


The Quirky Rot Fungi: Underexploited Potential for Soil Remediation and Rehabilitation


{kind=link}
{kind=link}
{kind=link}
Abstract
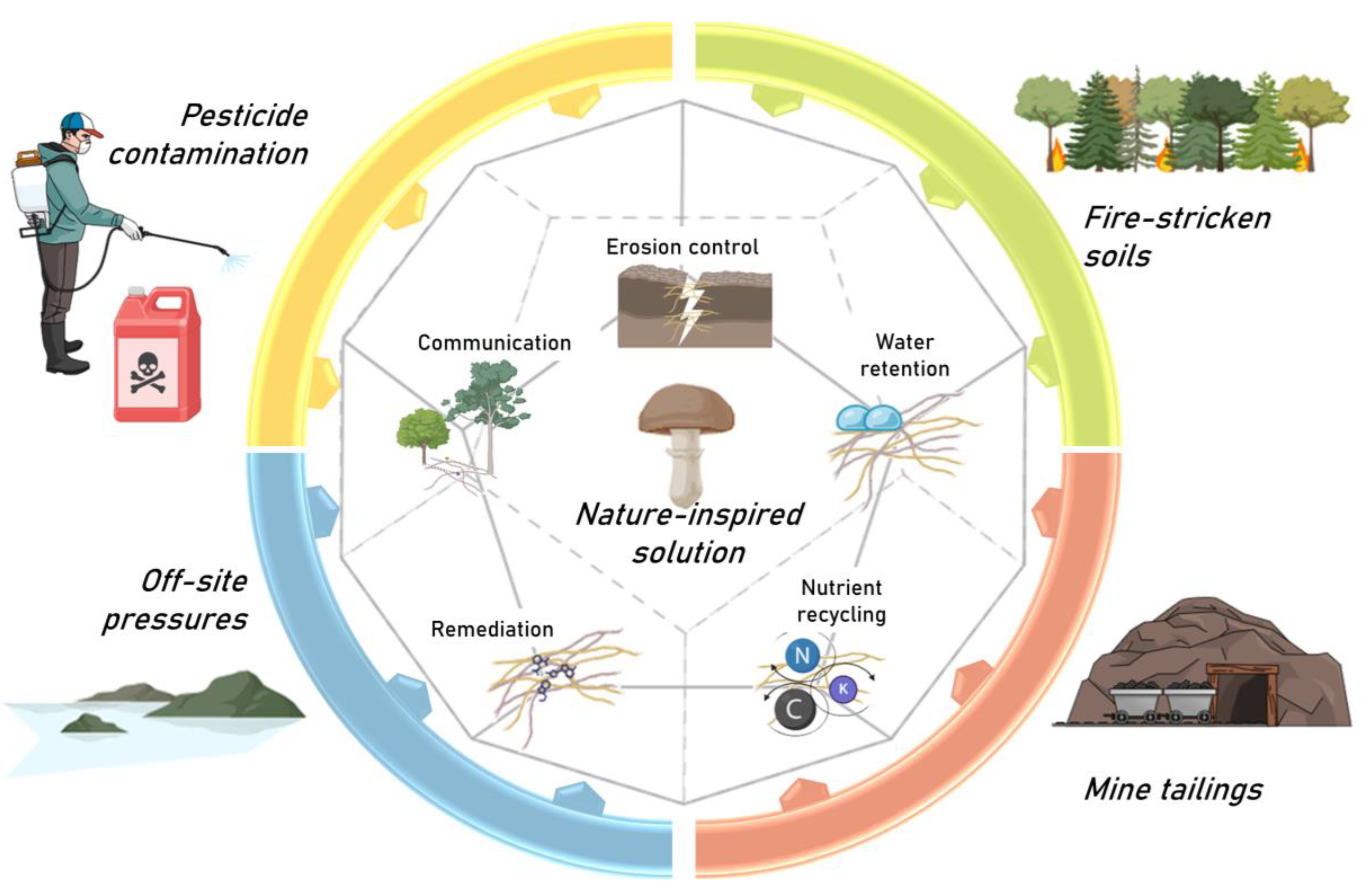
1. Soil Ecology, Health, and Erosion Control
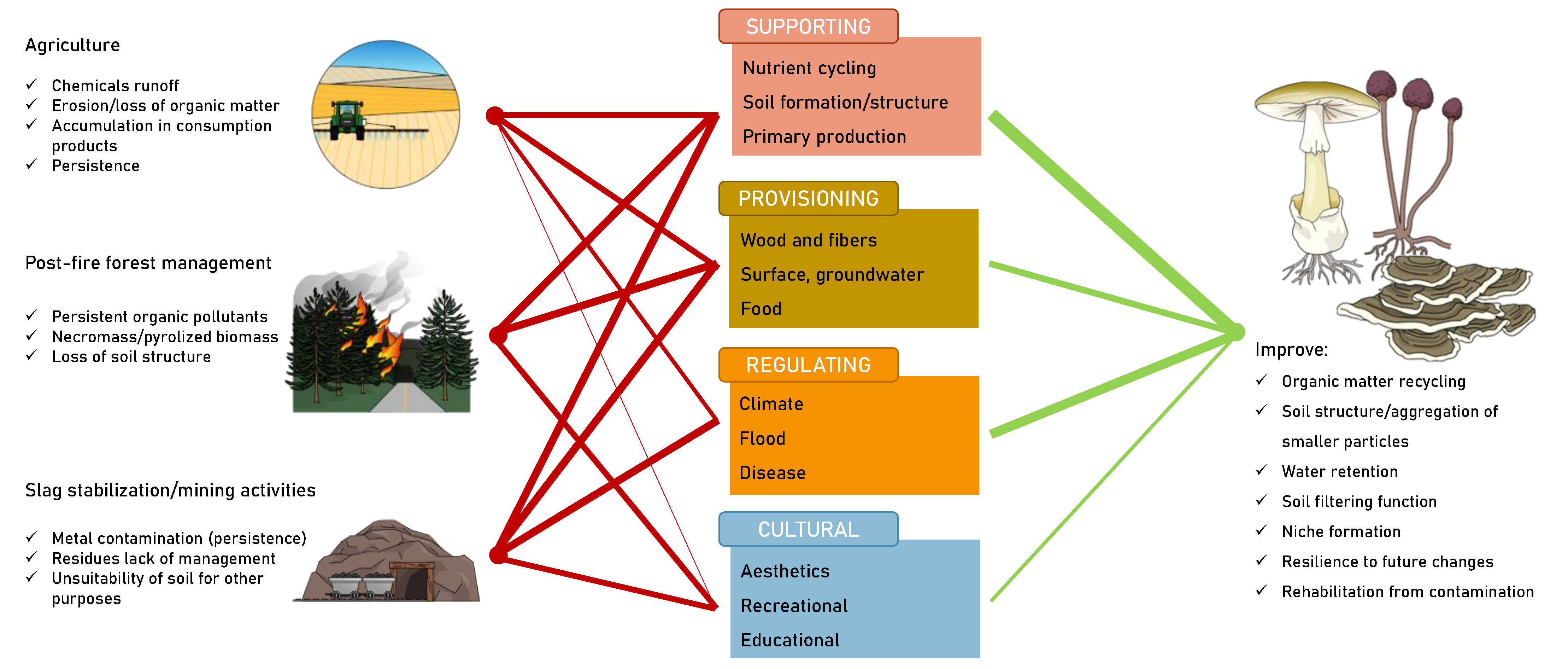
2. Agricultural Sustainability, Post-Fire Forest Management, and Ore-Affected Soils: Shared Challenges with Common Goals
2.1. Pesticides
2.2. Slag Stabilization and Mining-Affected Soil
2.3. Fire-Stricken Soils
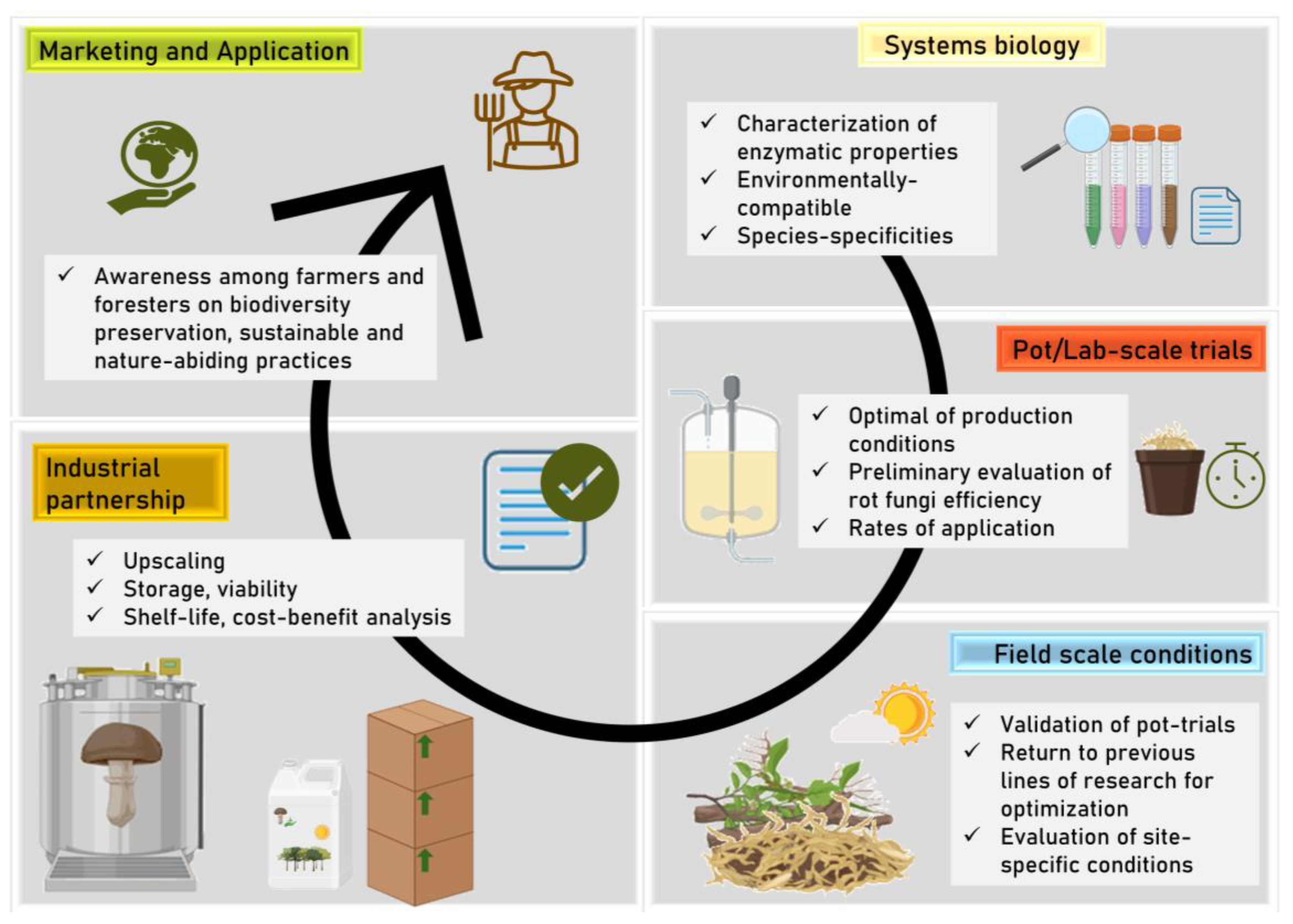
3. Drawbacks and Weaknesses
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goodell, B. Fungi involved in the biodeterioration and bioconversion of lignocellulose substrates. Genet. Bio-Technol. 2020, 2, 369–397. [Google Scholar] [CrossRef]
- Kang, P.; Pan, Y.; Ran, Y.; Li, W.; Shao, M.; Zhang, Y.; Ji, Q.; Ding, X. Soil saprophytic fungi could be used as an important ecological indicator for land management in desert steppe. Ecol. Indic. 2023, 150, 110224. [Google Scholar] [CrossRef]
- Li, J.; Zhang, T.; Meng, B.; Rudgers, J.A.; Cui, N.; Zhao, T.; Chai, H.; Yang, X.; Sternberg, M.; Sun, W. Disruption of fungal hyphae suppressed litter-derived C retention in soil and N translocation to plants under drought-stressed temperate grassland. Geoderma 2023, 432, 116396. [Google Scholar] [CrossRef]
- Ritz, K.; Young, I.M. Interactions between soil structure and fungi. Mycologist 2004, 18, 52–59. [Google Scholar] [CrossRef]
- Tláskal, V.; Brabcová, V.; Větrovský, T.; Jomura, M.; López-Mondéjar, R.; Oliveira Monteiro, L.M.; Saraiva, J.P.; Human, Z.R.; Cajthaml, T.; Nunes da Rocha, U.; et al. Complementary roles of wood-inhabiting fungi and bacteria facilitate deadwood decomposition. Msystems 2021, 6, 10–1128. [Google Scholar] [CrossRef]
- Arellano-Caicedo, C.; Ohlsson, P.; Bengtsson, M.; Beech, J.P.; Hammer, E.C. Habitat geometry in artificial microstructure affects bacterial and fungal growth, interactions, and substrate degradation. Commun. Biol. 2021, 4, 1226. [Google Scholar] [CrossRef]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions: Remaping the Benefits of Healthy Soils for People, Food, Nature and Climate. EU Soil Strategy for 2030. 2021. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52021DC0699 (accessed on 16 January 2025).
- United Nations. The Sustainable Development Goals Report. 2021. Available online: https://unstats.un.org/sdgs/report/2021/The-Sustainable-Development-Goals-Report-2021.pdf (accessed on 16 January 2025).
- Santos, I.; Ferreira da Silva, E.; Patinha, C.; Dias, A.C.; Reis, A.P. Definition of areas of probable risk to human health posed by As and Pb in soils and ground-level dusts of the surrounding area of an abandoned As-sulfide mine in the north of Portugal: Part 1. Environ. Earth Sci. 2013, 69, 1649–1660. [Google Scholar] [CrossRef]
- López, R.; Hallat, J.; Castro, A.; Miras, A.; Burgos, P. Heavy metal pollution in soils and urban-grown organic vegetables in the province of Sevilla, Spain. Biol. Agric. Hortic. 2019, 35, 219–237. [Google Scholar] [CrossRef]
- Reis, M.M.; Tuffi Santos, L.D.; da Silva, A.J.; de Pinho, G.P.; Montes, W.G. Metal contamination of water and sediments of the Vieira River, Montes Claros, Brazil. Arch. Environ. Contam. Toxicol. 2019, 77, 527–536. [Google Scholar] [CrossRef]
- Verma, S.; Jayakumar, S. Impact of forest fire on physical, chemical and biological properties of soil: A review. Proc. Int. Acad. Ecol. Environ. Sci. 2012, 2, 168. [Google Scholar]
- Wang, S.; Li, W.; Liu, L.; Qi, H.; You, H. Biodegradation of decabromodiphenyl ethane (DBDPE) by white-rot fungus Pleurotus ostreatus: Characteristics, mechanisms, and toxicological response. J. Hazard. Mater. 2022, 424, 127716. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Communication from the Commission to the European Parliament, The Council, The Europea Economic and Social Committee and the Committee of the Regions Managing Climate Risks—Protecting People and Prosperity. Strasbourg, France. 2024. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52024DC0091 (accessed on 16 January 2025).
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Wu, M.; Xu, Y.; Ding, W.; Li, Y.; Xu, H. Mycoremediation of manganese and phenanthrene by Pleurotus eryngii mycelium enhanced by Tween 80 and saponin. Appl. Microbiol. Biotechnol. 2016, 100, 7249–7261. [Google Scholar] [CrossRef]
- Harms, H.; Wick, L.Y.; Schlosser, D. The fungal community in organically polluted systems. In Mycology; Dighton, J., White, J.F.Y., Eds.; CRC Press-Taylor & Francis Group: Boca Raton, FL, USA, 2017; pp. 459–470. [Google Scholar]
- Ceci, A.; Pinzari, F.; Russo, F.; Persiani, A.M.; Gadd, G.M. Roles of saprotrophic fungi in biodegradation or transformation of organic and inorganic pollutants in co-contaminated sites. Appl. Microbiol. Biotechnol. 2019, 103, 53–68. [Google Scholar] [CrossRef]
- Miyauchi, S.; Navarro, D.; Grisel, S.; Chevret, D.; Berrin, J.G.; Rosso, M.N. The integrative omics of white-rot fungus Pycnoporus coccineus reveals co-regulated CAZymes for orchestrated lignocellulose breakdown. PLoS ONE 2017, 12, e0175528. [Google Scholar] [CrossRef]
- Venâncio, C.; Pereira, R.; Freitas, A.C.; Rocha-Santos, T.A.P.; da Costa, J.P.; Duarte, A.C.; Lopes, I. Salinity induced effects on the growth rates and mycelia composition of basidiomycete and zygomycete fungi. Environ. Pollut. 2017, 231, 1633–1641. [Google Scholar] [CrossRef]
- Borges, J.; Cardoso, P.; Lopes, I.; Figueira, E.; Venâncio, C. Exploring the Potential of White-Rot Fungi Exudates on the Amelioration of Salinized Soils. Agriculture 2023, 13, 382. [Google Scholar] [CrossRef]
- Verce, M.F.; Ulrich, R.L.; Freedman, D.L. Characterization of an isolate that uses vinyl chloride as a growth substrate under aerobic conditions. Appl. Environ. Microbiol. 2000, 66, 3535–3542. [Google Scholar] [CrossRef]
- Feijao, C.; d’Angelo, C.; Flanagan, I.; Abellan, B.; Gloinson, E.R.; Smith, E.; Traon, D.; Gehrt, D.; Teare, H.; Dunkerley, F. Directorate General for Health and Food Safety. In Development of Future Scenarios for Sustainable Pesticide Use and Achievement of Pesticide-Use and Risk Reduction Targets Announced in the Farm to Fork and Biodiversity Strategies by 2030; European Commission: Brussels, Belgium; Luxembourg, 2022; p. 135. [Google Scholar]
- Cherlet, M.; Hutchinson, C.; Reynolds, J.; Sommer, H.; Graham, V.M. World Atlas of Desertification, 3rd ed.; Publications Office of the European Union: Luxembourg, 2018; p. 248. [Google Scholar]
- Silva, V.; Mol, H.G.; Zomer, P.; Tienstra, M.; Ritsema, C.J.; Geissen, V. Pesticide residues in European agricultural soils–A hidden reality unfolded. Sci. Total Environ. 2019, 653, 1532–1545. [Google Scholar] [CrossRef]
- Adenipekun, C.O.; Lawal, R. Uses of mushrooms in bioremediation: A review. Biotechnol. Mol. Biol. Rev. 2012, 7, 62–68. [Google Scholar] [CrossRef]
- Bumpus, J.A. White rot fungi and their potential use in soil bioremediation processes. In Soil Biochemistry; Bollag, J.M., Stotzky, G., Eds.; Marcel Dekker: New York, NY, USA, 1993; pp. 65–100. [Google Scholar]
- Venâncio, C.; Cardoso, P.; Ekner-Grzyb, A.; Chmielowska-Bąk, J.; Grzyb, T.; Lopes, I. Sources, sinks, and solutions: How decaying fungi may devise sustainable farming practices for plastics degradation in terrestrial ecosystems. TrAC Trends Anal. Chem. 2024, 180, 117898. [Google Scholar] [CrossRef]
- Kijpornyongpan, T.; Schwartz, A.; Yaguchi, A.; Salvachúa, D. Systems biology-guided understanding of white-rot fungi for biotechnological applications: A review. Iscience 2022, 25, 104640. [Google Scholar] [CrossRef] [PubMed]
- Mileski, G.J.; Bumpus, J.A.; Jurek, M.A.; Aust, S.D. Biodegradation of pentachlorophenol by the white rot fungus Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1988, 54, 2885–2889. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Xue, C.; Chen, A.; Shang, C.; Luo, S. Phanerochaete chrysosporium-driven quinone redox cycling promotes degradation of imidacloprid. Int. Biodeterior. Biodegrad. 2020, 151, 104965. [Google Scholar] [CrossRef]
- Abo-State, M.A.M.; Osman, M.E.; Khattab, O.H.; El-Kelani, T.A.; Abdel-Rahman, Z.M. Degradative pathways of polycyclic aromatic hydrocarbons (PAHs) by Phanerochaete chrysosporium under optimum conditions. J. Radiat. Res. Appl. Sci. 2021, 14, 507–520. [Google Scholar] [CrossRef]
- Adenipekun, C.O.; Fasidi, I.O. Bioremediation of oil-polluted soil by Lentinus subnudus, a Nigerian white-rot fungus. Afr. J. Biotechnol. 2005, 4, 796–798. [Google Scholar]
- Okparanma, R.N.; Ayotamuno, J.M.; Davis, D.D.; Allagoa, M. Mycoremediation of polycyclic aromatic hydrocarbons (PAH)-contaminated oil-based drill-cuttings. Afr. J. Biotechnol. 2011, 10, 5149–5156. [Google Scholar]
- Liu, G.; Shi, Y.; Guo, G.; Zhao, L.; Niu, J.; Zhang, C. Soil pollution characteristics and systemic environmental risk assessment of a large-scale arsenic slag contaminated site. J. Clean. Prod. 2020, 251, 119721. [Google Scholar] [CrossRef]
- Santana, C.S.; Olivares, D.M.M.; Silva, V.H.; Luzardo, F.H.; Velasco, F.G.; de Jesus, R.M. Assessment of water resources pollution associated with mining activity in a semi-arid region. J. Environ. Manag. 2020, 273, 111148. [Google Scholar] [CrossRef]
- Brotons, J.M.; Díaz, A.R.; Sarría, F.A.; Serrato, F.B. Wind erosion on mining waste in southeast Spain. Land Degrad. Dev. 2010, 21, 196–209. [Google Scholar] [CrossRef]
- Sun, C.; Wu, P.; Wang, G.; Kong, X. Improvement of plant diversity along the slope of an historical Pb–Zn slag heap ameliorates the negative effect of heavy metal on microbial communities. Plant Soil 2022, 473, 473–487. [Google Scholar] [CrossRef]
- Rajapaksha, R.M.C.P.; Tobor-Kapłon, M.A.; Baath, E. Metal toxicity affects fungal and bacterial activities in soil differently. Appl. Environ. Microbiol. 2004, 70, 2966–2973. [Google Scholar] [CrossRef] [PubMed]
- Xiong, B.J.; Kleinsteuber, S.; Sträuber, H.; Dusny, C.; Harms, H.; Wick, L.Y. Impact of fungal hyphae on growth and dispersal of obligate anaerobic bacteria in aerated habitats. Mbio 2022, 13, e00769-22. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Bi, R.; Guo, X.; Chen, D.; Guo, Y.; Xu, Z. Erosion characteristics of different reclaimed substrates on iron tailings slopes under simulated rainfall. Sci. Rep. 2020, 10, 4275. [Google Scholar] [CrossRef]
- Udom, B.E.; Nuga, B.O.; Adesodun, J.K. Water-stable aggregates and aggregate-associated organic carbon and nitrogen after three annual applications of poultry manure and spent mushroom wastes. Appl. Soil Ecol. 2016, 101, 5–10. [Google Scholar] [CrossRef]
- Shakya, M.; Sharma, P.; Meryem, S.S.; Mahmood, Q.; Kumar, A. Heavy metal removal from industrial wastewater using fungi: Uptake mechanism and biochemical aspects. J. Environ. Eng. 2016, 142, C6015001. [Google Scholar] [CrossRef]
- Ceci, A.; Spinelli, V.; Massimi, L.; Canepari, S.; Persiani, A.M. Fungi and arsenic: Tolerance and bioaccumulation by soil saprotrophic species. Appl. Sci. 2020, 10, 3218. [Google Scholar] [CrossRef]
- Canonica, L.; Cecchi, G.; Capra, V.; Di Piazza, S.; Girelli, A.; Zappatore, S.; Zotti, M. Fungal Arsenic Tolerance and Bioaccumulation: Local Strains from Polluted Water vs. Alloch. Strains. Environ. 2024, 11, 23. [Google Scholar]
- Wang, Y.; Yi, B.; Sun, X.; Yu, L.; Wu, L.; Liu, W.; Wang, D.; Li, Y.; Jia, R.; Yu, H.; et al. Removal and tolerance mechanism of Pb by a filamentous fungus: A case study. Chemosphere 2019, 225, 200–208. [Google Scholar] [CrossRef]
- Benavides, V.; Ciudad, G.; Pinto-Ibieta, F.; Robledo, T.; Rubilar, O.; Serrano, A. Enhancing Laccase and Manganese Peroxidase Activity in White-Rot Fungi: The Role of Copper, Manganese, and Lignocellulosic Substrates. Agronomy 2024, 14, 2562. [Google Scholar] [CrossRef]
- Agbeshie, A.A.; Abugre, S.; Atta-Darkwa, T.; Awuah, R. A review of the effects of forest fire on soil properties. J. For. Res. 2022, 33, 1419–1441. [Google Scholar] [CrossRef]
- Moazeni, S.; Cerda, A. The Impacts of Forest Fires on Watershed Hydrological Response. A review. Trees. For. People 2024, 18, 100707. [Google Scholar]
- García-Carmona, M.; Lepinay, C.; García-Orenes, F.; Baldrian, P.; Arcenegui, V.; Cajthaml, T.; Mataix-Solera, J. Moss biocrust accelerates the recovery and resilience of soil microbial communities in fire-affected semi-arid Mediterranean soils. Sci. Total Environ. 2022, 846, 157467. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.S.; Stark, F.G.; Berry, T.D.; Zeba, N.; Whitman, T.; Traxler, M.F. Pyrolyzed substrates induce aromatic compound metabolism in the post-fire fungus, Pyronema domesticum. Front. Microbiol. 2021, 12, 729289. [Google Scholar] [CrossRef] [PubMed]
- Filialuna, O.; Cripps, C. Evidence that pyrophilous fungi aggregate soil after forest fire. For. Ecol. Manag. 2021, 498, 119579. [Google Scholar] [CrossRef]
- Ascough, P.L.; Sturrock, C.J.; Bird, M.I. Investigation of growth responses in saprophytic fungi to charred bio-mass. Isot. Environ. Health Studies 2010, 46, 64–77. [Google Scholar] [CrossRef]
- Kouki, J.; Salo, K. Forest disturbances affect functional groups of macrofungi in young successional forests–harvests and fire lead to different fungal assemblages. For. Ecol. Manag. 2020, 463, 118039. [Google Scholar] [CrossRef]
- Hadibarata, T.; Kristanti, R.A.; Bilal, M.; Al-Mohaimeed, A.M.; Chen, T.W.; Lam, M.K. Microbial degradation and transformation of benzo [a] pyrene by using a white-rot fungus Pleurotus eryngii F032. Chemosphere 2022, 307, 136014. [Google Scholar] [CrossRef]
- Omoni, V.T.; Bankole, P.O.; Semple, K.T.; Ojo, A.S.; Ibeto, C.; Okekporo, S.E.; Harrison, I.A. Enhanced remediation of polycyclic aromatic hydrocarbons in soil through fungal delignification strategy and organic waste amendment: A review. Indian J. Microbiol. 2024, 1–23. [Google Scholar] [CrossRef]
- Zhang, W.; Li, Q.; Wang, J.; Wang, Z.; Zhan, H.; Yu, X.; Zheng, Y.; Xiao, T.; Zhou, L.W. Biodegradation of Benzo [a] pyrene by a White-Rot Fungus Phlebia acerina: Surfactant-Enhanced Degradation and Possible Genes Involved. J. Fungi 2023, 9, 978. [Google Scholar] [CrossRef]
- Harris, K.; Young, I.M.; Gilligan, C.A.; Otten, W.; Ritz, K. Effect of bulk density on the spatial organisation of the fungus Rhizoctonia solani in soil. FEMS Microbiol. Ecol. 2003, 44, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.; Raaijmakers, J.M.; Kuramas, E.E. Microbial extracellular polymeric substances: Ecological function and impact on soil aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef] [PubMed]
- Maillard, F.; Schilling, J.; Andrews, E.; Schreiner, K.M.; Kennedy, P. Functional convergence in the decomposition of fungal necromass in soil and wood. FEMS Microbiol. Ecol. 2020, 96, fiz209. [Google Scholar] [CrossRef]
- Morrissey, J.P.; Walsh, U.F.; O’donnell, A.; Moënne-Loccoz, Y.; O’gara, F. Exploitation of genetically modified inoculants for industrial ecology applications. Antonie Van Leeuwenhoek 2002, 81, 599–606. [Google Scholar] [CrossRef]
- Wozniak, C.A.; McClung, G.; Gagliardi, J.; Segal, M.; Matthews, K. Regulation of genetically engineered microorganisms under FIFRA, FFDCA and TSCA. In Regulation of Agricultural Biotechnology: The United States and Canada; Wozniak, C., McHughen, A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 57–94. [Google Scholar] [CrossRef]
- Tyagi, M.; da Fonseca, M.M.R.; de Carvalho, C.C. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation 2011, 22, 231–241. [Google Scholar] [CrossRef]
- Magan, N.; Fragoeiro, S.; Bastos, C. Environmental factors and bioremediation of xenobiotics using white rot fungi. Mycobiology 2010, 38, 238–248. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venâncio, C. The Quirky Rot Fungi: Underexploited Potential for Soil Remediation and Rehabilitation. Appl. Sci. 2025, 15, 1039. https://doi.org/10.3390/app15031039
Venâncio C. The Quirky Rot Fungi: Underexploited Potential for Soil Remediation and Rehabilitation. Applied Sciences. 2025; 15(3):1039. https://doi.org/10.3390/app15031039
Chicago/Turabian StyleVenâncio, Cátia. 2025. "The Quirky Rot Fungi: Underexploited Potential for Soil Remediation and Rehabilitation" Applied Sciences 15, no. 3: 1039. https://doi.org/10.3390/app15031039
APA StyleVenâncio, C. (2025). The Quirky Rot Fungi: Underexploited Potential for Soil Remediation and Rehabilitation. Applied Sciences, 15(3), 1039. https://doi.org/10.3390/app15031039