Distinct and Overlapping Patterns of Acute Ethanol-Induced C-Fos Activation in Two Inbred Replicate Lines of Mice Selected for Drinking to High Blood Ethanol Concentrations


 , , ,
, , ,
Abstract
1. Introduction
2. Methods
2.1. Animals
2.2. Immunohistochemistry
2.3. C-Fos Quantification
2.4. Statistics
3. Results
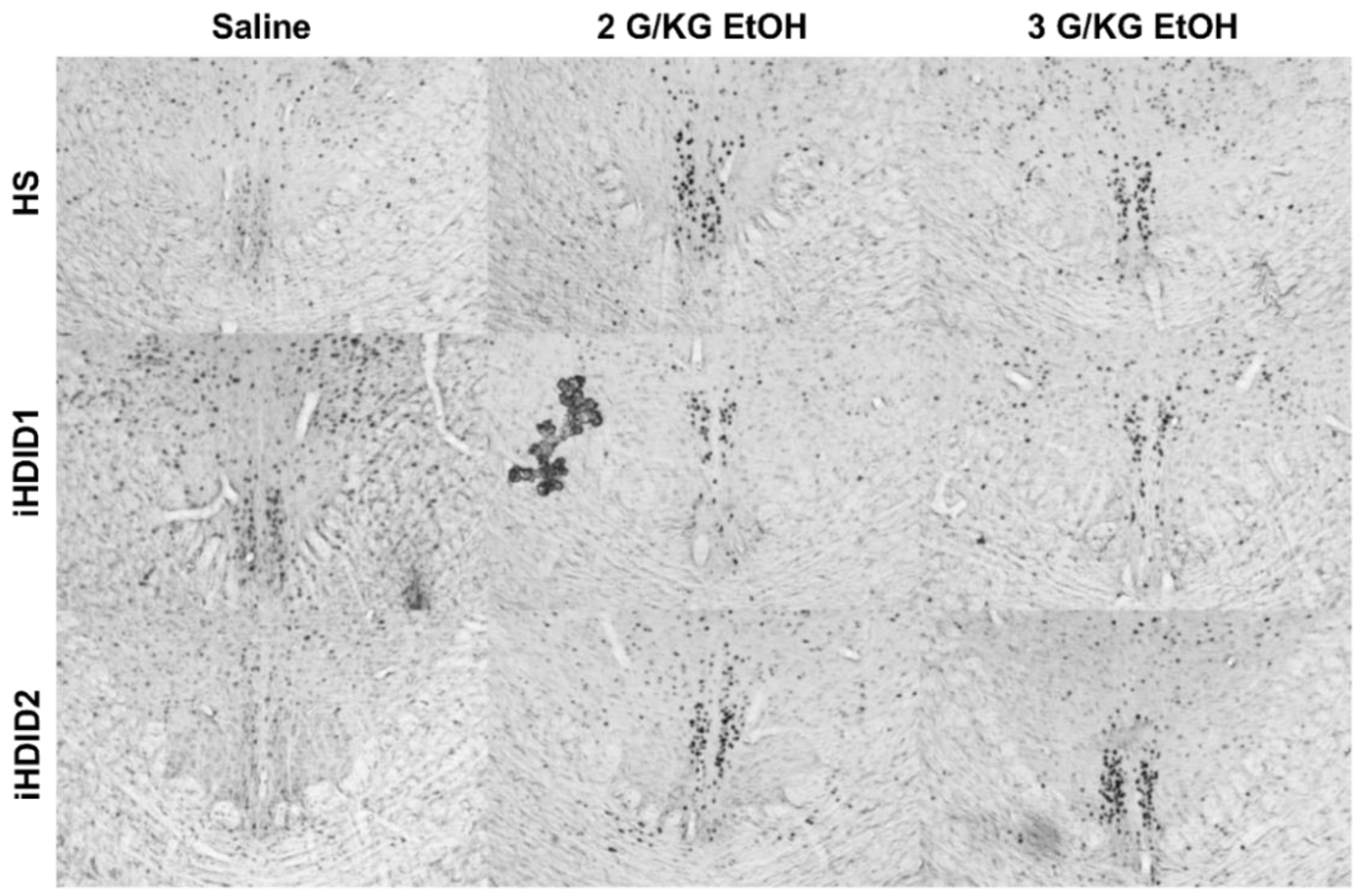
3.1. C-Fos Response in the Hindbrain
3.2. C-Fos Response in the Extended Amygdala
3.3. C-Fos Response in the Thalamic Area
3.4. C-Fos Response in Other Regions Examined
4. Discussion
4.1. Hindbrain Regions
4.2. Extended Amygdala Regions
4.3. Midbrain Regions
4.4. Other Regions Examined
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thiele, T.E.; Navarro, M. “Drinking in the dark” (DID) procedures: A model of binge-like ethanol drinking in non-dependent mice. Alcohol 2014, 48, 235–241. [Google Scholar] [CrossRef]
- Sprow, G.M.; Thiele, T.E. The neurobiology of binge-like ethanol drinking: Evidence from rodent models. Physiol. Behav. 2012, 106, 325–331. [Google Scholar] [CrossRef]
- Barkley-Levenson, A.M.; Crabbe, J.C. High Drinking in the Dark Mice: A genetic model of drinking to intoxication. Alcohol 2014, 48, 217–223. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Metten, P.; Savarese, A.M.; Ozburn, A.R.; Schlumbohm, J.P.; Spence, S.E.; Hack, W.R. Ethanol conditioned taste aversion in high drinking in the dark mice. Brain Sci. 2019, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Barkley-Levenson, A.M.; Crabbe, J.C. Distinct ethanol drinking microstructures in two replicate lines of mice selected for drinking to intoxication. Genes Brain Behav. 2015, 14, 398–410. [Google Scholar] [CrossRef]
- Dan Iancu, O.; Oberbeck, D.; Darakjian, P.; Metten, P.; Mcweeney, S.; Crabbe, J.C.; Hitzemann, R. Selection for Drinking in the Dark Alters Brain Gene Coexpression Networks. Alcohol. Clin. Exp. Res. 2013, 37, 1295–1303. [Google Scholar] [CrossRef]
- Grant, K.A.; Leng, X.; Green, H.L.; Szeliga, K.T.; Rogers, L.S.; Gonzales, S.W. Drinking typography established by scheduled induction predicts chronic heavy drinking in a monkey model of ethanol self-administration. Alcohol. Clin. Exp. Res. 2008, 32, 1824–1838. [Google Scholar] [CrossRef]
- Berridge, K.C. Measuring hedonic impact in animals and infants: Microstructure of affective taste reactivity patterns. Neurosci. Biobehav. Rev. 2000, 24, 173–198. [Google Scholar] [CrossRef]
- Robinson, S.L.; McCool, B.A. Microstructural analysis of rat ethanol and water drinking patterns using a modified operant self-administration model. Physiol. Behav. 2015, 149, 119–130. [Google Scholar] [CrossRef]
- Barkley-Levenson, A.M.; Cunningham, C.L.; Smitasin, P.J.; Crabbe, J.C. Rewarding and aversive effects of ethanol in High Drinking in the Dark selectively bred mice. Addict. Biol. 2015, 20, 80–90. [Google Scholar] [CrossRef]
- Barkley-Levenson, A.M.; Crabbe, J.C. Genotypic and sex differences in anxiety-like behavior and alcohol-induced anxiolysis in High Drinking in the Dark selected mice. Alcohol 2015, 49, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.C.; Lusk, R.; Saba, L.M. Alcohol Sensitivity as an Endophenotype of Alcohol Use Disorder: Exploring Its Translational Utility between Rodents and Humans. Brain Sci. 2020, 10, 725. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, Compact, 3rd ed.; Academic Press Inc.: Cambridge, MA, USA, 2008; ISBN 9780123742445. [Google Scholar]
- De Chaumont, F.; Dallongeville, S.; Chenouard, N.; Hervé, N.; Pop, S.; Provoost, T.; Meas-Yedid, V.; Pankajakshan, P.; Lecomte, T.; le Montagner, Y.; et al. Icy: An open bioimage informatics platform for extended reproducible research. Nat. Methods 2012, 9, 690–696. [Google Scholar] [CrossRef]
- Barkley-Levenson, A.M.; Crabbe, J.C. Ethanol drinking microstructure of a high drinking in the dark selected mouse line. Alcohol. Clin. Exp. Res. 2012, 36, 1330–1339. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Spence, S.E.; Brown, L.L.; Metten, P. Alcohol preference drinking in a mouse line selectively bred for high drinking in the dark. Alcohol 2011, 45, 427–440. [Google Scholar] [CrossRef]
- Bernstein, I.L.; Koh, M.T. Molecular signaling during taste aversion learning. Chem. Senses 2007, 32, 99–103. [Google Scholar] [CrossRef]
- Palmiter, R.D. The Parabrachial Nucleus: CGRP Neurons Function as a General Alarm. Trends Neurosci. 2018, 41, 280–293. [Google Scholar] [CrossRef]
- Smith, R.J.; Aston-Jones, G. Noradrenergic transmission in the extended amygdala: Role in increased drug-seeking and relapse during protracted drug abstinence. Brain Struct. Funct. 2008, 213, 43–61. [Google Scholar] [CrossRef]
- Aimino, M.A.; Coker, C.R.; Silberman, Y. Acute ethanol modulation of neurocircuit function in the nucleus of the tractus solitarius. Brain Res. Bull. 2018, 138, 5–11. [Google Scholar] [CrossRef]
- Burnham, N.W.; Thiele, T.E. Voluntary Binge-like Ethanol Consumption Site-specifically Increases c-Fos Immunoexpression in Male C57BL6/J Mice. Neuroscience 2017, 367, 159–168. [Google Scholar] [CrossRef]
- Thiele, T.E.; Roitman, M.F.; Bernstein, I.L. c-Fos induction in rat brainstem in response to ethanol- and lithium chloride-induced conditioned taste aversions. Alcohol. Clin. Exp. Res. 1996, 20, 1023–1028. [Google Scholar] [CrossRef]
- Lindvall, O.; Bjorklund, A. The organization of the ascending catecholamine neuron systems in the rat brain. As revealed by the glyoxylic acid fluorescence method. Acta Physiol. Scand. 1974, 92, 1–48. [Google Scholar]
- Aston-Jones, G.; Waterhouse, B. Locus coeruleus: From global projection system to adaptive regulation of behavior. Brain Res. 2016, 1645, 75–78. [Google Scholar] [CrossRef]
- Benarroch, E.E. Locus coeruleus. Cell Tissue Res. 2018, 373, 221–232. [Google Scholar] [CrossRef]
- Deal, A.L.; Mikhailova, M.A.; Grinevich, V.P.; Weiner, J.L.; Gainetdinov, R.R.; Budygin, E.A. In vivo voltammetric evidence that locus coeruleus activation predominantly releases norepinephrine in the infralimbic cortex: Effect of acute ethanol. Synapse 2019, 73, e22080. [Google Scholar] [CrossRef]
- Vilpoux, C.; Warnault, V.; Pierrefiche, O.; Daoust, M.; Naassila, M. Ethanol-sensitive brain regions in rat and mouse: A cartographic review, using immediate early gene expression. Alcohol. Clin. Exp. Res. 2009, 33, 945–969. [Google Scholar] [CrossRef]
- Thiele, T.E.; Cubero, I.; van Dijk, G.; Mediavilla, C.; Bernstein, I.L. Ethanol-Induced c-Fos Expression in Catecholamine- and Neuropeptide Y-Producing Neurons in Rat Brainstem. Alcohol. Clin. Exp. Res. 2000, 24, 802–809. [Google Scholar] [CrossRef]
- Hirschberg, S.; Li, Y.; Randall, A.; Kremer, E.J.; Pickering, A.E. Functional dichotomy in spinal- vs prefrontal-projecting locus coeruleus modules splits descending noradrenergic analgesia from ascending aversion and anxiety in rats. eLife 2017, 6, e29808. [Google Scholar] [CrossRef]
- Bonaz, B.; Taché, Y. Water-avoidance stress-induced c-fos expression in the rat brain and stimulation of fecal output: Role of corticotropin-releasing factor. Brain Res. 1994, 641, 21–28. [Google Scholar] [CrossRef]
- Thiele, T.E.; van Dijk, G.; Bernstein, I.L. Ethanol-induced c-Fos expression in rat lines selected for low and high alcohol consumption. Brain Res. 1997, 756, 278–282. [Google Scholar] [CrossRef]
- Canales, J.J. Catalase-independent early-gene expression in rat brain following acute ethanol exposure. Brain Res. 2004, 1016, 96–101. [Google Scholar] [CrossRef]
- Chang, S.L.; Patel, N.A.; Romero, A.A. Activation and desensitization of Fos immunoreactivity in the rat brain following ethanol administration. Brain Res. 1995, 679, 89–98. [Google Scholar] [CrossRef]
- Kolodziejska-Akiyama, K.M.; Young, M.C.; Jiang, Y.; Loh, H.H.; Chang, S.L. Ethanol-induced FOS immunoreactivity in the brain of μ-opioid receptor knockout mice. Drug Alcohol Depend. 2005, 80, 161–168. [Google Scholar] [CrossRef]
- Torruella-Suárez, M.L.; Vandenberg, J.R.; Cogan, E.S.; Tipton, G.J.; Teklezghi, A.; Dange, K.; Patel, G.K.; McHenry, J.A.; Andrew Hardaway, J.; Kantak, P.A.; et al. Manipulations of central amygdala neurotensin neurons alter the consumption of ethanol and sweet fluids in mice. J. Neurosci. 2020, 40, 632–647. [Google Scholar] [CrossRef]
- Kim, J.; Zhang, X.; Muralidhar, S.; LeBlanc, S.A.; Tonegawa, S. Basolateral to Central Amygdala Neural Circuits for Appetitive Behaviors. Neuron 2017, 93, 1464–1479. [Google Scholar] [CrossRef]
- Gilpin, N.W.; Herman, M.A.; Roberto, M. The central amygdala as an integrative hub for anxiety and alcohol use disorders. Biol. Psychiatry 2015, 77, 859–869. [Google Scholar] [CrossRef]
- Roberto, M.; Gilpin, N.W.; Siggins, G.R. The central amygdala and alcohol: Role of γ-aminobutyric acid, glutamate, and neuropeptides. Cold Spring Harb. Perspect. Med. 2012, 2, a012195. [Google Scholar] [CrossRef]
- Roberto, M.; Madamba, S.G.; Stouffer, D.G.; Parsons, L.H.; Siggins, G.R. Increased GABA release in the central amygdala of ethanol-dependent rats. J. Neurosci. 2004, 24, 10159–10166. [Google Scholar] [CrossRef]
- McCool, B.A.; Christian, D.T.; Diaz, M.R.; Läck, A.K. Glutamate plasticity in the drunken amygdala: The making of an anxious synapse. Int. Rev. Neurobiol. 2010, 91, 205–233. [Google Scholar] [CrossRef]
- Rainnie, D.G.; Asprodini, E.K.; Shinnick-Gallagher, P. Excitatory transmission in the basolateral amygdala. J. Neurophysiol. 1991, 66, 986–998. [Google Scholar] [CrossRef]
- Ch’ng, S.; Fu, J.; Brown, R.M.; McDougall, S.J.; Lawrence, A.J. The intersection of stress and reward: BNST modulation of aversive and appetitive states. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 87, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Sharko, A.C.; Kaigler, K.F.; Fadel, J.R.; Wilson, M.A. Ethanol-induced anxiolysis and neuronal activation in the amygdala and bed nucleus of the stria terminalis. Alcohol 2016, 50, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Saalfield, J.; Spear, L. Fos activation patterns related to acute ethanol and conditioned taste aversion in adolescent and adult rats. Alcohol 2019, 78, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Ryabinin, A.E.; Criado, J.R.; Henriksen, S.J.; Bloom, F.E.; Wilson, M.C. Differential sensitivity of c-Fos expression in hippocampus and other brain regions to moderate and low doses of alcohol. Mol. Psychiatry 1997, 2, 32–43. [Google Scholar] [CrossRef]
- Crankshaw, D.L.; Briggs, J.E.; Olszewski, P.K.; Shi, Q.; Grace, M.K.; Billington, C.J.; Levine, A.S. Effects of intracerebroventricular ethanol on ingestive behavior and induction of c-Fos immunoreactivity in selected brain regions. Physiol. Behav. 2003, 79, 113–120. [Google Scholar] [CrossRef]
- Schreiber, A.L.; McGinn, M.A.; Edwards, S.; Gilpin, N.W. Predator odor stress blunts alcohol conditioned aversion. Neuropharmacology 2019, 144, 82–90. [Google Scholar] [CrossRef]
- Campbell, R.R.; Domingo, R.D.; Williams, A.R.; Wroten, M.G.; McGregor, H.A.; Waltermire, R.S.; Greentree, D.I.; Goulding, S.P.; Thompson, A.B.; Lee, K.M.; et al. Increased alcohol-drinking induced by manipulations of mGlu5 phosphorylation within the bed nucleus of the stria terminalis. J. Neurosci. 2019, 39, 2745–2761. [Google Scholar] [CrossRef]
- Centanni, S.W.; Bedse, G.; Patel, S.; Winder, D.G. Driving the Downward Spiral: Alcohol-Induced Dysregulation of Extended Amygdala Circuits and Negative Affect HHS Public Access. Alcohol. Clin. Exp. Res. 2019, 43, 2000–2013. [Google Scholar] [CrossRef]
- Robinson, S.L.; Thiele, T.E. The Role of Neuropeptide Y (NPY) in Alcohol and Drug Abuse Disorders. Int. Rev. Neurobiol. 2017, 136, 177–197. [Google Scholar] [CrossRef]
- Barkley-Levenson, A.M.; Ryabinin, A.E.; Crabbe, J.C. Neuropeptide Y response to alcohol is altered in nucleus accumbens of mice selectively bred for drinking to intoxication. Behav. Brain Res. 2016, 302, 160–170. [Google Scholar] [CrossRef]
- Dhaher, R.; Finn, D.; Snelling, C.; Hitzemann, R. Lesions of the extended amygdala in C57BL/6J mice do not block the intermittent ethanol vapor-induced increase in ethanol consumption. Alcohol. Clin. Exp. Res. 2008, 32, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Rinker, J.A.; Marshall, S.A.; Mazzone, C.M.; Lowery-Gionta, E.G.; Gulati, V.; Pleil, K.E.; Kash, T.L.; Navarro, M.; Thiele, T.E. Extended Amygdala to Ventral Tegmental Area Corticotropin-Releasing Factor Circuit Controls Binge Ethanol Intake. Biol. Psychiatry 2017, 81, 930–940. [Google Scholar] [CrossRef] [PubMed]
- Companion, M.A.; Thiele, T.E. Assessment of ventral tegmental area-projecting GABAergic neurons from the bed nucleus of the stria terminalis in modulating binge-like ethanol intake. Eur. J. Neurosci. 2018, 48, 3335–3343. [Google Scholar] [CrossRef] [PubMed]
- Tandon, S.; Keefe, K.A.; Taha, S.A. Excitation of lateral habenula neurons as a neural mechanism underlying ethanol-induced conditioned taste aversion. J. Physiol. 2017, 595, 1393–1412. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Zuo, W.; Kang, S.; Li, J.; Fu, R.; Zhang, H.; Bekker, A.; Ye, J.H. The lateral habenula and alcohol: Role of glutamate and M-type potassium channels. Pharmacol. Biochem. Behav. 2017, 162, 94–102. [Google Scholar] [CrossRef]
- Haack, A.K.; Sheth, C.; Schwager, A.L.; Sinclair, M.S.; Tandon, S.; Taha, S.A. Lesions of the Lateral Habenula Increase Voluntary Ethanol Consumption and Operant Self-Administration, Block Yohimbine-Induced Reinstatement of Ethanol Seeking, and Attenuate Ethanol-Induced Conditioned Taste Aversion. PLoS ONE 2014, 9, e92701. [Google Scholar] [CrossRef]
- Li, J.; Zuo, W.; Fu, R.; Xie, G.; Kaur, A.; Bekker, A.; Hong Ye, J.; Ye, J.-H. High Frequency Electrical Stimulation of Lateral Habenula Reduces Voluntary Ethanol Consumption in Rats. Int. J. Neuropsychopharmacol. 2016, 19, pyw050. [Google Scholar] [CrossRef]
- Fu, R.; Mei, Q.; Zuo, W.; Li, J.; Gregor, D.; Bekker, A.; Ye, J. Low-dose ethanol excites lateral habenula neurons projecting to VTA, RMTg, and raphe. Int. J. Physiol. Pathophysiol. Pharmacol. 2017, 9, 217–230. [Google Scholar]
- McLaughlin, I.; Dani, J.A.; de Biasi, M. The medial habenula and interpeduncular nucleus circuitry is critical in addiction, anxiety, and mood regulation. J. Neurochem. 2017, 142, 130–143. [Google Scholar] [CrossRef]
- Sutherland, R.J. The dorsal diencephalic conduction system: A review of the anatomy and functions of the habenular complex. Neurosci. Biobehav. Rev. 1982, 6, 1–13. [Google Scholar] [CrossRef]
- Hwang, B.H.; Suzuki, R.; Lumeng, L.; Li, T.K.; McBride, W.J. Innate differences in neuropeptide Y (NPY) mRNA expression in discrete brain regions between alcohol-preferring (P) and -nonpreferring (NP) rats: A significantly low level of NPY mRNA in dentate gyrus of the hippocampus and absence of NPY mRNA in the medial habenular nucleus of P rats. Neuropeptides 2004, 38, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.G.; Learn, J.E.; McBride, W.J.; Lumeng, L.; Li, T.K.; Murphy, J.M. Alcohol-naïve alcohol-preferring (P) rats exhibit higher local cerebral glucose utilization than alcohol-nonpreferring (NP) and wistar rats. Alcohol. Clin. Exp. Res. 2001, 25, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.W.A.; Wang, S.D.; Wang, S.; Morton, G.; Zariwala, H.A.; de la Iglesia, H.O.; Turner, E.E. Role of the dorsal medial habenula in the regulation of voluntary activity, motor function, hedonic state, and primary reinforcement. J. Neurosci. 2014, 34, 11366–11384. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sun, Y.; Cai, X.; You, T.; Zhao, H.; Li, Y.; Zhao, H. Medial habenula-interpeduncular nucleus circuit contributes to anhedonia-like behavior in a rat model of depression. Front. Behav. Neurosci. 2018, 12, 238. [Google Scholar] [CrossRef] [PubMed]
- Bourdy, R.; Barrot, M. A new control center for dopaminergic systems: Pulling the VTA by the tail. Trends Neurosci. 2012, 35, 681–690. [Google Scholar] [CrossRef]
- Sheth, C.; Furlong, T.M.; Keefe, K.A.; Taha, S.A. Lesion of the rostromedial tegmental nucleus increases voluntary ethanol consumption and accelerates extinction of ethanol-induced conditioned taste aversion. Psychopharmacology 2016, 233, 3737–3749. [Google Scholar] [CrossRef]
- Glover, E.J.; McDougle, M.J.; Siegel, G.S.; Jhou, T.C.; Chandler, L.J. Role for the Rostromedial Tegmental Nucleus in Signaling the Aversive Properties of Alcohol. Alcohol. Clin. Exp. Res. 2016, 40, 1651–1661. [Google Scholar] [CrossRef]
- Fu, R.; Chen, X.; Zuo, W.; Li, J.; Kang, S.; Zhou, L.H.; Siegel, A.; Bekker, A.; Ye, J.H. Ablation of μ opioid receptor-expressing GABA neurons in rostromedial tegmental nucleus increases ethanol consumption and regulates ethanol-related behaviors. Neuropharmacology 2016, 107, 58–67. [Google Scholar] [CrossRef]
- Ye, J.H.; Fu, R.; He, W. The rostromedial tegmental nucleus and alcohol addiction. Oncotarget 2017, 8, 18624–18625. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Ozburn, A.R.; Metten, P.; Barkley-Levenson, A.; Schlumbohm, J.P.; Spence, S.E.; Hack, W.R.; Huang, L.C. High Drinking in the Dark (HDID) mice are sensitive to the effects of some clinically relevant drugs to reduce binge-like drinking. Pharmacol. Biochem. Behav. 2017, 160, 55–62. [Google Scholar] [CrossRef]
- Li, C.; McCall, N.M.; Lopez, A.J.; Kash, T.L. Alcohol effects on synaptic transmission in periaqueductal gray dopamine neurons. Alcohol 2013, 47, 279–287. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mcclintick, J.N.; Mcbride, W.J.; Bell, R.L.; Ding, Z.M.; Liu, Y.; Xuei, X.; Edenberg, H.J. Gene Expression Changes in Glutamate and GABA-A Receptors, Neuropeptides, Ion Channels, and Cholesterol Synthesis in the Periaqueductal Gray Following Binge-Like Alcohol Drinking by Adolescent Alcohol-Preferring (P) Rats. Alcohol. Clin. Exp. Res. 2016, 40, 955–968. [Google Scholar] [CrossRef]
- Motta, S.C.; Carobrez, A.P.; Canteras, N.S. The periaqueductal gray and primal emotional processing critical to influence complex defensive responses, fear learning and reward seeking. Neurosci. Biobehav. Rev. 2017, 76, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Topple, A.N.; Hunt, G.E.; McGregor, I.S. Possible neural substrates of beer-craving in rats. Neurosci. Lett. 1998, 252, 99–102. [Google Scholar] [CrossRef]
- Turek, V.F.; Ryabinin, A.E. Expression of c-Fos in the mouse Edinger-Westphal nucleus following ethanol administration is not secondary to hypothermia or stress. Brain Res. 2005, 1063, 132–139. [Google Scholar] [CrossRef]
- Zuniga, A.; Ryabinin, A.E. Involvement of centrally projecting edinger-westphal nucleus neuropeptides in actions of addictive drugs. Brain Sci. 2020, 10, 67. [Google Scholar] [CrossRef]
- Ryabinin, A.E.; Weitemier, A.Z. The urocortin 1 neurocircuit: Ethanol-sensitivity and potential involvement in alcohol consumption. Brain Res. Rev. 2006, 52, 368–380. [Google Scholar] [CrossRef]
- Ryabinin, A.E.; Galvan-Rosas, A.; Bachtell, R.K.; Risinger, F.O. High alcohol/sucrose consumption during dark circadian phase in C57Bl/6J mice: Involvement of hippocampus, lateral septum and urocortin-positive cells of the Edinger-Westphal nucleus. Psychopharmacology 2003, 165, 296–305. [Google Scholar] [CrossRef]
- Martin-Fardon, R.; Boutrel, B. Orexin/hypocretin (Orx/Hcrt) transmission and drug-seeking behavior: Is the paraventricular nucleus of the thalamus (PVT) part of the drug seeking circuitry? Front. Behav. Neurosci. 2012, 6, 75. [Google Scholar] [CrossRef]
- Matzeu, A.; Zamora-Martinez, E.R.; Martin-Fardon, R. The paraventricular nucleus of the thalamus is recruited by both natural rewards and drugs of abuse: Recent evidence of a pivotal role for orexin/hypocretin signaling in this thalamic nucleus in drug-seeking behavior. Front. Behav. Neurosci. 2014, 8, 117. [Google Scholar] [CrossRef]
- James, M.H.; Dayas, C.V. What about me…? The PVT: A role for the paraventricular thalamus (PVT) in drug-seeking behavior. Front. Behav. Neurosci. 2013, 7, 18. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Otis, J.M.; Namboodiri, V.M.K.; Matan, A.M.; Voets, E.S.; Mohorn, E.P.; Kosyk, O.; McHenry, J.A.; Robinson, J.E.; Resendez, S.L.; Rossi, M.A.; et al. Prefrontal cortex output circuits guide reward seeking through divergent cue encoding. Nature 2017, 543, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; van den Pol, A.N. Rapid binge-like eating and body weight gain driven by zona incerta GABA neuron activation. Science 2017, 356, 853–859. [Google Scholar] [CrossRef]
- Otis, J.M.; Zhu, M.H.; Namboodiri, V.M.K.; Cook, C.A.; Kosyk, O.; Matan, A.M.; Ying, R.; Hashikawa, Y.; Hashikawa, K.; Trujillo-Pisanty, I.; et al. Paraventricular Thalamus Projection Neurons Integrate Cortical and Hypothalamic Signals for Cue-Reward Processing. Neuron 2019, 103, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Marchant, N.J.; Furlong, T.M.; McNally, G.P. Medial dorsal hypothalamus mediates the inhibition of reward seeking after extinction. J. Neurosci. 2010, 30, 14102–14115. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, A.S.; Clemens, K.J.; Choi, E.A.; McNally, G.P. Paraventricular thalamus mediates context-induced reinstatement (renewal) of extinguished reward seeking. Eur. J. Neurosci. 2009, 29, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Do-Monte, F.H.; Quinõnes-Laracuente, K.; Quirk, G.J. A temporal shift in the circuits mediating retrieval of fear memory. Nature 2015, 519, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Schuckit, M.A. Biological markers in alcoholism. Prog. Neuropsychopharmacol. Biol. Psychiatry 1986, 10, 191–199. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Genotype | VEH c-Fos vs. HS VEH | 2 G/KG EtOH c-Fos vs. VEH | 3 G/KG EtOH c-Fos vs. VEH |
|---|---|---|---|---|
| A2 | HS | ↑ *** | ↑ **** | |
| iHDID1 | - | ↑ * | - | |
| iHDID2 | - | ↑ **** | ↑ **** | |
| LC | HS | - | ↑ * | |
| iHDID1 | - | - | - | |
| iHDID2 | - | ↑ ** | ↑ ** | |
| PBN | HS | ↑ ** | ↑ * | |
| iHDID1 | - | ↑ * | ↑ ** | |
| iHDID2 | - | ↑ ** | ↑ *** | |
| BLA | HS | - | - | |
| iHDID1 | - | - | - | |
| iHDID2 | - | - | - | |
| CeA | HS | ↑ **** | ↑ **** | |
| iHDID1 | - | ↑ **** | ↑ **** | |
| iHDID2 | - | ↑ *** | ↑ **** | |
| BNST | HS | ↑ * | - | |
| iHDID1 | - | - | - | |
| iHDID2 | - | ↑ ** | ↑ * | |
| LHb | HS | - | - | |
| iHDID1 | - | - | - | |
| iHDID2 | - | - | - | |
| MHb | HS | - | - | |
| iHDID1 | - | - | - | |
| iHDID2 | ↑ ** | - | - | |
| PVT | HS | ↑ * | ↑ * | |
| iHDID1 | - | ↑ ** | ↑ **** | |
| iHDID2 | ↑ * | - | - | |
| PAG | HS | - | - | |
| iHDID1 | - | - | - | |
| iHDID2 | - | - | - | |
| EW | HS | - | ↑ **** | |
| iHDID1 | ↑ ** | - | - | |
| iHDID2 | - | ↑ *** | ↑ *** | |
| RMTg | HS | - | - | |
| iHDID1 | ↓ * | - | - | |
| iHDID2 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, S.L.; Dornellas, A.P.S.; Burnham, N.W.; Houck, C.A.; Luhn, K.L.; Bendrath, S.C.; Companion, M.A.; Brewton, H.W.; Thomas, R.D.; Navarro, M.; et al. Distinct and Overlapping Patterns of Acute Ethanol-Induced C-Fos Activation in Two Inbred Replicate Lines of Mice Selected for Drinking to High Blood Ethanol Concentrations. Brain Sci. 2020, 10, 988. https://doi.org/10.3390/brainsci10120988
Robinson SL, Dornellas APS, Burnham NW, Houck CA, Luhn KL, Bendrath SC, Companion MA, Brewton HW, Thomas RD, Navarro M, et al. Distinct and Overlapping Patterns of Acute Ethanol-Induced C-Fos Activation in Two Inbred Replicate Lines of Mice Selected for Drinking to High Blood Ethanol Concentrations. Brain Sciences. 2020; 10(12):988. https://doi.org/10.3390/brainsci10120988
Chicago/Turabian StyleRobinson, Stacey L., Ana Paula S. Dornellas, Nathan W. Burnham, Christa A. Houck, Kendall L. Luhn, Sophie C. Bendrath, Michel A. Companion, Honoreé W. Brewton, Rhiannon D. Thomas, Montserrat Navarro, and et al. 2020. "Distinct and Overlapping Patterns of Acute Ethanol-Induced C-Fos Activation in Two Inbred Replicate Lines of Mice Selected for Drinking to High Blood Ethanol Concentrations" Brain Sciences 10, no. 12: 988. https://doi.org/10.3390/brainsci10120988
APA StyleRobinson, S. L., Dornellas, A. P. S., Burnham, N. W., Houck, C. A., Luhn, K. L., Bendrath, S. C., Companion, M. A., Brewton, H. W., Thomas, R. D., Navarro, M., & Thiele, T. E. (2020). Distinct and Overlapping Patterns of Acute Ethanol-Induced C-Fos Activation in Two Inbred Replicate Lines of Mice Selected for Drinking to High Blood Ethanol Concentrations. Brain Sciences, 10(12), 988. https://doi.org/10.3390/brainsci10120988