
Cognitive Control of Working Memory: A Model-Based Approach

 , , , and
, , , and
Abstract
1. Introduction
1.1. Measuring WM Control Processes with the Reference-Back Paradigm
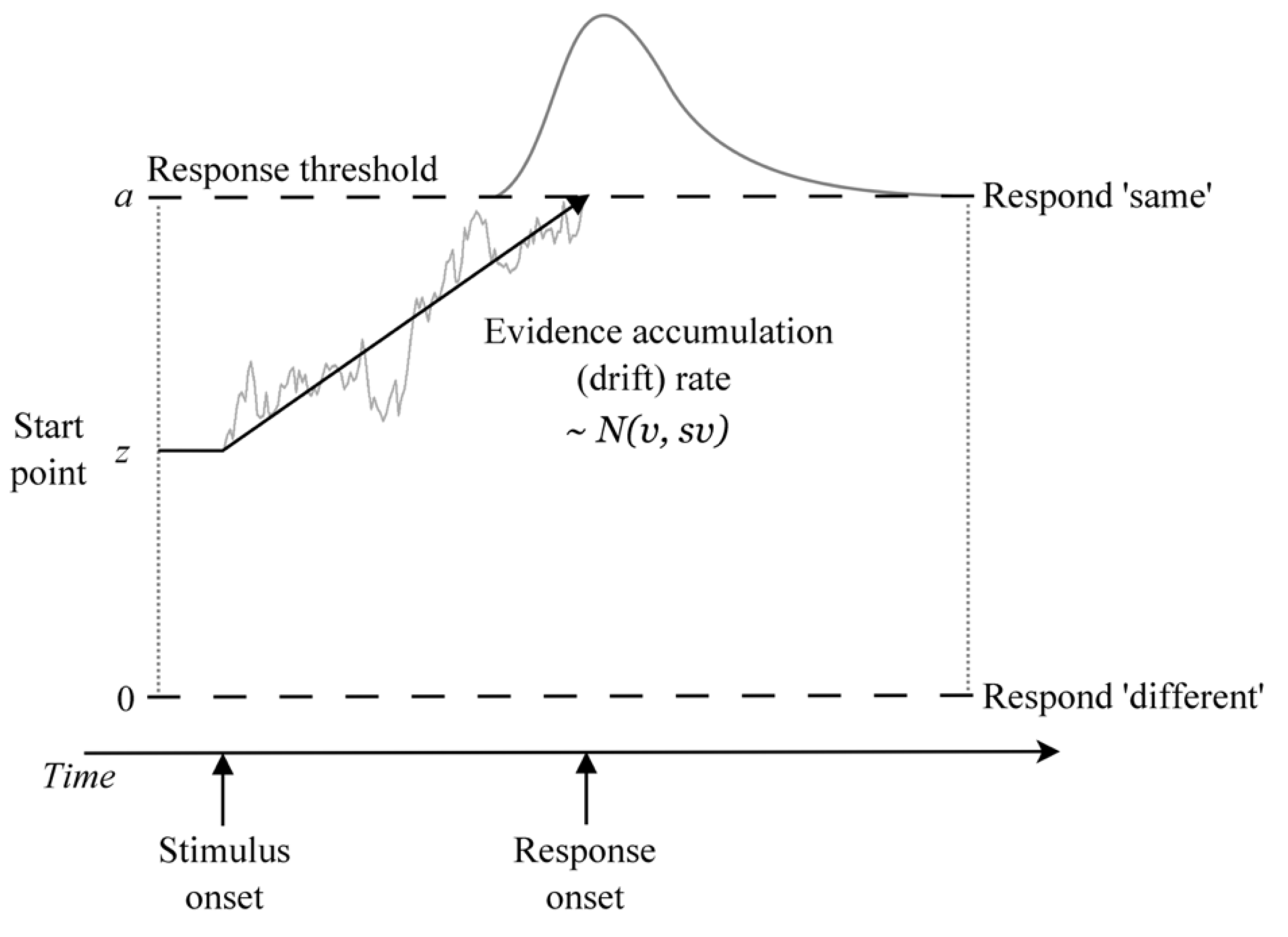
1.2. The Diffusion Decision Model
1.3. Current Study
2. Materials and Methods
2.1. Participants
2.2. Stimuli and Procedure
2.3. Analysis Methods
3. Results
3.1. Conventional Analyses
3.2. Diffusion Decision Model Analysis
3.3. Model Fit and Parameter Recovery
3.4. Parameter Effects
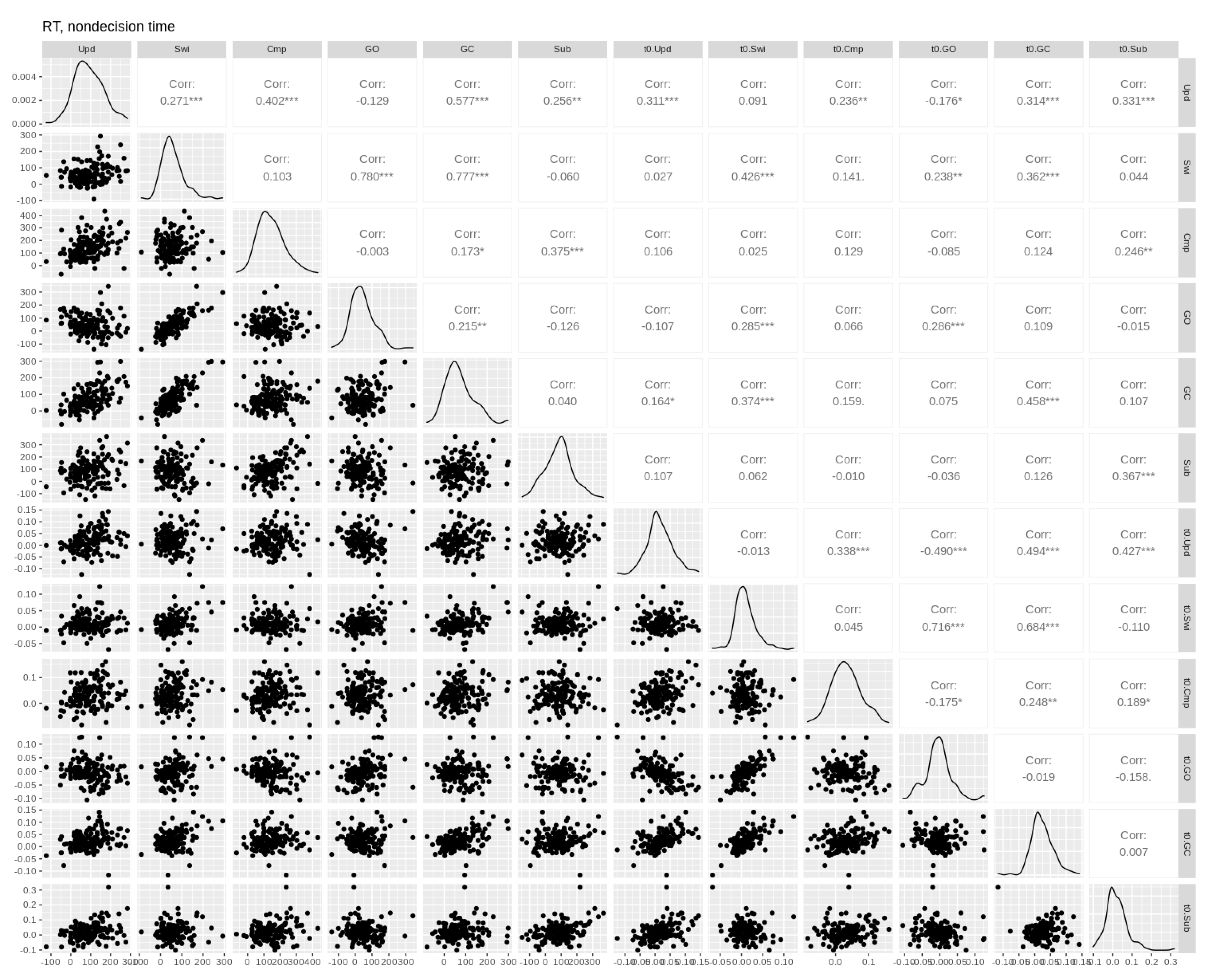
3.5. Individual Differences
4. Discussion
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Significance Tests of Behavioral Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | SS | dfeffect | Residual SS | dfwithin | F | p |
|---|---|---|---|---|---|---|
| Intercept | 996.33 | 1 | 9.59 | 149 | 15,485.87 | <0.001 |
| U | 0.05 | 1 | 0.32 | 149 | 23.48 | <0.001 |
| C | 0.03 | 1 | 0.86 | 149 | 4.91 | 0.028 |
| S | 0.00 | 1 | 0.26 | 149 | 0.54 | 0.466 |
| U × C | 0.05 | 1 | 0.61 | 149 | 11.80 | <0.001 |
| U × S | 0.04 | 1 | 0.33 | 149 | 16.28 | <0.001 |
| C × S | 0.03 | 1 | 0.22 | 149 | 22.59 | <0.001 |
| U × C × S | 0.00 | 1 | 0.27 | 149 | 1.54 | 0.217 |
| Effect | SS | dfeffect | Residual SS | dfwithin | F | p |
|---|---|---|---|---|---|---|
| Intercept | 623.25 | 1 | 33.94 | 149 | 2736.57 | <0.001 |
| U | 1.81 | 1 | 1.07 | 149 | 253.14 | <0.001 |
| C | 7.34 | 1 | 2.27 | 149 | 482.69 | <0.001 |
| S | 1.04 | 1 | 0.89 | 149 | 173.75 | <0.001 |
| U × C | 0.60 | 1 | 0.68 | 149 | 131.42 | <0.001 |
| U × S | 0.06 | 1 | 0.57 | 149 | 14.44 | <0.001 |
| C × S | 0.04 | 1 | 0.36 | 149 | 18.29 | <0.001 |
| U × C × S | 0.14 | 1 | 0.49 | 149 | 41.28 | <0.001 |
Appendix B
Appendix B.1. Sampling
Appendix B.2. Priors
Appendix C
Parameter Recovery

Appendix D
Individual Difference Correlations






References
- Durstewitz, D.; Seamans, J.K. The computational role of dopamine D1 receptors in working memory. Neural Netw. 2002, 15, 561–572. [Google Scholar] [CrossRef]
- Dreisbach, G. Mechanisms of Cognitive Control. Curr. Dir. Psychol. Sci. 2012, 21, 227–231. [Google Scholar] [CrossRef]
- Dreisbach, G.; Fröber, K. On How to Be Flexible (or Not): Modulation of the Stability-Flexibility Balance. Curr. Dir. Psychol. Sci. 2019, 28, 3–9. [Google Scholar] [CrossRef]
- Hommel, B. Between persistence and flexibility: The Yin and Yang of action control. In Advances in Motivation Science; Elsevier: Amsterdam, The Netherlands, 2015; Volume 2, pp. 33–67. [Google Scholar]
- Oberauer, K. Design for a Working Memory. Psychol. Learn. Motiv. Adv. Res. Theory 2009, 51, 45–100. [Google Scholar]
- Bledowski, C.; Kaiser, J.; Rahm, B. Basic operations in working memory: Contributions from functional imaging studies. Behav. Brain Res. 2010, 214, 172–179. [Google Scholar] [CrossRef]
- Ecker, U.K.H.; Oberauer, K.; Lewandowsky, S. Working memory updating involves item-specific removal. J. Mem. Lang. 2014, 74, 1–15. [Google Scholar] [CrossRef]
- Ecker, U.K.H.; Lewandowsky, S.; Oberauer, K. Removal of information from working memory: A specific updating process. J. Mem. Lang. 2014, 74, 77–90. [Google Scholar] [CrossRef]
- Murty, V.P.; Sambataro, F.; Radulescu, E.; Altamura, M.; Iudicello, J.; Zoltick, B.; Weinberger, D.R.; Goldberg, T.E.; Mattay, V.S. Selective updating of working memory content modulates meso-cortico-striatal activity. Neuroimage 2011, 57, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, R.C. Biologically based computational models of high-level cognition. Science 2006, 314, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Rac-Lubashevsky, R.; Kessler, Y. Dissociating working memory updating and automatic updating: The reference-back paradigm. J. Exp. Psychol. Learn. Mem. Cogn. 2016, 42, 951–969. [Google Scholar] [CrossRef] [PubMed]
- Rac-Lubashevsky, R.; Kessler, Y. Decomposing the n-back task: An individual differences study using the reference-back paradigm. Neuropsychologia 2016, 90, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.K.; Serences, J.T.; Courtney, S.M. Neural system for controlling the contents of object working memory in humans. Cereb. Cortex 2006, 16, 1595–1603. [Google Scholar] [CrossRef] [PubMed]
- Verschooren, S.; Kessler, Y.; Egner, T. Evidence for a single mechanism gating perceptual and long-term memory information into working memory. Cognition 2021, 212, 104668. [Google Scholar] [CrossRef] [PubMed]
- Trutti, A.C.; Verschooren, S.; Forstmann, B.U.; Boag, R.J. Understanding subprocesses of working memory through the lens of model-based cognitive neuroscience. Curr. Opin. Behav. Sci. 2021, 38, 57–65. [Google Scholar] [CrossRef]
- Frank, M.J.; Loughry, B.; O’Reilly, R.C. Interactions between frontal cortex and basal ganglia in working memory: A computational model. Cogn. Affect. Behav. Neurosci. 2001, 1, 137–160. [Google Scholar] [CrossRef]
- Hazy, T.E.; Frank, M.J.; O’Reilly, R.C. Banishing the homunculus: Making working memory work. Neuroscience 2006, 139, 105–118. [Google Scholar] [CrossRef]
- O’Reilly, R.C.; Frank, M.J. Making working memory work: A computational model of learning in the prefrontal cortex and basal ganglia. Neural Comput. 2006, 18, 283–328. [Google Scholar] [CrossRef]
- Braver, T.S.; Cohen, J.D. On the control of control: The role of dopamine in regulating prefrontal function and working memory. In Control of Cognitive Processes: Attention and Performance XVIII; MIT Press: Cambridge, MA, USA, 2000; pp. 713–737. [Google Scholar]
- O’Reilly, R.C.; Braver, T.S.; Cohen, J.D. A Biologically Based Computational Model of Working Memory. In Models of Working Memory; Cambridge University Press: Cambridge, UK, 1999; pp. 375–411. [Google Scholar]
- Jongkees, B.J. Baseline-dependent effect of dopamine’s precursor L-tyrosine on working memory gating but not updating. Cogn. Affect. Behav. Neurosci. 2020, 1–15. [Google Scholar] [CrossRef]
- Nir-Cohen, G.; Kessler, Y.; Egner, T. Neural substrates of working memory updating. J. Cogn. Neurosci. 2020, 32, 2285–2302. [Google Scholar] [CrossRef]
- Rac-Lubashevsky, R.; Kessler, Y. Oscillatory correlates of control over working memory gating and updating: An EEG study using the reference-back paradigm. J. Cogn. Neurosci. 2018, 30, 1870–1882. [Google Scholar] [CrossRef]
- Rac-Lubashevsky, R.; Kessler, Y. Revisiting the relationship between the P3b and working memory updating. Biol. Psychol. 2019, 148, 107769. [Google Scholar] [CrossRef]
- Rac-Lubashevsky, R.; Slagter, H.A.; Kessler, Y. Tracking Real-time changes in working memory updating and gating with the event-based eye-blink rate. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Ratcliff, R. A theory of memory retrieval. Psychol. Rev. 1978, 85, 59. [Google Scholar] [CrossRef]
- Forstmann, B.U.; Ratcliff, R.; Wagenmakers, E.-J. Sequential sampling models in cognitive neuroscience: Advantages, applications, and extensions. Annu. Rev. Psychol. 2016, 67, 641–666. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, R.; Rouder, J.N. Modeling response times for two-choice decisions. Psychol. Sci. 1998, 9, 347–356. [Google Scholar] [CrossRef]
- Donders, F.C. On the speed of mental processes. Acta Psychol. 1969, 30, 412–431. [Google Scholar] [CrossRef]
- Braver, T.S. The variable nature of cognitive control: A dual mechanisms framework. Trends Cogn. Sci. 2012, 16, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Boag, R.J.; Strickland, L.; Loft, S.; Heathcote, A. Strategic attention and decision control support prospective memory in a complex dual-task environment. Cognition 2019, 191, 103974. [Google Scholar] [CrossRef]
- Boag, R.J.; Strickland, L.; Heathcote, A.; Neal, A.; Loft, S. Cognitive control and capacity for prospective memory in complex dynamic environments. J. Exp. Psychol. Gen. 2019, 148, 2181. [Google Scholar] [CrossRef] [PubMed]
- Strickland, L.; Loft, S.; Remington, R.W.; Heathcote, A. Racing to remember: A theory of decision control in event-based prospective memory. Psychol. Rev. 2018, 125, 851–887. [Google Scholar] [CrossRef] [PubMed]
- Schall, J.D.; Palmeri, T.J.; Logan, G.D. Models of inhibitory control. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.M.; Forstmann, B.U.; Love, B.C.; Palmeri, T.J.; Van Maanen, L. Approaches to analysis in model-based cognitive neuroscience. J. Math. Psychol. 2017, 76, 65–79. [Google Scholar] [CrossRef]
- Ratcliff, R.; Smith, P.L.; Brown, S.D.; McKoon, G. Diffusion decision model: Current issues and history. Trends Cogn. Sci. 2016, 20, 260–281. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, R.; McKoon, G. The diffusion decision model: Theory and data for two-choice decision tasks. Neural Comput. 2008, 20, 873–922. [Google Scholar] [CrossRef] [PubMed]
- Green, D.M.; Swets, J.A. Signal Detection Theory and Psychophysics; Wiley & Sons: New York, NY, USA, 1966. [Google Scholar]
- Sternberg, S. High-speed scanning in human memory. Science 1966, 153, 652–654. [Google Scholar] [CrossRef]
- Ratcliff, R.; Strayer, D. Modeling simple driving tasks with a one-boundary diffusion model. Psychon. Bull. Rev. 2014, 21, 577–589. [Google Scholar] [CrossRef]
- Voss, A.; Rothermund, K.; Voss, J. Interpreting the parameters of the diffusion model: An empirical validation. Mem. Cogn. 2004, 32, 1206–1220. [Google Scholar] [CrossRef]
- Schmitz, F.; Voss, A. Components of task switching: A closer look at task switching and cue switching. Acta Psychol. 2014, 151, 184–196. [Google Scholar] [CrossRef]
- Schmitz, F.; Voss, A. Decomposing task-switching costs with the diffusion model. J. Exp. Psychol. Hum. Percept. Perform. 2012, 38, 222–250. [Google Scholar] [CrossRef] [PubMed]
- Voss, A.; Nagler, M.; Lerche, V. Diffusion models in experimental psychology: A practical introduction. Exp. Psychol. 2013, 60, 385–402. [Google Scholar] [CrossRef]
- Bogacz, R.; Wagenmakers, E.J.; Forstmann, B.U.; Nieuwenhuis, S. The neural basis of the speed-accuracy tradeoff. Trends Neurosci. 2010, 33, 10–16. [Google Scholar] [CrossRef]
- Forstmann, B.U.; Dutilh, G.; Brown, S.; Neumann, J.; Von Cramon, D.Y.; Ridderinkhof, K.R.; Wagenmakers, E.J. Striatum and pre-SMA facilitate decision-making under time pressure. Proc. Natl. Acad. Sci. USA 2008, 105, 17538–17542. [Google Scholar] [CrossRef]
- Boehm, U.; Van Maanen, L.; Forstmann, B.; Van Rijn, H. Trial-by-trial fluctuations in CNV amplitude reflect anticipatory adjustment of response caution. Neuroimage 2014, 96, 95–105. [Google Scholar] [CrossRef]
- van Maanen, L.; Brown, S.D.; Eichele, T.; Wagenmakers, E.J.; Ho, T.; Serences, J.; Forstmann, B.U. Neural correlates of trial-to-trial fluctuations in response caution. J. Neurosci. 2011, 31, 17488–17495. [Google Scholar] [CrossRef]
- de Leeuw, J.R. jsPsych: A JavaScript library for creating behavioral experiments in a Web browser. Behav. Res. Methods 2015, 47, 1–12. [Google Scholar] [CrossRef]
- Heathcote, A.; Lin, Y.S.; Reynolds, A.; Strickland, L.; Gretton, M.; Matzke, D. Dynamic models of choice. Behav. Res. Methods 2019, 51, 961–985. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Navarro, D. Learning Statistics with R: A Tutorial for Psychology Students and Other Beginners; University of Adelaide: Adelaide, Australia, 2015. [Google Scholar]
- Fox, J.; Weisberg, S. An R companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- Donkin, C.; Brown, S.D.; Heathcote, A. The overconstraint of response time models: Rethinking the scaling problem. Psychon. Bull. Rev. 2009, 16, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; van der Linde, A. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B Stat. Methodol. 2002, 64, 583–639. [Google Scholar] [CrossRef]
- Klauer, K.C. Hierarchical multinomial processing tree models: A latent-trait approach. Psychometrika 2010, 75, 70–98. [Google Scholar] [CrossRef]
- Schultz, W.; Dayan, P.; Montague, P.R. A neural substrate of prediction and reward. Science 1997, 275, 1593–1599. [Google Scholar] [CrossRef]
- Schultz, W. Predictive reward signal of dopamine neurons. J. Neurophysiol. 1998, 80, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Monsell, S.; Sumner, P.; Waters, H. Task-set reconfiguration with predictable and unpredictable task switches. Mem. Cognit. 2003, 31, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Allport, A.; Wylie, G. Task switching, stimulus-response bindings, and negative priming. In Control of Cognitive Processes: Attention and Performance XVIII; Monsell, S., Driver, J., Eds.; MIT Press: Cambridge, MA, USA, 2000; pp. 35–70. [Google Scholar]
- Hartmann, E.M.; Rey-Mermet, A.; Gade, M. Same same but different? Modeling N-1 switch cost and N-2 repetition cost with the diffusion model and the linear ballistic accumulator model. Acta Psychol. 2019, 198, 102858. [Google Scholar] [CrossRef]
- Schneider, D.W.; Anderson, J.R. Asymmetric switch costs as sequential difficulty effects. Q. J. Exp. Psychol. 2010, 63, 1873–1894. [Google Scholar] [CrossRef]
- Gilbert, S.J.; Shallice, T. Task switching: A PDP model. Cogn. Psychol. 2002, 44, 297–337. [Google Scholar] [CrossRef]
- Miletić, S.; Boag, R.J.; Forstmann, B.U. Mutual benefits: Combining reinforcement learning with sequential sampling models. Neuropsychologia 2020, 136, 107261. [Google Scholar] [CrossRef] [PubMed]
- Miletić, S.; Boag, R.J.; Trutti, A.C.; Stevenson, N.; Forstmann, B.U.; Heathcote, A. A new model of decision processing in instrumental learning tasks. Elife 2021, 10, e63055. [Google Scholar] [CrossRef] [PubMed]
- Druey, M.D. Stimulus-category and response-repetition effects in task switching: An evaluation of four explanations. J. Exp. Psychol. Learn. Mem. Cogn. 2014, 40, 125–146. [Google Scholar] [CrossRef]
- Notebaert, W.; Soetens, E. The influence of irrelevant stimulus changes on stimulus and response repetition effects. Acta Psychol. 2003, 112, 143–156. [Google Scholar] [CrossRef]
- Waszak, F.; Hommel, B.; Allport, A. Interaction of task readiness and automatic retrieval in task switching: Negative priming and competitor priming. Mem. Cogn. 2005, 33, 595–610. [Google Scholar] [CrossRef][Green Version]
- Szmalec, A.; Verbruggen, F.; Vandierendonck, A.; Kemps, E. Control of Interference During Working Memory Updating. J. Exp. Psychol. Hum. Percept. Perform. 2011, 37, 137–151. [Google Scholar] [CrossRef]
- Logan, G.D.; Gordon, R.D. Executive control of visual attention in dual-task situations. Psychol. Rev. 2001, 108, 393–434. [Google Scholar] [CrossRef] [PubMed]
- Wylie, G.; Allport, A. Task switching and the measurement of ‘switch costs’. Psychol. Res. 2000, 63, 212–233. [Google Scholar] [CrossRef] [PubMed]
- Busemeyer, J.R.; Gluth, S.; Rieskamp, J.; Turner, B.M. Cognitive and neural bases of multi-attribute, multi-alternative, value-based decisions. Trends Cogn. Sci. 2019, 23, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Forstmann, B.U.; Wagenmakers, E.-J.; Eichele, T.; Brown, S.; Serences, J.T. Reciprocal relations between cognitive neuroscience and formal cognitive models: Opposites attract? Trends Cogn. Sci. 2011, 15, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Love, B.C. Cognitive models as bridge between brain and behavior. Trends Cogn. Sci. 2016, 20, 247–248. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Turner, B.M.; Wang, T.; Merkle, E.C. Factor analysis linking functions for simultaneously modeling neural and behavioral data. Neuroimage 2017, 153, 28–48. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.M.; Forstmann, B.U.; Wagenmakers, E.J.; Brown, S.D.; Sederberg, P.B.; Steyvers, M. A Bayesian framework for simultaneously modeling neural and behavioral data. Neuroimage 2013, 72, 193–206. [Google Scholar] [CrossRef]
- Turner, B.M.; Palestro, J.J.; Miletić, S.; Forstmann, B.U. Advances in techniques for imposing reciprocity in brain-behavior relations. Neurosci. Biobehav. Rev. 2019, 102, 327–336. [Google Scholar] [CrossRef]
- Möller, M.; Bogacz, R. Learning the payoffs and costs of actions. PLoS Comput. Biol. 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Cools, R. Chemistry of the Adaptive Mind: Lessons from Dopamine. Neuron 2019, 104, 113–131. [Google Scholar] [CrossRef]
- Brown, S.D.; Heathcote, A. The simplest complete model of choice response time: Linear ballistic accumulation. Cogn. Psychol. 2008, 57, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Tillman, G.; Van Zandt, T.; Logan, G.D. Sequential sampling models without random between-trial variability: The racing diffusion model of speeded decision making. Psychon. Bull. Rev. 2020, 27, 911–936. [Google Scholar] [CrossRef]
- Donkin, C.; Brown, S.; Heathcote, A.; Wagenmakers, E.J. Diffusion versus linear ballistic accumulation: Different models but the same conclusions about psychological processes? Psychon. Bull. Rev. 2011, 18, 61–69. [Google Scholar] [CrossRef]
- Heathcote, A.; Hayes, B. Diffusion versus linear ballistic accumulation: Different models for response time with different conclusions about psychological mechanisms? Can. J. Exp. Psychol. 2012, 66, 125–136. [Google Scholar] [CrossRef]
- Turner, B.M.; Sederberg, P.B.; Brown, S.D.; Steyvers, M. A Method for efficiently sampling from distributions with correlated dimensions. Psychol. Methods 2013, 18, 368–384. [Google Scholar] [CrossRef]
- Brooks, S.P.; Gelman, A. General methods for monitoring convergence of iterative simulations. J. Comput. Graph. Stat. 1998, 7, 434–455. [Google Scholar]
- Gelman, A.; Rubin, D.B. Inference from Iterative Simulation Using Multiple Sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]









| Measure | Derivation | Interpretation |
|---|---|---|
| Updating | Difference between no-switch/reference and no-switch/comparison trials | Cost of updating WM |
| Comparison | Difference between no-switch/same (probe-referent match) and no-switch/different (probe-referent mismatch) trials | Cost of a mismatch between the probe stimulus and the WM referent |
| Switching | Difference between switch and no-switch trials | Cost of switching between WM modes |
| Gate opening | Difference between reference/switch and reference/no-switch trials | Cost specific to opening the gate to WM |
| Gate closing | Difference between comparison/switch and comparison/no-switch trials | Cost specific to closing the gate to WM |
| Substitution | Interaction of updating and comparison factors; difference between the cost of updating a new/mismatching item into WM and the cost of responding to a mismatching item without updating; (reference/different − reference/same) − (comparison/different − comparison/same) | Cost of updating a new item into WM |
| Model | Parameters | DIC Difference from Top Model | n | % |
|---|---|---|---|---|
| Top model | 21 | 0 | 56 | 37.3 |
| Threshold fixed | 20 | 179 | 46 | 30.7 |
| Drift rate fixed | 14 | 880 | 35 | 23.3 |
| Non-decision time fixed | 14 | 2720 | 13 | 8.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boag, R.J.; Stevenson, N.; van Dooren, R.; Trutti, A.C.; Sjoerds, Z.; Forstmann, B.U. Cognitive Control of Working Memory: A Model-Based Approach. Brain Sci. 2021, 11, 721. https://doi.org/10.3390/brainsci11060721
Boag RJ, Stevenson N, van Dooren R, Trutti AC, Sjoerds Z, Forstmann BU. Cognitive Control of Working Memory: A Model-Based Approach. Brain Sciences. 2021; 11(6):721. https://doi.org/10.3390/brainsci11060721
Chicago/Turabian StyleBoag, Russell J., Niek Stevenson, Roel van Dooren, Anne C. Trutti, Zsuzsika Sjoerds, and Birte U. Forstmann. 2021. "Cognitive Control of Working Memory: A Model-Based Approach" Brain Sciences 11, no. 6: 721. https://doi.org/10.3390/brainsci11060721
APA StyleBoag, R. J., Stevenson, N., van Dooren, R., Trutti, A. C., Sjoerds, Z., & Forstmann, B. U. (2021). Cognitive Control of Working Memory: A Model-Based Approach. Brain Sciences, 11(6), 721. https://doi.org/10.3390/brainsci11060721