

Resveratrol Supplementation in Obese Pregnant Rats Improves Maternal Metabolism and Prevents Increased Placental Oxidative Stress

 , ,
, ,  ,
,  ,
,  ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Standardization of Females Recruited for Breeding as Mothers
2.2. Maternal (F0) and Fetal Tissue Collection at 19dG
2.3. F0 Biochemical and Hormonal Analysis
2.4. F0 Liver Fat Content and Triglycerides Content
2.5. F0 Adipose Tissue Histology
2.6. F0 Adipocyte Size (AS) Distribution
2.7. F0 Liver Gene Expression by Reverse Transcription Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
2.8. Oxidative Stress Biomarkers in Liver and Placenta
2.8.1. Lipoperoxidation Assay
2.8.2. Carbonylated Proteins
2.8.3. 8-Oxo-2′-Deoxyguanosine (8-oxo-dG) Quantification
2.8.4. Reactive Oxygen Species (ROS) Assay
2.9. Superoxide Dismutase (SOD) Activity
2.10. Glutathione Peroxidase (GPx) Activity
2.11. Immunohistochemistry (Nitrotyrosine, SOD and GPx)
2.12. Statistical Analysis
3. Results
3.1. F0 Body Weight Prior and during Pregnancy
Placenta and Fetal Weight at 19dG
3.2. F0 Food and Calorie Intake during Pregnancy
3.3. F0 Body Weight and Fat Distribution at 19dG
3.4. F0 Metabolic Parameters at 19dG
3.5. F0 Adipocyte Size (AS) Distribution
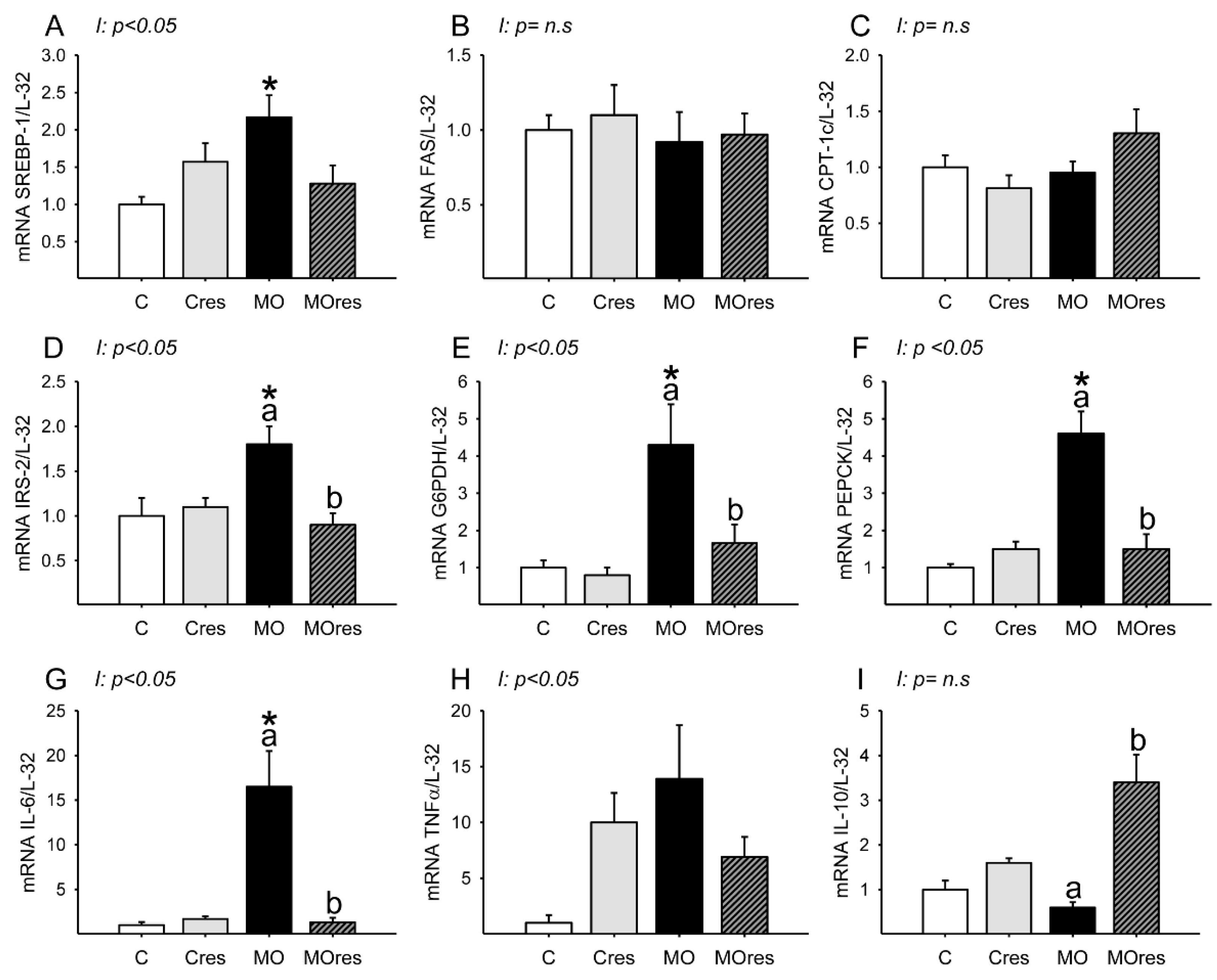
3.6. F0 Expression of Hepatic Metabolic and Inflammatory Genes
3.7. F0 Hepatic Oxidative Stress Biomarkers, Antioxidant Enzyme Activity and Gene Expression at 19dG
3.8. Placental Oxidative Stress Biomarkers and Antioxidant Enzyme Activity at 19dG
3.8.1. Male Placenta
3.8.2. Female Placenta
3.9. Fetal Hepatic Oxidative Stress Biomarkers and Antioxidant Enzyme Activity at 19dG
3.9.1. Male Liver
3.9.2. Female Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reichetzeder, C. Overweight and obesity in pregnancy: Their impact on epigenetics. Eur. J. Clin. Nutr. 2021, 75, 1710–1722. [Google Scholar] [CrossRef]
- Snelgrove-Clarke, E.; Macdonald, D.; Helwig, M.; Alsius, A. Women’s experiences of living with obesity during pregnancy, birthing, and postpartum: A qualitative systematic review protocol. JBI Evid. Synth. 2021, 19, 3183–3189. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.; Munoz, Y.; Ortiz, M.; Maliqueo, M.; Chouinard-Watkins, R.; Valenzuela, R. Impact of Maternal Obesity on the Metabolism and Bioavailability of Polyunsaturated Fatty Acids during Pregnancy and Breastfeeding. Nutrients 2020, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Mission, J.F.; Marshall, N.E.; Caughey, A.B. Pregnancy risks associated with obesity. Obstet. Gynecol. Clin. N. Am. 2015, 42, 335–353. [Google Scholar] [CrossRef] [PubMed]
- Downs, D.S. Obesity in Special Populations: Pregnancy. Prim. Care 2016, 43, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, E.; Nathanielsz, P.W. Mechanisms by which maternal obesity programs offspring for obesity: Evidence from animal studies. Nutr. Rev. 2013, 71 (Suppl. 1), S42–S54. [Google Scholar] [CrossRef]
- Zambrano, E.; Nathanielsz, P.W.; Rodriguez-Gonzalez, G.L. Developmental programming and ageing of male reproductive function. Eur. J. Clin. Investig. 2021, 51, e13637. [Google Scholar] [CrossRef]
- De Gusmao Correia, M.L.; Volpato, A.M.; Aguila, M.B.; Mandarim-de-Lacerda, C.A. Developmental origins of health and disease: Experimental and human evidence of fetal programming for metabolic syndrome. J. Hum. Hypertens. 2012, 26, 405–419. [Google Scholar] [CrossRef]
- Nathanielsz, P.W.; Ford, S.P.; Long, N.M.; Vega, C.C.; Reyes-Castro, L.A.; Zambrano, E. Interventions to prevent adverse fetal programming due to maternal obesity during pregnancy. Nutr. Rev. 2013, 71 (Suppl. 1), S78–S87. [Google Scholar] [CrossRef]
- Catalgol, B.; Batirel, S.; Taga, Y.; Ozer, N.K. Resveratrol: French paradox revisited. Front. Pharmacol. 2012, 3, 141. [Google Scholar] [CrossRef]
- Ros, P.; Argente, J.; Chowen, J.A. Effects of Maternal Resveratrol Intake on the Metabolic Health of the Offspring. Int. J. Mol. Sci. 2021, 22, 4792. [Google Scholar] [CrossRef] [PubMed]
- Vidavalur, R.; Otani, H.; Singal, P.K.; Maulik, N. Significance of wine and resveratrol in cardiovascular disease: French paradox revisited. Exp. Clin. Cardiol. 2006, 11, 217–225. [Google Scholar] [PubMed]
- Yao, L.; Wan, J.; Li, H.; Ding, J.; Wang, Y.; Wang, X.; Li, M. Resveratrol relieves gestational diabetes mellitus in mice through activating AMPK. Reprod. Biol. Endocrinol. 2015, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Zuo, Q.; Huang, S.; Yu, X.; Jiang, Z.; Zou, S.; Fan, M.; Sun, L. Resveratrol inhibits trophoblast apoptosis through oxidative stress in preeclampsia-model rats. Molecules 2014, 19, 20570–20579. [Google Scholar] [CrossRef] [PubMed]
- Poudel, R.; Stanley, J.L.; Rueda-Clausen, C.F.; Andersson, I.J.; Sibley, C.P.; Davidge, S.T.; Baker, P.N. Effects of resveratrol in pregnancy using murine models with reduced blood supply to the uterus. PLoS ONE 2013, 8, e64401. [Google Scholar] [CrossRef]
- Hsu, M.H.; Chen, Y.C.; Sheen, J.M.; Huang, L.T. Maternal Obesity Programs Offspring Development and Resveratrol Potentially Reprograms the Effects of Maternal Obesity. Int. J. Environ. Res. Public Health 2020, 17, 1610. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Developmental Programming of the Metabolic Syndrome: Can We Reprogram with Resveratrol? Int. J. Mol. Sci. 2018, 19, 2584. [Google Scholar] [CrossRef]
- Vega, C.C.; Reyes-Castro, L.A.; Rodriguez-Gonzalez, G.L.; Bautista, C.J.; Vazquez-Martinez, M.; Larrea, F.; Chamorro-Cevallos, G.A.; Nathanielsz, P.W.; Zambrano, E. Resveratrol partially prevents oxidative stress and metabolic dysfunction in pregnant rats fed a low protein diet and their offspring. J. Physiol. 2016, 594, 1483–1499. [Google Scholar] [CrossRef]
- Zheng, S.; Feng, Q.; Cheng, J.; Zheng, J. Maternal resveratrol consumption and its programming effects on metabolic health in offspring mechanisms and potential implications. Biosci. Rep. 2018, 38, BSR20171741. [Google Scholar] [CrossRef]
- Singh, C.K.; Kumar, A.; Hitchcock, D.B.; Fan, D.; Goodwin, R.; LaVoie, H.A.; Nagarkatti, P.; DiPette, D.J.; Singh, U.S. Resveratrol prevents embryonic oxidative stress and apoptosis associated with diabetic embryopathy and improves glucose and lipid profile of diabetic dam. Mol. Nutr. Food Res. 2011, 55, 1186–1196. [Google Scholar] [CrossRef]
- Trejo-Gonzalez, N.L.; Chirino-Galindo, G.; Palomar-Morales, M. Antiteratogenic capacity of resveratrol in streptozotocin-induced diabetes in rats. Rev. Peru. Med. Exp. Salud Publ. 2015, 32, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Grundy, D. Principles and standards for reporting animal experiments in The Journal of Physiology and Experimental Physiology. Exp. Physiol. 2015, 100, 755–758. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. Osteoarthr. Cartil. 2012, 20, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Gonzalez, G.L.; Reyes-Castro, L.A.; Bautista, C.J.; Beltran, A.A.; Ibanez, C.A.; Vega, C.C.; Lomas-Soria, C.; Castro-Rodriguez, D.C.; Elias-Lopez, A.L.; Nathanielsz, P.W.; et al. Maternal obesity accelerates rat offspring metabolic ageing in a sex-dependent manner. J. Physiol. 2019, 597, 5549–5563. [Google Scholar] [CrossRef]
- Cacho, J.; Sevillano, J.; de Castro, J.; Herrera, E.; Ramos, M.P. Validation of simple indexes to assess insulin sensitivity during pregnancy in Wistar and Sprague-Dawley rats. Am. J. Physiol.-Endocrinol. Metab. 2008, 295, E1269–E1276. [Google Scholar] [CrossRef]
- Vega, C.C.; Reyes-Castro, L.A.; Bautista, C.J.; Larrea, F.; Nathanielsz, P.W.; Zambrano, E. Exercise in obese female rats has beneficial effects on maternal and male and female offspring metabolism. Int. J. Obes. 2015, 39, 712–719. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef]
- Lecoutre, S.; Deracinois, B.; Laborie, C.; Eberle, D.; Guinez, C.; Panchenko, P.E.; Lesage, J.; Vieau, D.; Junien, C.; Gabory, A.; et al. Depot-and sex-specific effects of maternal obesity in offspring’s adipose tissue. J. Endocrinol. 2016, 230, 39–53. [Google Scholar] [CrossRef]
- Ibanez, C.A.; Vazquez-Martinez, M.; Leon-Contreras, J.C.; Reyes-Castro, L.A.; Rodriguez-Gonzalez, G.L.; Bautista, C.J.; Nathanielsz, P.W.; Zambrano, E. Different Statistical Approaches to Characterization of Adipocyte Size in Offspring of Obese Rats: Effects of Maternal or Offspring Exercise Intervention. Front. Physiol. 2018, 9, 1571. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Perez-Severiano, F.; Salvatierra-Sanchez, R.; Rodriguez-Perez, M.; Cuevas-Martinez, E.Y.; Guevara, J.; Limon, D.; Maldonado, P.D.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Santamaria, A. S-Allylcysteine prevents amyloid-beta peptide-induced oxidative stress in rat hippocampus and ameliorates learning deficits. Eur. J. Pharm. 2004, 489, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Mills, H.L.; Patel, N.; White, S.L.; Pasupathy, D.; Briley, A.L.; Santos Ferreira, D.L.; Seed, P.T.; Nelson, S.M.; Sattar, N.; Tilling, K.; et al. The effect of a lifestyle intervention in obese pregnant women on gestational metabolic profiles: Findings from the UK Pregnancies Better Eating and Activity Trial (UPBEAT) randomised controlled trial. BMC Med. 2019, 17, 15. [Google Scholar] [CrossRef]
- Groves, A.M.; Price, A.N.; Russell-Webster, T.; Jhaveri, S.; Yang, Y.; Battersby, E.E.; Shahid, S.; Costa Vieira, M.; Hughes, E.; Miller, F.; et al. Impact of maternal obesity on neonatal heart rate and cardiac size. Arch. Dis. Child.-Fetal Neonatal Ed. 2021, 107, 481–487. [Google Scholar] [CrossRef]
- Rodriguez-Gonzalez, G.L.; Castro-Rodriguez, D.C.; Zambrano, E. Pregnancy and Lactation: A Window of Opportunity to Improve Individual Health. In Investigations of Early Nutrition Effects on Long-Term Health; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1735, pp. 115–144. [Google Scholar] [CrossRef]
- Rodriguez, J.S.; Rodriguez-Gonzalez, G.L.; Reyes-Castro, L.A.; Ibanez, C.; Ramirez, A.; Chavira, R.; Larrea, F.; Nathanielsz, P.W.; Zambrano, E. Maternal obesity in the rat programs male offspring exploratory, learning and motivation behavior: Prevention by dietary intervention pre-gestation or in gestation. Int. J. Dev. Neurosci. 2012, 30, 75–81. [Google Scholar] [CrossRef]
- Zambrano, E.; Martinez-Samayoa, P.M.; Rodriguez-Gonzalez, G.L.; Nathanielsz, P.W. Dietary intervention prior to pregnancy reverses metabolic programming in male offspring of obese rats. J. Physiol. 2010, 588, 1791–1799. [Google Scholar] [CrossRef]
- Silva, L.; Pinheiro-Castro, N.; Novaes, G.M.; Pascoal, G.F.L.; Ong, T.P. Bioactive food compounds, epigenetics and chronic disease prevention: Focus on early-life interventions with polyphenols. Food Res. Int. 2019, 125, 108646. [Google Scholar] [CrossRef]
- Chan, R.S.; Woo, J. Prevention of overweight and obesity: How effective is the current public health approach. Int. J. Environ. Res. Public Health 2010, 7, 765–783. [Google Scholar] [CrossRef]
- Bautista, C.J.; Montano, S.; Ramirez, V.; Morales, A.; Nathanielsz, P.W.; Bobadilla, N.A.; Zambrano, E. Changes in milk composition in obese rats consuming a high-fat diet. Br. J. Nutr. 2016, 115, 538–546. [Google Scholar] [CrossRef]
- Mendez-del Villar, M.; Gonzalez-Ortiz, M.; Martinez-Abundis, E.; Perez-Rubio, K.G.; Lizarraga-Valdez, R. Effect of resveratrol administration on metabolic syndrome, insulin sensitivity, and insulin secretion. Metab. Syndr. Relat. Disord. 2014, 12, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Delpino, F.M.; Figueiredo, L.M.; Caputo, E.L.; Mintem, G.C.; Gigante, D.P. What is the effect of resveratrol on obesity? A systematic review and meta-analysis. Clin. Nutr. ESPEN 2021, 41, 59–67. [Google Scholar] [CrossRef]
- Malvasi, A.; Kosmas, I.; Mynbaev, O.A.; Sparic, R.; Gustapane, S.; Guido, M.; Tinelli, A. Can trans resveratrol plus d-chiro-inositol and myo-inositol improve maternal metabolic profile in overweight pregnant patients? Clin. Ter. 2017, 168, e240–e247. [Google Scholar] [CrossRef] [PubMed]
- Brawerman, G.M.; Kereliuk, S.M.; Brar, N.; Cole, L.K.; Seshadri, N.; Pereira, T.J.; Xiang, B.; Hunt, K.L.; Fonseca, M.A.; Hatch, G.M.; et al. Maternal resveratrol administration protects against gestational diabetes-induced glucose intolerance and islet dysfunction in the rat offspring. J. Physiol. 2019, 597, 4175–4192. [Google Scholar] [CrossRef]
- Castro-Rodriguez, D.C.; Reyes-Castro, L.A.; Vargas-Hernandez, L.; Itani, N.; Nathanielsz, P.W.; Taylor, P.D.; Zambrano, E. Maternal obesity (MO) programs morphological changes in aged rat offspring small intestine in a sex dependent manner: Effects of maternal resveratrol supplementation. Exp. Gerontol. 2021, 154, 111511. [Google Scholar] [CrossRef] [PubMed]
- Roy, H.J.; Keenan, M.J.; Zablah-Pimentel, E.; Hegsted, M.; Bulot, L.; O’Neil, C.E.; Bunting, L.D.; Fernandez, J.M. Adult female rats defend “appropriate” energy intake after adaptation to dietary energy. Obes. Res. 2003, 11, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef]
- Yang, X.; Li, L.; Xu, C.; Pi, M.; Wang, C.; Zhang, Y. Analysis of the different characteristics between omental preadipocytes and differentiated white adipocytes using bioinformatics methods. Adipocyte 2022, 11, 227–238. [Google Scholar] [CrossRef]
- Haider, N.; Larose, L. Harnessing adipogenesis to prevent obesity. Adipocyte 2019, 8, 98–104. [Google Scholar] [CrossRef]
- Vishvanath, L.; Gupta, R.K. Contribution of adipogenesis to healthy adipose tissue expansion in obesity. J. Clin. Investig. 2019, 129, 4022–4031. [Google Scholar] [CrossRef]
- Slawik, M.; Vidal-Puig, A.J. Lipotoxicity, overnutrition and energy metabolism in aging. Ageing Res. Rev. 2006, 5, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, Z.; Li, W.; Shan, Z.; Zhu, W. Resveratrol inhibits cell differentiation in 3T3-L1 adipocytes via activation of AMPK. Can. J. Physiol. Pharmacol. 2011, 89, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yu, R.; Xiong, Y.; Du, F.; Zhu, S. A vicious circle between insulin resistance and inflammation in nonalcoholic fatty liver disease. Lipids Health Dis. 2017, 16, 203. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Smith, G.I.; Shankaran, M.; Yoshino, M.; Schweitzer, G.G.; Chondronikola, M.; Beals, J.W.; Okunade, A.L.; Patterson, B.W.; Nyangau, E.; Field, T.; et al. Insulin resistance drives hepatic de novo lipogenesis in nonalcoholic fatty liver disease. J. Clin. Investig. 2020, 130, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Ferre, P.; Foufelle, F. SREBP-1c transcription factor and lipid homeostasis: Clinical perspective. Horm. Res. Paediatr. 2007, 68, 72–82. [Google Scholar] [CrossRef]
- Shimomura, I.; Bashmakov, Y.; Horton, J.D. Increased levels of nuclear SREBP-1c associated with fatty livers in two mouse models of diabetes mellitus. J. Biol. Chem. 1999, 274, 30028–30032. [Google Scholar] [CrossRef]
- Andrade, J.M.; Paraiso, A.F.; de Oliveira, M.V.; Martins, A.M.; Neto, J.F.; Guimaraes, A.L.; de Paula, A.M.; Qureshi, M.; Santos, S.H. Resveratrol attenuates hepatic steatosis in high-fat fed mice by decreasing lipogenesis and inflammation. Nutrition 2014, 30, 915–919. [Google Scholar] [CrossRef]
- Li, J.; Xue, Y.M.; Zhu, B.; Pan, Y.H.; Zhang, Y.; Wang, C.; Li, Y. Rosiglitazone Elicits an Adiponectin-Mediated Insulin-Sensitizing Action at the Adipose Tissue-Liver Axis in Otsuka Long-Evans Tokushima Fatty Rats. J. Diabetes Res. 2018, 2018, 4627842. [Google Scholar] [CrossRef]
- Michael, M.D.; Kulkarni, R.N.; Postic, C.; Previs, S.F.; Shulman, G.I.; Magnuson, M.A.; Kahn, C.R. Loss of insulin signaling in hepatocytes leads to severe insulin resistance and progressive hepatic dysfunction. Mol. Cell 2000, 6, 87–97. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Yang, J.; Zhou, S.; Wang, Y.; Li, Y.; Tong, X. The Role of the Pentose Phosphate Pathway in Diabetes and Cancer. Front. Endocrinol. 2020, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Choi, A.H.; Ham, M.; Kim, J.W.; Choe, S.S.; Park, J.; Lee, G.Y.; Yoon, K.H.; Kim, J.B. G6PD up-regulation promotes pancreatic β-cell dysfunction. Endocrinology 2011, 152, 793–803. [Google Scholar] [CrossRef]
- Riganti, C.; Gazzano, E.; Polimeni, M.; Aldieri, E.; Ghigo, D. The pentose phosphate pathway: An antioxidant defense and a crossroad in tumor cell fate. Free Radic. Biol. Med. 2012, 53, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Rattanatray, L.; Muhlhausler, B.S.; Nicholas, L.M.; Morrison, J.L.; McMillen, I.C. Impact of maternal overnutrition on gluconeogenic factors and methylation of the phosphoenolpyruvate carboxykinase promoter in the fetal and postnatal liver. Pediatr. Res. 2014, 75, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Gupte, R.S.; Floyd, B.C.; Kozicky, M.; George, S.; Ungvari, Z.I.; Neito, V.; Wolin, M.S.; Gupte, S.A. Synergistic activation of glucose-6-phosphate dehydrogenase and NAD(P)H oxidase by Src kinase elevates superoxide in type 2 diabetic, Zucker fa/fa, rat liver. Free Radic. Biol. Med. 2009, 47, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Rho, H.K.; Kim, K.H.; Choe, S.S.; Lee, Y.S.; Kim, J.B. Overexpression of glucose-6-phosphate dehydrogenase is associated with lipid dysregulation and insulin resistance in obesity. Mol. Cell. Biol. 2005, 25, 5146–5157. [Google Scholar] [CrossRef] [PubMed]
- Gupte, S.A. Glucose-6-phosphate dehydrogenase: A novel therapeutic target in cardiovascular diseases. Curr. Opin. Investig. Drugs 2008, 9, 993–1000. [Google Scholar]
- Nijland, M.J.; Mitsuya, K.; Li, C.; Ford, S.; McDonald, T.J.; Nathanielsz, P.W.; Cox, L.A. Epigenetic modification of fetal baboon hepatic phosphoenolpyruvate carboxykinase following exposure to moderately reduced nutrient availability. J. Physiol. 2010, 588, 1349–1359. [Google Scholar] [CrossRef]
- Yu, S.; Meng, S.; Xiang, M.; Ma, H. Phosphoenolpyruvate carboxykinase in cell metabolism: Roles and mechanisms beyond gluconeogenesis. Mol. Metab. 2021, 53, 101257. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, S.; Ferguson, S.; Wang, L.; Klepcyk, P.; Yun, J.S.; Friedman, J.E. Phosphoenolpyruvate carboxykinase overexpression selectively attenuates insulin signaling and hepatic insulin sensitivity in transgenic mice. J. Biol. Chem. 2002, 277, 23301–23307. [Google Scholar] [CrossRef] [PubMed]
- Valera, A.; Pujol, A.; Pelegrin, M.; Bosch, F. Transgenic mice overexpressing phosphoenolpyruvate carboxykinase develop non-insulin-dependent diabetes mellitus. Proc. Natl. Acad. Sci. USA 1994, 91, 9151–9154. [Google Scholar] [CrossRef] [PubMed]
- Elnagar, A.; El-Dawy, K.; El-Belbasi, H.I.; Rehan, I.F.; Embark, H.; Al-Amgad, Z.; Shanab, O.; Mickdam, E.; Batiha, G.E.; Alamery, S.; et al. Ameliorative Effect of Oxytocin on FBN1 and PEPCK Gene Expression, and Behavioral Patterns in Rats’ Obesity-Induced Diabetes. Front. Public Health 2022, 10, 777129. [Google Scholar] [CrossRef]
- O’Reilly, J.R.; Reynolds, R.M. The risk of maternal obesity to the long-term health of the offspring. Clin. Endocrinol. 2013, 78, 9–16. [Google Scholar] [CrossRef]
- King, J.C. Maternal obesity, metabolism, and pregnancy outcomes. Annu. Rev. Nutr. 2006, 26, 271–291. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, J.E.; Ferrell, W.R.; Crawford, L.; Wallace, A.M.; Greer, I.A.; Sattar, N. Maternal obesity is associated with dysregulation of metabolic, vascular, and inflammatory pathways. J. Clin. Endocrinol. Metab. 2002, 87, 4231–4237. [Google Scholar] [CrossRef]
- Charles, B.A.; Doumatey, A.; Huang, H.; Zhou, J.; Chen, G.; Shriner, D.; Adeyemo, A.; Rotimi, C.N. The roles of IL-6, IL-10, and IL-1RA in obesity and insulin resistance in African-Americans. J. Clin. Endocrinol. Metab. 2011, 96, E2018–E2022. [Google Scholar] [CrossRef] [PubMed]
- Bertola, A.; Bonnafous, S.; Anty, R.; Patouraux, S.; Saint-Paul, M.C.; Iannelli, A.; Gugenheim, J.; Barr, J.; Mato, J.M.; Le Marchand-Brustel, Y.; et al. Hepatic expression patterns of inflammatory and immune response genes associated with obesity and NASH in morbidly obese patients. PLoS ONE 2010, 5, e13577. [Google Scholar] [CrossRef]
- Hu, F.B.; Meigs, J.B.; Li, T.Y.; Rifai, N.; Manson, J.E. Inflammatory markers and risk of developing type 2 diabetes in women. Diabetes 2004, 53, 693–700. [Google Scholar] [CrossRef]
- Bravo-Flores, E.; Mancilla-Herrera, I.; Espino, Y.S.S.; Ortiz-Ramirez, M.; Flores-Rueda, V.; Ibarguengoitia-Ochoa, F.; Ibanez, C.A.; Zambrano, E.; Solis-Paredes, M.; Perichart-Perera, O.; et al. Macrophage Populations in Visceral Adipose Tissue from Pregnant Women: Potential Role of Obesity in Maternal Inflammation. Int. J. Mol. Sci. 2018, 19, 1074. [Google Scholar] [CrossRef]
- Radaelli, T.; Uvena-Celebrezze, J.; Minium, J.; Huston-Presley, L.; Catalano, P.; Hauguel-de Mouzon, S. Maternal interleukin-6: Marker of fetal growth and adiposity. J. Soc. Gynecol. Investig. 2006, 13, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Van Exel, E.; Gussekloo, J.; de Craen, A.J.; Frolich, M.; Bootsma-Van Der Wiel, A.; Westendorp, R.G.; Leiden 85 Plus, S. Low production capacity of interleukin-10 associates with the metabolic syndrome and type 2 diabetes: The Leiden 85-Plus Study. Diabetes 2002, 51, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 inhibits cytokine production by activated macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar] [PubMed]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Solis Paredes, J.M.; Perichart Perera, O.; Montoya Estrada, A.; Reyes Munoz, E.; Espino, Y.S.S.; Ortega Castillo, V.; Medina Bastidas, D.; Tolentino Dolores, M.; Sanchez Martinez, M.; Nava Salazar, S.; et al. Gestational Weight Gain Influences the Adipokine-Oxidative Stress Association during Pregnancy. Obes. Facts 2021, 14, 604–612. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Malti, N.; Merzouk, H.; Merzouk, S.A.; Loukidi, B.; Karaouzene, N.; Malti, A.; Narce, M. Oxidative stress and maternal obesity: Feto-placental unit interaction. Placenta 2014, 35, 411–416. [Google Scholar] [CrossRef]
- Alcala, M.; Gutierrez-Vega, S.; Castro, E.; Guzman-Gutierrez, E.; Ramos-Alvarez, M.P.; Viana, M. Antioxidants and Oxidative Stress: Focus in Obese Pregnancies. Front. Physiol. 2018, 9, 1569. [Google Scholar] [CrossRef]
- Li, S.; Eguchi, N.; Lau, H.; Ichii, H. The Role of the Nrf2 Signaling in Obesity and Insulin Resistance. Int. J. Mol. Sci. 2020, 21, 6973. [Google Scholar] [CrossRef]
- Perez-Cerezales, S.; Ramos-Ibeas, P.; Rizos, D.; Lonergan, P.; Bermejo-Alvarez, P.; Gutierrez-Adan, A. Early sex-dependent differences in response to environmental stress. Reproduction 2018, 155, R39–R51. [Google Scholar] [CrossRef]
- Myatt, L.; Muralimanoharan, S.; Maloyan, A. Effect of preeclampsia on placental function: Influence of sexual dimorphism, microRNA’s and mitochondria. In Advances in Fetal and Neonatal Physiology; Advances in Experimental Medicine and Biology; Springer Nature: London, UK, 2014; Volume 814, pp. 133–146. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; Rodríguez-Rodríguez, P.; de Pablo, Á.L.L.; López-Giménez, M.R.; González, M.C.; Arribas, S.M. Fetal Undernutrition and Oxidative Stress: Influence of Sex and Gender. In Handbook of Famine, Starvation, and Nutrient Deprivation: From Biology to Policy; Preedy, V., Patel, V.B., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–19. [Google Scholar] [CrossRef]
- Tondreau, M.Y.; Boucher, E.; Simard, M.; Tremblay, Y.; Bilodeau, J.F. Sex-specific perinatal expression of glutathione peroxidases during mouse lung development. Mol. Cell. Endocrinol. 2012, 355, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, S.A.; Wellen, K.E.; Henderson, G.C. Sexual dimorphism in the hepatic protein response to a moderate trans fat diet in senescence-accelerated mice. Lipids Health Dis. 2017, 16, 243. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jin, H.; Qiu, Y.; Liu, Y.; Wen, L.; Fu, Y.; Qi, H.; Baker, P.N.; Tong, C. Reactive Oxygen Species are Essential for Placental Angiogenesis During Early Gestation. Oxid. Med. Cell. Longev. 2022, 2022, 4290922. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Gene | Sequence |
|---|---|---|
| NM_001276707 | Sterol Regulatory Element-Binding Transcription Factor 1 (SREBP-1) | F: 5′-TGCGCAAGACAGCAGATTTA-3′ R: 5′-ACAAGATTGTGGAGCTCAAGG-3′ |
| NM_017332 | Fatty acid synthase (FAS) | F: 5′-GGCCACCTCAGTCCTGTTAT-3′ R: 5′-AGGGTCCAGCTAGAGGGTACA-3′ |
| NM_001034925 | Carnitine palmitoyltransferase 1C (CPT-1c) | F: 5′-TGTCCACAATTACCCGGATT-3′ R: 5′-GACGCCATACCCATGGTC-3′ |
| NM_001168633 | Insulin receptor substrate 2 (IRS-2) | F: 5′-CCAGGCACTGGAGCCTTA-3′ R: 5′-GCCCGCAGCACTTTACTC-3′ |
| NM_017006 | Glucose-6-phosphate dehydrogenase (G6PDH) | F: 5′-TTATCATCATGGGTGCATCG-3′ R: 5′-AAGGTGTCTTCGGGTAGAAGG-3′ |
| NM_001108377 | Phosphoenolpyruvate carboxykinase 2 (PEPCK2) | F: 5′-CCGACTGCACTGGTTCCT-3′ R: 5′-TCAGCCTGTGCCAGCTAAG-3′ |
| NM_012589 | Interleukin 6 (IL-6) | F: 5′-CCACTGCCTTCCCTACTTCA-3′ R: 5′-CTGGTCTGTTGTGGGTGGTA-3′ |
| NM_012675 | Tumor necrosis factor-alpha (TNF-alpha) | F: 5′-CACTTGGCTGAGAGGAAAGG-3′ R: 5′-CAAATGAGTGTCCCGCAGA-3′ |
| NM_012854 | Interleukin 10 (IL-10) | F: 5′-CAGATTCCTTACTGCAGGACTTTA-3′ R: 5′-CAAATGCTCCTTGATTTCTGG-3′ |
| NM_031789 | Nuclear factor erythroid 2–related factor 2 (Nrf2) | F: 5′-CAACAGTATTTCTGCCGCTGT-3′ R: 5′-CACAGGGAGGACTTTGTGAGT-3′ |
| NM_017050 | Cu/Zn cytosolic superoxide dismutase-1 (SOD-1) | F: 5′-GGTCCAGCGGATGAAGAG-3′ R: 5′-GGACACATTGGCCACACC-3′ |
| NM_030826 | Glutathione peroxidase-1 (GPX-1) | F: 5′-CGACATCGAACCCGATATAGA-3′ R: 5′-ATGCCTTAGGGGTTGCTAGG-3′ |
| NM_013226 | Ribosomal protein L32 (Rn-L32) | F: 5′-CCGGAAGTTTCTGGTCCAC-3′ R: 5′-CAGCACAGTAAGATTTGTTGCAC-3′ |
| C | Cres | MO | MOres | |
|---|---|---|---|---|
| Body weight (g) | 452 ± 7 | 429 ± 8 | 537 ± 14 * | 514 ± 16 * |
| Total fat (g) | 20.3 ± 2.4 | 12.4 ± 1 | 56.5 ± 5.5 * | 50.7 ± 4.9 * |
| Adiposity index | 4.5 ± 0.5 | 2.9 ± 0.3 | 10.5 ± 1 * | 9.8 ± 0.9 * |
| Fat depot (g) | ||||
| Mediastinal | 0.3 ± 0.07 | 0.3 ± 0.03 | 3.8 ± 2.7 | 1 ± 0.2 |
| Retroperitoneal and perirenal | 3.4 ± 0.5 | 2.7 ± 0.2 | 14.5 ± 1.2 * | 14.1 ± 1.2 * |
| Omental | 3.8 ± 0.5 | 3.9 ± 0.6 | 15.6 ± 3.2 * | 13.6 ± 2.1 * |
| Parametrial and perivescical | 4.3 ± 0.8 | 2.9 ± 0.2 | 11.9 ± 1.2 * | 11.8 ± 1.3 * |
| Periovarian | 2.8 ± 0.3 | 2.6 ± 0.3 | 10.8 ± 0.6 * | 10.1 ± 0.8 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-González, G.L.; Vargas-Hernández, L.; Reyes-Castro, L.A.; Ibáñez, C.A.; Bautista, C.J.; Lomas-Soria, C.; Itani, N.; Estrada-Gutierrez, G.; Espejel-Nuñez, A.; Flores-Pliego, A.; et al. Resveratrol Supplementation in Obese Pregnant Rats Improves Maternal Metabolism and Prevents Increased Placental Oxidative Stress. Antioxidants 2022, 11, 1871. https://doi.org/10.3390/antiox11101871
Rodríguez-González GL, Vargas-Hernández L, Reyes-Castro LA, Ibáñez CA, Bautista CJ, Lomas-Soria C, Itani N, Estrada-Gutierrez G, Espejel-Nuñez A, Flores-Pliego A, et al. Resveratrol Supplementation in Obese Pregnant Rats Improves Maternal Metabolism and Prevents Increased Placental Oxidative Stress. Antioxidants. 2022; 11(10):1871. https://doi.org/10.3390/antiox11101871
Chicago/Turabian StyleRodríguez-González, Guadalupe L., Lilia Vargas-Hernández, Luis A. Reyes-Castro, Carlos A. Ibáñez, Claudia J. Bautista, Consuelo Lomas-Soria, Nozomi Itani, Guadalupe Estrada-Gutierrez, Aurora Espejel-Nuñez, Arturo Flores-Pliego, and et al. 2022. "Resveratrol Supplementation in Obese Pregnant Rats Improves Maternal Metabolism and Prevents Increased Placental Oxidative Stress" Antioxidants 11, no. 10: 1871. https://doi.org/10.3390/antiox11101871
APA StyleRodríguez-González, G. L., Vargas-Hernández, L., Reyes-Castro, L. A., Ibáñez, C. A., Bautista, C. J., Lomas-Soria, C., Itani, N., Estrada-Gutierrez, G., Espejel-Nuñez, A., Flores-Pliego, A., Montoya-Estrada, A., Reyes-Muñoz, E., Taylor, P. D., Nathanielsz, P. W., & Zambrano, E. (2022). Resveratrol Supplementation in Obese Pregnant Rats Improves Maternal Metabolism and Prevents Increased Placental Oxidative Stress. Antioxidants, 11(10), 1871. https://doi.org/10.3390/antiox11101871