaccines Designed by Immunoinformatics-Activated Polyfunctional and Memory T Cells That Trigger Protection against Experimental Visceral Leishmaniasis. Vaccines 2020, 8, 252. [Google Scholar] [CrossRef]
- Ostolin, T.; Gusmao, M.R.; Mathias, F.A.S.; Cardoso, J.M.O.; Roatt, B.M.; Aguiar-Soares, R.D.O.; Ruiz, J.C.; Resende, D.M.; de Brito, R.C.F.; Reis, A.B. A chimeric vaccine combined with adjuvant system induces immunogenicity and protection against visceral leishmaniasis in BALB/c mice. Vaccine 2021, 39, 2755–2763. [Google Scholar] [CrossRef] [PubMed]
- Athanasiou, E.; Agallou, M.; Tastsoglou, S.; Kammona, O.; Hatzigeorgiou, A.; Kiparissides, C.; Karagouni, E. A Poly(Lactic-co-Glycolic) Acid Nanovaccine Based on Chimeric Peptides from Different Leishmania infantum Proteins Induces Dendritic Cells Maturation and Promotes Peptide-Specific IFNgamma-Producing CD8(+) T Cells Essential for the Protection against Experimental Visceral Leishmaniasis. Front. Immunol. 2017, 8, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, D.S.; Ribeiro, P.A.F.; Martins, V.T.; Lage, D.P.; Costa, L.E.; Chavez-Fumagalli, M.A.; Ramos, F.F.; Santos, T.T.O.; Ludolf, F.; Oliveira, J.S.; et al. Vaccination with a CD4(+) and CD8(+) T-cell epitopes-based recombinant chimeric protein derived from Leishmania infantum proteins confers protective immunity against visceral leishmaniasis. Transl. Res. 2018, 200, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Lage, D.P.; Ribeiro, P.A.F.; Dias, D.S.; Mendonca, D.V.C.; Ramos, F.F.; Carvalho, L.M.; de Oliveira, D.; Steiner, B.T.; Martins, V.T.; Perin, L.; et al. A candidate vaccine for human visceral leishmaniasis based on a specific T cell epitope-containing chimeric protein protects mice against Leishmania infantum infection. NPJ Vaccines 2020, 5, 75. [Google Scholar] [CrossRef]
- Martins, V.T.; Duarte, M.C.; Lage, D.P.; Costa, L.E.; Carvalho, A.M.; Mendes, T.A.; Roatt, B.M.; Menezes-Souza, D.; Soto, M.; Coelho, E.A. A recombinant chimeric protein composed of human and mice-specific CD4(+) and CD8(+) T-cell epitopes protects against visceral leishmaniasis. Parasite Immunol. 2017, 39, e12359. [Google Scholar] [CrossRef]
- Ashwin, H.; Sadlova, J.; Vojtkova, B.; Becvar, T.; Lypaczewski, P.; Schwartz, E.; Greensted, E.; Van Bocxlaer, K.; Pasin, M.; Lipinski, K.S.; et al. Characterization of a new Leishmania major strain for use in a controlled human infection model. Nat. Commun. 2021, 12, 215. [Google Scholar] [CrossRef]
- Martins, V.T.; Machado, A.S.; Humbert, M.V.; Christodoulides, M.; Coelho, E.A.F. Preclinical Assessment of the Immunogenicity of Experimental Leishmania Vaccines. Methods Mol. Biol. 2022, 2410, 481–502. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Coelho, E.A.; Machado-Coelho, G.L.; Grimaldi, G., Jr.; Gazzinelli, R.T. Making an anti-amastigote vaccine for visceral leishmaniasis: Rational, update and perspectives. Curr. Opin. Microbiol. 2012, 15, 476–485. [Google Scholar] [CrossRef]
- Machado, A.S.; Lage, D.P.; Vale, D.L.; Freitas, C.S.; Linhares, F.P.; Cardoso, J.M.O.; Pereira, I.A.G.; Ramos, F.F.; Tavares, G.S.V.; Ludolf, F.; et al. A recombinant Leishmania amastigote-specific protein, rLiHyG, with adjuvants, protects against infection with Leishmania infantum. Acta Trop. 2022, 230, 106412. [Google Scholar] [CrossRef]
- Coelho, E.A.; Tavares, C.A.; Carvalho, F.A.; Chaves, K.F.; Teixeira, K.N.; Rodrigues, R.C.; Charest, H.; Matlashewski, G.; Gazzinelli, R.T.; Fernandes, A.P. Immune responses induced by the Leishmania (Leishmania) donovani A2 antigen, but not by the LACK antigen, are protective against experimental Leishmania (Leishmania) amazonensis infection. Infect. Immun. 2003, 71, 3988–3994. [Google Scholar] [CrossRef] [Green Version]
- Reche, P.A.; Reinherz, E.L. Prediction of peptide-MHC binding using profiles. Methods Mol. Biol. 2007, 409, 185–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malvolti, S.; Malhame, M.; Mantel, C.F.; Le Rutte, E.A.; Kaye, P.M. Human leishmaniasis vaccines: Use cases, target population and potential global demand. PLoS Negl. Trop. Dis. 2021, 15, e0009742. [Google Scholar] [CrossRef] [PubMed]
- Jain, K.; Jain, N.K. Vaccines for visceral leishmaniasis: A review. J. Immunol. Methods 2015, 422, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R. Stage-Specific Class I Nucleases of Leishmania Play Important Roles in Parasite Infection and Survival. Front. Cell Infect. Microbiol. 2021, 11, 769933. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Sarkar, R.R. Perspectives on Leishmania Species and Stage-specific Adaptive Mechanisms. Trends Parasitol. 2018, 34, 1068–1081. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.T.; Chavez-Fumagalli, M.A.; Costa, L.E.; Canavaci, A.M.; Martins, A.M.; Lage, P.S.; Lage, D.P.; Duarte, M.C.; Valadares, D.G.; Magalhaes, R.D.; et al. Antigenicity and protective efficacy of a Leishmania amastigote-specific protein, member of the super-oxygenase family, against visceral leishmaniasis. PLoS Negl. Trop. Dis. 2013, 7, e2148. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.T.; Chavez-Fumagalli, M.A.; Lage, D.P.; Duarte, M.C.; Garde, E.; Costa, L.E.; da Silva, V.G.; Oliveira, J.S.; Magalhaes-Soares, D.F.; Teixeira, S.M.; et al. Antigenicity, Immunogenicity and Protective Efficacy of Three Proteins Expressed in the Promastigote and Amastigote Stages of Leishmania infantum against Visceral Leishmaniasis. PLoS ONE 2015, 10, e0137683. [Google Scholar] [CrossRef]
- Machado, A.S.; Lage, D.P.; Vale, D.L.; Freitas, C.S.; Linhares, F.P.; Cardoso, J.M.O.; Oliveira-da-Silva, J.A.; Pereira, I.A.G.; Ramos, F.F.; Tavares, G.S.V.; et al. Leishmania LiHyC protein is immunogenic and induces protection against visceral leishmaniasis. Parasite Immunol. 2022, 230, e12921. [Google Scholar] [CrossRef]
- Poot, J.; Janssen, L.H.; van Kasteren-Westerneng, T.J.; van der Heijden-Liefkens, K.H.; Schijns, V.E.; Heckeroth, A. Vaccination of dogs with six different candidate leishmaniasis vaccines composed of a chimerical recombinant protein containing ribosomal and histone protein epitopes in combination with different adjuvants. Vaccine 2009, 27, 4439–4446. [Google Scholar] [CrossRef]
- Rodrigues, V.; Cordeiro-da-Silva, A.; Laforge, M.; Silvestre, R.; Estaquier, J. Regulation of immunity during visceral Leishmania infection. Parasit Vectors 2016, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Ratnapriya, S.; Keerti; Sahasrabuddhe, A.A.; Dube, A. Visceral leishmaniasis: An overview of vaccine adjuvants and their applications. Vaccine 2019, 37, 3505–3519. [Google Scholar] [CrossRef] [PubMed]
- Askarizadeh, A.; Badiee, A.; Khamesipour, A. Development of nano-carriers for Leishmania vaccine delivery. Expert Opin. Drug Deliv. 2020, 17, 167–187. [Google Scholar] [CrossRef]
- Badiee, A.; Heravi Shargh, V.; Khamesipour, A.; Jaafari, M.R. Micro/nanoparticle adjuvants for antileishmanial vaccines: Present and future trends. Vaccine 2013, 31, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Askarizadeh, A.; Jaafari, M.R.; Khamesipour, A.; Badiee, A. Liposomal adjuvant development for leishmaniasis vaccines. Ther. Adv. Vaccines 2017, 5, 85–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palatnik-de-Sousa, C.B.; Barbosa Ade, F.; Oliveira, S.M.; Nico, D.; Bernardo, R.R.; Santos, W.R.; Rodrigues, M.M.; Soares, I.; Borja-Cabrera, G.P. FML vaccine against canine visceral leishmaniasis: From second-generation to synthetic vaccine. Expert Rev. Vaccines 2008, 7, 833–851. [Google Scholar] [CrossRef]
- Gorman, M.J.; Patel, N.; Guebre-Xabier, M.; Zhu, A.L.; Atyeo, C.; Pullen, K.M.; Loos, C.; Goez-Gazi, Y.; Carrion, R., Jr.; Tian, J.H.; et al. Fab and Fc contribute to maximal protection against SARS-CoV-2 following NVX-CoV2373 subunit vaccine with Matrix-M vaccination. Cell Rep. Med. 2021, 2, 100405. [Google Scholar] [CrossRef]
- Kaur, T.; Thakur, A.; Kaur, S. Protective immunity using MPL-A and autoclaved Leishmania donovani as adjuvants along with a cocktail vaccine in murine model of visceral leishmaniasis. J. Parasit. Dis. 2013, 37, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Margaroni, M.; Agallou, M.; Athanasiou, E.; Kammona, O.; Kiparissides, C.; Gaitanaki, C.; Karagouni, E. Vaccination with poly(D,L-lactide-co-glycolide) nanoparticles loaded with soluble Leishmania antigens and modified with a TNFalpha-mimicking peptide or monophosphoryl lipid A confers protection against experimental visceral leishmaniasis. Int. J. Nanomed. 2017, 12, 6169–6184. [Google Scholar] [CrossRef] [Green Version]
- Margaroni, M.; Agallou, M.; Kontonikola, K.; Karidi, K.; Kammona, O.; Kiparissides, C.; Gaitanaki, C.; Karagouni, E. PLGA nanoparticles modified with a TNFalpha mimicking peptide, soluble Leishmania antigens and MPLA induce T cell priming in vitro via dendritic cell functional differentiation. Eur. J. Pharm. Biopharm. 2016, 105, 18–31. [Google Scholar] [CrossRef]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. Microbiol. 2011, 9, 604–615. [Google Scholar] [CrossRef]
- Kaushal, H.; Bras-Goncalves, R.; Negi, N.S.; Lemesre, J.L.; Papierok, G.; Salotra, P. Role of CD8(+) T cells in protection against Leishmania donovani infection in healed Visceral Leishmaniasis individuals. BMC Infect. Dis. 2014, 14, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, L.S.; Barreto, A.S.; Bomfim, L.G.S.; Gomes, M.C.; Ferreira, N.L.C.; da Cruz, G.S.; Magalhaes, L.S.; de Jesus, A.R.; Palatnik-de-Sousa, C.B.; Correa, C.B.; et al. Multifunctional, TNF-alpha and IFN-gamma-Secreting CD4 and CD8 T Cells and CD8(High) T Cells Are Associated With the Cure of Human Visceral Leishmaniasis. Front. Immunol. 2021, 12, 773983. [Google Scholar] [CrossRef]
- Kumar, R.; Bhatia, M.; Pai, K. Role of Cytokines in the Pathogenesis of Visceral Leishmaniasis. Clin. Lab. 2017, 63, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Nylen, S.; Sacks, D. Interleukin-10 and the pathogenesis of human visceral leishmaniasis. Trends Immunol. 2007, 28, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Madhubala, R. Ubiquitin conjugation of open reading frame F DNA vaccine leads to enhanced cell-mediated immune response and induces protection against both antimony-susceptible and-resistant strains of Leishmania donovani. J. Immunol. 2009, 183, 7719–7731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islamuddin, M.; Chouhan, G.; Want, M.Y.; Tyagi, M.; Abdin, M.Z.; Sahal, D.; Afrin, F. Corrigendum: Leishmanicidal activities of Artemisia annua leaf essential oil against Visceral Leishmaniasis. Front. Microbiol. 2015, 6, 1015. [Google Scholar] [CrossRef] [Green Version]
- Soto, M.; Ramirez, L.; Solana, J.C.; Cook, E.C.L.; Hernandez-Garcia, E.; Charro-Zanca, S.; Redondo-Urzainqui, A.; Reguera, R.M.; Balana-Fouce, R.; Iborra, S. Resistance to Experimental Visceral Leishmaniasis in Mice Infected With Leishmania infantum Requires Batf3. Front. Immunol. 2020, 11, 590934. [Google Scholar] [CrossRef]
- Kumar, P.; Misra, P.; Yadav, N.K.; Joshi, S.; Sahasrabuddhe, A.A.; Dube, A.; Rishi, N.; Mitra, D.K. Prophylactic interferon-gamma and interleukin-17 facilitate parasite clearance in experimental visceral leishmaniasis. Trop. Parasitol. 2019, 9, 30–35. [Google Scholar] [CrossRef]
- Carrion, J.; Nieto, A.; Iborra, S.; Iniesta, V.; Soto, M.; Folgueira, C.; Abanades, D.R.; Requena, J.M.; Alonso, C. Immunohistological features of visceral leishmaniasis in BALB/c mice. Parasite Immunol. 2006, 28, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Goto, Y.; Gidwani, K.; Cowgill, K.D.; Sundar, S.; Reed, S.G. Evaluation of ex vivo human immune response against candidate antigens for a visceral leishmaniasis vaccine. Am. J. Trop. Med. Hyg. 2010, 82, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Samant, M.; Misra, P.; Khare, P.; Sisodia, B.; Shasany, A.K.; Dube, A. Th1-stimulatory polyproteins of soluble Leishmania donovani promastigotes ranging from 89.9 to 97.1 kDa offers long-lasting protection against experimental visceral leishmaniasis. Vaccine 2008, 26, 5700–5711. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Kushawaha, P.K.; Tripathi, C.D.; Sundar, S.; Dube, A. A novel recombinant Leishmania donovani p45, a partial coding region of methionine aminopeptidase, generates protective immunity by inducing a Th1 stimulatory response against experimental visceral leishmaniasis. Int. J. Parasitol. 2012, 42, 429–435. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ChimT Sequence | KKKKG-LFVNQKLVGTIADVRSYEK (XP_001470432.1; LiHyC)-GKKG-YIMSGPARYVYFHMVLPVEAQ (XP_001468941.1; LiHyp1)-GKKKG-GVCVANTNVAAGAHTAALAAAVCVV (XP_001462854.1; LiHyV)-GKKKG-SLFVLYMYVTCRGGYTYLQL (XP_001467126.1; LiHyG)-GKKKK |
|---|---|
| Physical–chemical characteristics | 113 amino acid residues |
| Molecular weight of 11.9 kDa | |
| Isoelectric point of 10.71 | |
| Instability index of 6.09 | |
| Aliphatic index of 93.27 | |
| Grand average of hydropathicity (GRAVY) of 0.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lage, D.P.; Vale, D.L.; Linhares, F.P.; Freitas, C.S.; Machado, A.S.; Cardoso, J.M.O.; de Oliveira, D.; Galvani, N.C.; de Oliveira, M.P.; Oliveira-da-Silva, J.A.; et al. A Recombinant Chimeric Protein-Based Vaccine Containing T-Cell Epitopes from Amastigote Proteins and Combined with Distinct Adjuvants, Induces Immunogenicity and Protection against Leishmania infantum Infection. Vaccines 2022, 10, 1146. https://doi.org/10.3390/vaccines10071146
Lage DP, Vale DL, Linhares FP, Freitas CS, Machado AS, Cardoso JMO, de Oliveira D, Galvani NC, de Oliveira MP, Oliveira-da-Silva JA, et al. A Recombinant Chimeric Protein-Based Vaccine Containing T-Cell Epitopes from Amastigote Proteins and Combined with Distinct Adjuvants, Induces Immunogenicity and Protection against Leishmania infantum Infection. Vaccines. 2022; 10(7):1146. https://doi.org/10.3390/vaccines10071146
Chicago/Turabian StyleLage, Daniela P., Danniele L. Vale, Flávia P. Linhares, Camila S. Freitas, Amanda S. Machado, Jamille M. O. Cardoso, Daysiane de Oliveira, Nathália C. Galvani, Marcelo P. de Oliveira, João A. Oliveira-da-Silva, and et al. 2022. "A Recombinant Chimeric Protein-Based Vaccine Containing T-Cell Epitopes from Amastigote Proteins and Combined with Distinct Adjuvants, Induces Immunogenicity and Protection against Leishmania infantum Infection" Vaccines 10, no. 7: 1146. https://doi.org/10.3390/vaccines10071146
APA StyleLage, D. P., Vale, D. L., Linhares, F. P., Freitas, C. S., Machado, A. S., Cardoso, J. M. O., de Oliveira, D., Galvani, N. C., de Oliveira, M. P., Oliveira-da-Silva, J. A., Ramos, F. F., Tavares, G. S. V., Ludolf, F., Bandeira, R. S., Pereira, I. A. G., Chávez-Fumagalli, M. A., Roatt, B. M., Machado-de-Ávila, R. A., Christodoulides, M., ... Martins, V. T. (2022). A Recombinant Chimeric Protein-Based Vaccine Containing T-Cell Epitopes from Amastigote Proteins and Combined with Distinct Adjuvants, Induces Immunogenicity and Protection against Leishmania infantum Infection. Vaccines, 10(7), 1146. https://doi.org/10.3390/vaccines10071146