
Towards Functional Insect Feeds: Agri-Food By-Products Enriched with Post-Distillation Residues of Medicinal Aromatic Plants in Tenebrio molitor (Coleoptera: Tenebrionidae) Breeding

 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Colony
2.2. Diet Preparation
2.3. Experimental Design
2.4. Proximate Composition
2.5. GC-FID Analysis
2.6. Total Phenol and Flavonoid Content and Antioxidant Potential
2.7. Statistical Analysis
3. Results
3.1. Alternative Substrates’ Composition
3.2. Growth Performance
3.3. TML Proximate Composition
3.4. Effect of Substrates’ Proximate Composition on Growth and TML Nutrient Content
3.5. TML Fatty Acids Profile
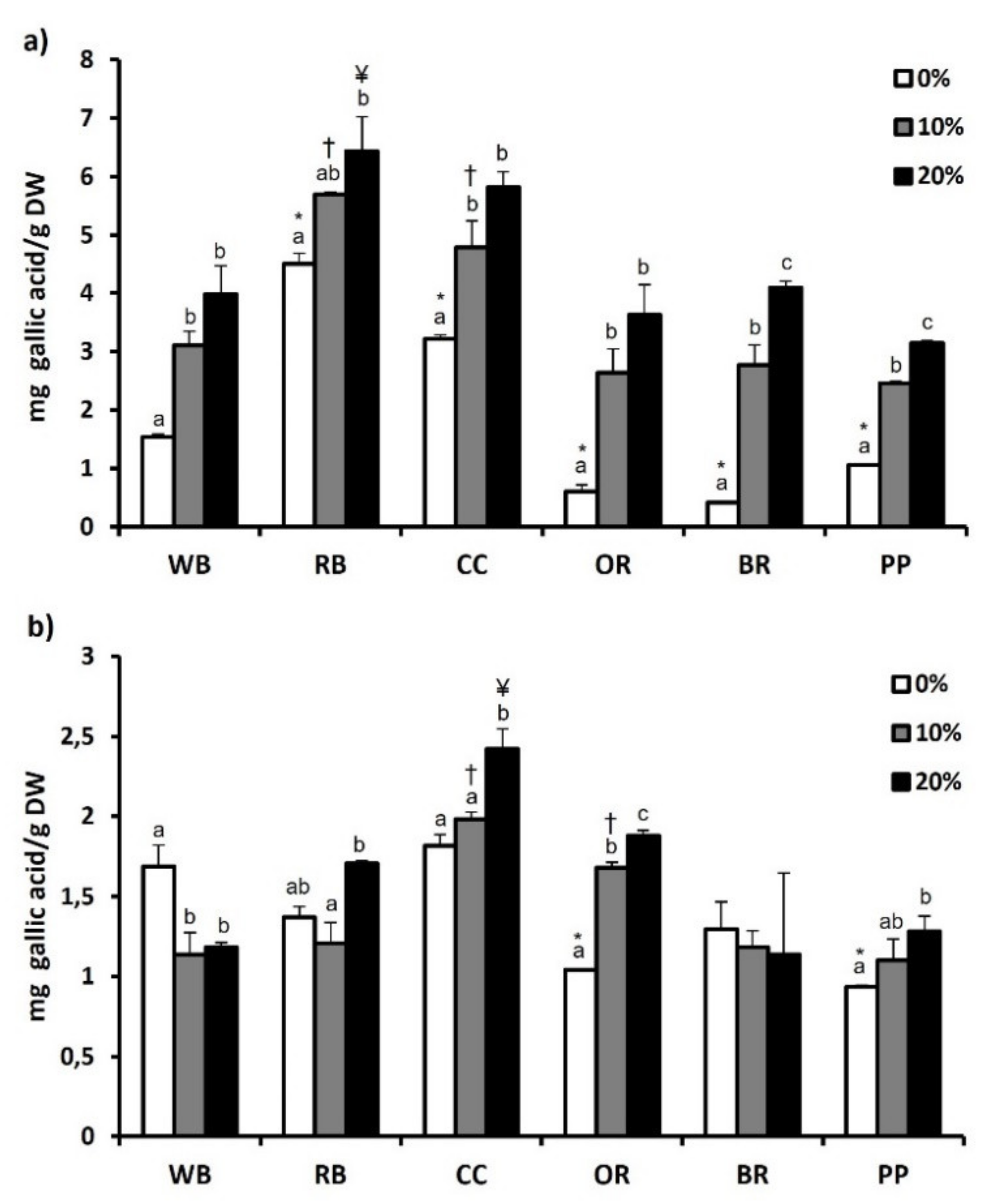
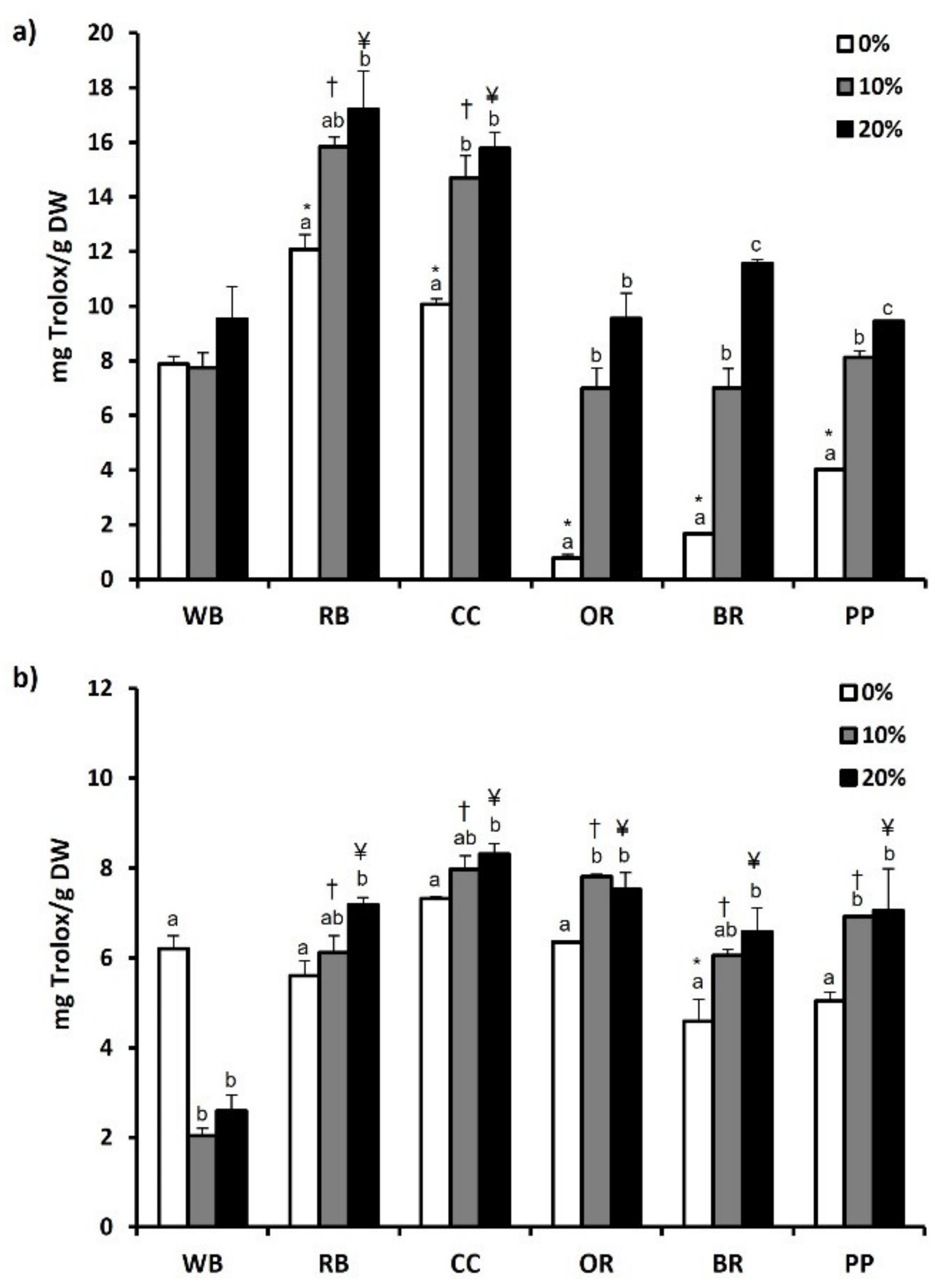
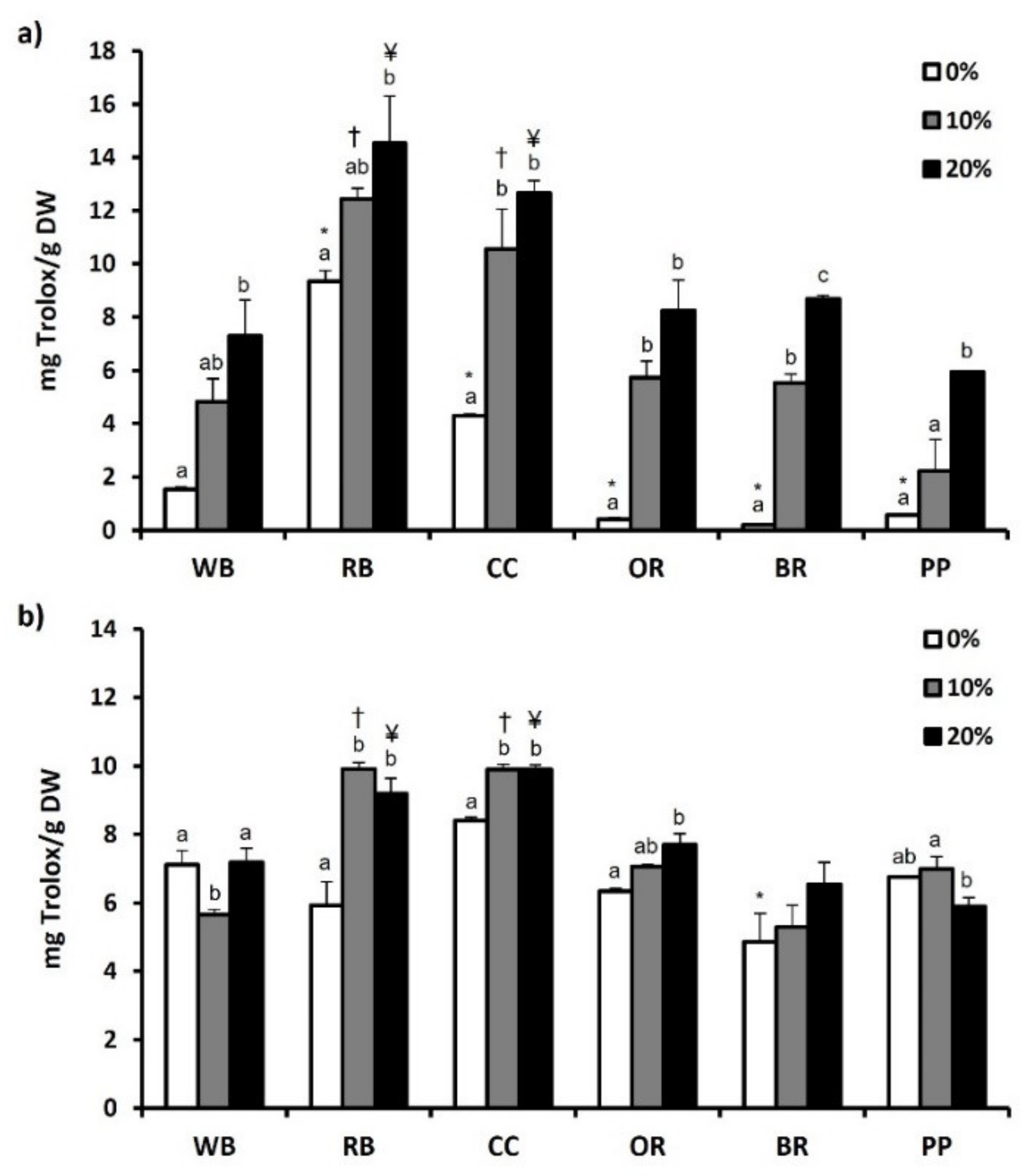
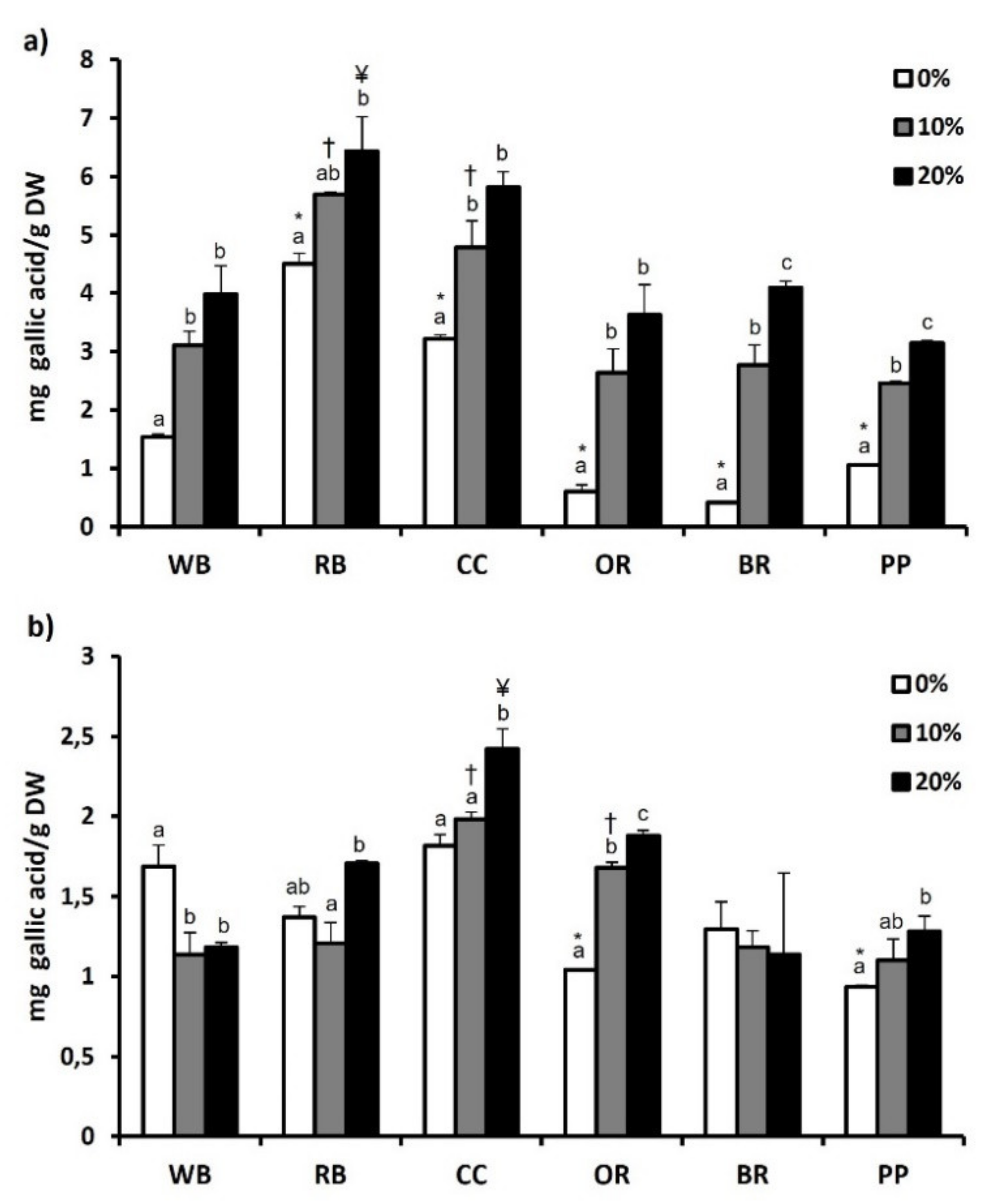
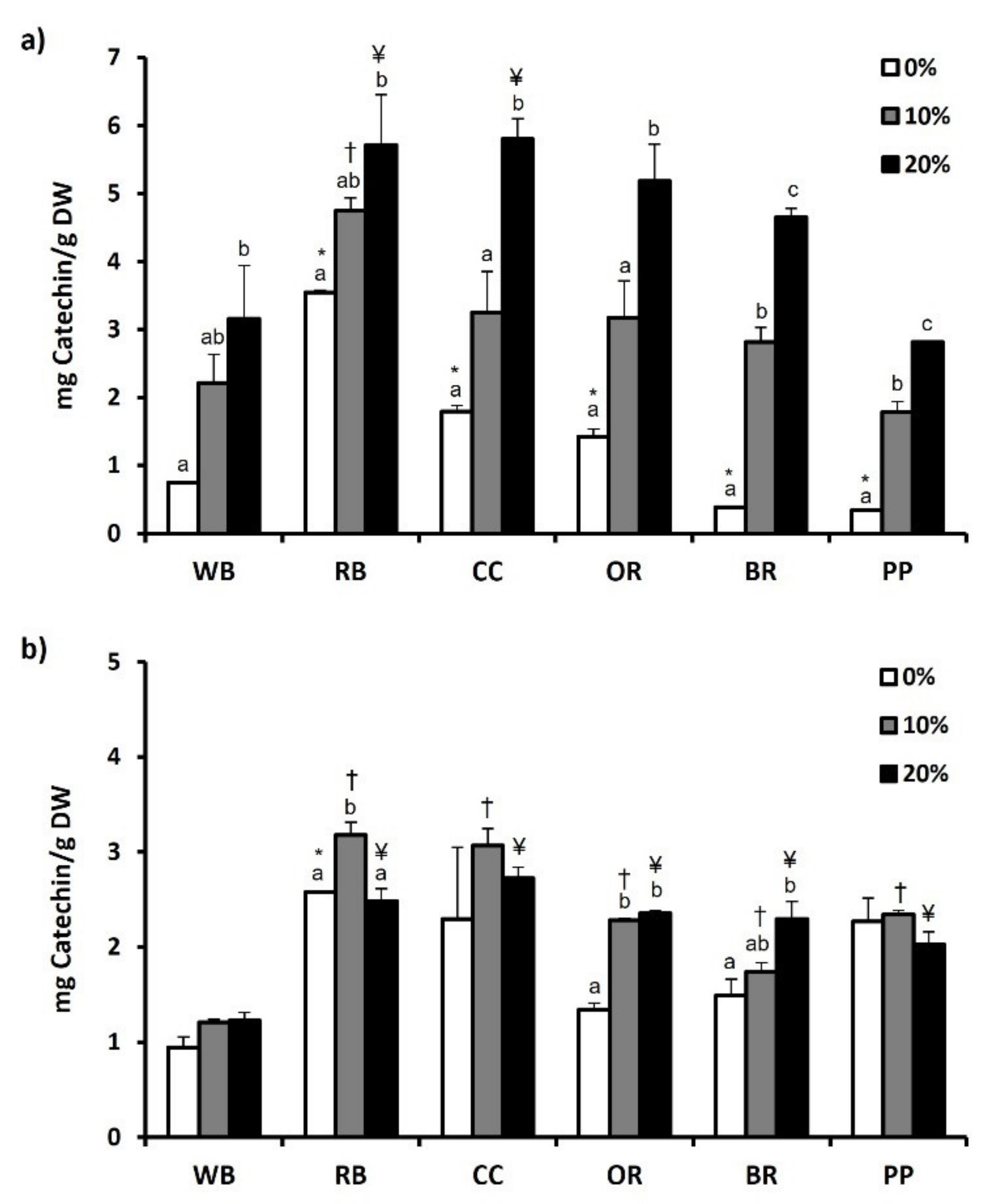
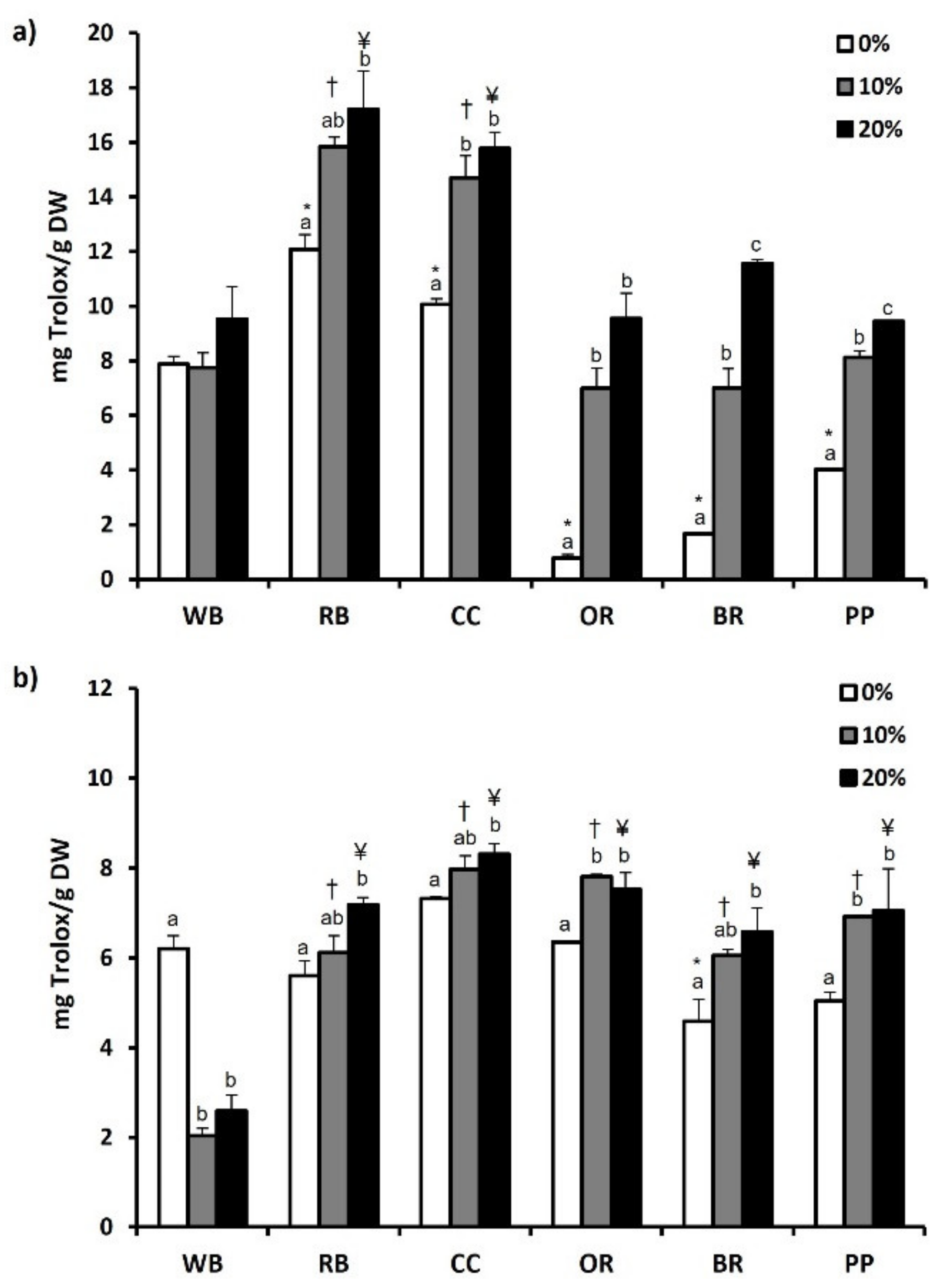
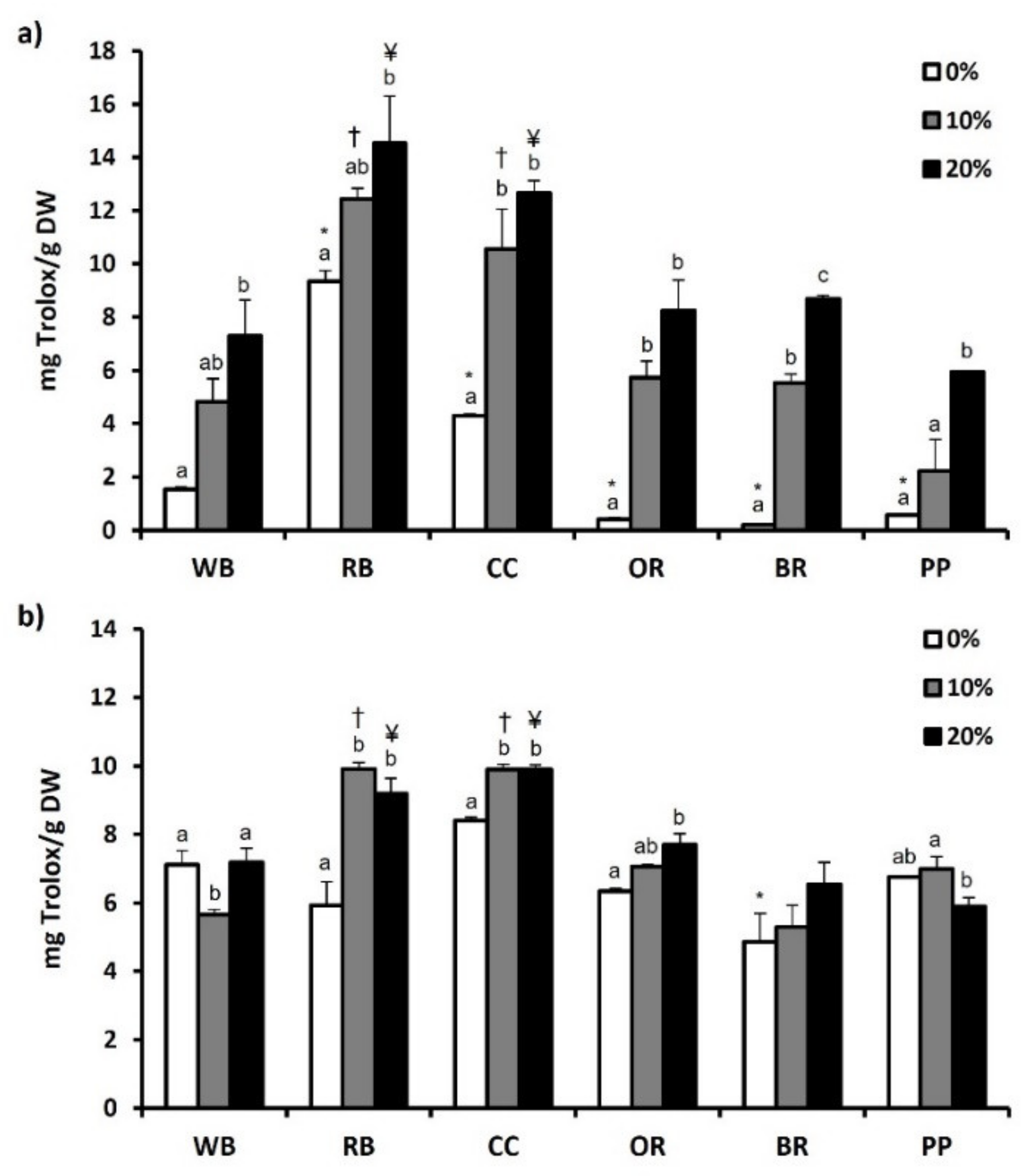
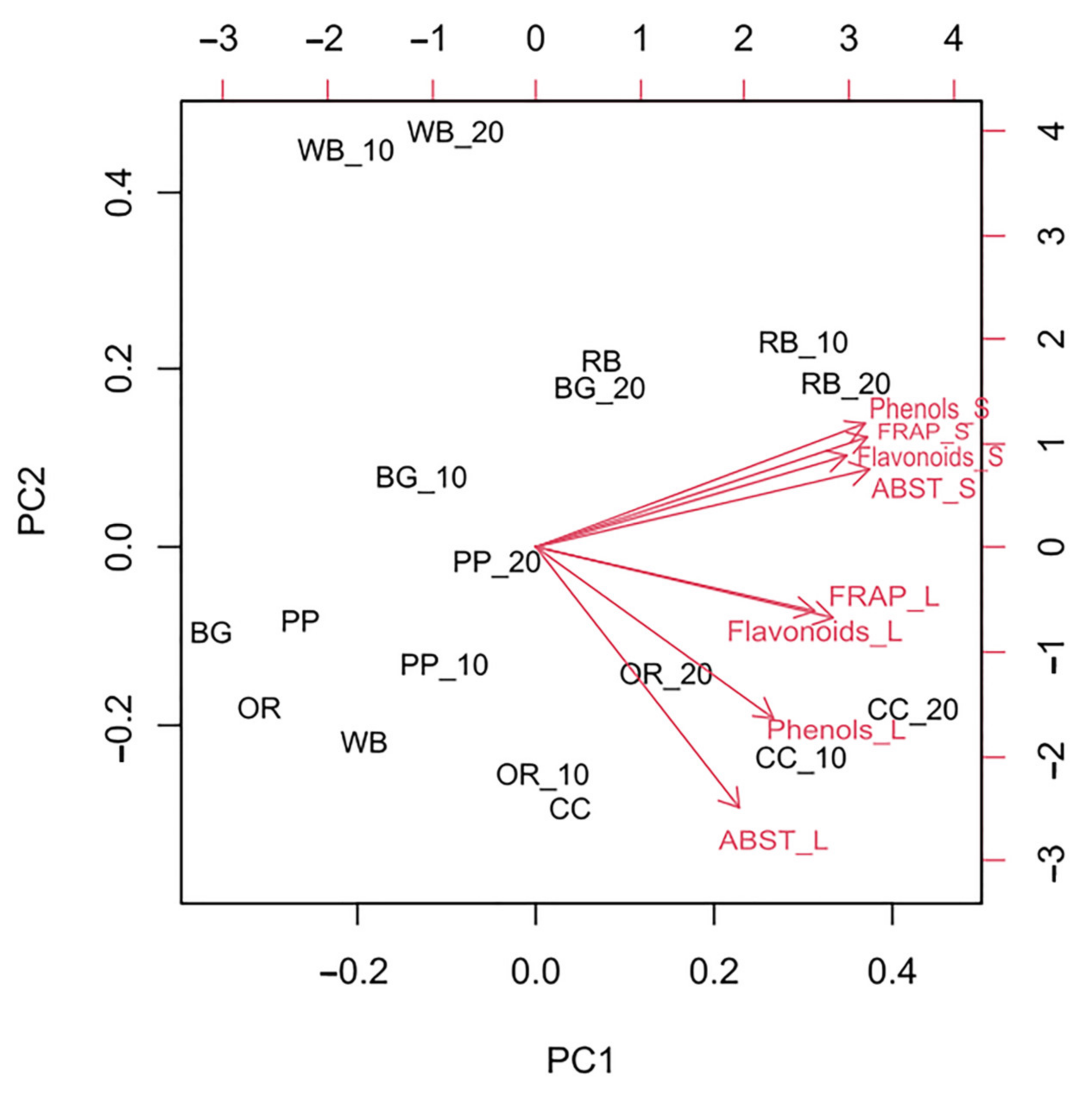
3.6. Total Phenol and Flavonoid Content and Antioxidant Potential
4. Discussion
4.1. Growth of TML
4.2. Composition of TML
4.3. Fatty Acids of TML
4.4. Phenols-Flavonoids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, Y.; Li, D. Impact of increased demand for animal protein products in Asian countries: Implications on global food security. Anim. Front. 2013, 3, 48–55. [Google Scholar] [CrossRef]
- Machovina, B.; Feeley, K.J.; Ripple, W.J. Biodiversity conservation: The key is reducing meat consumption. Sci. Total Environ. 2015, 536, 419–431. [Google Scholar] [CrossRef]
- Abbasi, T.; Abbasi, T.; Abbasi, S. Reducing the global environmental impact of livestock production: The minilivestock option. J. Clean. Prod. 2016, 112, 1754–1766. [Google Scholar] [CrossRef]
- Lenzen, M.; Moran, D.; Kanemoto, K.; Foran, B.; Lobefaro, L.; Geschke, A. International trade drives biodiversity threats in developing nations. Nature 2012, 486, 109–112. [Google Scholar] [CrossRef]
- Mateo-Sagasta, J.; Zadeh, S.M.; Turral, H.; Burke, J. Water Pollution from Agriculture: A Global Review. Executive Summary; FAO: Rome, Italy, 2017. [Google Scholar]
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock: A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; p. 115. [Google Scholar]
- UN. General Assembly United Nations Official Document—Transforming Our World: The 2030 Agenda for Sustainable Development. Available online: https://www.un.org/ga/search/view_doc.asp?symbol=A/RES/70/1&Lang=E (accessed on 20 October 2021).
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; p. 187. [Google Scholar]
- Yen, A.L. Edible insects: Traditional knowledge or western phobia? Entomol. Res. 2009, 39, 289–298. [Google Scholar] [CrossRef]
- Finke, M.D.; Oonincx, D.G.A.B. Insects as food for insectivores. In Mass Production of Beneficial Organisms; Morales-Ramos, J.A., Rojas, M.G., Shapiro-Ilan, D.I., Eds.; Elsevier: San Diego, CA, USA, 2014; pp. 583–616. [Google Scholar]
- Chia, S.Y.; Tanga, C.M.; van Loon, J.J.A.; Dicke, M. Insects for sustainable animal feed: Inclusive business models involving smallholder farmers. Curr. Opin. Environ. Sustain. 2019, 41, 23–30. [Google Scholar] [CrossRef]
- Dobermann, D.; Swift, J.A.; Field, L.M. Opportunities and hurdles of edible insects for food and feed. Nutr. Bull. 2017, 42, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jurgilevich, A.; Birge, T.; Kentala-Lehtonen, J.; Korhonen-Kurki, K.; Pietikäinen, J.; Saikku, L.; Schösler, H. Transition towards circular economy in the food system. Sustainability 2016, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- Fontana, A.R.; Antoniolli, A.; Bottini, R.n. Grape pomace as a sustainable source of bioactive compounds: Extraction, characterization, and biotechnological applications of phenolics. J. Agric. Food Chem. 2013, 61, 8987–9003. [Google Scholar] [CrossRef]
- Brewer, L.R.; Kubola, J.; Siriamornpun, S.; Herald, T.J.; Shi, Y.-C. Wheat bran particle size influence on phytochemical extractability and antioxidant properties. Food Chem. 2014, 152, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P. Role of food and nutrition in cancer. In The Role of Functional Food Security in Global Health; Singh, R.B., Watson, R.R., Takahashi, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 193–203. [Google Scholar]
- Schabel, H.G. Forest insects as food: A global review. In Proceedings of the Forest Insects as Food: Humans Bite Back/Asia-Pacific Resources and their Potential for Development, Chiang Mai, Thailand, 19–21 February 2008; pp. 37–40. [Google Scholar]
- Adámková, A.; Mlček, J.; Adámek, M.; Borkovcová, M.; Bednářová, M.; Hlobilová, V.; Knížková, I.; Juríková, T. Tenebrio molitor (Coleoptera: Tenebrionidae)—Optimization of rearing conditions to obtain desired nutritional values. J. Insect Sci. 2020, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- EU. 2017/893; Commission Regulation (EU) 2017/893 of 24 May 2017 Amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein; EU: Maastricht, The Netherlands, 2017. [Google Scholar]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A. Safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06343. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed. Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Silva, L.B.; de Souza, R.G.; da Silva, S.R.; Feitosa, A.d.C.; Lopes, E.C.; Lima, S.B.P.; Dourado, L.R.B.; Pavan, B.E. Development of Tenebrio molitor (Coleoptera: Tenebrionidae) on poultry litter-based diets: Effect on chemical composition of larvae. J. Insect Sci. 2021, 21, 7. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Bliamplias, D.; Gourgouta, M.; Michail, V.; Athanassiou, C.G. Rearing Tenebrio molitor and Alphitobius diaperinus larvae on seed cleaning process byproducts. Insects 2021, 12, 293. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, M.; Dall’Asta, C.; Galaverna, G.; del Castillo Bilbao, M.D. Rice Bran By-Product: From Valorization Strategies to Nutritional Perspectives. Foods 2021, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Sharif, M.K.; Butt, M.S.; Anjum, F.M.; Khan, S.H. Rice bran: A novel functional ingredient. Crit. Rev. Food Sci. Nutr. 2014, 54, 807–816. [Google Scholar] [CrossRef]
- Gebrechristos, H.Y.; Chen, W. Utilization of potato peel as eco-friendly products: A review. Food Sci. Nutr. 2018, 6, 1352–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, P.D.; Mandavgane, S.A.; Puranik, N.M.; Jambhulkar, S.J.; Kulkarni, B.D. Valorization of potato peel: A biorefinery approach. Crit. Rev. Biotechnol. 2018, 38, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Negro, M.J.; Manzanares, P.; Ruiz, E.; Castro, E.; Ballesteros, M. The biorefinery concept for the industrial valorization of residues from olive oil industry. In Olive Mill Waste; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 57–78. [Google Scholar]
- Castillo, S.; Rodríguez, A.; Bautista-Villarreal, M.; García-Solano, N.; Gallardo-Rivera, C.; Báez-González, J.G.; Sánchez-García, E.; García-Alanis, K.G. Rheological and Nutritional Characterization of Sweet Corn By-Product (Cob) to Develop a Functional Ingredient Applied in Dressings. Front. Nutr. 2021, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lau, T.; Harbourne, N.; Oruña-Concha, M.J. Valorisation of sweet corn (Zea mays) cob by extraction of valuable compounds. Int. J. Food Sci. Technol. 2019, 54, 1240–1246. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Fang, L.; Wang, J.; Zhang, C. Use of biogas solid residue from anaerobic digestion as an effective amendment to remediate Cr (VI)-contaminated soils. Environ. Sci. Pollut. Res. 2019, 26, 13041–13053. [Google Scholar] [CrossRef] [PubMed]
- Aravani, V.P.; Sun, H.; Yang, Z.; Liu, G.; Wang, W.; Anagnostopoulos, G.; Syriopoulos, G.; Charisiou, N.D.; Goula, M.A.; Kornaros, M. Agricultural and livestock sector’s residues in Greece & China: Comparative qualitative and quantitative characterization for assessing their potential for biogas production. Renew. Sustain. Energy Rev. 2022, 154, 111821. [Google Scholar] [CrossRef]
- De Elguea-Culebras, G.O.; Bravo, E.M.; Sánchez-Vioque, R. Potential sources and methodologies for the recovery of phenolic compounds from distillation residues of Mediterranean aromatic plants. An approach to the valuation of by-products of the essential oil market–A review. Ind. Crops Prod. 2022, 175, 114261. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant activity of medicinal and aromatic plants. A review. Flavour Fragr. J. 2010, 25, 291–312. [Google Scholar] [CrossRef]
- Mandalakis, M.; Anastasiou, T.I.; Martou, N.; Keisaris, S.; Greveniotis, V.; Katharios, P.; Lazari, D.; Krigas, N.; Antonopoulou, E. Antibacterial Effects of Essential Oils of Seven Medicinal-Aromatic Plants Against the Fish Pathogen Aeromonas veronii bv. sobria: To Blend or Not to Blend? Molecules 2021, 26, 2731. [Google Scholar] [CrossRef]
- Anastasiou, T.I.; Mandalakis, M.; Krigas, N.; Vézignol, T.; Lazari, D.; Katharios, P.; Dailianis, T.; Antonopoulou, E. Comparative evaluation of essential oils from medicinal-aromatic plants of Greece: Chemical composition, antioxidant capacity and antimicrobial activity against bacterial fish pathogens. Molecules 2020, 25, 148. [Google Scholar] [CrossRef] [Green Version]
- Mancini, S.; Fratini, F.; Turchi, B.; Mattioli, S.; Dal Bosco, A.; Tuccinardi, T.; Nozic, S.; Paci, G. Former foodstuff products in Tenebrio molitor rearing: Effects on growth, chemical composition, microbiological load, and antioxidant status. Animals 2019, 9, 484. [Google Scholar] [CrossRef] [Green Version]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of various commodities for the development of the yellow mealworm, Tenebrio molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasel, N.J.; Mène-Safrané, L.; Ruczynski, I.; Komar, E.; Christe, P. Diet induced modifications of fatty-acid composition in mealworm larvae (Tenebrio molitor ). J. Food Res. 2017, 6, 22–31. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists Inc.: Washington, DC, USA, 1990. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- FAO. Food Agriculture Organization of the United Nations. Amino Acid Content of Foods and Biological Data on Proteins; FAO: Rome, Italy, 1970; Volume 24. [Google Scholar]
- Maloney, K.P.; Truong, V.D.; Allen, J.C. Chemical optimization of protein extraction from sweet potato (Ipomoea batatas) peel. J. Food Sci. 2012, 77, E307–E312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Criscioni, P.; Fernández, C. Effect of rice bran as a replacement for oat grain in energy and nitrogen balance, methane emissions, and milk performance of Murciano-Granadina goats. J. Dairy Sci. 2016, 99, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, F.E.M.; Van Eylen, D.; Kabel, M.A. Characterization of substituents in xylans from corn cobs and stover. Carbohydr. Polym. 2011, 86, 722–731. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- AOCS. Official Methods and Recommended Practices of the American Oil Chemists’ Society, 4th ed.; American Oil Chemists’ Society: Urbana, IL, USA, 1989. [Google Scholar]
- Panteli, N.; Mastoraki, M.; Nikouli, E.; Lazarina, M.; Antonopoulou, E.; Kormas, K.A. Imprinting statistically sound conclusions for gut microbiota in comparative animal studies: A case study with diet and teleost fishes. Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 36, 100738. [Google Scholar] [CrossRef]
- Scalbert, A.; Monties, B.; Janin, G. Tannins in wood: Comparison of different estimation methods. J. Agric. Food Chem. 1989, 37, 1324–1329. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free. Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Jensen, K.; Kristensen, T.N.; Heckmann, L.H.; Sørensen, J.G. Breeding and Maintaining High-Quality Insects; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017; pp. 175–198. [Google Scholar]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-llan, D.I.; Tedders, W.L. Use of nutrient self-selection as a diet refining tool in Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2013, 48, 206–221. [Google Scholar] [CrossRef]
- Urrejola, S.; Nespolo, R.; Lardies, M.A. Diet-induced developmental plasticity in life histories and energy metabolism in a beetle. Rev. Chil. Hist. Nat. 2011, 84, 523–533. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Self-selection of two diet components by Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and its impact on fitness. Environ. Entomol. 2011, 40, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Rho, M.S.; Lee, K.P. Geometric analysis of nutrient balancing in the mealworm beetle, Tenebrio molitor L. (Coleoptera: Tenebrionidae). J. Insect Physiol. 2014, 71, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Sideris, V.; Georgiadou, M.; Papadoulis, G.; Mountzouris, K.; Tsagkarakis, A. Effect of processed beverage by-product-based diets on biological parameters, conversion efficiency and body composition of Hermetia illucens (L) (Diptera: Stratiomyidae). Insects 2021, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Minor, M.; Morel, P.C.; Najar-Rodriguez, A.J. Bioconversion of three organic wastes by black soldier fly (Diptera: Stratiomyidae) larvae. Environ. Entomol. 2018, 47, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Przemieniecki, S.W.; Kosewska, A.; Ciesielski, S.; Kosewska, O. Changes in the gut microbiome and enzymatic profile of Tenebrio molitor larvae biodegrading cellulose, polyethylene and polystyrene waste. Environ. Pollut. 2020, 256, 113265. [Google Scholar] [CrossRef] [PubMed]
- Van Broekhoven, S.; Oonincx, D.G.A.B.; Van Huis, A.; Van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Arapoglou, D.; Varzakas, T.; Vlyssides, A.; Israilides, C. Ethanol production from potato peel waste (PPW). Waste Manag. 2010, 30, 1898–1902. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G. Rice Bran and Other Rice By-Products. Available online: https://www.feedipedia.org/node/750 (accessed on 7 October 2021).
- Heuzé, V.; Tran, G.; Baumont, R.; Noblet, J.; Renaudeau, D.; Lessire, M.; Lebas, F. Wheat Bran. Available online: https://www.feedipedia.org/node/726 (accessed on 7 October 2021).
- Heuzé, V.; Tran, G.; Lebas, F. Maize Cobs. Available online: https://www.feedipedia.org/node/718 (accessed on 7 October 2021).
- Pavela, R. The feeding effect of polyphenolic compounds on the Colorado potato beetle (Leptinotarsa decemlineata Say). Pest Technol. 2007, 1, 81–84. [Google Scholar]
- Regnault-Roger, C.; Ribodeau, M.; Hamraoui, A.; Bareau, I.; Blanchard, P.; Gil-Munoz, M.-I.; Barberan, F.T. Polyphenolic compounds of Mediterranean Lamiaceae and investigation of orientational effects on Acanthoscelides obtectus (Say). J. Stored Prod. Res. 2004, 40, 395–408. [Google Scholar] [CrossRef]
- Fuso, A.; Barbi, S.; Macavei, L.I.; Luparelli, A.V.; Maistrello, L.; Montorsi, M.; Sforza, S.; Caligiani, A. Effect of the rearing substrate on total protein and amino acid composition in black soldier fly. Foods 2021, 10, 1773. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downer, R.G.H.; Matthews, J.R. Patterns of lipid distribution and utilisation in insects. Am. Zool. 1976, 16, 733–745. [Google Scholar] [CrossRef]
- Costa, S.; Pedro, S.; Lourenço, H.; Batista, I.; Teixeira, B.; Bandarra, N.M.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio molitor larvae as an alternative food source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Siemianowska, E.; Kosewska, A.; Aljewicz, M.; Skibniewska, K.A.; Polak-Juszczak, L.; Jarocki, A.; Jedras, M. Larvae of mealworm (Tenebrio molitor L.) as European novel food. Agric. Sci. 2013, 4, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.A.; Ding, Q.; Yin, L.; Chi, X.; Sun, N.; He, R.; Luo, L.; Ma, H.; Li, Z. Comparison of the nutritional value of mysore thorn borer (Anoplophora chinensis) and mealworm larva (Tenebrio molitor ): Amino acid, fatty acid, and element profiles. Food Chem. 2020, 323, 126818. [Google Scholar] [CrossRef] [PubMed]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.-J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422, 193–201. [Google Scholar] [CrossRef]
- Paul, A.; Frederich, M.; Megido, R.C.; Alabi, T.; Malik, P.; Uyttenbroeck, R.; Francis, F.; Blecker, C.; Haubruge, E.; Lognay, G. Insect fatty acids: A comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. J. Asia-Pac. Entomol. 2017, 20, 337–340. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquacult. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Cappellozza, S.; Leonardi, M.G.; Savoldelli, S.; Carminati, D.; Rizzolo, A.; Cortellino, G.; Terova, G.; Moretto, E.; Badaile, A.; Concheri, G. A first attempt to produce proteins from insects by means of a circular economy. Animals 2019, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.S.V.C.; Reddy, S.G.; Babu, P.P.; Reddy, A.R. The antioxidant and antiproliferative activities of methanolic extracts from Njavara rice bran. BMC Complement. Altern. Med. 2010, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Devi, R.R.; Arumughan, C. Phytochemical characterization of defatted rice bran and optimization of a process for their extraction and enrichment. Bioresour. Technol. 2007, 98, 3037–3043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.W.; Zhang, R.F.; Zhang, F.X.; Liu, R.H. Phenolic profiles and antioxidant activity of black rice bran of different commercially available varieties. J. Agric. Food Chem. 2010, 58, 7580–7587. [Google Scholar] [CrossRef]
- Burghardt, F.; Knüttel, H.; Becker, M.; Fiedler, K. Flavonoid wing pigments increase attractiveness of female common blue (Polyommatus icarus) butterflies to mate-searching males. Naturwissenschaften 2000, 87, 304–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmonds, M.S.J. Flavonoid–insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Yang, T.; Guang Hu, J.; Yu, Y.; Li, G.; Guo, X.; Li, T.; Liu, R.H. Comparison of phenolics, flavonoids, and cellular antioxidant activities in ear sections of sweet corn (Zea mays L. saccharata Sturt). J. Food Processing Preserv. 2019, 43, e13855. [Google Scholar] [CrossRef] [Green Version]
- Balandrán-Quintana, R.R.; Mercado-Ruiz, J.N.; Mendoza-Wilson, A.M. Wheat bran proteins: A review of their uses and potential. Food Rev. Int. 2015, 31, 279–293. [Google Scholar] [CrossRef]
- Friedman, M.; Kozukue, N.; Kim, H.-J.; Choi, S.-H.; Mizuno, M. Glycoalkaloid, phenolic, and flavonoid content and antioxidative activities of conventional nonorganic and organic potato peel powders from commercial gold, red, and Russet potatoes. J. Food Compos. Anal. 2017, 62, 69–75. [Google Scholar] [CrossRef]
- Singh, P.P.; Saldaña, M.D.A. Subcritical water extraction of phenolic compounds from potato peel. Food Res. Int. 2011, 44, 2452–2458. [Google Scholar] [CrossRef]
- Mohdaly, A.A.A.; Sarhan, M.A.; Smetanska, I.; Mahmoud, A. Antioxidant properties of various solvent extracts of potato peel, sugar beet pulp and sesame cake. J. Sci. Food Agric. 2010, 90, 218–226. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Rice By-Products: Phytochemicals and Food Products Application, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2020; p. 129. [Google Scholar]
- Friedman, M.; Jürgens, H.S. Effect of pH on the stability of plant phenolic compounds. J. Agric. Food Chem. 2000, 48, 2101–2110. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.; Baser, K.H.C.; Windisch, W. Essential oils and aromatic plants in animal feeding—A European perspective. A review. Flavour Fragr. J. 2010, 25, 327–340. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Wasowicz, E.; Gramza, A.; Hes, M.; Jelen, H.; Korczak, J.; Malecka, M.; Mildner-Szkudlarz, S.; Rudzinska, M.; Samotyja, U.; Zawirska-Wojtasiak, R. Oxidation of lipids in food. Pol. J. Food Nutr. Sci. 2004, 13, 87–100. [Google Scholar]
- Nino, M.C.; Reddivari, L.; Osorio, C.; Kaplan, I.; Liceaga, A.M. Insects as a source of phenolic compounds and potential health benefits. J. Insects Food Feed. 2021, 7, 1077–1087. [Google Scholar] [CrossRef]
- Ruuhola, T.; Tikkanen, O.-P.; Tahvanainen, J. Differences in host use efficiency of larvae of a generalist moth, Operophtera brumata on three chemically divergent Salix species. J. Chem. Ecol. 2001, 27, 1595–1615. [Google Scholar] [CrossRef]
- Movva, V.; Pathipati, U.R. Feeding-induced phenol production in Capsicum annuum L. influences Spodoptera litura F. larval growth and physiology. Arch. Insect Biochem. Physiol. 2017, 95, e21387. [Google Scholar] [CrossRef]
- Di Mattia, C.; Battista, N.; Sacchetti, G.; Serafini, M. Antioxidant activities in vitro of water and liposoluble extracts obtained by different species of edible insects and invertebrates. Front. Nutr. 2019, 6, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Dry Matter | Protein | Fat | Ash | Energy (MJ/kg) | Fiber and NFE | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Addition of MAP (%) | Addition of MAP (%) | Addition of MAP (%) | Addition of MAP (%) | Addition of MAP (%) | Addition of MAP (%) | |||||||||||||
| 0 | 10 | 20 | 0 | 10 | 20 | 0 | 10 | 20 | 0 | 10 | 20 | 0 | 10 | 20 | 0 | 10 | 20 | |
| WB | 86.0 | 88.7 | 88.4 | 21.5 | 19.8 | 19.7 | 5.4 | 5.2 | 6.1 | 5.6 | 5.0 | 5.1 | 19.1 | 19.4 | 19.7 | 67.5 | 70.0 | 69.0 |
| RB | 89.6 | 93.5 | 91.4 | 19.8 | 18.5 | 18.7 | 19.1 | 20.9 | 20.4 | 10.3 | 9.5 | 9.1 | 21.8 | 22.6 | 22.2 | 50.8 | 51.2 | 51.9 |
| CC | 88.1 | 91.0 | 90.5 | 7.3 | 5.9 | 6.2 | 2.2 | 1.7 | 2.3 | 1.5 | 1.9 | 2.8 | 18.3 | 18.6 | 18.9 | 88.9 | 90.5 | 88.8 |
| OR | 78.1 | 83.1 | 81.4 | 12.5 | 10.9 | 12.4 | 1.2 | 1.1 | 2.4 | 70.7 | 69.2 | 62.7 | 5.8 | 6.8 | 8.6 | 15.6 | 18.8 | 22.5 |
| BR | 90.4 | 89.1 | 89.7 | 25.3 | 24.3 | 22.0 | 2.0 | 2.4 | 2.5 | 27.3 | 24.8 | 24.9 | 14.4 | 15.6 | 15.4 | 45.4 | 48.4 | 50.5 |
| PP | 87.3 | 86.1 | 86.8 | 13.8 | 13.1 | 13.0 | 1.1 | 1.8 | 3.1 | 6.0 | 6.6 | 6.9 | 16.6 | 17.3 | 18.1 | 79.1 | 78.5 | 77.0 |
| Substrate | Number of Larvae | Total Dry Weight | ||||
|---|---|---|---|---|---|---|
| Addition of MAP (%) | Addition of MAP (%) | |||||
| 0 | 10 | 20 | 0 | 10 | 20 | |
| WB | 37.0 ab ± 9.7b | 82.0 a ± 14.6a | 72.6 a ± 5.2ab | 4.63 a ± 0.87a | 6.49 a ± 0.67a | 5.74 a ± 0.36a |
| RB | 26.0 ab ± 5.2b | 58.2 ab ± 9.4a | 45.2 ab ± 13.2ab | 0.68 b ± 0.11b | 2.51b ± 0.73a | 2.02 b ± 0.43a |
| CC | 48.6 a ± 9.7a | 43.0 ab ± 13.1a | 39.2 ab ± 9.8a | 1.50 b ± 0.15a | 1.73 bc ± 0.52a | 1.40 bc ± 0.25a |
| OR | 11.8 b ± 2.5b | 35.0 b ± 6.6a | 26.0 b ± 5.2ab | 0.05 c ± 0.01b | 0.32 c ± 0.02a | 0.38 cd ± 0.09a |
| BR | 12.0 b ± 0.5b | 13.6 c ± 2.2ab | 22.8 b ± 3.7a | 0.05 c ± 0.01b | 0.27 c ± 0.03a | 0.29 d ± 0.02a |
| PP | 25.0 ab ± 3.1a | 44.8 ab ± 13.6a | 28.0 b ± 4.4a | 0.15 c ± 0.02b | 0.62 bc ± 0.11a | 0.43 cd ± 0.12ab |
| % MAPs | WB | RB | CC | OR | BR | PP | |
| Dry matter | 0 | 39.3 ± 1.6 b | 39.5 ± 1.1 b | 31.6 ± 0.2 c | 32.9 ± 0.6 c, A | 33.3 ± 0.8 c | 45.1 ± 1.3 a, A |
| 10 | 40.3 ± 0.6 ab | 42.8 ± 0.2 a | 30.6 ± 0.5 c | 28.5 ± 2.1 c,B | 31.9 ± 2.0 c | 38.1 ± 0.3 b, B | |
| 20 | 39.5 ± 1.1 ab | 41.8 ± 0.2 a | 31.6 ± 0.3 c | 35.3 ± 1.8 b, A | 30.1 ± 0.4 c | 37.4 ± 0.0 ab, B | |
| Protein (N x 6.25) | 0 | 51.2 ± 0.9 | 53.4 ± 0.2 | 50.7 ± 0.6 | 57.5 ± 0.9 A | 55.8 ± 2.0 B | 59.5 ± 2.0 A |
| 10 | 52.1 ± 1.4 ab | 47.7 ± 0.5 bc | 51.3 ± 1.8 ac | 56.0 ± 4.2 ab, A | 61.7 ± 3.8 a, A | 44.1 ± 2.0 c, B | |
| 20 | 50.6 ± 0.1 | 47.5 ± 1.6 | 44.4 ± 4.1 | 42.8 ± 0.4 B | 52.9 ± 6.5 B | 43.6 ± 1.2 B | |
| Protein (N x 4.76) | 0 | 39.0 ± 0.7 | 40.7 ± 0.1 | 38.6 ± 0.4 | 43.8 ± 0.7 A | 42.5 ± 1.5 B | 45.3 ± 1.5 A |
| 10 | 39.7 ± 1.1 ab | 36.4 ± 0.4 bc | 39.1 ± 1.3 ac | 42.7 ± 3.2 ab, A | 47.0 ± 2.9 a, A | 33.6 ± 1.5 c, B | |
| 20 | 38.5 ± 0.1 | 36.2 ± 1.2 | 33.8 ± 3.1 | 32.6 ± 0.3 B | 40.3 ± 4.9 B | 33.2 ± 0.9 B | |
| Fat | 0 | 25.2 ± 2.5 b | 31.3 ± 0.3 a | 16.6 ± 1.1 c | 23.9 ± 1.6 b, A | 20.9 ± 0.5 bc | 21.7 ± 2.2 bc, B |
| 10 | 26.6 ± 0.6 ab | 30.7 ± 0.3 ab | 15.5 ± 0.7 c | 10.4 ± 0.4 c, B | 22.2 ± 0.7 b | 29.6 ± 1.4 a, A | |
| 20 | 28.2 ± 3.1 a | 31.3 ± 0.2 a | 16.0 ± 0.1 bc | 11.5 ± 0.6 c, B | 17.8 ± 1.6 bc | 29.7 ± 0.9 a, A | |
| Ash | 0 | 5.5 ± 0.2 b | 5.0 ± 0.4 b | 5.3 ± 0.1 b, A | 6.7 ± 0.2 b, C | 10.2 ± 0.1 a, A | 10.3 ± 0.5 a, A |
| 10 | 4.8 ± 0.2 c | 5.3 ± 0.3 d | 2.5 ± 0.1 e, B | 9.8 ± 0.4 a, B | 7.7 ± 0.2 b, B | 5.4 ± 0.9 c, B | |
| 20 | 5.2 ± 0.4 d | 6.1 ± 0.4 bc | 5.2 ± 0.3 c, A | 13.1 ± 0.7 a, A | 7.8 ± 0.5 b, B | 5.3 ± 0.4 c, B | |
| Fiber & NFE | 0 | 18.1 ± 3.2 ab | 10.3 ± 0.8 b | 27.5 ± 1.6 a | 11.9 ± 1.0 b, B | 13.2 ± 2.2 b, AB | 8.5 ± 3.8 b, B |
| 10 | 16.4 ± 2.1 bc | 16.2 ± 0.8 bc | 30.7 ± 2.5 a | 23.7 ± 3.5 ab, A | 8.4 ± 4.1 c, B | 20.9 ± 1.6 bc, A | |
| 20 | 16.0 ± 2.7 c | 15.1 ± 1.0 c | 34.4 ± 3.8 a | 32.6 ± 1.7 ab, A | 21.5 ± 6.7 bc, A | 21.4 ± 0.5 c, A | |
| Two-way ANOVA p-values | |||||||
| Substate type | MAP addition | Substrate x MAP | |||||
| Dry matter | <0.001 | 0.046 | <0.001 | ||||
| Protein | 0.004 | <0.001 | 0.005 | ||||
| Fat | <0.001 | Non-significant | <0.001 | ||||
| Ash | <0.001 | <0.001 | <0.001 | ||||
| Fiber & NFE | <0.001 | <0.001 | <0.010 | ||||
| no. of TML | Total Dry Larval Weight | TML Dry Matter | TML Protein | TML Fat | TML Ash | TML Fiber & NFE | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substrates’ | r | Sig. | r | Sig. | r | Sig. | r | Sig. | r | Sig. | r | Sig. | r | Sig. |
| Dry matter | 0.270 | NS | 0.304 | NS | 0.112 | NS | −0.049 | NS | 0.228 | NS | −0.397 | NS | −0.067 | NS |
| Protein | −0.252 | NS | −0.069 | NS | 0.333 | NS | 0.368 | NS | 0.439 | NS | 0.222 | NS | −0.657 | ** |
| Fat | 0.427 | NS | 0.636 | ** | 0.496 | * | −0.377 | NS | 0.684 | ** | −0.368 | NS | −0.162 | NS |
| Ash | −0.682 | ** | −0.671 | ** | −0.164 | NS | 0.315 | NS | −0.051 | NS | 0.699 | *** | −0.302 | NS |
| Energy | 0.727 | *** | 0.843 | *** | 0.591 | ** | −0.362 | NS | 0.643 | ** | −0.703 | *** | −0.090 | NS |
| Fiber and NFE | 0.561 | NS | 0.483 | * | 0.139 | NS | −0.354 | NS | 0.046 | NS | −0.602 | ** | 0.323 | NS |
| Fatty Acid | % MAPs | WB | CC | RB | OR | BG | PP | SEMpooled |
|---|---|---|---|---|---|---|---|---|
| C10:0 | 0 | 0.26 f, B | 0.66 d, B | 0.36 e, A | 0.85 c, B | 1.31 b, A | 2.62 a, A | 0.01 |
| 10 | 0.38 e, A | 0.64 b, C | 0.21 f, B | 0.61 c, C | 0.87 a, C | 0.44 d, C | ||
| 20 | 0.23 f, C | 0.71 c, A | 0.35 e, A | 1.14 a, A | 0.89 b, B | 0.46 d, B | ||
| C14:0 | 0 | 3.62 e, B | 4.72 b, A | 2.78 f, C | 4.10 d, B | 5.09 a, A | 4.13 c, A | 0.01 |
| 10 | 4.11 b, A | 4.63 a, B | 3.73 f, A | 4.02 d, C | 4.11 c, B | 3.99 e, B | ||
| 20 | 3.14 e, C | 4.12 b, C | 2.89 f, B | 4.99 a, A | 3.57 d, C | 3.81 c, C | ||
| C16:0 | 0 | 11.0 f, B | 14.4 c, A | 12.1 d, B | 16.9 a, A | 16.2 b, A | 16.3 b, A | 0.03 |
| 10 | 11.7 e, A | 13.1 cd, B | 13.0 d, A | 13.7 a, B | 13.4 bc, B | 13.5 ab, B | ||
| 20 | 6.76 f, C | 13.0 a, B | 10.1 d, C | 11.9 c, C | 12.2 b, C | 8.85 a, C | ||
| C16:1 | 0 | 2.18 e, C | 2.84 d, A | 1.36 f, C | 2.94 c, A | 3.10 b, A | 3.57 a, A | 0.11 |
| 10 | 2.95 b, A | 2.50 de, C | 2.77 c, A | 2.51 d, B | 1.92 e, C | 3.47 a, C | ||
| 20 | 2.66 b, B | 2.56 c, B | 1.81 f, B | 2.15 e, C | 2.21 d, B | 3.54 a, B | ||
| C17:0 | 0 | 1.30 e, B | 3.13 d, C | 1.20 f, C | 6.37 c, A | 8.09 a, A | 6.81 b, A | 0.02 |
| 10 | 1.91 d, A | 4.47 b, A | 1.29 e, A | 1.94 d, C | 5.36 a, C | 2.14 c, B | ||
| 20 | 1.25 e, C | 3.32 c, B | 1.24 e, B | 5.76 b, B | 6.35 a, B | 1.36 d, C | ||
| C18:0 | 0 | 2.48 e, A | 3.75 c, A | 3.62 c, A | 3.35 d, C | 5.05 b, A | 6.68 a, A | 0.08 |
| 10 | 2.17 e, B | 3.38 c, B | 2.44 d, C | 3.58 b, A | 3.88 a, B | 2.20 e, B | ||
| 20 | 1.16 d, C | 3.20 a, C | 2.70 b, B | 3.35 a, B | 3.36 a, C | 1.51 c, C | ||
| C18:1 | 0 | 47.4 a, B | 46.0 b, A | 44.8 c, C | 46.9 a, A | 41.8 d, B | 38.4 e, C | 0.28 |
| 10 | 45.8 d, C | 42.3 e, C | 48.7 b, B | 46.6 c, A | 39.3 f, C | 52.5 a, B | ||
| 20 | 49.6 b, A | 45.0 c, B | 49.4 b, A | 42.1 e, B | 43.0 d, A | 56.0 a, A | ||
| C18:2 ω6 | 0 | 27.6 b, B | 19.1 c, C | 28.8 a, A | 15.6 d, C | 15.0 e, C | 14.0 f, C | 0.05 |
| 10 | 26.1 a, C | 18.8 d, A | 25.3 b, C | 20.7 c, A | 18.5 e, A | 15.5 f, B | ||
| 20 | 30.0 a, A | 20.1 c, B | 28.4 b, B | 18.0 d, B | 18.0 d, B | 17.6 e, A | ||
| C20:1 | 0 | 0.57 a, C | 0.20 e, C | 0.33 d, C | 0.38 bc, C | 0.41 b, C | 0.34 cd, C | 0.02 |
| 10 | 0.75 e, B | 2.18 b, B | 0.62 f, A | 1.03 d, B | 8.30 a, A | 1.65 c, B | ||
| 20 | 0.94 d, A | 2.36 b, A | 0.57 e, B | 2.09 c, A | 4.85 a, B | 2.34 b, A | ||
| SFAs | 0 | 18.9 e, B | 27.0 c, A | 20.2 d, B | 31.8 b, A | 36.6 a, A | 37.0 a, A | 0.20 |
| 10 | 20.6 e, A | 26.9 b, A | 20.9 e, A | 24.3 c, C | 27.8 a, B | 22.7 d, B | ||
| 20 | 12.9 f, C | 24.9 c, B | 17.5 d, C | 27.6 a, B | 26.8 b, C | 16.5 e, C | ||
| MUFAs | 0 | 50.2 a, B | 49.4 b, B | 46.5 c, C | 50.7 a, A | 46.6 c, C | 44.0 d, B | 0.25 |
| 10 | 49.9 cd, B | 47.3 e, C | 52.4 b, A | 50.5 c, A | 49.6 d, B | 58.1 a, A | ||
| 20 | 53.6 b, A | 50.3 d, A | 52.0 c, B | 46.3 e, B | 50.4 d, A | 62.3 a, A | ||
| PUFAs | 0 | 27.8 b, B | 19.4 c, B | 29.1 a, A | 15.8 e, C | 16.1 d, C | 14.4 f, C | 0.06 |
| 10 | 26.7 a, C | 18.8 d, C | 25.6 b, B | 21.3 c, A | 18.5 e, A | 15.9 f, B | ||
| 20 | 30.5 a, A | 20.5 c, A | 29.0 b, A | 18.5 d, B | 18.4 d, B | 18.0 e, A | ||
| Omega 6 | 0 | 27.7 b, B | 19.3 c, B | 29.0 a, A | 15.7 d, C | 15.7 d, C | 14.2 e, C | 0.06 |
| 10 | 26.3 a, C | 18.8 d, C | 25.4 b, C | 21.1 c, A | 18.5 e, A | 15.7 f, B | ||
| 20 | 30.2 a, A | 20.4 c, A | 28.8 b, B | 18.2 d, B | 18.2 d, B | 17.9 e, A |
| Substrate | N | No. of Larvae x Total Weight | Total Weight x Phenols | Total Weight x Flavonoids | Total Weight x ABTS | Total Weight x FRAP | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| r | Sig. | r | Sig. | r | Sig. | r | Sig. | r | Sig. | ||
| WB | 9 | 0.915 | ** | −0.52 | NS | 0.761 | * | −0.719 | * | −0.387 | NS |
| RB | 9 | 0.896 | ** | 0.293 | NS | 0.166 | NS | 0.646 | NS | 0.766 | * |
| CC | 9 | 0.578 | * | 0.031 | NS | −0.131 | NS | −0.402 | NS | 0.278 | NS |
| OR | 9 | 0.444 | NS | 0.910 | ** | 0.911 | ** | 0.916 | ** | 0.786 | ** |
| BR | 9 | 0.177 | NS | 0.359 | NS | 0.681 | * | 0.773 | * | 0.470 | NS |
| PP | 9 | 0.404 | NS | 0.271 | NS | 0.163 | NS | 0.341 | NS | 0.312 | NS |
| All substrates | 54 | 0.814 | ** | −0.016 | NS | −0.416 | ** | −0.584 | ** | 0.980 | NS |
| Basic substrate | 17 | 0.571 | * | 0.649 | ** | −0.374 | NS | 0.371 | NS | 0.393 | NS |
| Substrate | N | Correlation between Substrates Contents x Larvae Contents | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Phenols | Total Flavonoids | ABTS | FRAP | ||||||
| r | Significance | r | Significance | r | Significance | r | Significance | ||
| WB | 9 | −0.673 | * | 0.283 | NS | −0.039 | NS | −0.226 | NS |
| RB | 9 | 0.385 | NS | 0.087 | NS | 0.833 | * | 0.786 | * |
| CC | 9 | 0.922 | ** | 0.169 | NS | 0.550 | NS | 0.922 | ** |
| OR | 9 | 0.951 | ** | 0.819 | ** | 0.851 | ** | 0.943 | ** |
| BR | 9 | 0.362 | NS | 0.841 | ** | 0.786 | ** | 0.579 | NS |
| PP | 9 | 0.690 | * | −0.352 | NS | 0.738 | * | −0.436 | NS |
| All substrates | 54 | 0.417 | ** | 0.504 | ** | 0.357 | ** | 0.641 | ** |
| Basic substrate | 17 | 0.450 | NS | 0.460 | NS | 0.353 | NS | 0.069 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreadis, S.S.; Panteli, N.; Mastoraki, M.; Rizou, E.; Stefanou, V.; Tzentilasvili, S.; Sarrou, E.; Chatzifotis, S.; Krigas, N.; Antonopoulou, E. Towards Functional Insect Feeds: Agri-Food By-Products Enriched with Post-Distillation Residues of Medicinal Aromatic Plants in Tenebrio molitor (Coleoptera: Tenebrionidae) Breeding. Antioxidants 2022, 11, 68. https://doi.org/10.3390/antiox11010068
Andreadis SS, Panteli N, Mastoraki M, Rizou E, Stefanou V, Tzentilasvili S, Sarrou E, Chatzifotis S, Krigas N, Antonopoulou E. Towards Functional Insect Feeds: Agri-Food By-Products Enriched with Post-Distillation Residues of Medicinal Aromatic Plants in Tenebrio molitor (Coleoptera: Tenebrionidae) Breeding. Antioxidants. 2022; 11(1):68. https://doi.org/10.3390/antiox11010068
Chicago/Turabian StyleAndreadis, Stefanos S., Nikolas Panteli, Maria Mastoraki, Eleftheria Rizou, Vassilia Stefanou, Sofia Tzentilasvili, Eirini Sarrou, Stavros Chatzifotis, Nikos Krigas, and Efthimia Antonopoulou. 2022. "Towards Functional Insect Feeds: Agri-Food By-Products Enriched with Post-Distillation Residues of Medicinal Aromatic Plants in Tenebrio molitor (Coleoptera: Tenebrionidae) Breeding" Antioxidants 11, no. 1: 68. https://doi.org/10.3390/antiox11010068
APA StyleAndreadis, S. S., Panteli, N., Mastoraki, M., Rizou, E., Stefanou, V., Tzentilasvili, S., Sarrou, E., Chatzifotis, S., Krigas, N., & Antonopoulou, E. (2022). Towards Functional Insect Feeds: Agri-Food By-Products Enriched with Post-Distillation Residues of Medicinal Aromatic Plants in Tenebrio molitor (Coleoptera: Tenebrionidae) Breeding. Antioxidants, 11(1), 68. https://doi.org/10.3390/antiox11010068