Investigation of Biochemical Alterations in Ischemic Stroke Using Fourier Transform Infrared Imaging Spectroscopy—A Preliminary Study

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Block and Tissue Preparation
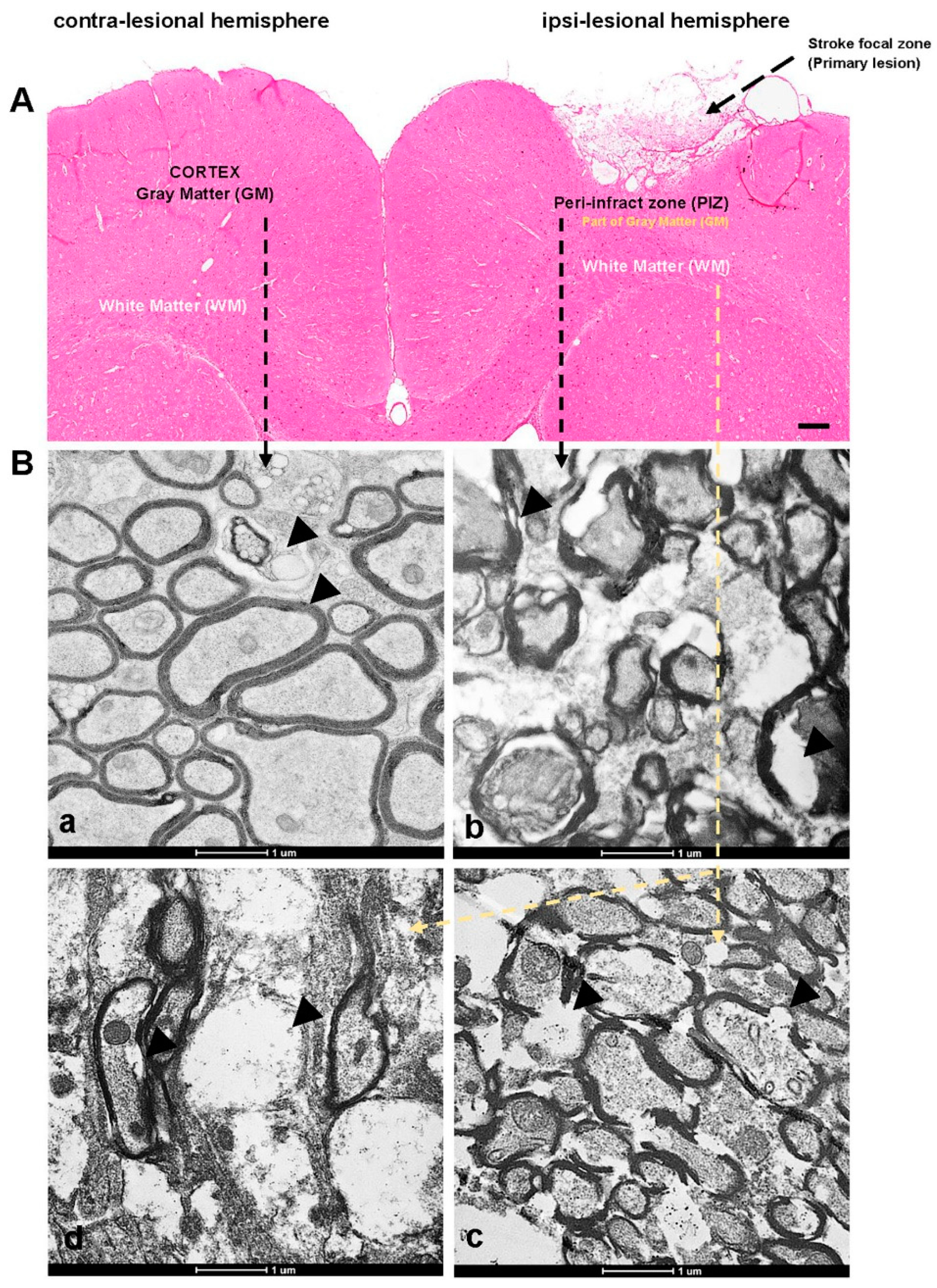
2.3. Transmission Electron Microscopy
2.4. FTIR Data Acquisition and Pre-Processing
2.5. Statistical Analysis
3. Results
3.1. Transmission Electron Microscope Studies to Define Ultrastructural Changes
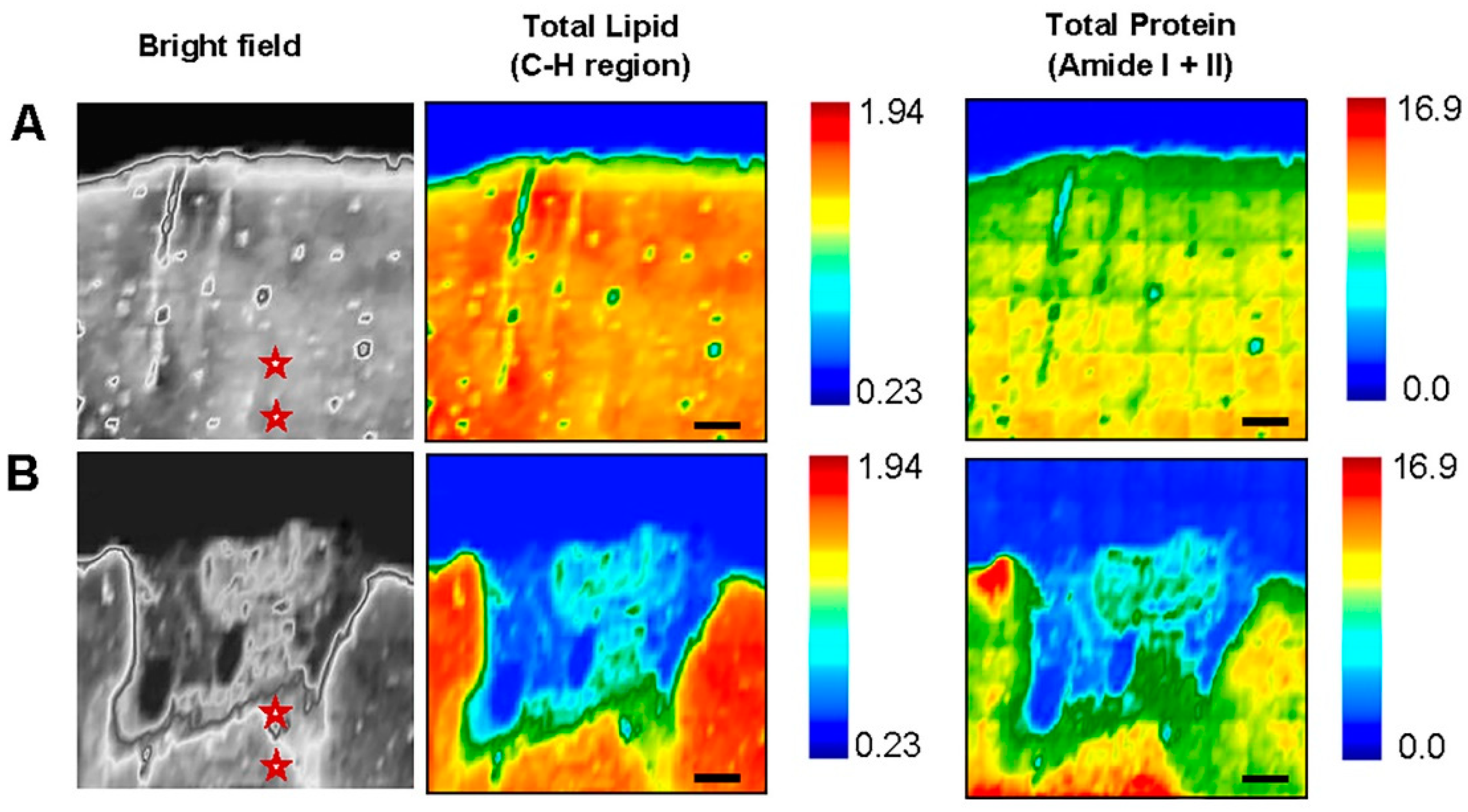
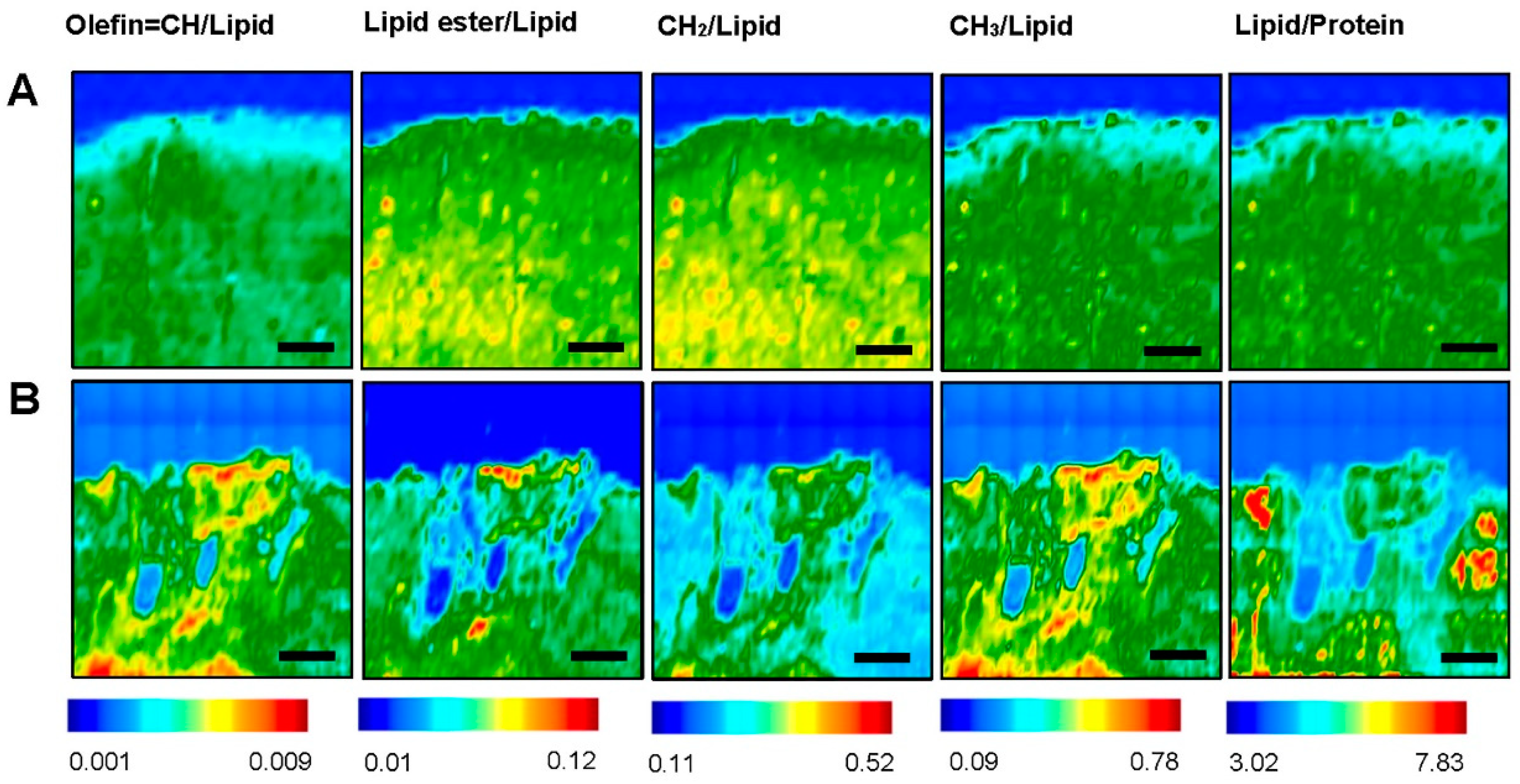
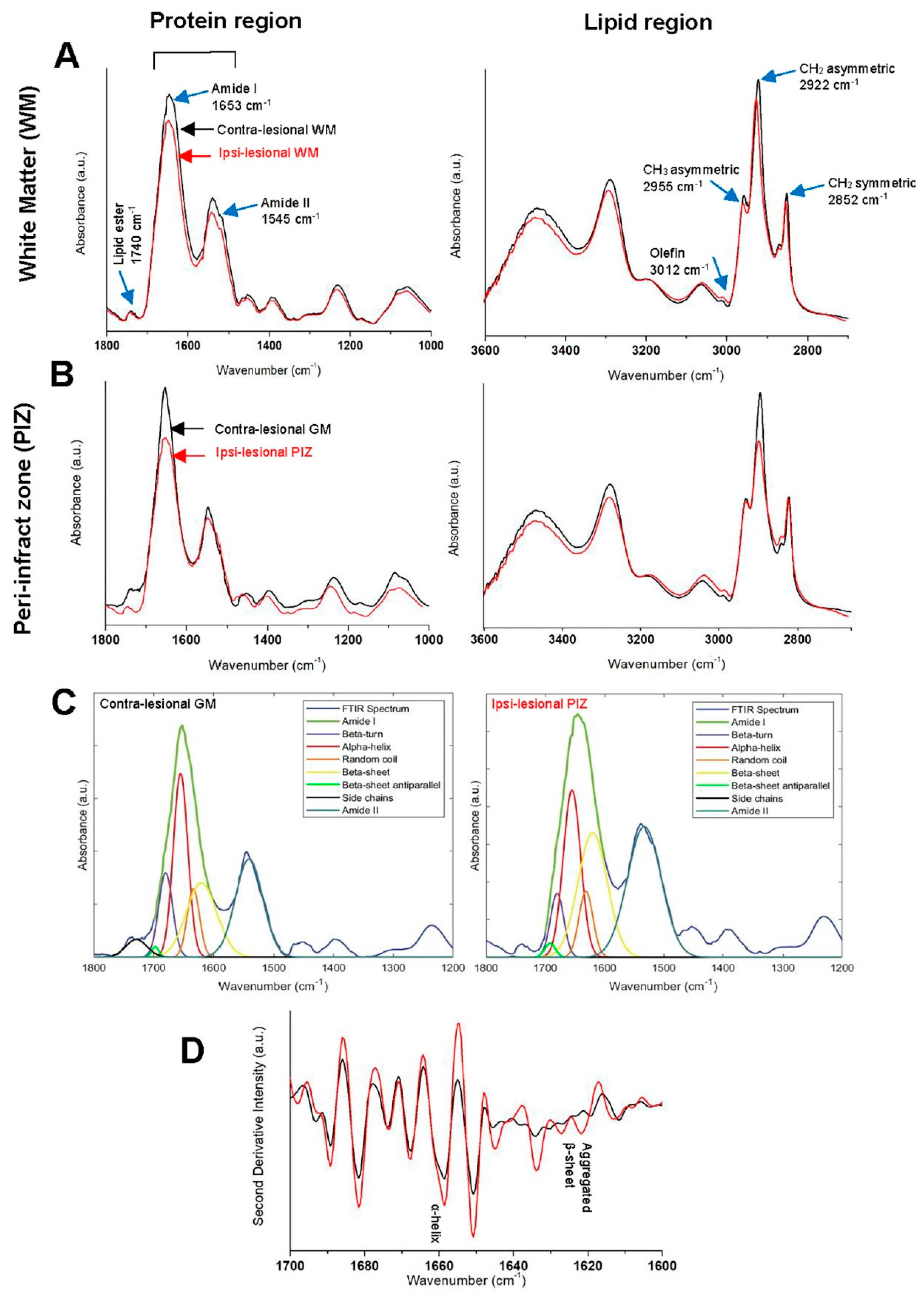
3.2. Fourier Transform Infrared Imaging Spectroscopic Studies to Identify Biochemical and Molecular Changes
3.2.1. Contra-Lesional WM vs Ipsi-Lesional WM
3.2.2. Contra-Lesional GM vs Ipsi-Lesional PIZ
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hossmann, K.-A. Pathophysiology and therapy of experimental stroke. Cell. Mol. Neurobiol. 2006, 26, 1055–1081. [Google Scholar] [CrossRef]
- Sommer, C.J. Ischemic stroke: Experimental models and reality. Acta Neuropathol. 2017, 133, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Mergenthaler, P.; Dirnagl, U.; Meisel, A. Pathophysiology of stroke: Lessons from animal models. Metab. Brain Dis. 2004, 19, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Dirnagl, U.; Iadecola, C.; Moskowitz, M.A. Pathobiology of ischaemic stroke. Trends Neurosci. 1999, 22, 391–397. [Google Scholar] [CrossRef]
- Ali, M.H.M.; Rakib, F.; Abdelalim, E.M.; Limbeck, A.; Mall, R.; Ullah, E.; Mesaeli, N.; McNaughton, D.; Ahmed, T.; Al-Saad, K. Fourier-Transform Infrared Imaging Spectroscopy and Laser Ablation -ICPMS New Vistas for Biochemical Analyses of Ischemic Stroke in Rat Brain. Front. Mol. Neurosci. 2018, 12, 647. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H.; Dalkara, T.; Moskowitz, M.A. Mechanisms, challenges and opportunities in stroke. Nat. Rev. Neurosci. 2003, 4, 399–414. [Google Scholar] [CrossRef]
- Siesjö, B.K. Acidosis and ischemic brain damage. Neurochem. Pathol. 1988, 9, 31–88. [Google Scholar] [CrossRef]
- Woodruff, T.M.; Thundyil, J.; Tang, S.-C.; Sobey, C.G.; Taylor, S.M.; Arumugam, T.V. Pathophysiology, treatment, and animal and cellular models of human ischemic stroke. Mol. Neurodegener. 2011, 6, 11. [Google Scholar] [CrossRef]
- Pekny, M.; Nilsson, M. Astrocyte activation and reactive gliosis. Glia 2005, 50, 427–434. [Google Scholar] [CrossRef]
- Wan, J.; Ren, H.; Wang, J. Iron toxicity, lipid peroxidation and ferroptosis after intracerebral haemorrhage. Stroke Vasc. Neurol. 2019, 4, 93–95. [Google Scholar] [CrossRef]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Westlake, K.P.; Nagarajan, S.S. Functional Connectivity in Relation to Motor Performance and Recovery After Stroke. Front. Syst. Neurosci. 2011, 5, 8. [Google Scholar] [CrossRef] [PubMed]
- Dijkhuizen, R.M.; van der Marel, K.; Otte, W.M.; Hoff, E.I.; van der Zijden, J.P.; van der Toorn, A.; van meer, M.P.A. Functional MRI and Diffusion Tensor Imaging of Brain Reorganization After Experimental Stroke. Transl. Stroke Res. 2012, 3, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Van Bruggen, N.; Cullen, B.M.; King, M.D.; Doran, M.; Williams, S.R.; Gadian, D.G.; Cremer, J.E. T2- and diffusion-weighted magnetic resonance imaging of a focal ischemic lesion in rat brain. Stroke 1992, 23, 576–582. [Google Scholar] [CrossRef]
- Dijkhuizen, R.M.; Singhal, A.B.; Mandeville, J.B.; Wu, O.; Halpern, E.F.; Finklestein, S.P.; Rosen, B.R.; Lo, E.H. Correlation between brain reorganization, ischemic damage, and neurologic status after transient focal cerebral ischemia in rats: A functional magnetic resonance imaging study. J. Neurosci. 2003, 23, 510–517. [Google Scholar] [CrossRef]
- Dijkhuizen, R.M.; Beekwilder, J.P.; Van Der Worp, H.B.; Van Der Sprenkel, J.W.B.; Tulleken, K.A.F.; Nicolay, K. Correlation between tissue depolarizations and damage in focal ischemic rat brain. Brain Res. 1999, 840, 194–205. [Google Scholar] [CrossRef]
- Dietrich, W.D.; Ginsberg, M.D.; Busto, R.; Watson, B.D. Photochemically induced cortical infarction in the Rat. 2. Acute and subacute alterations in local glucose utilization. Br. J. Pharmacol. 1986, 6, 195–202. [Google Scholar] [CrossRef]
- Pevsner, P.H.; Eichenbaum, J.W.; Miller, D.C.; Pivawer, G.; Eichenbaum, K.D.; Stern, A.; Zakian, K.L.; Koutcher, J.A. A photothrombotic model of small early ischemic infarcts in the rat brain with histologic and MRI correlation. J. Pharmacol. Toxicol. Methods 2001, 45, 227–233. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Xi, G.; Hua, Y.; Nagaraja, T.N.; Fenstermacher, J.D.; Keep, R.F. Development of a rat model of photothrombotic ischemia and infarction within the caudoputamen. Stroke 2009, 40, 248–253. [Google Scholar] [CrossRef]
- Ali, M.H.M.; Rakib, F.; Al-Saad, K.; Al-Saady, R.; Goormaghtigh, E. An Innovative Platform Merging Elemental Analysis and Ftir Imaging for Breast Tissue Analysis. Sci. Rep. 2019, 9, 9854. [Google Scholar] [CrossRef]
- Levin, I.W.; Bhargava, R. Fourier transform infrared vibrational spectroscopic imaging: Integrating microscopy and molecular recognition. Annu. Rev. Phys. Chem. 2005, 56, 429–474. [Google Scholar] [CrossRef] [PubMed]
- Caine, S.; Hackett, M.J.; Hou, H.; Kumar, S.; Maley, J.; Ivanishvili, Z.; Suen, B.; Szmigielski, A.; Jiang, Z.; Sylvain, N.J.; et al. A novel multi-modal platform to image molecular and elemental alterations in ischemic stroke. Neurobiol. Dis. 2016, 91, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.H.M.; Rakib, F.; Nischwitz, V.; Ullah, E.; Mall, R.; Shraim, A.M.; Ahmad, M.I.; Ghouri, Z.K.; McNaughton, D.; Küppers, S.; et al. Application of FTIR and LA-ICPMS Spectroscopies as a Possible Approach for Biochemical Analyses of Different Rat Brain Regions. Appl. Sci. 2018, 8, 2436. [Google Scholar] [CrossRef]
- Petibois, C.; Wehbe, K.; Belbachir, K.; Noreen, R.; Déléris, G. Current trends in the development of FTIR imaging for the quantitative analysis of biological samples. Acta Phys. Pol. A 2009, 115, 507. [Google Scholar] [CrossRef]
- Miller, L.M.; Bourassa, M.W.; Smith, R.J. FTIR spectroscopic imaging of protein aggregation in living cells. Biochim. et Biophys. Acta Bioenergy 2013, 1828, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, F.S.; Adamcik, J.; Jeong, J.S.; Lashuel, H.A.; Mezzenga, R.; Dietler, G. Influence of the β-Sheet Content on the Mechanical Properties of Aggregates during Amyloid Fibrillization. Angew. Chem. Int. Ed. 2015, 54, 2462–2466. [Google Scholar] [CrossRef]
- Hackett, M.J.; Aitken, J.B.; El-Assaad, F.; McQuillan, J.A.; Carter, E.A.; Ball, H.J.; Tobin, M.J.; Paterson, D.; De Jonge, M.D.; Siegele, R.; et al. Mechanisms of murine cerebral malaria: Multimodal imaging of altered cerebral metabolism and protein oxidation at hemorrhage sites. Sci. Adv. 2015, 1, e1500911. [Google Scholar] [CrossRef]
- Szczerbowska-Boruchowska, M.; Dumas, P.; Kastyak, M.Z.; Chwiej, J.; Lankosz, M.; Adamek, D.; Krygowska-Wajs, A. Biomolecular investigation of human substantia nigra in Parkinson’s disease by synchrotron radiation Fourier transform infrared microspectroscopy. Arch. Biochem. Biophys. 2007, 459, 241–248. [Google Scholar] [CrossRef]
- Sarroukh, R.; Goormaghtigh, E.; Ruysschaert, J.-M.; Raussens, V. ATR-FTIR: A “rejuvenated” tool to investigate amyloid proteins. Biochim. Biophys. Acta Biomembr. 2013, 1828, 2328–2338. [Google Scholar] [CrossRef]
- Choo, L.-P.; Mansfield, J.R.; Pizzi, N.; Somorjai, R.L.; Jackson, M.; Halliday, W.C.; Mantsch, H.H. Infrared spectra of human central nervous system tissue: Diagnosis of alzheimer’s disease by multivariate analyses. Biospectroscopy 1995, 1, 141–148. [Google Scholar] [CrossRef]
- Maquelin, K.; Kirschner, C.; Choo-Smith, L.-P.; Ngo-Thi, N.A.; Van Vreeswijk, T.; Stämmler, M.; Endtz, H.P.; Bruining, H.A.; Naumann, D.; Puppels, G.J. Prospective study of the performance of vibrational spectroscopies for rapid identification of bacterial and fungal pathogens recovered from blood cultures. J. Clin. Microbiol. 2003, 41, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, S.T. Cellular and molecular mechanisms of neural repair after stroke: Making waves. Ann. Neurol. 2006, 59, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Molecular basis of long-term plasticity underlying addiction. Nat. Rev. Neurosci. 2001, 2, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, N.; Niwa, S.; Tanaka, Y. Molecular motors in neurons: Transport mechanisms and roles in brain function, development, and disease. Neuron 2010, 68, 610–638. [Google Scholar] [CrossRef]
- Hackett, M.J.; Sylvain, N.J.; Hou, H.; Caine, S.; Alaverdashvili, M.; Pushie, M.J.; Kelly, M.E. Concurrent glycogen and lactate imaging with FTIR spectroscopy to spatially localize metabolic parameters of the glial response following brain ischemia. Anal. Chem. 2016, 88, 10949–10956. [Google Scholar] [CrossRef]
- Gazi, E.; Baker, M.; Dwyer, J.; Lockyer, N.P.; Gardner, P.; Shanks, J.H.; Reeve, R.S.; Hart, C.A.; Clarke, N.W.; Brown, M.D. A Correlation of FTIR Spectra Derived from Prostate Cancer Biopsies with Gleason Grade and Tumour Stage. Eur. Urol. 2006, 50, 750–761. [Google Scholar] [CrossRef]
- Sahu, R.; Mordechai, S. Fourier transform infrared spectroscopy in cancer detection. Futur. Oncol. 2005, 1, 635–647. [Google Scholar] [CrossRef]
- Richter, T.; Steiner, G.; Abu-Id, M.H.; Salzer, R.; Bergmann, R.; Rodig, H.; Johannsen, B. Identification of tumor tissue by FTIR spectroscopy in combination with positron emission tomography. Vib. Spectrosc. 2002, 28, 103–110. [Google Scholar] [CrossRef]
- Uzdensky, A.B. Photothrombotic Stroke as a Model of Ischemic Stroke. Transl. Stroke Res. 2018, 9, 437–451. [Google Scholar] [CrossRef]
- Liu, W.; Wang, X.; Yang, S.; Huang, J.; Xue, X.; Zheng, Y.; Shang, G.; Tao, J.; Chen, L. Electroacupunctre improves motor impairment via inhibition of microglia-mediated neuroinflammation in the sensorimotor cortex after ischemic stroke. Life Sci. 2016, 151, 313–322. [Google Scholar] [CrossRef]
- Sinke, M.R.T.; Otte, W.M.; van Meer, M.P.A.; van der Toorn, A.; Dijkhuizen, R.M. Modified structural network backbone in the contralesional hemisphere chronically after stroke in rat brain. J. Cereb. Blood Flow Metab. 2018, 38, 1642–1653. [Google Scholar] [CrossRef] [PubMed]
- Grefkes, C.; Fink, G.R. Reorganization of cerebral networks after stroke: New insights from neuroimaging with connectivity approaches. Brain 2011, 134, 1264–1276. [Google Scholar] [CrossRef] [PubMed]
- Çakmak, G.; Miller, L.M.; Zorlu, F.; Severcan, F. Amifostine, a radioprotectant agent, protects rat brain tissue lipids against ionizing radiation induced damage: An FTIR microspectroscopic imaging study. Arch. Biochem. Biophys. 2012, 520, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Petibois, C.; Desbat, B. Clinical application of FTIR imaging: New reasons for hope. Trends Biotechnol. 2010, 28, 495–500. [Google Scholar] [CrossRef]
- Hackett, M.J.; DeSouza, M.; Caine, S.; Bewer, B.; Nichol, H.; Paterson, P.G.; Colbourne, F. A new method to image heme-Fe, total Fe, and aggregated protein levels after intracerebral hemorrhage. ACS Chem. Neurosci. 2015, 6, 761–770. [Google Scholar] [CrossRef]
- Shirley, R.; Ord, E.N.J.; Work, L.M. Oxidative stress and the use of antioxidants in stroke. Antioxidants 2014, 3, 472–501. [Google Scholar] [CrossRef]
- Cipolla, M.J.; Crete, R.; Vitullo, L.; Rix, R.D. Transcellular transport as a mechanism of blood-brain barrier disruption during stroke. Front. Biosci. 2004, 9, 777–785. [Google Scholar] [CrossRef]
- Solenski, N.J.; Dipierro, C.G.; Trimmer, P.A.; Kwan, A.-L.; Helms, G.A. Ultrastructural changes of neuronal mitochondria after transient and permanent cerebral ischemia. Stroke 2002, 33, 816–824. [Google Scholar] [CrossRef]
- Szalontai, B.; Nishiyama, Y.; Gombos, Z.; Murata, N. Membrane dynamics as seen by Fourier transform infrared spectroscopy in a cyanobacterium, Synechocystis PCC 6803—The effects of lipid unsaturation and the protein-to-lipid ratio. Biochim. Biophys. Acta Biomembr. 2000, 1509, 409–419. [Google Scholar] [CrossRef]
- Toyran, N.; Zorlu, F.; Dönmez, G.; Öǧe, K.; Severcan, F. Chronic hypoperfusion alters the content and structure of proteins and lipids of rat brain homogenates: A Fourier transform infrared spectroscopy study. Eur. Biophys. J. 2004, 33, 549–554. [Google Scholar] [CrossRef]
- Schreibelt, G.; Kooij, G.; Reijerkerk, A.; Van Doorn, R.; Gringhuis, S.I.; Van Der Pol, S.; Weksler, B.B.; Romero, I.A.; Couraud, P.O.; Piontek, J.; et al. Reactive oxygen species alter brain endothelial tight junction dynamics via RhoA, PI3 kinase, and PKB signaling. FASEB J. 2007, 21, 3666–3676. [Google Scholar] [CrossRef] [PubMed]
- Magura, I.S.; Rozhmanova, O.M. Oxidative stress and neurodegenerative disorders. Biopolym. Cell 1997, 13, 513–515. [Google Scholar] [CrossRef]
- Emerit, J.; Edeas, M.; Bricaire, F. Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 2004, 58, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative Stress in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Lamba, O.P.; Borchman, D.; Garner, W.H. Spectral characterization of lipid peroxidation in rabbit lens membranes induced by hydrogen peroxide in the presence of Fe2+ Fe3+ cations: A site-specific catalyzed oxidation. Free Radic. Biol. Med. 1994, 16, 591–601. [Google Scholar] [CrossRef]
- De Zwart, L.L.; Meerman, J.H.N.; Commandeur, J.N.M.; Vermeulen, N.P.E. Biomarkers of free radical damage applications in experimental animals and in humans. Free Radic. Biol. Med. 1999, 26, 202–226. [Google Scholar] [CrossRef]
- Adibhatla, R.M.; Hatcher, J.F. Altered Lipid Metabolism in Brain Injury and Disorders. In Lipids in Health and Disease; Springer: Dordrecht, The Netherlands, 2008; pp. 241–268. [Google Scholar]
- Romont, C.; Marie, C.; Bralet, J. Increased lipid peroxidation in vulnerable brain regions after transient forebrain ischemia in rats. Stroke 1989, 20, 918–924. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Slater, T.F. Free-radical mechanisms in tissue injury. Biochem. J. 1984, 222, 1–15. [Google Scholar] [CrossRef]
- Riley, P. Free radicals in biology: Oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Boil. 1994, 65, 27–33. [Google Scholar] [CrossRef]
- Spitz, D.R.; Azzam, E.I.; Li, J.J.; Gius, D. Metabolic oxidation/reduction reactions and cellular responses to ionizing radiation: A unifying concept in stress response biology. Cancer Metastasis Rev. 2004, 23, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Sowa, M.G.; Mantsch, H.H. Infrared spectroscopy: A new frontier in medicine. Biophys. Chem. 1997, 68, 109–125. [Google Scholar] [CrossRef]
- Xing, C.; Arai, K.; Lo, E.H.; Hommel, M. Pathophysiologic cascades in ischemic stroke. Int. J. Stroke 2012, 7, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Lipton, P. Ischemic cell death in brain neurons. Physiol. Rev. 1999, 79, 1431–1568. [Google Scholar] [CrossRef]
- Sen, N.; Hara, M.R.; Ahmad, A.S.; Cascio, M.B.; Kamiya, A.; Ehmsen, J.T.; Aggrawal, N.; Hester, L.; Doré, S.; Snyder, S.H.; et al. GOSPEL: A neuroprotective protein that binds to GAPDH upon S-Nitrosylation. Neuron 2009, 63, 709. [Google Scholar] [CrossRef]
- Squitti, R.; Siotto, M.; Assenza, G.; Giannantoni, N.M.; Rongioletti, M.; Zappasodi, F.; Tecchio, F. Prognostic value of serum copper for post-stroke clinical recovery: A pilot study. Front. Neurol. 2018, 9, 333. [Google Scholar] [CrossRef]
- DeGregorio-Rocasolano, N.; Martí-Sistac, O.; Ponce, J.; Castelló-Ruiz, M.; Millán, M.; Guirao, V.; García-Yébenes, I.; Salom, J.B.; Ramos-Cabrer, P.; Alborch, E.; et al. Iron-loaded transferrin (Tf) is detrimental whereas iron-free Tf confers protection against brain ischemia by modifying blood Tf saturation and subsequent neuronal damage. Redox Biol. 2018, 15, 143–158. [Google Scholar] [CrossRef]
- Qin, A.-P.; Zhang, H.-L.; Qin, Z.-H. Mechanisms of lysosomal proteases participating in cerebral ischemia-induced neuronal death. Neurosci. Bull. 2008, 24, 117–123. [Google Scholar] [CrossRef][Green Version]
- Ozek, N.S.; Tuna, S.; Erson-Bensan, A.E.; Severcan, F. Characterization of microRNA-125b expression in MCF7 breast cancer cells by ATR-FTIR spectroscopy. Analyst 2010, 135, 3094–3102. [Google Scholar] [CrossRef]
- Wang, Q.; Kretlow, A.; Beekes, M.; Naumann, D.; Miller, L. In situ characterization of prion protein structure and metal accumulation in scrapie-infected cells by synchrotron infrared and X-ray imaging. Vib. Spectrosc. 2005, 38, 61–69. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infrared Band Assignment | Spectral Range (cm−1) | Comments | |
|---|---|---|---|
| Amide I + II | 1700–1500 | Total protein region | |
| Protein components | Amide I | 1700–1600 | Specifically sensitive to protein secondary structure |
| Amide II | 1555–1535 | Mainly ν(C–N) associated with proteins | |
| β-sheet | 1610–1635 | Functional protein, protein secondary structure | |
| α-helix | 1660–1650 | Functional protein, protein secondary structure | |
| Random coil | 1645–1630 | protein secondary structure | |
| C-H region | 3100–2800 | Total lipid region | |
| Lipid component | νsCH2 | 2852–2800 | to measure lipid acyl chain length |
| νasCH2 | 2915–2930 | ||
| νasCH3 | 2950–2960 | to measure concentration of methyl content | |
| ν (C = O) | 1755–1715 | to measure oxidative stress | |
| ν (olefinic = CH) | 3000–3027 | to measure unsaturated lipid |
| ROI | Total Lipid | Total Protein | Lipid/Protein | CH2/Lipid | CH3/Lipid | Olefin/Lipid | Lipid Ester/Lipid | |
|---|---|---|---|---|---|---|---|---|
| Contra-lesional | GM | 0.69 ± 0.022 * | 14.881 ± 0.043 * | 5.887 ± 0.0013 ** | 0.233 ± 0.0001 ** | 0.165 ± 0.0003 ** | 0.0032 ± 0.0003 ** | 0.063 ± 0.0051 ** |
| WM | 1.39 ± 0.072 * | 9.061 ± 0.0014 * | 6.991 ± 0.0013 ** | 0.341 ± 0.0002 ** | 0.281 ± 0.0002 ** | 0.0045 ± 0.0003 ** | 0.086 ± 0.0031 ** | |
| Ipsi-lesional | PIZ | 0.59 ± 0.032 ** | 11.003 ± 0.012 ** | 5.256 ± 0.0033 ** | 0.194 ± 0.0002 ** | 0.394 ± 0.0003 ** | 0.0052 ± 0.0005 ** | 0.057 ± 0.0062 ** |
| WM | 0.93 ± 0.052 ** | 8.995 ± 0.0014 ** | 6.367 ± 0.0013 ** | 0.125 ± 0.0001 ** | 0.399 ± 0.0006 ** | 0.0062 ± 0.0003 ** | 0.067 ± 0.0033 ** |
| Location of Brain | Contra-Lesional GM | Ipsi-Lesional PIZ | |
|---|---|---|---|
| Ratios | |||
| α-helix/Amide I | 51% * | 32% * | |
| β-sheet/Amide I | 37% * | 66% * | |
| Random coil/Amide I | 12% * | 10% * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakib, F.; Ali, C.M.; Yousuf, M.; Afifi, M.; Bhatt, P.R.; Ullah, E.; Al-Saad, K.; Ali, M.H.M. Investigation of Biochemical Alterations in Ischemic Stroke Using Fourier Transform Infrared Imaging Spectroscopy—A Preliminary Study. Brain Sci. 2019, 9, 293. https://doi.org/10.3390/brainsci9110293
Rakib F, Ali CM, Yousuf M, Afifi M, Bhatt PR, Ullah E, Al-Saad K, Ali MHM. Investigation of Biochemical Alterations in Ischemic Stroke Using Fourier Transform Infrared Imaging Spectroscopy—A Preliminary Study. Brain Sciences. 2019; 9(11):293. https://doi.org/10.3390/brainsci9110293
Chicago/Turabian StyleRakib, Fazle, Carmen M. Ali, Mohammed Yousuf, Mohammed Afifi, Pooja R. Bhatt, Ehsan Ullah, Khalid Al-Saad, and Mohamed H. M. Ali. 2019. "Investigation of Biochemical Alterations in Ischemic Stroke Using Fourier Transform Infrared Imaging Spectroscopy—A Preliminary Study" Brain Sciences 9, no. 11: 293. https://doi.org/10.3390/brainsci9110293
APA StyleRakib, F., Ali, C. M., Yousuf, M., Afifi, M., Bhatt, P. R., Ullah, E., Al-Saad, K., & Ali, M. H. M. (2019). Investigation of Biochemical Alterations in Ischemic Stroke Using Fourier Transform Infrared Imaging Spectroscopy—A Preliminary Study. Brain Sciences, 9(11), 293. https://doi.org/10.3390/brainsci9110293