
Effects of Compound Polysaccharides Derived from Astragalus and Glycyrrhiza on Growth Performance, Meat Quality and Antioxidant Function of Broilers Based on Serum Metabolomics and Cecal Microbiota

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Astragalus Polysaccharides and Glycyrrhiza Polysaccharides
2.2. Experimental Design and Animal Management
2.3. Growth Performance
2.4. Serum and Breast Muscle Antioxidant Function
2.5. Meat Quality
2.6. qRT-PCR of Intestinal Antioxidant Enzyme Genes
2.7. Cecal Microbiota
2.8. Metabolite Extraction and Analysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Apparent Metabolic Rate
3.3. Meat Quality
3.4. Breast Muscle Fatty Acid
3.5. Serum and Breast Muscle Antioxidant Function
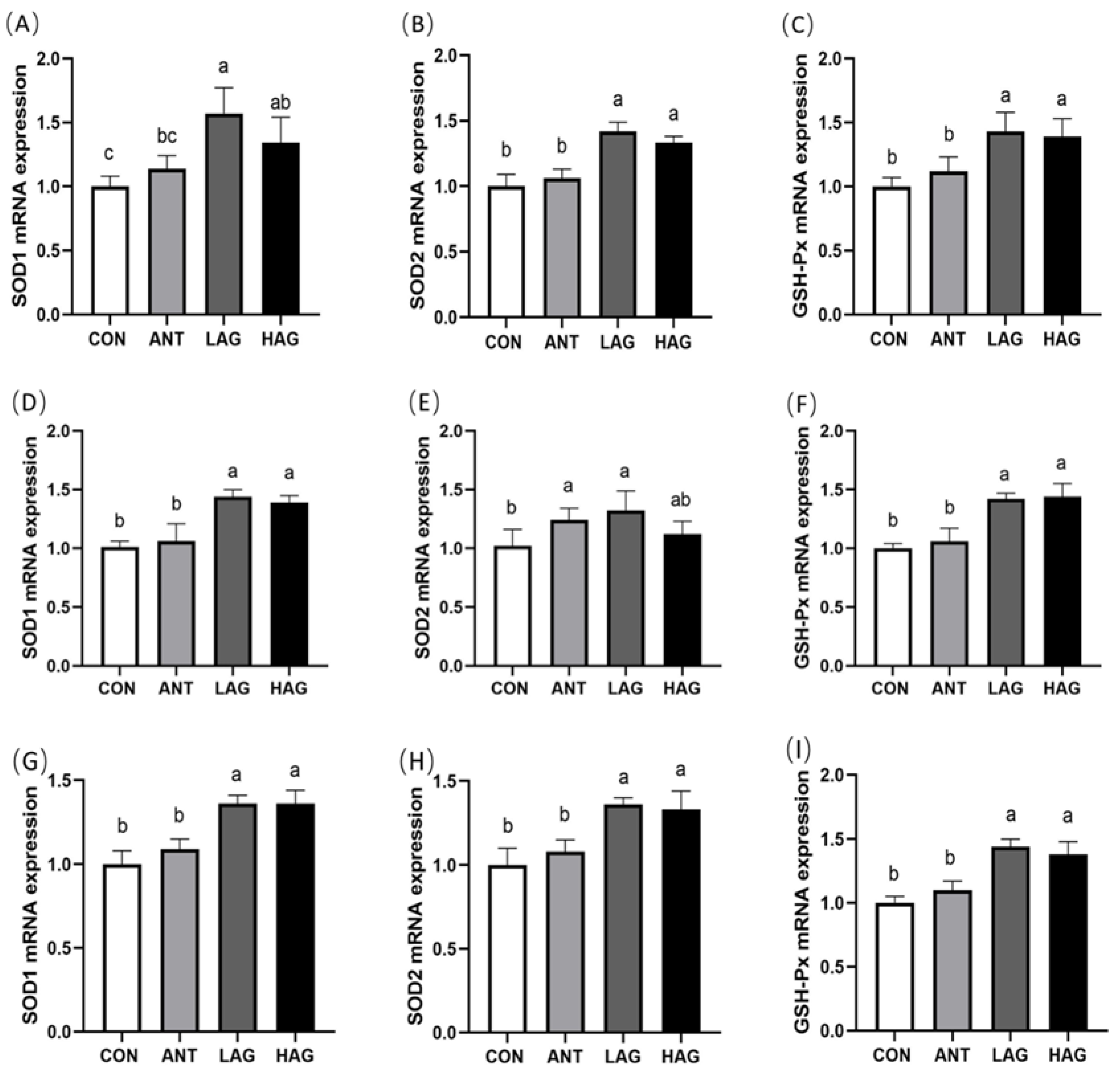
3.6. Intestine Antioxidant Enzyme mRNA Expression
3.7. Cecal Microbial Diversity
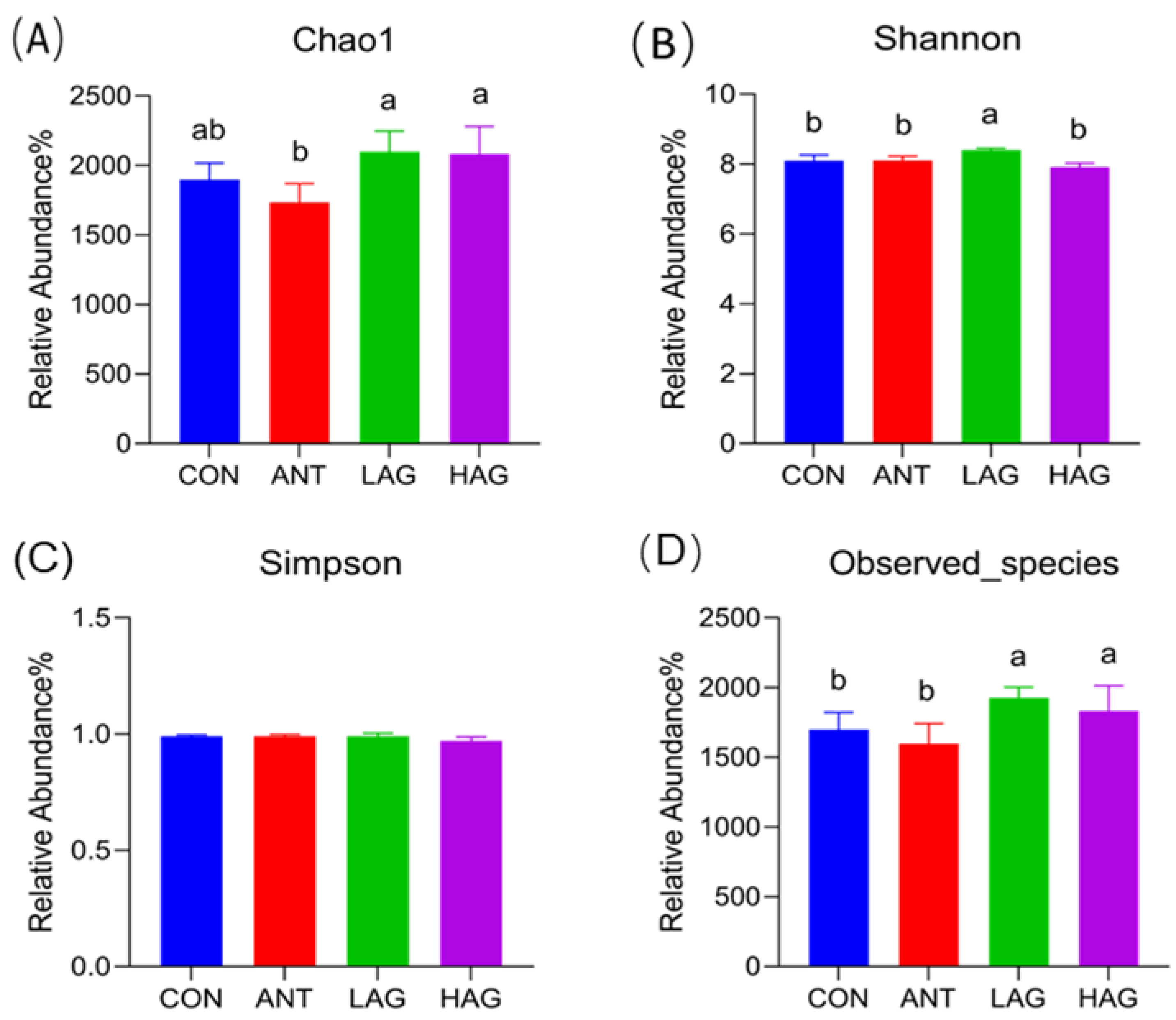
3.7.1. Alpha Diversity Analysis
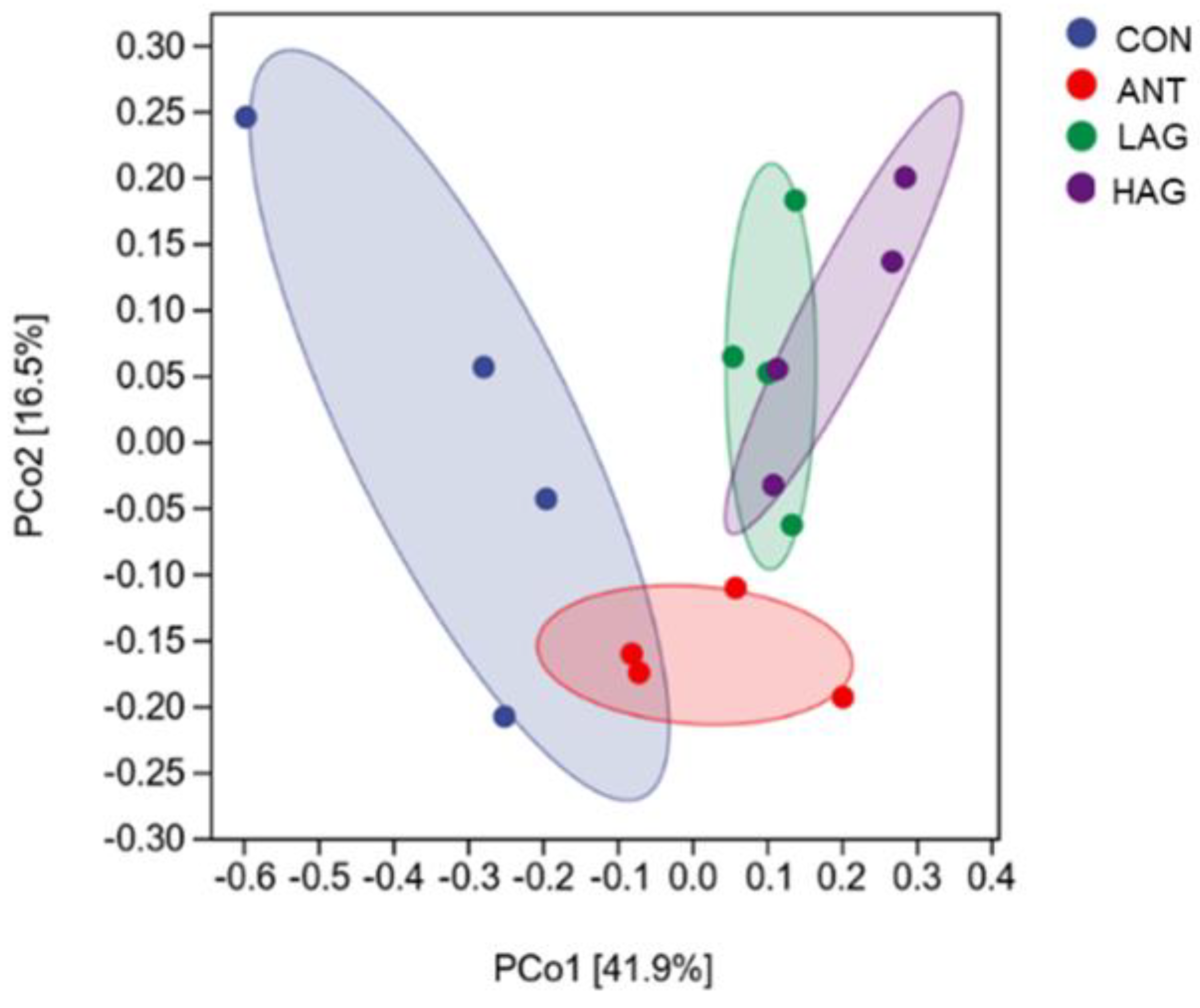
3.7.2. Beta Diversity Analysis
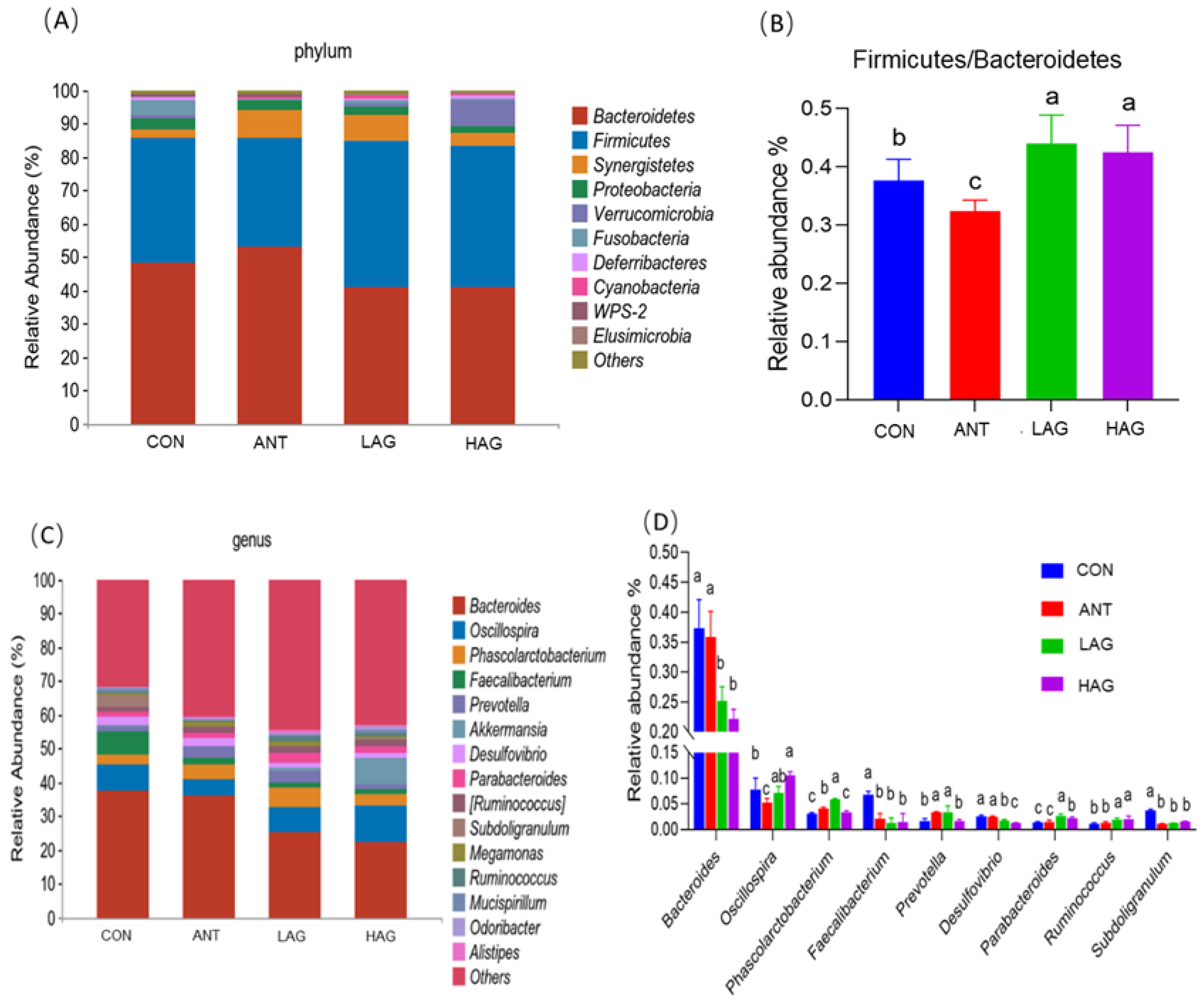
3.7.3. Microflora Structure
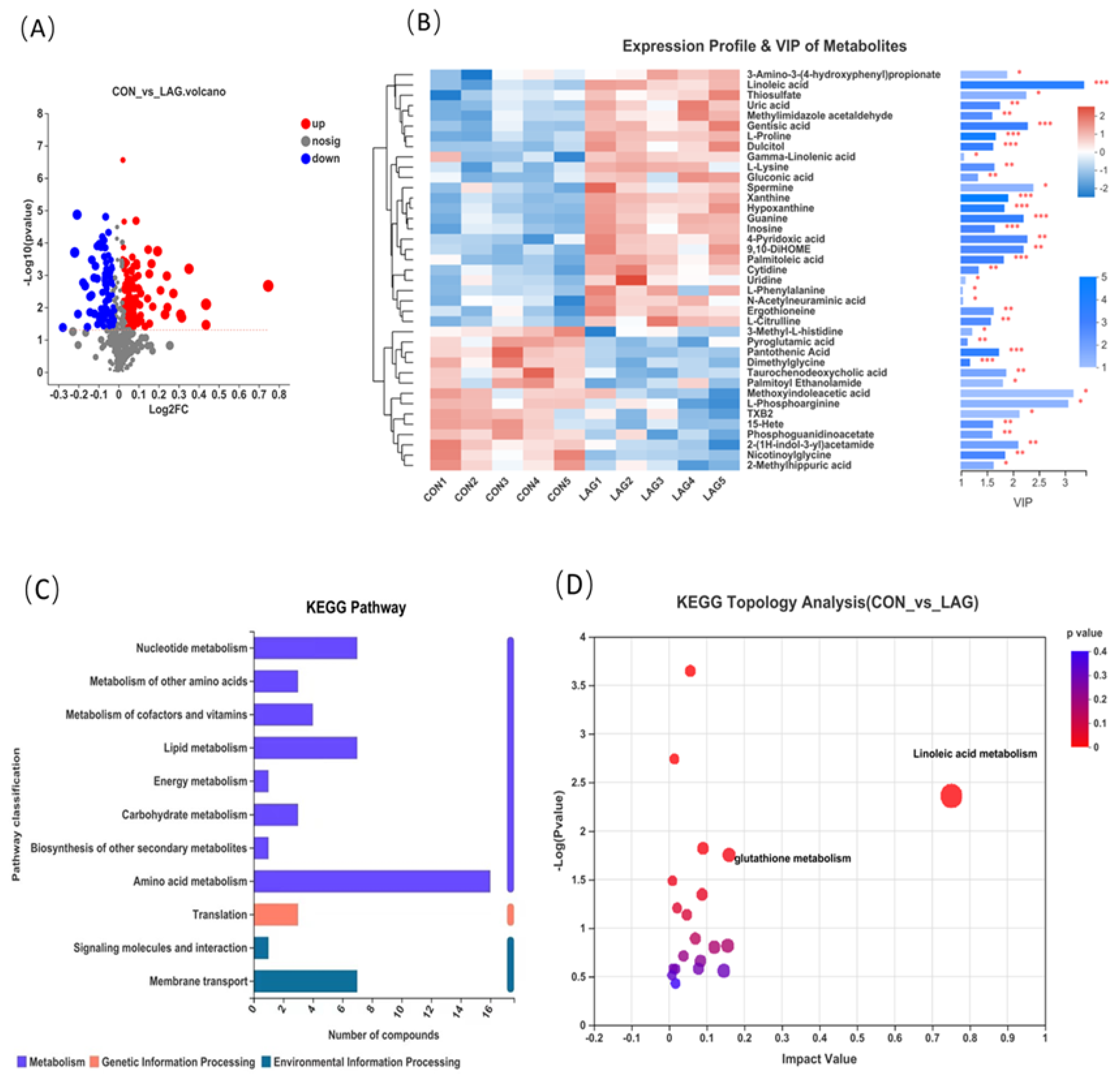
3.8. Serum Metabolomic Analysis
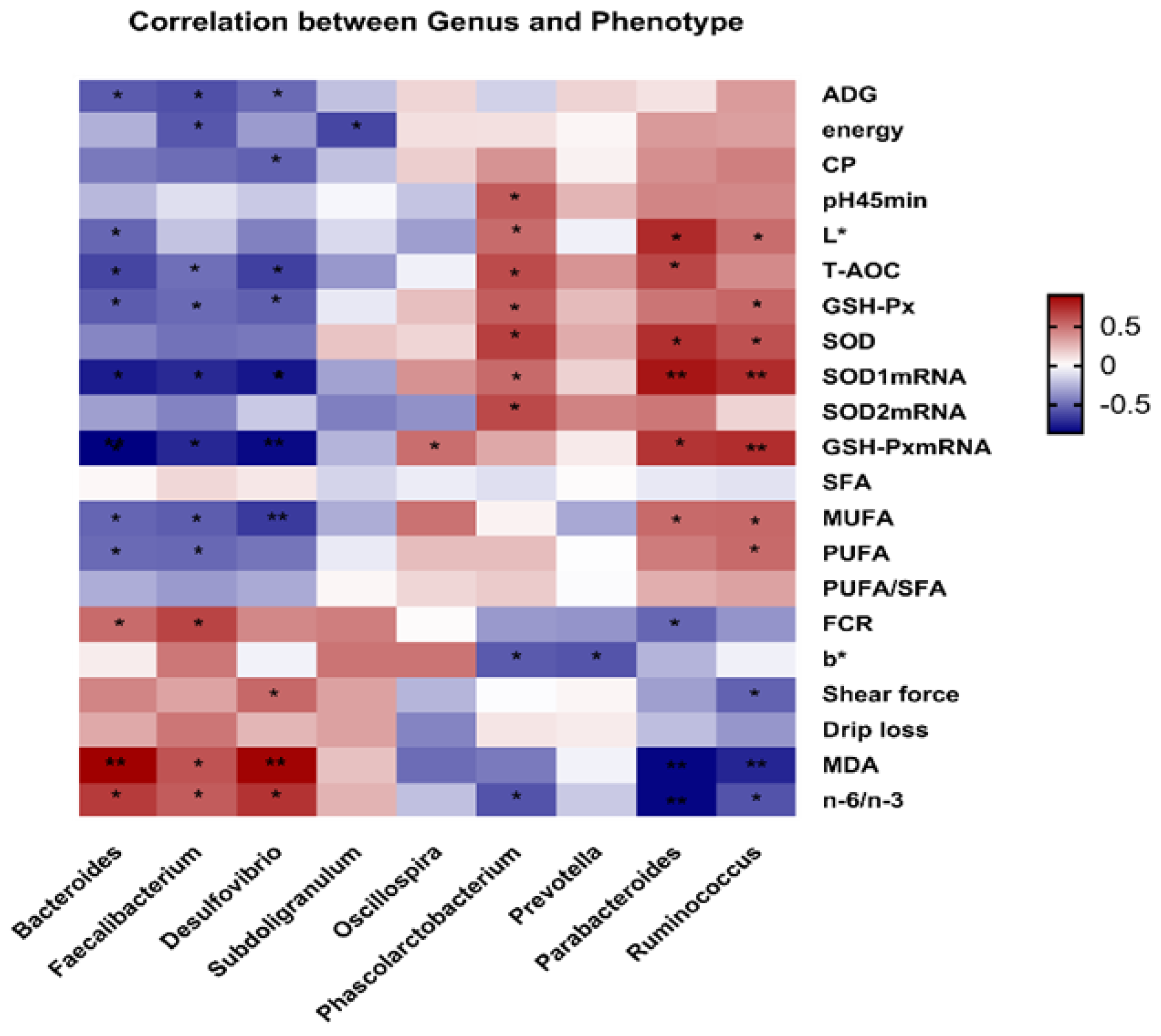
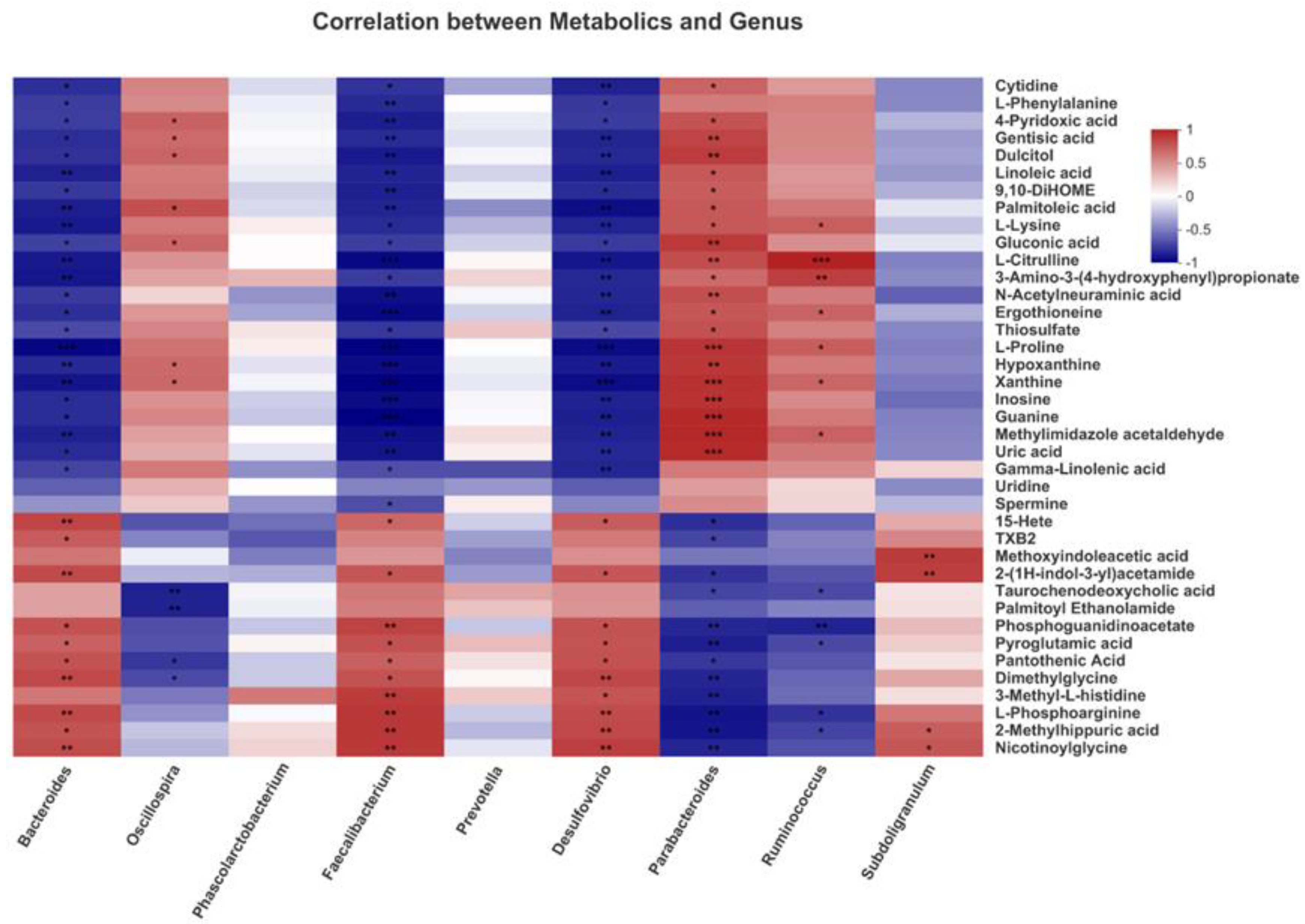
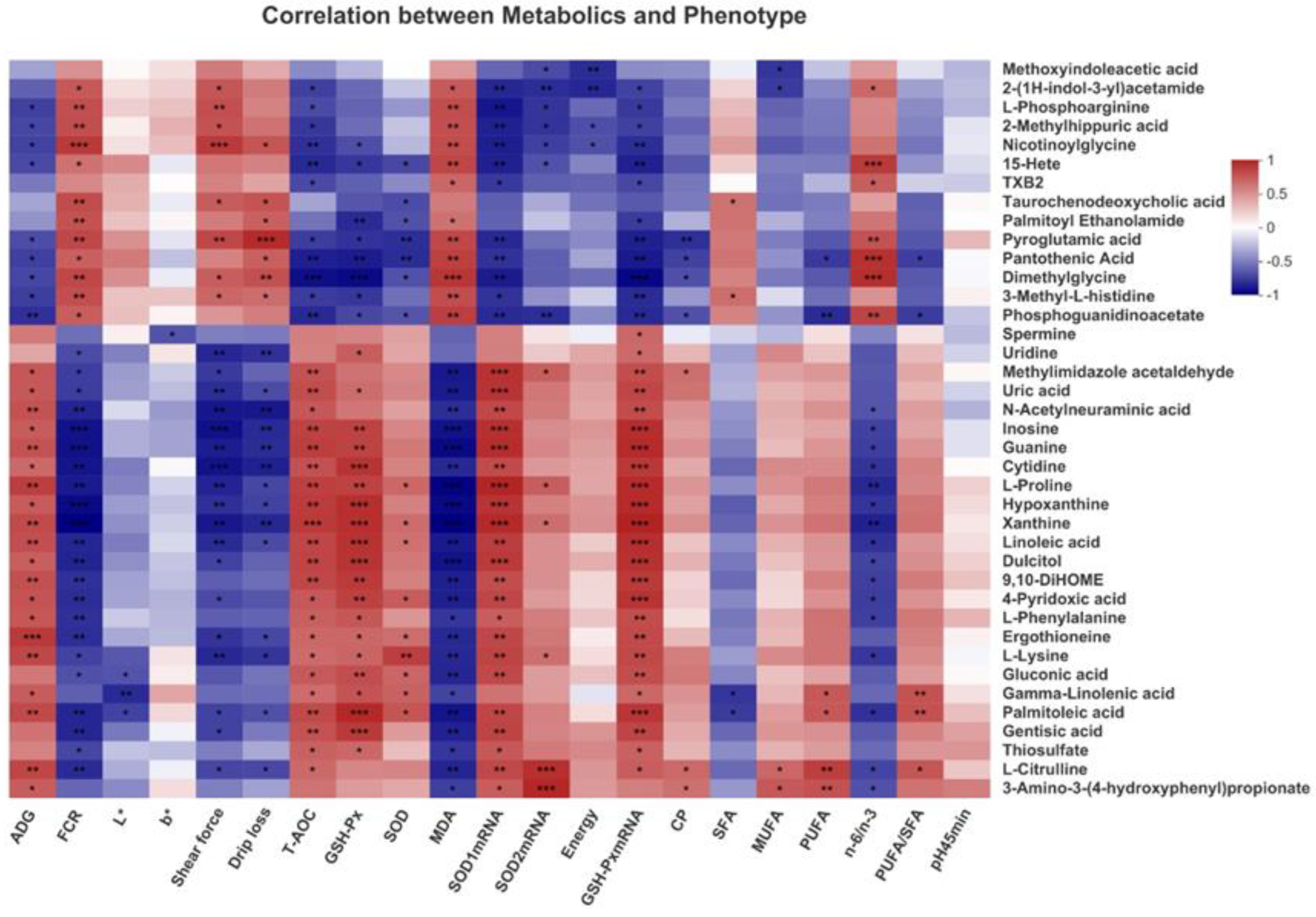
3.9. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Boeckel, T.P.; Glennon, E.E.; Chen, D.; Gilbert, M.; Robinson, T.P.; Grenfell, B.T.; Levin, S.A.; Bonhoeffer, S.; Laxminarayan, R. Reducing antimicrobial use in food animals. Science 2017, 357, 1350. [Google Scholar] [CrossRef]
- Suresh, G.; Das, R.K.; Kaur Brar, S.; Rouissi, T.; Avalos Ramirez, A.; Chorfi, Y.; Godbout, S. Alternatives to antibiotics in poultry feed: Molecular perspectives. Crit. Rev. Microbiol. 2018, 44, 318. [Google Scholar] [CrossRef]
- Haque, M.; Sarker, S.; Islam, M.; Karim, M.; Kayesh, M.E.H.; Shiddiky, M.J.; Anwer, M.S. Sustainable Antibiotic-Free Broiler Meat Production: Current Trends, Challenges, and Possibilities in a Developing Country Perspective. Biology 2020, 9, 411. [Google Scholar] [CrossRef]
- Salim, H.M.; Huque, K.S.; Kamaruddin, K.M.; Haque Beg, A. Global restriction of using antibiotic growth promoters and alternative strategies in poultry production. Sci. Prog. 2018, 101, 52. [Google Scholar] [CrossRef]
- Zhang, X.H.; Sun, Z.Y.; Cao, F.L.; Ahmad, H.; Yang, X.H.; Zhao, L.G.; Wang, T. Effects of dietary supplementation with fermented ginkgo leaves on antioxidant capacity, intestinal morphology and microbial ecology in broiler chicks. Br. Poult. Sci. 2015, 56, 370. [Google Scholar] [CrossRef]
- Diaz-Sanchez, S.; D’Souza, D.; Biswas, D.; Hanning, I. Botanical alternatives to antibiotics for use in organic poultry production. Poult. Sci. 2015, 94, 1419. [Google Scholar] [CrossRef]
- Abolhasani, A.; Barzegar, M.; Sahari, M.A. Effect of gamma irradiation on the extraction yield, antioxidant, and antityrosinase activities of pistachio green hull extract. Radiat. Phys. Chem. 2018, 144, 373. [Google Scholar] [CrossRef]
- Hamed, M.; Bougatef, H.; Karoud, W.; Krichen, F.; Haddar, A.; Bougatef, A.; Sila, A. Polysaccharides extracted from pistachio external hull: Characterization, antioxidant activity and potential application on meat as preservative. Ind. Crops Prod. 2020, 148, 112315. [Google Scholar] [CrossRef]
- Barreca, D.; Laganà, G.; Leuzzi, U.; Smeriglio, A.; Trombetta, D.; Bellocco, E. Evaluation of the nutraceutical, antioxidant and cytoprotective properties of ripe pistachio (Pistacia vera L., variety Bronte) hulls. Food Chem. 2016, 196, 493. [Google Scholar] [CrossRef]
- Zhao, Z.T.; Ye, X.M.; Ouyang, K.H.; Wang, W.J. Effects of polysaccharides from Yingshan Yunwu tea on meat quality, immune status and intestinal microflora in chickens. Int. J. Biol. Macromol. 2020, 155, 61. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G. Antioxidant activity of polysaccharides from different sources of ginseng. Int. J. Biol. Macromol. 2019, 125, 906. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ren, W.; Zhang, L.; Zhang, Y.; Liu, D.; Liu, Y. A review of the pharmacological action of Astragalus polysaccharide. Front. Pharmacol. 2020, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Wu, S. Effect of dietary Astragalus membranaceus polysaccharide on the growth performance and immunity of juvenile broilers. Poult. Sci. 2018, 97, 3489. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, X.F.; Xing, T.; Li, J.L.; Zhu, X.D.; Zhang, L.; Gao, F. The combined impact of xylo-oligosaccharides and gamma-irradiated Astragalus polysaccharides on growth performance and intestinal mucosal barrier function of broilers. Poult. Sci. 2021, 100, 100909. [Google Scholar] [CrossRef]
- Shan, L.; Ren, L.; Zhu, X.; Li, J.; Zhou, G. Immunomodulatory effect of γ-irradiated astragalus polysaccharides on immunosuppressed broilers. Anim. Sci. J. 2018, 90, 13133. [Google Scholar] [CrossRef]
- Liu, L.; Shen, J.; Zhao, C.; Wang, X.; Yao, J.; Gong, Y.; Yang, X. Dietary Astragalus polysaccharide alleviated immunological stress in broilers exposed to lipopolysaccharide. Int. J. Biol. Macromol. 2015, 72, 624. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Li, B.; Zheng, N.; Wu, G.; Ma, J.; Tao, X.; Chen, L.; Zhong, J.; Sheng, L.; Li, H. Integrated metagenomic and metabolomic analyses of the effect of astragalus polysaccharides on alleviating high-fat diet–induced metabolic disorders. Front. Pharmacol. 2020, 11, 833. [Google Scholar] [CrossRef]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M.B.P. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.X.; Shao, Q.; Chen, W.B.; Ma, L.; Xu, W.H.; Ma, Y.B. Effects of Glycyrrhiza polysaccharide in diet on growth performance, serum antioxidant capacity, and biochemistry of broilers. Poult. Sci. 2021, 100, 25. [Google Scholar] [CrossRef]
- Qiao, Y.; Liu, C.; Guo, Y.; Zhang, W.; Guo, W.; Oleksandr, K.; Wang, Z. Polysaccharides derived from Astragalus membranaceus and Glycyrrhiza uralensis improve growth performance of broilers by enhancing intestinal health and modulating gut microbiota. Poult. Sci. 2022, 101, 101905. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R. Use of liquorice (Glycyrrhiza glabra) in poultry nutrition: Global impacts on performance, carcass and meat quality. World’s Poult. Sci. J. 2019, 75, 293. [Google Scholar] [CrossRef]
- GB 5009.168-2016.2016-12-23; National Food Safety Standard Determination of Fatty Acids in Food. Standards Press of China: Beijing, China, 2016. (In Chinese)
- Wallace, R.J.; Oleszek, W.; Franz, C.; Hahn, I.; Baser, K.H.C.; Mathe, A.; Teichmann, K. Dietary plant bioactives for poultry health and productivity. Br. Poult. Sci. 2010, 51, 461. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.F.; Yang, W.R.; Yang, H.W.; Wang, Y.; Yang, Z.B.; Jiang, S.Z.; Zhang, G.G. Effects of Astragalus membranaceus on growth performance, carcass characteristics, and antioxidant status of broiler chickens. Acta Agric. Scand. Sect. A 2010, 60, 151. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Abdelnour, S.A.; Taha, A.E.; Khafaga, A.F.; Arif, M.; Ayasan, T.; Abdel-Daim, M.M. Herbs as thermoregulatory agents in poultry: An overview. Sci. Total Environ. 2020, 703, 134399. [Google Scholar] [CrossRef] [PubMed]
- Rashidi, N.; Ghorbani, M.R.; Tatar, A.; Salari, S. Response of broiler chickens reared at high density to dietary supplementation with licorice extract and probiotic. J. Anim. Physiol. Anim. Nutr. 2019, 103, 100. [Google Scholar] [CrossRef]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; Abd El-Fattah, A.H.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of Glycyrrhiza glabra extract on growth, gene expression of gut integrity, and Campylobacter jejuni colonization in broiler chickens. Front. Vet. Sci. 2020, 22, 612063. [Google Scholar] [CrossRef]
- Ali, M.; Kang, G.H.; Joo, S.T. A review: Influences of pre-slaughter stress on poultry meat quality. Asian-Australas. J. Anim. Sci. 2008, 21, 912. [Google Scholar] [CrossRef]
- Galli, G.M.; Aniecevski, E.; Petrolli, T.G.; da Rosa, G.; Boiago, M.M.; Simões, C.A.; Da Silva, A.S. Growth performance and meat quality of broilers fed with microencapsulated organic acids. Anim. Feed Sci. Technol. 2021, 271, 114706. [Google Scholar] [CrossRef]
- Kim, T.W.; Kim, C.W.; Kwon, S.G.; Hwang, J.H.; Park, D.H.; Kang, D.G.; Kim, I.S. pH as analytical indicator for managing pork meat quality. Sains Malays. 2016, 45, 1097–1103. [Google Scholar]
- Zhang, X.; Shi, H.; Niu, X.; Song, Y.; Tang, F.; Bian, C.; Qiao, H. Effect of aqueous extract of fermented Astragalia and Glycyrrhizaon on growth performance, immune organ indexes and meat quality of 817 broilers. China Anim. Husb. Vet. Med. 2018, 145, 933–939. (In Chinese) [Google Scholar] [CrossRef]
- Brewer, S. Irradiation effects on meat color–a review. Meat Sci. 2004, 68, 1. [Google Scholar] [CrossRef]
- Mingyuan, W.; Xiping, Y.; Xi, Z.; Bizhi, H.; Jianyong, J.; Zhe, W. Study of banana stems and leaves silage on beef quality and growth performance of Yunnan yellow cattle. Feed Ind. 2015, 36, 12. (In Chinese) [Google Scholar] [CrossRef]
- Oh, H.J.; Song, M.H.; Yun, W.; Lee, J.H.; An, J.S.; Kim, Y.J.; Cho, J.H. Effects of replacing soybean meal with perilla seed meal on growth performance, and meat quality of broilers. J. Anim. Sci. Technol. 2020, 62, 495. [Google Scholar] [CrossRef]
- Manilla, H.; Husvéth, F. N-3 fatty acid enrichment and oxidative stability of broiler chicken (A review). Acta Aliment. 1999, 28, 235. [Google Scholar] [CrossRef]
- Kouba, M.; Enser, M.; Whittington, F.M.; Nute, G.R.; Wood, J.D. Effect of a high-linolenic acid diet on lipogenic enzyme activities, fatty acid composition, and meat quality in the growing pig. J. Anim. Sci. 2003, 81, 1967. [Google Scholar] [CrossRef]
- Corino, C.; Rossi, R.; Cannata, S.; Ratti, S. Effect of dietary linseed on the nutritional value and quality of pork and pork products: Systematic review and meta-analysis. Meat Sci. 2014, 98, 679–688. [Google Scholar] [CrossRef]
- Hu, F.B.; Manson, J.E.; Willett, W.C. Types of dietary fat and risk of coronary heart disease: A critical review. J. Am. Coll. Nutr. 2001, 20, 5. [Google Scholar] [CrossRef]
- Siriwardhana, N.; Kalupahana, N.S.; Moustaid-Moussa, N. Health benefits of n-3 polyunsaturated fatty acids: Eicosapentaenoic acid and docosahexaenoic acid. Adv. Food Nutr. Res. 2012, 65, 211. [Google Scholar] [CrossRef]
- Alagawany, M.; Ashour, E.A.; El-Fakhrany, H.H.H.; Ismail, T.A.; Nasr, M. Early nutrition programming with Astragalus membranaceus polysaccharide: Its effect on growth, carcasses, immunity, antioxidants, lipid profile and liver and kidney functions in broiler chickens. Anim. Biotechnol. 2022, 33, 362–368. [Google Scholar] [CrossRef]
- Zhang, G.G.; Yang, Z.B.; Wang, Y.; Yang, W.R. Effects of Astragalus membranaceus root processed to different particle sizes on growth performance, antioxidant status, and serum metabolites of broiler chickens. Poult. Sci. 2013, 92, 178. [Google Scholar] [CrossRef] [PubMed]
- El-Shafei, A.A.; Al-Gamal, M.A.; Abdelrahman, A.S.; Arafa, M.M. Influence of different levels of astragalus root powder in broiler chick diets on the physiological and biochemical changes. J. Appl. Sci. Res. 2013, 9, 2104–2118. [Google Scholar]
- Aslam, M.N.; Sohaib, M.; Khan, A.U.; Ali, S.; Amjad, A.; Ahmed, S. Lipids Oxidative Stability and Microbial Shelf Life Quality of Licorice (Glycyrrhiza glabra L.) Extract Supplemented Chicken Patties. Braz. J. Poult. Sci. 2020, 22, 1316. [Google Scholar] [CrossRef]
- Hoerr, F.J. Clinical aspects of immunosuppression in poultry. Avian Dis. 2010, 54, 2–15. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, Y.; Liang, Y.; Jiao, X.; Zhao, C. Beneficial effect of intestinal fermentation of natural polysaccharides. Nutrients 2018, 10, 1055. [Google Scholar] [CrossRef]
- Liu, Y.S.; Li, S.; Wang, X.F.; Xing, T.; Li, J.L.; Zhu, X.D.; Gao, F. Microbiota populations and short-chain fatty acids production in cecum of immunosuppressed broilers consuming diets containing γ-irradiated Astragalus polysaccharides. Poult. Sci. 2021, 100, 273. [Google Scholar] [CrossRef]
- Downs, I.A.; Aroniadis, O.C.; Kelly, L.; Brandt, L.J. Postinfection irritable bowel syndrome. J. Clin. Gastroenterol. 2017, 51, 869. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Kwok, L.Y.; Zheng, Y.; Wang, L.; Guo, Z.; Zhang, J.; Zhang, H. Differential fecal microbiota are retained in broiler chicken lines divergently selected for fatness traits. Sci. Rep. 2016, 6, 1. [Google Scholar] [CrossRef]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Takase, K. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 1. [Google Scholar] [CrossRef]
- Houston, S.; Blakely, G.W.; McDowell, A.; Martin, L.; Patrick, S. Binding and degradation of fibrinogen by Bacteroides fragilis and characterization of a 54 kDa fibrinogen-binding protein. Microbiology 2010, 156, 2516. [Google Scholar] [CrossRef]
- Gophna, U.; Konikoff, T.; Nielsen, H.B. Oscillospira and related bacteria–From metagenomic species to metabolic features. Environ. Microbiol. 2017, 19, 835. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A central, enigmatic component of the human gut microbiota. Trends Microbiol. 2016, 24, 523. [Google Scholar] [CrossRef]
- Liu, Y.L.; Yan, T.; Li, X.Y.; Duan, Y.L.; Yang, X.; Yang, X.J. Effects of Bacillus subtilis and antibiotic growth promoters on the growth performance, intestinal function and gut microbiota of pullets from 0 to 6 weeks. Animal 2020, 14, 1619. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Huang, Z.; Zeng, S.; Liu, J.; Wei, D.; Deng, X.; He, J. Intestinal bacterial signatures of white feces syndrome in shrimp. Appl. Microbiol. Biotechnol. 2018, 102, 3701. [Google Scholar] [CrossRef] [PubMed]
- Jalanka, J.; Major, G.; Murray, K.; Singh, G.; Nowak, A.; Kurtz, C.; Spiller, R. The effect of psyllium husk on intestinal microbiota in constipated patients and healthy controls. Int. J. Mol. Sci. 2019, 20, 433. [Google Scholar] [CrossRef] [PubMed]
- Maier, E.; Anderson, R.C.; Roy, N.C. Live Faecalibacterium prausnitzii does not enhance epithelial barrier integrity in an apical anaerobic co-culture model of the large intestine. Nutrients 2017, 9, 1349. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Warth, B.; Spilker, M.E.; Siuzdak, G. Metabolomics activity screening for identifying metabolites that modulate phenotype. Nat. Biotechnol. 2018, 36, 316. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Ren, D.; Yan, W.; Wang, Y.; Liu, H.; Shen, M. Linoleic acid inhibits Lactobacillus activity by destroying cell membrane and affecting normal metabolism. J. Sci. Food Agric. 2020, 100, 2057. [Google Scholar] [CrossRef]
- Whelan, J.; Fritsche, K. Linoleic acid. Adv. Nutr. 2013, 4, 311. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.Q.; Ma, X.W.; Dong, X.Y.; Tao, Z.R.; Lu, L.Z.; Zou, X.T. Effects of parental dietary linoleic acid on growth performance, antioxidant capacity, and lipid metabolism in domestic pigeons (Columba livia). Poult. Sci. 2020, 99, 1471. [Google Scholar] [CrossRef]
- Qi, X.; Wu, S.; Zhang, H.; Yue, H.; Xu, S.; Ji, F.; Qi, G. Effects of dietary conjugated linoleic acids on lipid metabolism and antioxidant capacity in laying hens. Arch. Anim. Nutr. 2011, 65, 354. [Google Scholar] [CrossRef]
- Du, M.; Ahn, D.U. Dietary CLA affects lipid metabolism in broiler chicks. Lipids 2003, 38, 505. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, W.; Dong, R.; Wang, S.; Lin, Y.; Zheng, C. Effects of dietary linoleic acid level on laying performance, egg quality and lipids metabolism of ducks during the early laying period. Chin. J. Anim. Nutr. 2015, 27, 731. [Google Scholar] [CrossRef]
- Belury, M.A.; Cole, R.M.; Snoke, D.B.; Banh, T.; Angelotti, A. Linoleic acid, glycemic control and Type 2 diabetes. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 30. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhou, Y.; Wu, M.; Li, X.; Mai, K.; Ai, Q. ω-6 Polyunsaturated fatty acids (linoleic acid) activate both autophagy and antioxidation in a synergistic feedback loop via TOR-dependent and TOR-independent signaling pathways. Cell Death Dis. 2020, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489. [Google Scholar] [CrossRef] [PubMed]
- Emmett, M. Acetaminophen toxicity and 5-oxoproline (pyroglutamic acid): A tale of two cycles, one an ATP-depleting futile cycle and the other a useful cycle. Clin. J. Am. Soc. Nephrol. 2014, 9, 191. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hyde, A.S.; Simpson, M.A.; Barycki, J.J. Emerging regulatory paradigms in glutathione metabolism. Adv. Cancer Res. 2014, 122, 69. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Items | CON | ANT | LAG | HAG | SEM | p-Value |
| BW | ||||||
| Initial (g) | 46.05 | 46.30 | 46.20 | 45.97 | 0.15 | 0.893 |
| 21 day (g) | 841.72 b | 866.47 a | 885.78 a | 870.14 a | 5.05 | 0.010 |
| 42 day (g) | 2490.91 b | 2581.66 a | 2657.41 a | 2585.83 a | 17.81 | 0.003 |
| Day1–21 | ||||||
| ADFI (g/d) | 48.16 | 50.20 | 47.24 | 46.56 | 0.67 | 0.244 |
| ADG (g/d) | 37.89 b | 39.06 a | 39.99 a | 39.23 a | 0.24 | 0.009 |
| FCR (g·g) | 1.27 | 1.28 | 1.18 | 1.19 | 0.02 | 0.121 |
| Day22–42 | ||||||
| ADFI (g/d) | 142.78 | 138.50 | 148.05 | 142.55 | 1.26 | 0.054 |
| ADG (g/d) | 78.53 b | 81.68 ab | 84.36 a | 81.70 ab | 0.73 | 0.029 |
| FCR (g·g) | 1.82 | 1.70 | 1.76 | 1.75 | 0.01 | 0.296 |
| Day1–42 | ||||||
| ADFI (g/d) | 95.11 | 94.52 | 97.64 | 94.55 | 0.58 | 0.168 |
| ADG (g/d) | 58.21 b | 60.37 a | 62.18 a | 60.47 a | 0.42 | 0.003 |
| FCR (g·g) | 1.63 a | 1.57 b | 1.57 b | 1.56 b | 0.01 | <0.001 |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Items | CON | ANT | LAG | HAG | SEM | p-Value |
| Energy | 70.76 b | 74.77 a | 74.90 a | 75.49 a | 0.68 | 0.043 |
| CP | 47.47 b | 50.46 ab | 52.75 a | 49.98 ab | 0.63 | 0.019 |
| EE | 73.73 | 75.09 | 75.13 | 75.93 | 0.43 | 0.355 |
| Ca | 44.66 | 45.70 | 47.20 | 46.29 | 0.53 | 0.399 |
| P | 44.16 | 45.61 | 46.07 | 47.15 | 0.62 | 0.411 |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Items | CON | ANT | LAG | HAG | SEM | p-Value |
| pH45min | 6.24 b | 6.24 b | 6.50 a | 6.24 b | 0.04 | 0.038 |
| pH24h | 5.75 b | 5.82 ab | 5.91 a | 5.83 ab | 0.02 | 0.032 |
| L* | 46.35 c | 49.51 b | 52.61 a | 51.81 ab | 0.65 | <0.001 |
| a* | 6.79 | 6.29 | 6.93 | 5.69 | 0.28 | 0.423 |
| b* | 8.51 a | 8.65 a | 4.34 b | 6.32 b | 0.51 | 0.001 |
| Shear force | 27.14 a | 26.65 a | 24.34 b | 26.39 a | 0.39 | 0.047 |
| Drip loss | 1.62 a | 1.08 b | 0.96 b | 1.39 ab | 0.09 | 0.028 |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Items | CON | ANT | LAG | HAG | SEM | p-Value |
| SFA | ||||||
| C16:0 | 20.25 | 19.49 | 18.93 | 19.01 | 0.21 | 0.078 |
| C18:0 | 9.73 | 9.12 | 8.86 | 8.74 | 0.17 | 0.189 |
| C22:0 | 0.72 a | 0.61 b | 0.56 b | 0.57 b | 0.02 | 0.004 |
| MUFA | ||||||
| C16:1 | 1.27 c | 1.40 b | 1.38 b | 1.54 a | 0.02 | <0.001 |
| C18:1n-9c | 22.25 c | 23.51 ab | 23.02 b | 24.04 a | 0.17 | <0.001 |
| C22:1n-9 | 5.80 | 5.07 | 6.16 | 6.48 | 0.26 | 0.265 |
| C24:1 | 1.30 bc | 1.16 c | 1.55 ab | 1.68 a | 0.07 | 0.017 |
| PUFA | ||||||
| C18:2n-6 | 33.31 b | 35.38 a | 35.04 a | 35.53 a | 0.30 | 0.023 |
| C18:3n-3 | 2.05 c | 2.02 c | 2.66 a | 2.32 b | 0.06 | <0.001 |
| C20:2 | 0.86 b | 0.75 b | 1.12 a | 1.22 a | 0.05 | <0.001 |
| C20:3n-6 | 0.34 | 0.33 | 0.35 | 0.37 | 0.01 | 0.58 |
| C22:6n-3 | 0.60 c | 0.69 c | 1.21 a | 1.00 b | 0.06 | <0.001 |
| Total SFA | 30.70 a | 29.21 ab | 28.35 b | 28.32 b | 0.34 | 0.037 |
| Total MUFA | 30.62 c | 31.13 bc | 32.10 b | 33.73 a | 0.33 | 0.001 |
| Total PUFA | 37.15 b | 39.15 a | 40.32 a | 40.43 a | 0.36 | 0.001 |
| PUFA/SFA | 1.22 b | 1.35 a | 1.43 a | 1.43 a | 0.03 | 0.004 |
| n-6/n-3 | 57.83 a | 54.42 a | 29.82 b | 36.80 b | 2.99 | <0.001 |
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Items | CON | ANT | LAG | HAG | SEM | p-Value |
| Serum | ||||||
| T-AOC (U/mL) | 8.15 b | 8. 37 b | 9.08 a | 8.89 a | 0.10 | <0.001 |
| GSH-Px (U/mL) | 886.94 b | 889.72 b | 938.34 a | 906.35 b | 5.64 | 0.001 |
| SOD (U/mL) | 72.40 b | 75.85 b | 89.19 a | 79.47 b | 1.76 | 0.001 |
| MDA (mmol/mL) | 5.57 a | 5.62 a | 3.61 b | 3.65 b | 0.21 | <0.001 |
| Breast muscle | ||||||
| T-AOC (U/mg prot) | 127.98 b | 127.74 b | 135.54 a | 133.24 a | 1.13 | 0.002 |
| GSH-Px (U/mg prot) | 3.53 | 3.51 | 3.43 | 3.60 | 0.04 | 0.374 |
| SOD (U/mg prot) | 76.29 | 77.44 | 76.63 | 78.69 | 0.78 | 0.728 |
| MDA (nmol/mg prot) | 0.25 a | 0.24 a | 0.16 b | 0.16 b | 0.01 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, Y.; Guo, Y.; Zhang, W.; Guo, W.; Oleksandr, K.; Bozhko, N.; Wang, Z.; Liu, C. Effects of Compound Polysaccharides Derived from Astragalus and Glycyrrhiza on Growth Performance, Meat Quality and Antioxidant Function of Broilers Based on Serum Metabolomics and Cecal Microbiota. Antioxidants 2022, 11, 1872. https://doi.org/10.3390/antiox11101872
Qiao Y, Guo Y, Zhang W, Guo W, Oleksandr K, Bozhko N, Wang Z, Liu C. Effects of Compound Polysaccharides Derived from Astragalus and Glycyrrhiza on Growth Performance, Meat Quality and Antioxidant Function of Broilers Based on Serum Metabolomics and Cecal Microbiota. Antioxidants. 2022; 11(10):1872. https://doi.org/10.3390/antiox11101872
Chicago/Turabian StyleQiao, Yingying, Yongpeng Guo, Wei Zhang, Weibing Guo, Kyselov Oleksandr, Nataliia Bozhko, Zhixiang Wang, and Changzhong Liu. 2022. "Effects of Compound Polysaccharides Derived from Astragalus and Glycyrrhiza on Growth Performance, Meat Quality and Antioxidant Function of Broilers Based on Serum Metabolomics and Cecal Microbiota" Antioxidants 11, no. 10: 1872. https://doi.org/10.3390/antiox11101872
APA StyleQiao, Y., Guo, Y., Zhang, W., Guo, W., Oleksandr, K., Bozhko, N., Wang, Z., & Liu, C. (2022). Effects of Compound Polysaccharides Derived from Astragalus and Glycyrrhiza on Growth Performance, Meat Quality and Antioxidant Function of Broilers Based on Serum Metabolomics and Cecal Microbiota. Antioxidants, 11(10), 1872. https://doi.org/10.3390/antiox11101872