
The Importance of Thioredoxin-1 in Health and Disease


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
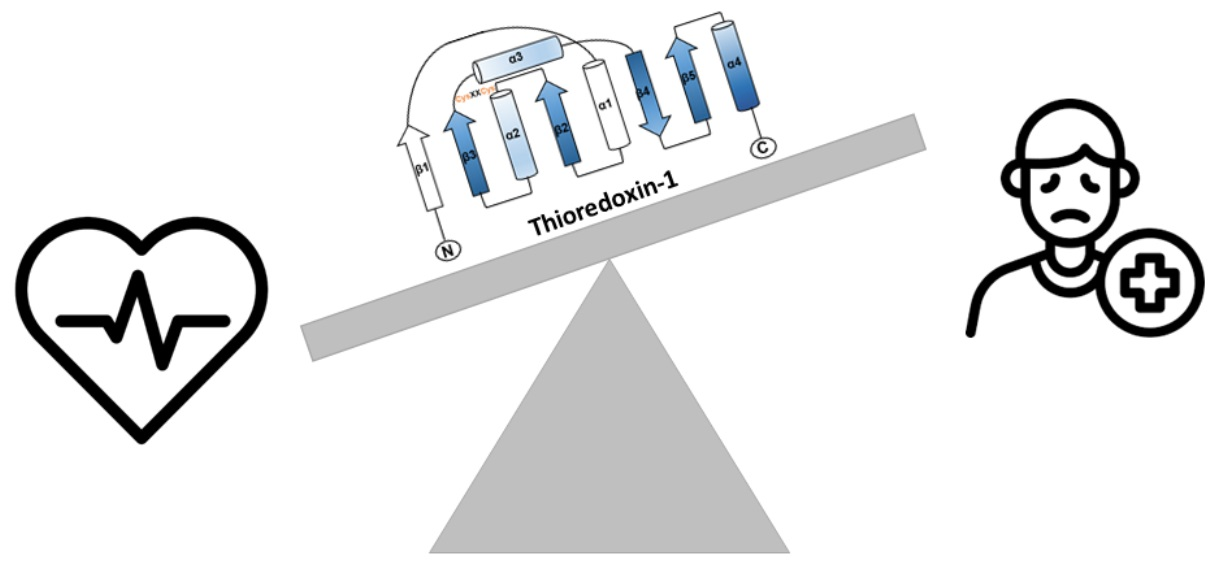
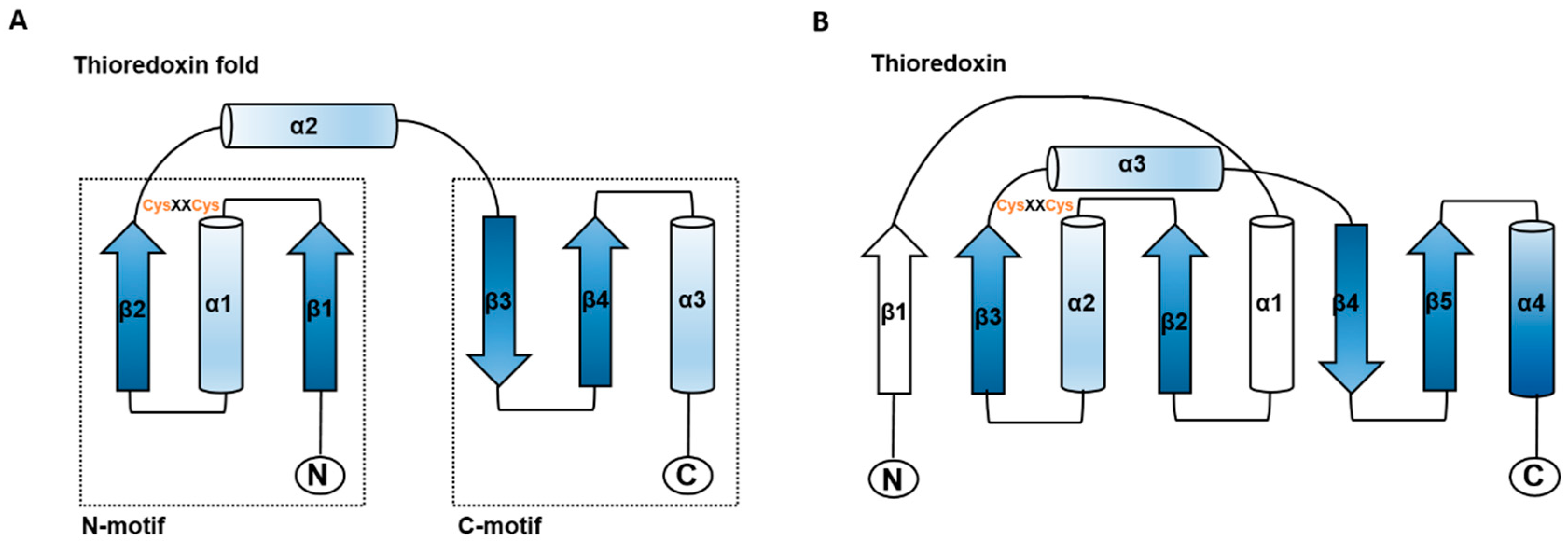
1.1. The Thioredoxin Family Proteins
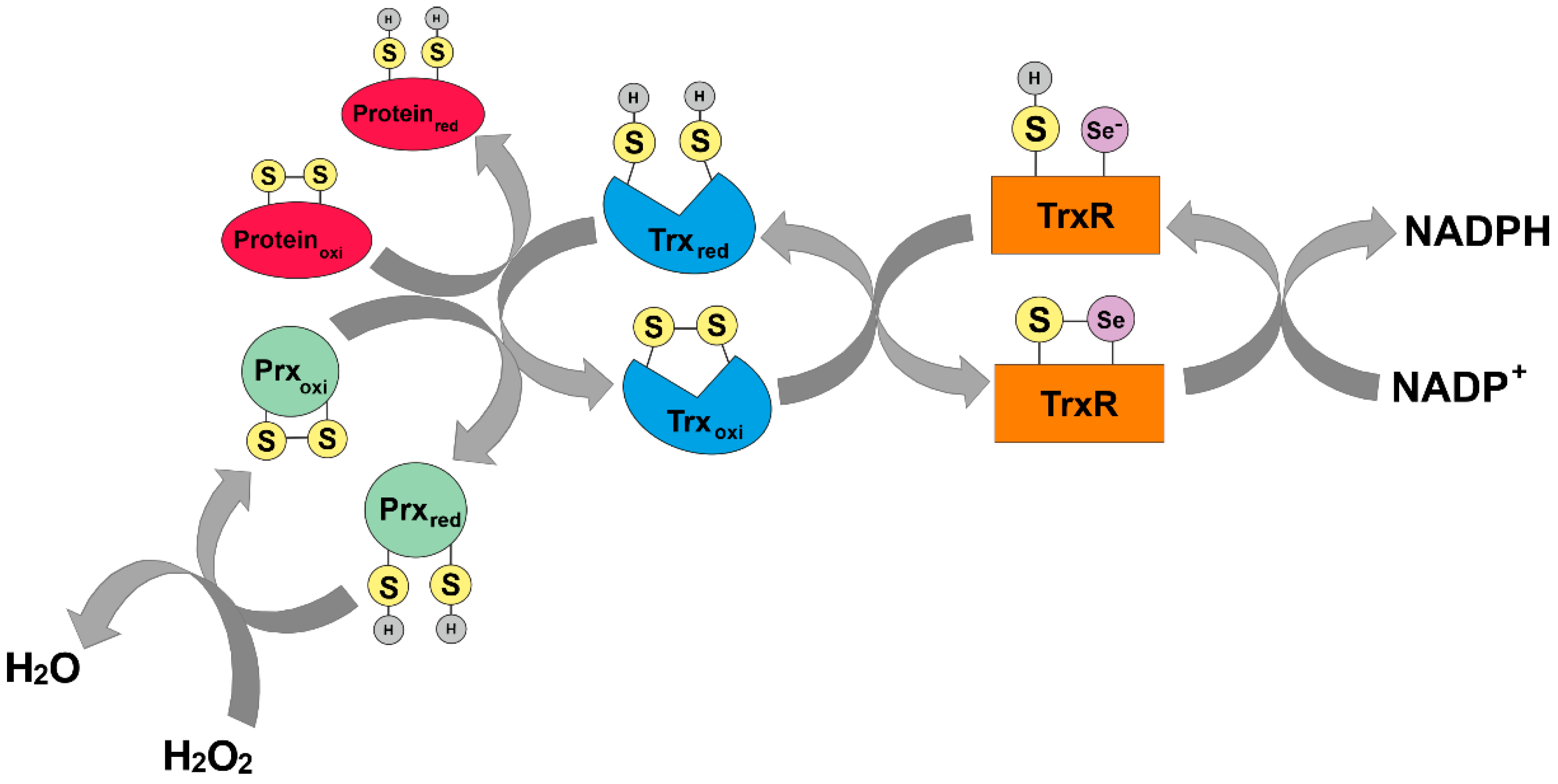
1.2. The Thioredoxin System
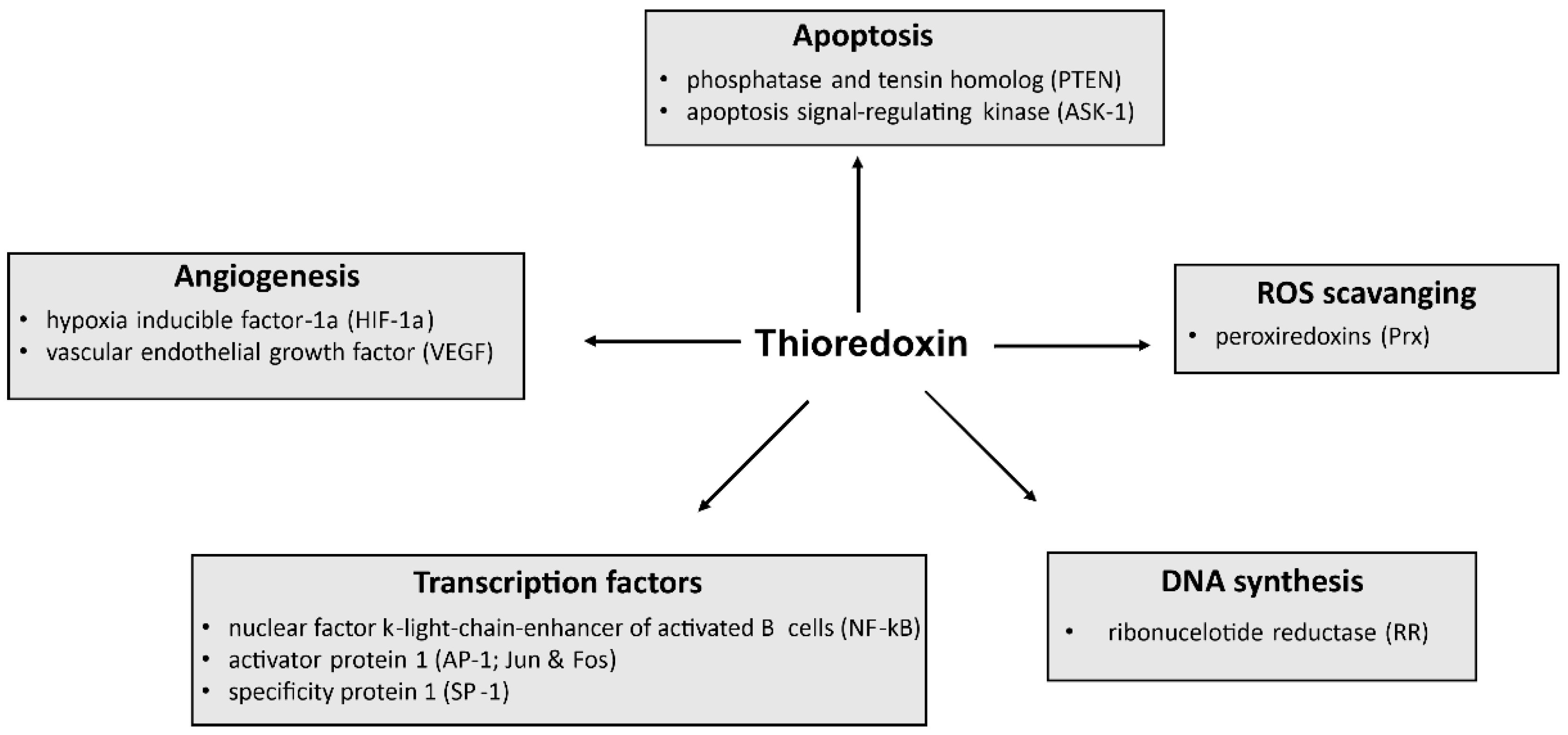
2. Functions of Trx-1
2.1. Antioxidant Defense
2.1.1. Regulation of Trx Activity
Gene Expression Level
Protein–Protein Interaction
Post-Translational Modifications
2.2. Regulation of Gene Expression
2.2.1. Activator Protein 1 (AP-1)
2.2.2. Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B-Cells (NF-κB)
2.3. Regulation of Cell Death
2.4. Inflammation
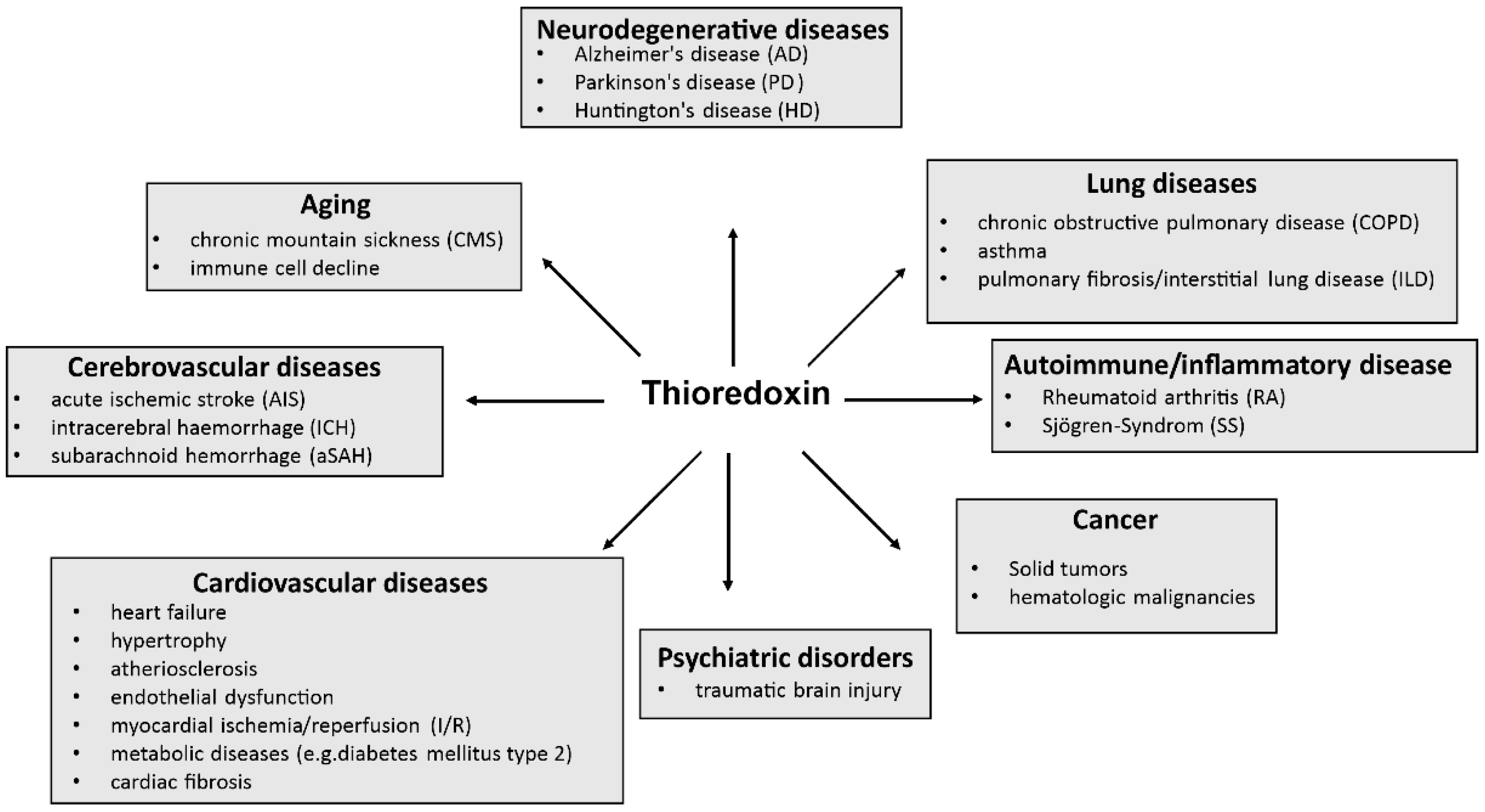
2.5. Trx-1 in Disease
3. Trx as a Predictive Biological and Prognostic Marker in Cancer
3.1. Lung
3.2. Gut
3.3. Urogenital
3.4. Hemato-Oncology
4. Trx as a Predictive Biological and Prognostic Marker in Non-Cancer Diseases
4.1. Autoimmune and Inflammatory Diseases
4.2. Lung
4.3. Abdominal Disease
4.4. Cardiovascular
4.5. Cerebrovascular
4.6. Trauma
4.7. Aging
5. Trx-1 in Health and Disease—Potential Therapeutic Approaches
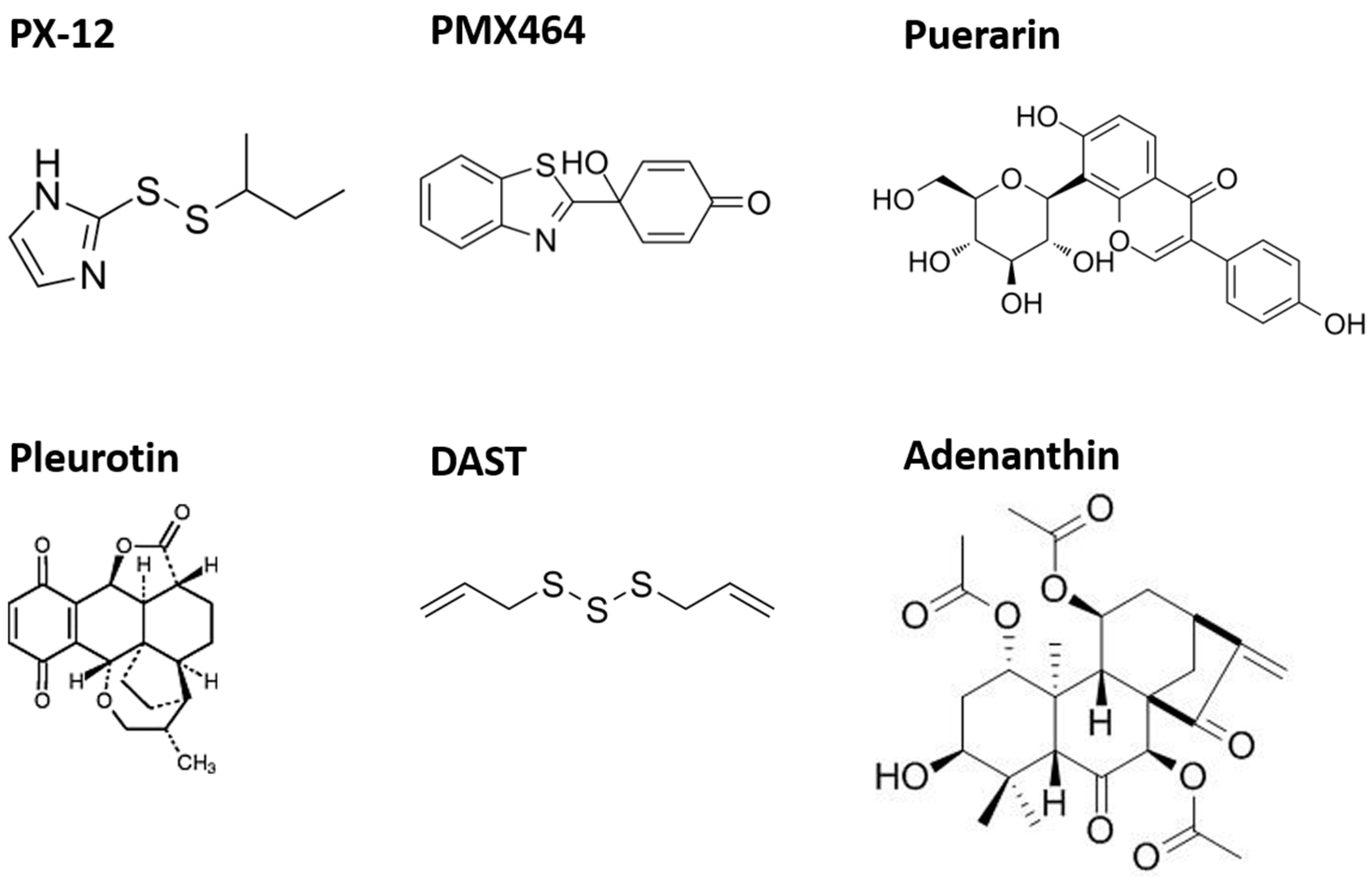
5.1. PX-12 (1-Methylpropyl 2-Imidazolyl Disulfide)
5.2. Quinols
5.3. PMX464 (4-(Benzothiazol-2-yl)- 4-Hydroxycyclohexa-2,5-dienone—Previously AW464; [123])
5.4. Secondary Metabolites
5.4.1. Puerarin
5.4.2. Pleurotin
5.4.3. Adenanthin
5.4.4. Diallyl Trisulfide (DAST)
5.4.5. Quinones (P-Benzoquinone, BQ)
5.5. Administration of Trx-1
5.6. Recombinant Trx-1
5.7. Human Serum Albumin-Trx Fusion Protein (HSA-Trx)
5.8. In Vitro or In Vivo Manipulation of Trx in Disease
5.8.1. In Vitro or In Vivo Deficiency of Trx
5.8.2. In Vitro or In Vivo Overexpression of Trx-1
5.9. Therapeutic Agents
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holmgren, A. Thioredoxin. Annu. Rev. Biochem. 1985, 54, 237–271. [Google Scholar] [CrossRef] [PubMed]
- Black, S.; Harte, E.M.; Hudson, B.; Wartofsky, L. A Specific Enzymatic Reduction of l(-) Methionine Sulfoxide and a Related Nonspecific Reduction of Disulfides. J. Biol. Chem. 1960, 235, 2910–2916. [Google Scholar] [CrossRef]
- Wilson, L.G.; Asahi, T.; Bandurski, R.S. Yeast sulfate-reducing system. I. Reduction of sulfate to sulfite. J. Biol. Chem. 1961, 236, 1822–1829. [Google Scholar] [CrossRef]
- Laurent, T.C.; Moore, E.C.; Reichard, P. Enzymatic Synthesis of Deoxyribonucleotides: IV. Isolation and characterization of thioredoxin, the hydrogen donor from Escherichia coli B. J. Biol. Chem. 1964, 239, 3436–3444. [Google Scholar] [CrossRef] [PubMed]
- Tagaya, Y.; Maeda, Y.; Mitsui, A.; Kondo, N.; Matsui, H.; Hamuro, J.; Brown, N.; Arai, K.; Yokota, T.; Wakasugi, H.; et al. ATL-derived factor (ADF), an IL-2 receptor/Tac inducer homologous to thioredoxin; possible involvement of dithiol-reduction in the IL-2 receptor induction. EMBO J. 1989, 8, 757–764. [Google Scholar] [CrossRef]
- Powis, G.; Montfort, W.R. Properties and biological activities of thioredoxins. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 421–455. [Google Scholar] [CrossRef] [PubMed]
- Arnér, E.S.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef]
- Nakamura, H.; Nakamura, K.; Yodoi, J. Redox regulation of cellular activation. Annu. Rev. Immunol. 1997, 15, 351–369. [Google Scholar] [CrossRef]
- Pekkari, K.; Holmgren, A. Truncated thioredoxin: Physiological functions and mechanism. Antioxid. Redox Signal. 2004, 6, 53–61. [Google Scholar] [CrossRef]
- Whayne, T.F., Jr.; Parinandi, N.; Maulik, N. Thioredoxins in cardiovascular disease. Can. J. Physiol. Pharmacol. 2015, 93, 903–911. [Google Scholar] [CrossRef]
- Marks, P.A. Thioredoxin in cancer--role of histone deacetylase inhibitors. Semin. Cancer Biol. 2006, 16, 436–443. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, J. [Thioredoxin and neurodegenerative diseases]. Sheng Li Ke Xue Jin Zhan [Prog. Physiol.] 2002, 33, 74–76. [Google Scholar] [PubMed]
- Matsui, M.; Oshima, M.; Oshima, H.; Takaku, K.; Maruyama, T.; Yodoi, J.; Taketo, M.M. Early embryonic lethality caused by targeted disruption of the mouse thioredoxin gene. Dev. Biol. 1996, 178, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Nonn, L.; Williams, R.R.; Erickson, R.P.; Powis, G. The absence of mitochondrial thioredoxin 2 causes massive apoptosis, exencephaly, and early embryonic lethality in homozygous mice. Mol. Cell. Biol. 2003, 23, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. The thioredoxin superfamily in oxidative protein folding. Antioxid. Redox Signal. 2014, 21, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Hirota, K.; Nakamura, H.; Yodoi, J. Redox Regulation by Thioredoxin and its Related Molecules. Drug News Perspect. 2002, 15, 575–580. [Google Scholar] [CrossRef]
- Martin, J.L. Thioredoxin—A fold for all reasons. Structure 1995, 3, 245–250. [Google Scholar] [CrossRef]
- Atkinson, H.J.; Babbitt, P.C. An atlas of the thioredoxin fold class reveals the complexity of function-enabling adaptations. PLoS Comput. Biol. 2009, 5, e1000541. [Google Scholar] [CrossRef]
- Holmgren, A. Thioredoxin and glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Holmgren, A. Antioxidant function of thioredoxin and glutaredoxin systems. Antioxid. Redox Signal. 2000, 2, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.P.; Holmgren, A. Glutaredoxins: Glutathione-dependent redox enzymes with functions far beyond a simple thioredoxin backup system. Antioxid. Redox Signal. 2004, 6, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Ladenstein, R.; Epp, O.; Bartels, K.; Jones, A.; Huber, R.; Wendel, A. Structure analysis and molecular model of the selenoenzyme glutathione peroxidase at 2.8 A resolution. J. Mol. Biol. 1979, 134, 199–218. [Google Scholar] [CrossRef] [PubMed]
- Wood, Z.A.; Schröder, E.; Robin Harris, J.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Rosen, A.; Lundman, P.; Carlsson, M.; Bhavani, K.; Srinivasa, B.R.; Kjellström, G.; Nilsson, K.; Holmgren, A. A CD4+ T cell line-secreted factor, growth promoting for normal and leukemic B cells, identified as thioredoxin. Int. Immunol. 1995, 7, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A.; Johansson, C.; Berndt, C.; Lönn, M.E.; Hudemann, C.; Lillig, C.H. Thiol redox control via thioredoxin and glutaredoxin systems. Biochem. Soc. Trans. 2005, 33, 1375–1377. [Google Scholar] [CrossRef]
- Kallis, G.B.; Holmgren, A. Differential reactivity of the functional sulfhydryl groups of cysteine-32 and cysteine-35 present in the reduced form of thioredoxin from Escherichia coli. J. Biol. Chem. 1980, 255, 10261–10265. [Google Scholar] [CrossRef]
- Karlenius, T.C.; Tonissen, K.F. Thioredoxin and Cancer: A Role for Thioredoxin in all States of Tumor Oxygenation. Cancers 2010, 2, 209–232. [Google Scholar] [CrossRef]
- Rhee, S.G.; Yang, K.S.; Kang, S.W.; Woo, H.A.; Chang, T.S. Controlled elimination of intracellular H2O2: Regulation of peroxiredoxin, catalase, and glutathione peroxidase via post-translational modification. Antioxid. Redox Signal. 2005, 7, 619–626. [Google Scholar] [CrossRef]
- Haendeler, J. Thioredoxin-1 and posttranslational modifications. Antioxid. Redox Signal. 2006, 8, 1723–1728. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, H.-J.K.; Karlenius, T.C.; Tonissen, K.F. Regulation of the human thioredoxin gene promoter and its key substrates: A study of functional and putative regulatory elements. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2014, 1840, 303–314. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Yamaguchi, Y.; Kondo, N.; Masutani, H.; Yodoi, J. Thioredoxin-dependent redox regulation of the antioxidant responsive element (ARE) in electrophile response. Oncogene 2003, 22, 1860–1865. [Google Scholar] [CrossRef]
- Nishiyama, A.; Matsui, M.; Iwata, S.; Hirota, K.; Masutani, H.; Nakamura, H.; Takagi, Y.; Sono, H.; Gon, Y.; Yodoi, J. Identification of thioredoxin-binding protein-2/vitamin D(3) up-regulated protein 1 as a negative regulator of thioredoxin function and expression. J. Biol. Chem. 1999, 274, 21645–21650. [Google Scholar] [CrossRef]
- Yoshihara, E. TXNIP/TBP-2: A Master Regulator for Glucose Homeostasis. Antioxidants 2020, 9, 765. [Google Scholar] [CrossRef]
- Yamanaka, H.; Maehira, F.; Oshiro, M.; Asato, T.; Yanagawa, Y.; Takei, H.; Nakashima, Y. A possible interaction of thioredoxin with VDUP1 in HeLa cells detected in a yeast two-hybrid system. Biochem. Biophys. Res. Commun. 2000, 271, 796–800. [Google Scholar] [CrossRef]
- Schulze, P.C.; Yoshioka, J.; Takahashi, T.; He, Z.; King, G.L.; Lee, R.T. Hyperglycemia Promotes Oxidative Stress through Inhibition of Thioredoxin Function by Thioredoxin-interacting Protein. J. Biol. Chem. 2004, 279, 30369–30374. [Google Scholar] [CrossRef]
- Patwari, P.; Higgins, L.J.; Chutkow, W.A.; Yoshioka, J.; Lee, R.T. The interaction of thioredoxin with Txnip. Evidence for formation of a mixed disulfide by disulfide exchange. J. Biol. Chem. 2006, 281, 21884–21891. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Masutani, H.; Nakamura, H.; Nishinaka, Y.; Yodoi, J. Redox regulation by thioredoxin and thioredoxin-binding proteins. IUBMB Life 2001, 52, 29–33. [Google Scholar] [CrossRef]
- Fould, B.; Lamamy, V.; Guenin, S.P.; Ouvry, C.; Cogé, F.; Boutin, J.A.; Ferry, G. Mutagenic analysis in a pure molecular system shows that thioredoxin-interacting protein residue Cys247 is necessary and sufficient for a mixed disulfide formation with thioredoxin. Protein Sci. Publ. Protein Soc. 2012, 21, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Suh, H.W.; Jeon, Y.H.; Hwang, E.; Nguyen, L.T.; Yeom, J.; Lee, S.G.; Lee, C.; Kim, K.J.; Kang, B.S.; et al. The structural basis for the negative regulation of thioredoxin by thioredoxin-interacting protein. Nat. Commun. 2014, 5, 2958. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Mukai, N.; Wang, B.F.; Yang, K.; Patwari, P.; Kitsis, R.N.; Yoshioka, J. Txnip C247S mutation protects the heart against acute myocardial infarction. J. Mol. Cell. Cardiol. 2021, 155, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Nakayama, Y.; Abdali, S.A.; Yoshioka, J. Cardiomyocyte-specific Txnip C247S mutation improves left ventricular functional reserve in streptozotocin-induced diabetic mice. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H259–H274. [Google Scholar] [CrossRef]
- Chen, X.; Tang, W.; Liu, S.; Yu, L.; Chen, Z. Thioredoxin-1 phosphorylated at T100 is needed for its anti-apoptotic activity in HepG2 cancer cells. Life Sci. 2010, 87, 254–260. [Google Scholar] [CrossRef]
- Casagrande, S.; Bonetto, V.; Fratelli, M.; Gianazza, E.; Eberini, I.; Massignan, T.; Salmona, M.; Chang, G.; Holmgren, A.; Ghezzi, P. Glutathionylation of human thioredoxin: A possible crosstalk between the glutathione and thioredoxin systems. Proc. Natl. Acad. Sci. USA 2002, 99, 9745–9749. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Marletta, M.A. Thioredoxin catalyzes the S-nitrosation of the caspase-3 active site cysteine. Nat. Chem. Biol. 2005, 1, 154–158. [Google Scholar] [CrossRef]
- Haendeler, J.; Hoffmann, J.; Tischler, V.; Berk, B.C.; Zeiher, A.M.; Dimmeler, S. Redox regulatory and anti-apoptotic functions of thioredoxin depend on S-nitrosylation at cysteine 69. Nat. Cell Biol. 2002, 4, 743–749. [Google Scholar] [CrossRef]
- Wu, C.; Liu, T.; Chen, W.; Oka, S.; Fu, C.; Jain, M.R.; Parrott, A.M.; Baykal, A.T.; Sadoshima, J.; Li, H. Redox regulatory mechanism of transnitrosylation by thioredoxin. Mol. Cell. Proteom. MCP 2010, 9, 2262–2275. [Google Scholar] [CrossRef]
- Holmgren, A.; Lu, J. Thioredoxin and thioredoxin reductase: Current research with special reference to human disease. Biochem. Biophys. Res. Commun. 2010, 396, 120–124. [Google Scholar] [CrossRef]
- Holmgren, A.; Luthman, M. Tissue distrubution and subcellular localization of bovine thioredoxin determined by radioimmunoassay. Biochemistry 1978, 17, 4071–4077. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Matsui, M.; Iwata, S.; Nishiyama, A.; Mori, K.; Yodoi, J. AP-1 transcriptional activity is regulated by a direct association between thioredoxin and Ref-1. Proc. Natl. Acad. Sci. USA 1997, 94, 3633–3638. [Google Scholar] [CrossRef]
- Abate, C.; Patel, L.; Rauscher, F.J., 3rd; Curran, T. Redox regulation of fos and jun DNA-binding activity in vitro. Science 1990, 249, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Xanthoudakis, S.; Curran, T. Identification and characterization of Ref-1, a nuclear protein that facilitates AP-1 DNA-binding activity. EMBO J. 1992, 11, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Murata, M.; Sachi, Y.; Nakamura, H.; Takeuchi, J.; Mori, K.; Yodoi, J. Distinct roles of thioredoxin in the cytoplasm and in the nucleus. A two-step mechanism of redox regulation of transcription factor NF-kappaB. J. Biol. Chem. 1999, 274, 27891–27897. [Google Scholar] [CrossRef] [PubMed]
- Das, K.C. c-Jun NH2-terminal kinase-mediated redox-dependent degradation of IkappaB: Role of thioredoxin in NF-kappaB activation. J. Biol. Chem. 2001, 276, 4662–4670. [Google Scholar] [CrossRef]
- Hayashi, T.; Ueno, Y.; Okamoto, T. Oxidoreductive regulation of nuclear factor kappa B. Involvement of a cellular reducing catalyst thioredoxin. J. Biol. Chem. 1993, 268, 11380–11388. [Google Scholar] [CrossRef]
- Okamoto, T.; Asamitsu, K.; Tetsuka, T. Thioredoxin and mechanism of inflammatory response. Methods Enzymol. 2002, 347, 349–360. [Google Scholar] [CrossRef]
- Yang, J.P.; Merin, J.P.; Nakano, T.; Kato, T.; Kitade, Y.; Okamoto, T. Inhibition of the DNA-binding activity of NF-kappa B by gold compounds in vitro. FEBS Lett. 1995, 361, 89–96. [Google Scholar] [CrossRef]
- Saitoh, M.; Nishitoh, H.; Fujii, M.; Takeda, K.; Tobiume, K.; Sawada, Y.; Kawabata, M.; Miyazono, K.; Ichijo, H. Mammalian thioredoxin is a direct inhibitor of apoptosis signal-regulating kinase (ASK) 1. EMBO J. 1998, 17, 2596–2606. [Google Scholar] [CrossRef]
- Liu, Y.; Min, W. Thioredoxin promotes ASK1 ubiquitination and degradation to inhibit ASK1-mediated apoptosis in a redox activity-independent manner. Circ. Res. 2002, 90, 1259–1266. [Google Scholar] [CrossRef]
- Hashemy, S.I.; Holmgren, A. Regulation of the catalytic activity and structure of human thioredoxin 1 via oxidation and S-nitrosylation of cysteine residues. J. Biol. Chem. 2008, 283, 21890–21898. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wan, A.; Xu, G.; Ye, D. Small changes huge impact: The role of thioredoxin 1 in the regulation of apoptosis by S-nitrosylation. Acta Biochim. Biophys. Sin. 2013, 45, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Muri, J.; Kopf, M. The thioredoxin system: Balancing redox responses in immune cells and tumors. Eur. J. Immunol. 2023, 53, e2249948. [Google Scholar] [CrossRef]
- Tamaki, H.; Nakamura, H.; Nishio, A.; Nakase, H.; Ueno, S.; Uza, N.; Kido, M.; Inoue, S.; Mikami, S.; Asada, M.; et al. Human thioredoxin-1 ameliorates experimental murine colitis in association with suppressed macrophage inhibitory factor production. Gastroenterology 2006, 131, 1110–1121. [Google Scholar] [CrossRef] [PubMed]
- Awan, M.U.N.; Yan, F.; Mahmood, F.; Bai, L.; Liu, J.; Bai, J. The Functions of Thioredoxin 1 in Neurodegeneration. Antioxid. Redox Signal. 2022, 36, 1023–1036. [Google Scholar] [CrossRef]
- Xu, J.; Li, T.; Wu, H.; Xu, T. Role of thioredoxin in lung disease. Pulm. Pharmacol. Ther. 2012, 25, 154–162. [Google Scholar] [CrossRef]
- Jabbar, S.; Mathews, P.; Kang, Y. Emerging Evidence of the Significance of Thioredoxin-1 in Hematopoietic Stem Cell Aging. Antioxidants 2022, 11, 1291. [Google Scholar] [CrossRef]
- Flores, L.C.; Ortiz, M.; Dube, S.; Hubbard, G.B.; Lee, S.; Salmon, A.; Zhang, Y.; Ikeno, Y. Thioredoxin, oxidative stress, cancer and aging. Longev. Healthspan 2012, 1, 4. [Google Scholar] [CrossRef]
- Li, H.; Wan, A. Apoptosis of rheumatoid arthritis fibroblast-like synoviocytes: Possible roles of nitric oxide and the thioredoxin 1. Mediat. Inflamm. 2013, 2013, 953462. [Google Scholar] [CrossRef]
- Griffiths, H.R.; Bennett, S.J.; Olofsson, P.; Dunston, C.R. Thioredoxin as a putative biomarker and candidate target in age-related immune decline. Biochem. Soc. Trans. 2014, 42, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.S.; Geng, W.S.; Chen, L.; Jia, J.J. Thioredoxin as a Therapeutic Target in Cerebral Ischemia. Curr. Pharm. Des. 2018, 24, 2986–2992. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.P.; Ogata, F.T.; Stern, A. Thioredoxin promotes survival signaling events under nitrosative/oxidative stress associated with cancer development. Biomed. J. 2017, 40, 189–199. [Google Scholar] [CrossRef]
- Miyazaki, K.; Noda, N.; Okada, S.; Hagiwara, Y.; Miyata, M.; Sakurabayashi, I.; Yamaguchi, N.; Sugimura, T.; Terada, M.; Wakasugi, H. Elevated serum level of thioredoxin in patients with hepatocellular carcinoma. Biotherapy 1998, 11, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Kakolyris, S.; Giatromanolaki, A.; Koukourakis, M.; Powis, G.; Souglakos, J.; Sivridis, E.; Georgoulias, V.; Gatter, K.C.; Harris, A.L. Thioredoxin expression is associated with lymph node status and prognosis in early operable non-small cell lung cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2001, 7, 3087–3091. [Google Scholar]
- Noike, T.; Miwa, S.; Soeda, J.; Kobayashi, A.; Miyagawa, S. Increased expression of thioredoxin-1, vascular endothelial growth factor, and redox factor-1 is associated with poor prognosis in patients with liver metastasis from colorectal cancer. Hum. Pathol. 2008, 39, 201–208. [Google Scholar] [CrossRef]
- Tamai, T.; Uto, H.; Takami, Y.; Oda, K.; Saishoji, A.; Hashiguchi, M.; Kumagai, K.; Kure, T.; Mawatari, S.; Moriuchi, A.; et al. Serum manganese superoxide dismutase and thioredoxin are potential prognostic markers for hepatitis C virus-related hepatocellular carcinoma. World J. Gastroenterol. 2011, 17, 4890–4898. [Google Scholar] [CrossRef]
- Nagano, M.; Hatakeyama, K.; Kai, M.; Nakamura, H.; Yodoi, J.; Asada, Y.; Chijiiwa, K. Nuclear expression of thioredoxin-1 in the invasion front is associated with outcome in patients with gallbladder carcinoma. HPB Off. J. Int. Hepato Pancreato Biliary Assoc. 2012, 14, 573–582. [Google Scholar] [CrossRef][Green Version]
- Lim, J.Y.; Yoon, S.O.; Hong, S.W.; Kim, J.W.; Choi, S.H.; Cho, J.Y. Thioredoxin and thioredoxin-interacting protein as prognostic markers for gastric cancer recurrence. World J. Gastroenterol. 2012, 18, 5581–5588. [Google Scholar] [CrossRef]
- Duangkumpha, K.; Stoll, T.; Phetcharaburanin, J.; Yongvanit, P.; Thanan, R.; Techasen, A.; Namwat, N.; Khuntikeo, N.; Chamadol, N.; Roytrakul, S.; et al. Discovery and Qualification of Serum Protein Biomarker Candidates for Cholangiocarcinoma Diagnosis. J. Proteome Res. 2019, 18, 3305–3316. [Google Scholar] [CrossRef]
- Tabata, C.; Terada, T.; Tabata, R.; Yamada, S.; Eguchi, R.; Fujimori, Y.; Nakano, T. Serum thioredoxin-1 as a diagnostic marker for malignant peritoneal mesothelioma. J. Clin. Gastroenterol. 2013, 47, e7–e11. [Google Scholar] [CrossRef] [PubMed]
- Shang, W.; Xie, Z.; Lu, F.; Fang, D.; Tang, T.; Bi, R.; Chen, L.; Jiang, L. Increased Thioredoxin-1 Expression Promotes Cancer Progression and Predicts Poor Prognosis in Patients with Gastric Cancer. Oxidative Med. Cell. Longev. 2019, 2019, 9291683. [Google Scholar] [CrossRef]
- Ribback, S.; Winter, S.; Klatte, T.; Schaeffeler, E.; Gellert, M.; Stühler, V.; Scharpf, M.; Bedke, J.; Burchardt, M.; Schwab, M.; et al. Thioredoxin 1 (Trx1) is associated with poor prognosis in clear cell renal cell carcinoma (ccRCC): An example for the crucial role of redox signaling in ccRCC. World J. Urol. 2022, 40, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Zhong, W.; Zhao, R.; Oberley, T.D. Thioredoxin 1 as a subcellular biomarker of redox imbalance in human prostate cancer progression. Free Radic. Biol. Med. 2010, 49, 2078–2087. [Google Scholar] [CrossRef] [PubMed]
- An, N.; Kang, Y. Thioredoxin and hematologic malignancies. Adv. Cancer Res. 2014, 122, 245–279. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Thompson, M.A.; Tamayo, A.T.; Zuo, Z.; Lee, J.; Vega, F.; Ford, R.J.; Pham, L.V. Over-expression of Thioredoxin-1 mediates growth, survival, and chemoresistance and is a druggable target in diffuse large B-cell lymphoma. Oncotarget 2012, 3, 314–326. [Google Scholar] [CrossRef]
- Peroja, P.; Pasanen, A.K.; Haapasaari, K.M.; Jantunen, E.; Soini, Y.; Turpeenniemi-Hujanen, T.; Bloigu, R.; Lilja, L.; Kuittinen, O.; Karihtala, P. Oxidative stress and redox state-regulating enzymes have prognostic relevance in diffuse large B-cell lymphoma. Exp. Hematol. Oncol. 2012, 1, 2. [Google Scholar] [CrossRef]
- Kari, E.J.M.; Kuusisto, M.E.L.; Honkavaara, P.; Hakalahti, A.; Haapasaari, K.M.; Bloigu, R.; Karihtala, P.; Teppo, H.R.; Pirinen, R.; Turpeenniemi-Hujanen, T.; et al. Thioredoxin-1 as a biological predictive marker for selecting diffuse large B-cell lymphoma patients for etoposide-containing treatment. Eur. J. Haematol. 2020, 105, 156–163. [Google Scholar] [CrossRef]
- Shao, L.; Diccianni, M.B.; Tanaka, T.; Gribi, R.; Yu, A.L.; Pullen, J.D.; Camitta, B.M.; Yu, J. Thioredoxin expression in primary T-cell acute lymphoblastic leukemia and its therapeutic implication. Cancer Res. 2001, 61, 7333–7338. [Google Scholar]
- Jikimoto, T.; Nishikubo, Y.; Koshiba, M.; Kanagawa, S.; Morinobu, S.; Morinobu, A.; Saura, R.; Mizuno, K.; Kondo, S.; Toyokuni, S.; et al. Thioredoxin as a biomarker for oxidative stress in patients with rheumatoid arthritis. Mol. Immunol. 2002, 38, 765–772. [Google Scholar] [CrossRef]
- Li, X.; Shen, H.; Zhou, T.; Cao, X.; Chen, Y.; Liang, Y.; Lu, T.; He, J.; Dou, Z.; Liu, C.; et al. Early Elevation of Thioredoxin-1 Serum Levels Predicts 28-Day Mortality in Patients with Sepsis. J. Inflamm. Res. 2021, 14, 3837–3848. [Google Scholar] [CrossRef]
- Yamada, Y.; Nakamura, H.; Adachi, T.; Sannohe, S.; Oyamada, H.; Kayaba, H.; Yodoi, J.; Chihara, J. Elevated serum levels of thioredoxin in patients with acute exacerbation of asthma. Immunol. Lett. 2003, 86, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.M.; Hu, H.; Lu, L.; Deem, A.; Akindipe, O.; Brantly, M.; Block, E.R.; Antony, V.B.; Baz, M.A. Thioredoxin as a biomarker for graft rejection in lung transplant recipients. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2008, 13, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, S.; Nishio, A.; Nakamura, H.; Kido, M.; Kiriya, K.; Asada, M.; Tamaki, H.; Fukui, T.; Kawasaki, K.; Watanabe, N.; et al. Clinical significance of serum thioredoxin 1 levels in patients with acute pancreatitis. Pancreas 2006, 32, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.M.; Li, M.J.; Tao, C. Serum and urinary thioredoxin concentrations are associated with severity of children hydronephrosis. Clin. Chim. Acta Int. J. Clin. Chem. 2017, 466, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Mongardon, N.; Lemiale, V.; Borderie, D.; Burke-Gaffney, A.; Perbet, S.; Marin, N.; Charpentier, J.; Pène, F.; Chiche, J.D.; Mira, J.P.; et al. Plasma thioredoxin levels during post-cardiac arrest syndrome: Relationship with severity and outcome. Crit. Care (Lond. Engl.) 2013, 17, R18. [Google Scholar] [CrossRef] [PubMed]
- Otaki, Y.; Watanabe, T.; Takahashi, H.; Kadowaki, S.; Narumi, T.; Honda, Y.; Wanezaki, M.; Sasaki, S.; Tamura, H.; Nishiyama, S.; et al. Association of plasma thioredoxin-1 with renal tubular damage and cardiac prognosis in patients with chronic heart failure. J. Cardiol. 2014, 64, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yang, L.; Zhong, L. Decreased serum levels of thioredoxin in patients with coronary artery disease plus hyperhomocysteinemia is strongly associated with the disease severity. Atherosclerosis 2010, 212, 351–355. [Google Scholar] [CrossRef]
- Yu, T.; Zhang, W.; Lin, Y.; Li, Q.; Xue, J.; Cai, Z.; Cheng, Y.; Shao, B. Prognostic value of serum thioredoxin levels in ischemic stroke. Neurol. Res. 2017, 39, 988–995. [Google Scholar] [CrossRef]
- Qi, A.Q.; Li, Y.; Liu, Q.; Si, J.Z.; Tang, X.M.; Zhang, Z.Q.; Qi, Q.D.; Chen, W.B. Thioredoxin is a novel diagnostic and prognostic marker in patients with ischemic stroke. Free Radic. Biol. Med. 2015, 80, 129–135. [Google Scholar] [CrossRef]
- Wu, M.H.; Song, F.Y.; Wei, L.P.; Meng, Z.Y.; Zhang, Z.Q.; Qi, Q.D. Serum Levels of Thioredoxin Are Associated with Stroke Risk, Severity, and Lesion Volumes. Mol. Neurobiol. 2016, 53, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.X.; Cai, J.Y.; Lin, Q.; Chen, X.D.; Lu, C.; Sun, J.; Ba, H.J. Thioredoxin as a marker for severity and prognosis of aneurysmal subarachnoid hemorrhage. J. Neurol. Sci. 2016, 363, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.Q.; Hu, X.C.; He, S.R.; Li, B.B.; Zheng, X.D.; Pan, G.H. Prognostic value of serum thioredoxin concentrations after intracerebral hemorrhage. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 455, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.; Gidlöf, A.; Eriksson, M.; Larsson, E.; Brattström, O.; Oldner, A. Thioredoxin a novel biomarker of post-injury sepsis. Free Radic. Biol. Med. 2017, 104, 138–143. [Google Scholar] [CrossRef]
- Pan, D.S.; Le, H.W.; Yan, M.; Hassan, M.; Gong, J.B.; Wang, H. Change of serum levels of thioredoxin in patients with severe traumatic brain injury. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 453, 62–66. [Google Scholar] [CrossRef]
- Zhang, P.; Li, Z.; Yang, F.; Ji, L.; Yang, Y.; Liu, C.; Liu, H.; Ma, J.; Liu, J.; Dang, Z.; et al. Novel insights into plasma biomarker candidates in patients with chronic mountain sickness based on proteomics. Biosci. Rep. 2021, 41, BSR20202219. [Google Scholar] [CrossRef]
- Bai, J.; Nakamura, H.; Hattori, I.; Tanito, M.; Yodoi, J. Thioredoxin suppresses 1-methyl-4-phenylpyridinium-induced neurotoxicity in rat PC12 cells. Neurosci. Lett. 2002, 321, 81–84. [Google Scholar] [CrossRef]
- Ehrenfeld, V.; Fulda, S. Thioredoxin inhibitor PX-12 induces mitochondria-mediated apoptosis in acute lymphoblastic leukemia cells. Biol. Chem. 2020, 401, 273–283. [Google Scholar] [CrossRef]
- Kinoshita, T.; Hoshino, T.; Imaoka, H.; Ichiki, H.; Okamoto, M.; Kawayama, T.; Yodoi, J.; Kato, S.; Aizawa, H. Thioredoxin prevents the development and progression of elastase-induced emphysema. Biochem. Biophys. Res. Commun. 2007, 354, 712–719. [Google Scholar] [CrossRef]
- Kinoshita, H.; Shimozato, O.; Ishii, T.; Kamoda, H.; Hagiwara, Y.; Ohtori, S.; Yonemoto, T. The Thioredoxin-1 Inhibitor, PX-12, Suppresses Local Osteosarcoma Progression. Anticancer Res. 2021, 41, 6013–6021. [Google Scholar] [CrossRef]
- Lin, Q.D.; Liu, L.N.; Liu, X.Y.; Yan, Y.; Fang, B.J.; Zhang, Y.L.; Zhou, J.; Li, Y.F.; Zuo, W.L.; Song, Y.P. Experimental study on thioredoxin redox inhibitor 1-methylpropyl 2-imidazolyl disulfide promoting apoptosis of multiple myeloma cells in vitro and in vivo. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, R.K.; Kirkpatrick, D.L.; Belani, C.P.; Friedland, D.; Green, S.B.; Chow, H.H.; Cordova, C.A.; Stratton, S.P.; Sharlow, E.R.; Baker, A.; et al. A Phase I pharmacokinetic and pharmacodynamic study of PX-12, a novel inhibitor of thioredoxin-1, in patients with advanced solid tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Bi, L.; Zhang, P.; Wang, F.; Lin, F.; Ni, W.; Wu, J.; Jiang, L. Thioredoxin-1 inhibitor PX-12 induces human acute myeloid leukemia cell apoptosis and enhances the sensitivity of cells to arsenic trioxide. Int. J. Clin. Exp. Pathol. 2014, 7, 4765–4773. [Google Scholar] [PubMed]
- Wang, F.; Lin, F.; Zhang, P.; Ni, W.; Bi, L.; Wu, J.; Jiang, L. Thioredoxin-1 inhibitor, 1-methylpropyl 2-imidazolyl disulfide, inhibits the growth, migration and invasion of colorectal cancer cell lines. Oncol. Rep. 2015, 33, 967–973. [Google Scholar] [CrossRef] [PubMed]
- You, B.R.; Shin, H.R.; Park, W.H. PX-12 inhibits the growth of A549 lung cancer cells via G2/M phase arrest and ROS-dependent apoptosis. Int. J. Oncol. 2014, 44, 301–308. [Google Scholar] [CrossRef]
- You, B.R.; Shin, H.R.; Han, B.R.; Park, W.H. PX-12 induces apoptosis in Calu-6 cells in an oxidative stress-dependent manner. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 2087–2095. [Google Scholar] [CrossRef]
- Ramanathan, R.K.; Stephenson, J.J.; Weiss, G.J.; Pestano, L.A.; Lowe, A.; Hiscox, A.; Leos, R.A.; Martin, J.C.; Kirkpatrick, L.; Richards, D.A. A phase I trial of PX-12, a small-molecule inhibitor of thioredoxin-1, administered as a 72-hour infusion every 21 days in patients with advanced cancers refractory to standard therapy. Investig. New Drugs 2012, 30, 1591–1596. [Google Scholar] [CrossRef]
- Jordan, B.F.; Runquist, M.; Raghunand, N.; Gillies, R.J.; Tate, W.R.; Powis, G.; Baker, A.F. The thioredoxin-1 inhibitor 1-methylpropyl 2-imidazolyl disulfide (PX-12) decreases vascular permeability in tumor xenografts monitored by dynamic contrast enhanced magnetic resonance imaging. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 529–536. [Google Scholar] [CrossRef]
- Chi, O.Z.; Barsoum, S.; Grayson, J.; Liu, X.; Weiss, H.R. Effects of the thioredoxin-1 inhibitor PX-12 on blood-brain barrier permeability in the early stage of focal cerebral ischemia. Pharmacology 2013, 92, 175–181. [Google Scholar] [CrossRef]
- Miyo, M.; Konno, M.; Colvin, H.; Nishida, N.; Koseki, J.; Kawamoto, K.; Tsunekuni, K.; Nishimura, J.; Hata, T.; Takemasa, I.; et al. The importance of mitochondrial folate enzymes in human colorectal cancer. Oncol. Rep. 2017, 37, 417–425. [Google Scholar] [CrossRef]
- Yu, L.; Guo, Q.; Luo, Z.; Wang, Y.; Weng, J.; Chen, Y.; Liang, W.; Li, Y.; Zhang, Y.; Chen, K.; et al. TXN inhibitor impedes radioresistance of colorectal cancer cells with decreased ALDH1L2 expression via TXN/NF-κB signaling pathway. Br. J. Cancer 2022, 127, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, T.D.; Matthews, C.S.; Cookson, J.; Chew, E.H.; Shah, M.; Bailey, K.; Monks, A.; Harris, E.; Westwell, A.D.; Wells, G.; et al. Elucidation of thioredoxin as a molecular target for antitumor quinols. Cancer Res. 2005, 65, 3911–3919. [Google Scholar] [CrossRef] [PubMed]
- Wells, G.; Berry, J.M.; Bradshaw, T.D.; Burger, A.M.; Seaton, A.; Wang, B.; Westwell, A.D.; Stevens, M.F. 4-Substituted 4-hydroxycyclohexa-2,5-dien-1-ones with selective activities against colon and renal cancer cell lines. J. Med. Chem. 2003, 46, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Martin, S.G. The thioredoxin system: A key target in tumour and endothelial cells. Br. J. Radiol. 2008, 1, S57–S68. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Westwell, A.D.; Bradshaw, T.D.; Stevens, M.F.; Carmichael, J.; Martin, S.G. Cytotoxic and antiangiogenic activity of AW464 (NSC 706704), a novel thioredoxin inhibitor: An in vitro study. Br. J. Cancer 2005, 92, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Huber, K.; Evans, H.; Lakhani, N.; Martin, S. A cellular and molecular investigation of the action of PMX464, a putative thioredoxin inhibitor, in normal and colorectal cancer cell lines. Br. J. Pharmacol. 2007, 151, 1167–1175. [Google Scholar] [CrossRef]
- Metcalfe, C.; Ramasubramoni, A.; Pula, G.; Harper, M.T.; Mundell, S.J.; Coxon, C.H. Thioredoxin Inhibitors Attenuate Platelet Function and Thrombus Formation. PLoS ONE 2016, 11, e0163006. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Y.; Yu, H.; Liu, X.; Jiao, B.; Lu, X. Libertellenone H, a Natural Pimarane Diterpenoid, Inhibits Thioredoxin System and Induces ROS-Mediated Apoptosis in Human Pancreatic Cancer Cells. Molecules 2021, 26, 315. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, D.; Osama, A.; Fang, J. Natural Molecules Targeting Thioredoxin System and Their Therapeutic Potential. Antioxid. Redox Signal. 2021, 34, 1083–1107. [Google Scholar] [CrossRef]
- Dal Piaz, F.; Braca, A.; Belisario, M.A.; De Tommasi, N. Thioredoxin system modulation by plant and fungal secondary metabolites. Curr. Med. Chem. 2010, 17, 479–494. [Google Scholar] [CrossRef]
- Zhang, L. Pharmacokinetics and drug delivery systems for puerarin, a bioactive flavone from traditional Chinese medicine. Drug Deliv. 2019, 26, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Zhang, H.; Peng, C. Puerarin: A review of pharmacological effects. Phytother. Res. PTR 2014, 28, 961–975. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Xu, L.H.; Zhao, W.; Tian, L.J.; Ji, X.L. The optimal therapeutic timing and mechanism of puerarin treatment of spinal cord ischemia-reperfusion injury in rats. J. Ethnopharmacol. 2011, 134, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, X.; Dong, D.; Lei, T.; Ou, H. Up-regulation of thioredoxin system by puerarin inhibits lipid uptake in macrophages. Free Radic. Biol. Med. 2021, 162, 542–554. [Google Scholar] [CrossRef]
- Robbins, W.J.; Kavanagh, F.; Hervey, A. Antibiotic Substances from Basidiomycetes. I. Pleurotus griseus. Proc. Natl. Acad. Sci. USA 1947, 33, 171–176. [Google Scholar] [CrossRef]
- Riondel, J.; Beriel, H.; Dardas, A.; Carraz, G.; Oddoux, L. Studies of antitumor activity of the culture filtrate of Hohenbuehelia geogenius (D.C. ex Fr.) Sing (basidiomycete). Arzneimittel-Forschung 1981, 31, 293–299. [Google Scholar]
- Wipf, P.; Hopkins, T.D.; Jung, J.K.; Rodriguez, S.; Birmingham, A.; Southwick, E.C.; Lazo, J.S.; Powis, G. New inhibitors of the thioredoxin-thioredoxin reductase system based on a naphthoquinone spiroketal natural product lead. Bioorg. Med. Chem. Lett. 2001, 11, 2637–2641. [Google Scholar] [CrossRef]
- Welsh, S.J.; Williams, R.R.; Birmingham, A.; Newman, D.J.; Kirkpatrick, D.L.; Powis, G. The thioredoxin redox inhibitors 1-methylpropyl 2-imidazolyl disulfide and pleurotin inhibit hypoxia-induced factor 1alpha and vascular endothelial growth factor formation. Mol. Cancer Ther. 2003, 2, 235–243. [Google Scholar]
- Berdicevsky, I.; Kaufman, G.; Newman, D.J.; Horwitz, B.A. Preliminary study of activity of the thioredoxin inhibitor pleurotin against Trichophyton mentagrophytes: A novel anti-dermatophyte possibility. Mycoses 2009, 52, 313–317. [Google Scholar] [CrossRef]
- Muchowicz, A.; Firczuk, M.; Chlebowska, J.; Nowis, D.; Stachura, J.; Barankiewicz, J.; Trzeciecka, A.; Kłossowski, S.; Ostaszewski, R.; Zagożdżon, R.; et al. Adenanthin targets proteins involved in the regulation of disulphide bonds. Biochem. Pharmacol. 2014, 89, 210–216. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Y.; Wei, Z.; Tao, L.; Sheng, X.; Wang, S.; Chen, J.; Ruan, J.; Liu, Z.; Cao, Y.; et al. Targeting Thioredoxin System with an Organosulfur Compound, Diallyl Trisulfide (DATS), Attenuates Progression and Metastasis of Triple-Negative Breast Cancer (TNBC). Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 50, 1945–1963. [Google Scholar] [CrossRef]
- Tian, Y.; Ge, Z.; Xu, M.; Ge, X.; Zhao, M.; Ding, F.; Yin, J.; Wang, X.; You, Y.; Shi, Z.; et al. Diallyl trisulfide sensitizes radiation therapy on glioblastoma through directly targeting thioredoxin 1. Free Radic. Biol. Med. 2022, 189, 157–168. [Google Scholar] [CrossRef]
- Shu, N.; Hägglund, P.; Cai, H.; Hawkins, C.L.; Davies, M.J. Modification of Cys residues in human thioredoxin-1 by p-benzoquinone causes inhibition of its catalytic activity and activation of the ASK1/p38-MAPK signalling pathway. Redox Biol. 2020, 29, 101400. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I. Functional Foods for Health: The Interrelated Antioxidant and Anti-Inflammatory Role of Fruits, Vegetables, Herbs, Spices and Cocoa in Humans. Curr. Pharm. Des. 2016, 22, 6701–6715. [Google Scholar] [CrossRef] [PubMed]
- Leitão, M.; Ribeiro, T.; García, P.A.; Barreiros, L.; Correia, P. Benefits of Fermented Papaya in Human Health. Foods 2022, 11, 563. [Google Scholar] [CrossRef] [PubMed]
- Marotta, F.; Chui, D.H.; Jain, S.; Polimeni, A.; Koike, K.; Zhou, L.; Lorenzetti, A.; Shimizu, H.; Yang, H. Effect of a fermented nutraceutical on thioredoxin level and TNF-alpha signalling in cirrhotic patients. J. Biol. Regul. Homeost. Agents 2011, 25, 37–45. [Google Scholar]
- Taketani, Y.; Kinugasa, K.; Kitajima, R.; Nishiumi, S.; Ashida, H.; Nakamura, H.; Fujita, T.; Kanzaki, K.; Masutani, H.; Yodoi, J. Protective effects of oral administration of yeast thioredoxin against gastric mucosal injury. Biosci. Biotechnol. Biochem. 2014, 78, 1221–1230. [Google Scholar] [CrossRef]
- Wang, W.; Fang, D.; Zhang, H.; Xue, J.; Wangchuk, D.; Du, J.; Jiang, L. Sodium Butyrate Selectively Kills Cancer Cells and Inhibits Migration in Colorectal Cancer by Targeting Thioredoxin-1. OncoTargets Ther. 2020, 13, 4691–4704. [Google Scholar] [CrossRef]
- Lee, R.L.; Rancourt, R.C.; del Val, G.; Pack, K.; Pardee, C.; Accurso, F.J.; White, C.W. Thioredoxin and dihydrolipoic acid inhibit elastase activity in cystic fibrosis sputum. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L875–L882. [Google Scholar] [CrossRef]
- Nakamura, T.; Hoshino, Y.; Yamada, A.; Teratani, A.; Furukawa, S.; Okuyama, H.; Ueda, S.; Wada, H.; Yodoi, J.; Nakamura, H. Recombinant human thioredoxin-1 becomes oxidized in circulation and suppresses bleomycin-induced neutrophil recruitment in the rat airway. Free Radic. Res. 2007, 41, 1089–1098. [Google Scholar] [CrossRef]
- Ueda, S.; Nakamura, T.; Yamada, A.; Teratani, A.; Matsui, N.; Furukawa, S.; Hoshino, Y.; Narita, M.; Yodoi, J.; Nakamura, H. Recombinant human thioredoxin suppresses lipopolysaccharide-induced bronchoalveolar neutrophil infiltration in rat. Life Sci. 2006, 79, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Ono, R.; Masaki, T.; Dien, S.; Yu, X.; Fukunaga, A.; Yodoi, J.; Nishigori, C. Suppressive effect of recombinant human thioredoxin on ultraviolet light-induced inflammation and apoptosis in murine skin. J. Dermatol. 2012, 39, 843–851. [Google Scholar] [CrossRef]
- Lee, B.W.; Jeon, B.S.; Yoon, B.I. Exogenous recombinant human thioredoxin-1 prevents acetaminophen-induced liver injury by scavenging oxidative stressors, restoring the thioredoxin-1 system and inhibiting receptor interacting protein-3 overexpression. J. Appl. Toxicol. 2018, 38, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, Q.; Lin, J.L.; Wang, Y.; Zhang, R.X.; Hou, J.B.; Yu, B. Recombinant Human Thioredoxin-1 Protects Macrophages from Oxidized Low-Density Lipoprotein-Induced Foam Cell Formation and Cell Apoptosis. Biomol. Ther. 2018, 26, 121–129. [Google Scholar] [CrossRef]
- Ikuta, S.; Chuang, V.T.; Ishima, Y.; Nakajou, K.; Furukawa, M.; Watanabe, H.; Maruyama, T.; Otagiri, M. Albumin fusion of thioredoxin--the production and evaluation of its biological activity for potential therapeutic applications. J. Control. Release: Off. J. Control. Release Soc. 2010, 147, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.I.; Kubota, M.; Shimoda, M.; Hayase, T.; Miyaguchi, M.; Kobayashi, N.; Ikeda, M.; Ishima, Y.; Kawahara, M. Thioredoxin-albumin fusion protein prevents urban aerosol-induced lung injury via suppressing oxidative stress-related neutrophil extracellular trap formation. Environ. Pollut. (Barking Essex 1987) 2021, 268, 115787. [Google Scholar] [CrossRef]
- Furukawa, M.; Tanaka, R.; Chuang, V.T.; Ishima, Y.; Taguchi, K.; Watanabe, H.; Maruyama, T.; Otagiri, M. Human serum albumin-thioredoxin fusion protein with long blood retention property is effective in suppressing lung injury. J. Control. Release Off. J. Control. Release Soc. 2011, 154, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kodama, A.; Watanabe, H.; Tanaka, R.; Tanaka, H.; Chuang, V.T.; Miyamoto, Y.; Wu, Q.; Endo, M.; Hamasaki, K.; Ishima, Y.; et al. A human serum albumin-thioredoxin fusion protein prevents experimental contrast-induced nephropathy. Kidney Int. 2013, 83, 446–454. [Google Scholar] [CrossRef]
- Tanaka, K.I.; Shimoda, M.; Chuang, V.T.G.; Nishida, K.; Kawahara, M.; Ishida, T.; Otagiri, M.; Maruyama, T.; Ishima, Y. Thioredoxin-albumin fusion protein prevents copper enhanced zinc-induced neurotoxicity via its antioxidative activity. Int. J. Pharm. 2018, 535, 140–147. [Google Scholar] [CrossRef]
- Nishida, K.; Watanabe, H.; Murata, R.; Tokumaru, K.; Fujimura, R.; Oshiro, S.; Nagasaki, T.; Miyahisa, M.; Hiramoto, Y.; Nosaki, H.; et al. Recombinant Long-Acting Thioredoxin Ameliorates AKI to CKD Transition via Modulating Renal Oxidative Stress and Inflammation. Int. J. Mol. Sci. 2021, 22, 5600. [Google Scholar] [CrossRef]
- Das, K.C. Thioredoxin-deficient mice, a novel phenotype sensitive to ambient air and hypersensitive to hyperoxia-induced lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L429–L442. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.I.; Chin, A.; Park, J.Y.; Ikeda, S.; Mizushima, W.; Ralda, G.; Zhai, P.; Tong, M.; Byun, J.; Tang, F.; et al. Thioredoxin-1 maintains mitochondrial function via mechanistic target of rapamycin signalling in the heart. Cardiovasc. Res. 2020, 116, 1742–1755. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Yamato, E.; Toyoda, S.; Tashiro, F.; Ikegami, H.; Yodoi, J.; Miyazaki, J. Transgenic expression of antioxidant protein thioredoxin in pancreatic beta cells prevents progression of type 2 diabetes mellitus. Antioxid. Redox Signal. 2008, 10, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Miyata, S.; Nii-Kono, T.; Kitazawa, R.; Kitazawa, S.; Higo, S.; Fukunaga, M.; Ueyama, S.; Nakamura, H.; Yodoi, J.; et al. Overexpression of thioredoxin1 in transgenic mice suppresses development of diabetic nephropathy. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2007, 22, 1547–1557. [Google Scholar] [CrossRef]
- Hamada, Y.; Fujii, H.; Kitazawa, R.; Yodoi, J.; Kitazawa, S.; Fukagawa, M. Thioredoxin-1 overexpression in transgenic mice attenuates streptozotocin-induced diabetic osteopenia: A novel role of oxidative stress and therapeutic implications. Bone 2009, 44, 936–941. [Google Scholar] [CrossRef]
- Zhou, F.; Gomi, M.; Fujimoto, M.; Hayase, M.; Marumo, T.; Masutani, H.; Yodoi, J.; Hashimoto, N.; Nozaki, K.; Takagi, Y. Attenuation of neuronal degeneration in thioredoxin-1 overexpressing mice after mild focal ischemia. Brain Res. 2009, 1272, 62–70. [Google Scholar] [CrossRef]
- Kong, L.; Zhou, X.; Li, F.; Yodoi, J.; McGinnis, J.; Cao, W. Neuroprotective effect of overexpression of thioredoxin on photoreceptor degeneration in Tubby mice. Neurobiol. Dis. 2010, 38, 446–455. [Google Scholar] [CrossRef]
- Sánchez-Villamil, J.P.; D’Annunzio, V.; Finocchietto, P.; Holod, S.; Rebagliati, I.; Pérez, H.; Peralta, J.G.; Gelpi, R.J.; Poderoso, J.J.; Carreras, M.C. Cardiac-specific overexpression of thioredoxin 1 attenuates mitochondrial and myocardial dysfunction in septic mice. Int. J. Biochem. Cell Biol. 2016, 81, 323–334. [Google Scholar] [CrossRef]
- Sugimoto, M.; Inoue, T.; Takeshita, K.; Nakamura, H.; Yodoi, J. Effects of a new anti-rheumatic drug KE-298 and its active metabolite: KE-758 on secretion of thioredoxin and on the level of intracellular glutathione in human monocytes and T cells. Mol. Immunol. 2002, 38, 793–799. [Google Scholar] [CrossRef]
- Schroeder, A.; Warnken, U.; Röth, D.; Klika, K.D.; Vobis, D.; Barnert, A.; Bujupi, F.; Oberacker, T.; Schnölzer, M.; Nicolay, J.P.; et al. Targeting Thioredoxin-1 by dimethyl fumarate induces ripoptosome-mediated cell death. Sci. Rep. 2017, 7, 43168. [Google Scholar] [CrossRef]
- Zhang, X.; Bai, L.; Zhang, S.; Zhou, X.; Li, Y.; Bai, J. Trx-1 ameliorates learning and memory deficits in MPTP-induced Parkinson’s disease model in mice. Free Radic. Biol. Med. 2018, 124, 380–387. [Google Scholar] [CrossRef] [PubMed]
- González, R.; Rodríguez-Hernández, M.A.; Negrete, M.; Ranguelova, K.; Rossin, A.; Choya-Foces, C.; Cruz-Ojeda, P.; Miranda-Vizuete, A.; Martínez-Ruiz, A.; Rius-Pérez, S.; et al. Downregulation of thioredoxin-1-dependent CD95 S-nitrosation by Sorafenib reduces liver cancer. Redox Biol. 2020, 34, 101528. [Google Scholar] [CrossRef] [PubMed]
- López-Grueso, M.J.; González, R.; Muntané, J.; Bárcena, J.A.; Padilla, C.A. Thioredoxin Downregulation Enhances Sorafenib Effects in Hepatocarcinoma Cells. Antioxidants 2019, 8, 501. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberacker, T.; Kraft, L.; Schanz, M.; Latus, J.; Schricker, S. The Importance of Thioredoxin-1 in Health and Disease. Antioxidants 2023, 12, 1078. https://doi.org/10.3390/antiox12051078
Oberacker T, Kraft L, Schanz M, Latus J, Schricker S. The Importance of Thioredoxin-1 in Health and Disease. Antioxidants. 2023; 12(5):1078. https://doi.org/10.3390/antiox12051078
Chicago/Turabian StyleOberacker, Tina, Leonie Kraft, Moritz Schanz, Jörg Latus, and Severin Schricker. 2023. "The Importance of Thioredoxin-1 in Health and Disease" Antioxidants 12, no. 5: 1078. https://doi.org/10.3390/antiox12051078
APA StyleOberacker, T., Kraft, L., Schanz, M., Latus, J., & Schricker, S. (2023). The Importance of Thioredoxin-1 in Health and Disease. Antioxidants, 12(5), 1078. https://doi.org/10.3390/antiox12051078