
Gonadal Development in European Eel Populations of North Adriatic Lagoons at Different Silvering Stages

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of Eels
2.2. Gonad Sampling
2.3. Statistical Analysis
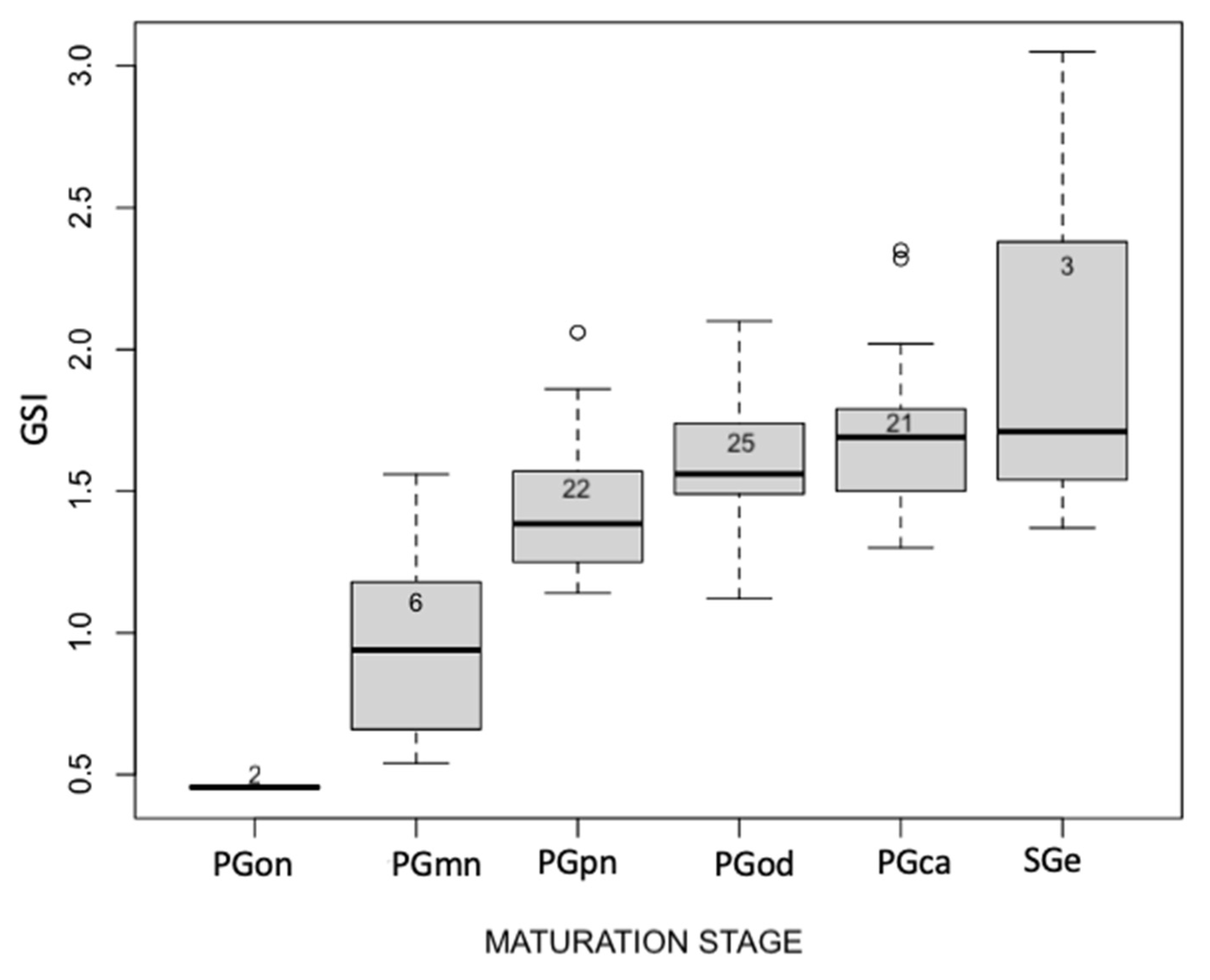
3. Results
3.1. Measurements
3.2. Histological Analysis
- -
- One-nucleoli (PGon): characterized by only one nucleolus inside the germinal vesicle, the absence of lipid droplets, and a reduced oocyte diameter of about 32.5 μm;
- -
- Multiple nucleoli (PGmn): characterized by multiple nucleoli inside the germinal vesicle arranged randomly. There is also the presence of globular masses called Balbiani’s bodies within the cytoplasm;
- -
- Perinucleolar (PGpn): characterized by the presence of the first lipid droplets and the increase of the oocyte diameter to around 60 μm;
- -
- Circumnuclear oil droplet (PGod): characterized both by the position of nucleoli that is arranged peripherally and by the increase of lipid droplets around the germinal vesicle. The oocyte diameter is around 75 μm;
- -
- The cortical stage (PGca) is characterized by an increase in lipid droplets arranged around the germinal vesicle and the presence of cortical alveoli in the periphery. The oocyte diameter is around 80 μm;
- -
- Early secondary growth (SGe): this stage is characterized by the presence of yolk at the periphery of the oocyte. The oocyte diameter is around 95 μm.
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ege, V. A revision of the genus Anguilla Shaw. Dana Rep. 1939, 16, 8–256. [Google Scholar]
- Watanabe, S.; Aoyama, J.; Tsukamoto, K. A new species of freshwater eel Anguilla luzonensis (Teleostei: Anguillidae) from Luzon Island of the Philippines. Fish. Sci. 2009, 75, 387–392. [Google Scholar] [CrossRef]
- Kuroki, M.; Aoyama, J.; Miller, M.; Yoshinaga, T.; Shinoda, A.; Hagihara, S.; Tsukamoto, K. Sympatric spawning of Anguilla marmorata and Anguilla japonica in the western North Pacific Ocean. J. Fish Biol. 2009, 74, 1853–1865. [Google Scholar] [CrossRef]
- Tesch, F.-W.; Bartsch, P. The Eel; Wiley Online Library: Hoboken, NJ, USA, 2003. [Google Scholar]
- Dekker, W. Status of the European eel stock and fisheries. In Eel Biology; Springer: Berlin, Germany, 2003; pp. 237–254. [Google Scholar]
- Dekker, W. The history of commercial fisheries for European eel commenced only a century ago. Fish. Manag. Ecol. 2019, 26, 6–19. [Google Scholar] [CrossRef]
- Drouineau, H.; Durif, C.; Castonguay, M.; Mateo, M.; Rochard, E.; Verreault, G.; Yokouchi, K.; Lambert, P. Freshwater eels: A symbol of the effects of global change. Fish Fish. 2018, 19, 903–930. [Google Scholar] [CrossRef]
- ICES. European Eel (Anguilla anguilla) Throughout Its Natural Range; ICES Advice: Copenhagen, Denmark, 2020. [Google Scholar]
- Pankhurst, N. Relation of visual changes to the onset of sexual maturation in the European eel Anguilla anguilla (L.). J. Fish Biol. 1982, 21, 127–140. [Google Scholar] [CrossRef]
- Fontaine, Y.; Pisam, M.; Le Moal, C.; Rambourg, A. Silvering and gill “mitochondria-rich” cells in the eel, Anguilla anguilla. Cell Tissue Res. 1995, 281, 465–471. [Google Scholar] [CrossRef]
- Waring, H. Color Change Mechanisms of Cold-Blooded Vertebrates; Academic Press: New York, NY, USA, 1963; Volume 266. [Google Scholar]
- Rohr, D.H.; Lokman, P.M.; Davie, P.S.; Young, G. 11-Ketotestosterone induces silvering-related changes in immature female short-finned eels, Anguilla australis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 701–714. [Google Scholar] [CrossRef]
- Boëtius, I.; Boétius, J. Lipid and protein content in Anguilla anguilla during growth and starvation. Dana 1985, 4, 1–17. [Google Scholar]
- Svedäng, H.; Wickström, H. Low fat contents in female silver eels: Indications of insufficient energetic stores for migration and gonadal development. J. Fish Biol. 1997, 50, 475–486. [Google Scholar] [CrossRef]
- Durif, C.; Dufour, S.; Elie, P. The silvering process of Anguilla anguilla: A new classification from the yellow resident to the silver migrating stage. J. Fish Biol. 2005, 66, 1025–1043. [Google Scholar] [CrossRef]
- Van Ginneken, V.J.; Maes, G.E. The European eel (Anguilla anguilla, Linnaeus), its lifecycle, evolution and reproduction: A literature review. Rev. Fish Biol. Fish. 2005, 15, 367–398. [Google Scholar] [CrossRef]
- Melià, P.; Bevacqua, D.; Crivelli, A.J.; Panfili, J.; De Leo, G.A.; Gatto, M. Sex differentiation of the European eel in brackish and freshwater environments: A comparative analysis. J. Fish Biol. 2006, 69, 1228–1235. [Google Scholar] [CrossRef]
- Van den Thillart, G.; Dufour, S. How to estimate the reproductive success of European silver eels. In Spawning Migration of the European Eel; Springer: Berlin, Germany, 2009; pp. 3–9. [Google Scholar]
- Okamura, A.; Horie, N.; Mikawa, N.; Yamada, Y.; Tsukamoto, K. Recent advances in artificial production of glass eels for conservation of anguillid eel populations. Ecol. Freshw. Fish 2014, 23, 95–110. [Google Scholar] [CrossRef]
- Gong, X.; Wang, D.; Bao, B.; Zhang, Q.; Jiang, X.; Liu, L. Gonadal development and silvering of the Japanese Eel (Anguilla japonica) in the Yangtze River during downstream migration. Aquac. Fish. 2017, 2, 173–178. [Google Scholar] [CrossRef]
- Mordenti, M.; Di Biase, A.; Sirri, R.; Modugno, S.; Tasselli, A. Induction of Sexual Maturation in Wild Female European Eels (Anguilla anguilla) in Darkness and Light. Isr. J. Aquac. 2012, 64, 1–9. [Google Scholar]
- Mordenti, O.; Di Biase, A.; Bastone, G.; Sirri, R.; Zaccaroni, A.; Parmeggiani, A. Controlled reproduction in the wild European eel (Anguilla anguilla): Two populations compared. Aquac. Int. 2013, 21, 1045–1063. [Google Scholar] [CrossRef]
- Fontaine, Y. L’argenture de l’anguille: Métamorphose, anticipation, adaptation. Bull. Français De La Pêche Et De La Piscic. 1994, 335, 171–185. [Google Scholar] [CrossRef][Green Version]
- Durif, C.M.; Dufour, S.; Elie, P. Impact of silvering stage, age, body size and condition on reproductive potential of the European eel. Mar. Ecol. Prog. Ser. 2006, 327, 171–181. [Google Scholar] [CrossRef]
- Durif, C.M.; Ginneken, V.v.; Dufour, S.; Müller, T.; Elie, P. Seasonal evolution and individual differences in silvering eels from different locations. In Spawning Migration of the European Eel; Springer: Berlin, Germany, 2009; pp. 13–38. [Google Scholar]
- Devlaming, V.; Grossman, G.; Chapman, F. On the use of the gonosomatic index. Comp. Biochem. Physiol. Part A Physiol. 1982, 73, 31–39. [Google Scholar] [CrossRef]
- West, G. Methods of assessing ovarian development in fishes: A review. Mar. Freshw. Res. 1990, 41, 199–222. [Google Scholar] [CrossRef]
- Ganias, K.; Somarakis, S.; Koutsikopoulos, C.; Machias, A. Factors affecting the spawning period of sardine in two highly oligotrophic Seas. Mar. Biol. 2007, 151, 1559–1569. [Google Scholar] [CrossRef]
- Hinton, M.G. Use of gonad indices to estimate the status of reproductive activity of female swordfish, Xiphias gladius: A validated classification method. Fish Bull. 1997, 95, 80–84. [Google Scholar]
- McQuinn, I.H. Identification of spring-and autumn-spawning herring (Clupea harengus harengus) using maturity stages assigned from a gonadosomatic index model. Can. J. Fish. Aquat. Sci. 1989, 46, 969–980. [Google Scholar] [CrossRef]
- Vitale, F.; Svedäng, H.; Cardinale, M. Histological analysis invalidates macroscopically determined maturity ogives of the Kattegat cod (Gadus morhua) and suggests new proxies for estimating maturity status of individual fish. ICES J. Mar. Sci. 2006, 63, 485–492. [Google Scholar] [CrossRef]
- Mordenti, O.; Di Biase, A.; Casalini, A.; Emmanuele, P.; Melotti, P.; Roncarati, A. Growth performances and natural diet of European eel (Anguilla anguilla L.) reared in muddy and sandy ponds. Aquat. Living Resour. 2016, 29, 105. [Google Scholar] [CrossRef]
- Ciccotti, E. Italy. In Management of the European Eel; Moriarty, C., Dekker, W., Eds.; Marine Institute: Dublin, Ireland, 1997; Volume 15, pp. 61–65. [Google Scholar]
- Grier, H.; Uribe-Aranzábal, M.; Patiño, R. The ovary, folliculogenesis, and oogenesis in teleosts. Reprod. Biol. Phylogeny Fishes 2009, 8, 25–84. [Google Scholar]
- Han, Y.-s.; Liao, I.-c.; Huang, Y.-s.; He, J.-t.; Chang, C.-W.; Tzeng, W.-N. Synchronous changes of morphology and gonadal development of silvering Japanese eel Anguilla japonica. Aquaculture 2003, 219, 783–796. [Google Scholar] [CrossRef]
- Pérez, L.; Peñaranda, D.; Dufour, S.; Baloche, S.; Palstra, A.; Van Den Thillart, G.; Asturiano, J. Influence of temperature regime on endocrine parameters and vitellogenesis during experimental maturation of European eel (Anguilla anguilla) females. Gen. Comp. Endocrinol. 2011, 174, 51–59. [Google Scholar] [CrossRef]
- Emmanuele, P.; Casalini, A.; Pisati, D.; Andreini, R.; Guercilena, N.; Parmeggiani, A.; Zaccaroni, A.; Mordenti, O. Artificial reproduction of Anguilla anguilla: Evaluation of biometrics characteristics of a population from Valle Campo Lagoon, Comacchio (Italy). Aquac. Int. 2020, 28, 777–790. [Google Scholar] [CrossRef]
- Di Biase, A.; Lokman, P.M.; Govoni, N.; Casalini, A.; Emmanuele, P.; Parmeggiani, A.; Mordenti, O. Co-treatment with androgens during artificial induction of maturation in female eel, Anguilla anguilla: Effects on egg production and early development. Aquaculture 2017, 479, 508–515. [Google Scholar] [CrossRef]
- De Leo, G.; Gatto, M. Trends in vital rates of the European eel: Evidence for density dependence? Ecol. Appl. 1996, 6, 1281–1294. [Google Scholar] [CrossRef]
- Ackermann, G.E.; Schwaiger, J.; Negele, R.D.; Fent, K. Effects of long-term nonylphenol exposure on gonadal development and biomarkers of estrogenicity in juvenile rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2002, 60, 203–221. [Google Scholar] [CrossRef]
- Palstra, A.; Cohen, E.; Niemantsverdriet, P.; Van Ginneken, V.; Van den Thillart, G. Artificial maturation and reproduction of European silver eel: Development of oocytes during final maturation. Aquaculture 2005, 249, 533–547. [Google Scholar] [CrossRef]
- Vøllestad, L.A. Tagging experiments with yellow eel, Anguilla anguilla (L.), in brackish water in Norway. Sarsia 1988, 73, 157–161. [Google Scholar] [CrossRef]
- Vøllestad, L.A. Geographic variation in age and length at metamorphosis of maturing European eel: Environmental effects and phenotypic plasticity. J. Anim. Ecol. 1992, 61, 41–48. [Google Scholar] [CrossRef]
- Hain, J. The behaviour of migratory eels, Anguilla rostrata, in response to current, salinity and lunar period. Helgoländer Wiss. Meeresunters. 1975, 27, 211–233. [Google Scholar] [CrossRef]
- Di Biase, A.; Bastone, G.; Casalini, A.; Parmeggiani, A.; Costantini, G.; Mordenti, O. Study of the morpho-physiological char-acteristics of two Anguilla anguilla populations selected for artificial reproduction. In Proceedings of the Global Aquaculture: Securing Our Future—AQUA, Prague, Czech Republic, 1–5 September 2012; pp. 1–5. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages | Codes | Steps |
|---|---|---|
| Oogonia proliferation | OP | Frequently form cell nest |
| Chromatin nucleolus (CN) | CNI | Leptotene |
| CNz | Zygotene | |
| CNp | Pachytene | |
| CNed | Early diplotene | |
| Primary Growth (PG) | PGon | One nucleolus (single nucleolus occupying the whole vesicle and presence of Balibiani’s body) |
| PGmn | Multiple nucleoli (multiple nucleoli scattered irregularly within the germinal vesicle) | |
| PGpn | Peronucleolar (all nucleoli begin to be oriented around the membrane of the germinal vesicle) | |
| PGod | Circumnuclear oil droplets (first lipid droplets begin to appear) | |
| PGca | Cortical alveolar (presence of cortical alveoli at the periphery of the ooplasm) | |
| Secondary Growth (SG) | SGe | Early secondary growth or early yolked oocytes (deposition on periphery of the first yolk globules) |
| SGI | Late secondary growth or late yolked oocytes (diameter of the oocyte significantly increases) | |
| SGfg | Full-growth oocytes (yolk distributed throughout the oocyte) | |
| Oocyte Maturation (OM) | OMegv | Eccentric germinal vesicle (oil droplets become one globule) |
| OMgvm | Germinal vesicle migration to animal pole (oocyte hydrates) | |
| OMgub | Germinal vesicle breakdown (oocyte hydration near completion) | |
| OMmr | Meiosis resumes, 2nd arrest | |
| Ovulation (OV) | OV | Oocyte emerges from the follicle, becomes an egg |
| Lagoon (Tot. Eels Sampled) | Parameter | Resident | Pre-Migrant | Migrant | |
|---|---|---|---|---|---|
| FII | FIII | FIV | FV | ||
| VALLE DI COMACCHIO (320) | % | 3.2 | 29.7 | 65.9 | 1.2 |
| K | 1.79 ± 0.17 | 2.06 ± 0.22 | 2.23 ± 0.23 | 1.95 ± 0.17 | |
| GSI n. 32 | 0.71 ± 0.17 | 1.65 ± 0.23 | 1.70 ± 0.16 | 1.74 ± 0.22 | |
| Body Weight (BW) | 404.3 ± 145.7 | 996.45 ± 119.36 | 1366.94 ± 221.99 | 755.00 ± 175.04 | |
| Total Body Length (BL) | 601.1 ± 52.7 | 786.12 ± 43.12 | 848.67 ± 50.56 | 726.25 ± 57.17 | |
| VALLE NUOVA (254) | % | 5.5 | 73.2 | - | 21.3 |
| K | 1.75 ± 0.22 | 1.81 ± 0.15 | - | 1.83 ± 0.23 | |
| GSI n. 25 | 0.62 ± 0.26 | 1.63 ± 0.19 | - | 1.96 ± 0.12 | |
| Body Weight (BW) | 433.92 ± 130.46 | 557.09 ± 89.00 | - | 554.29 ± 113.00 | |
| Total Body Length (BL) | 620.00 ± 62.52 | 673.61 ± 36.20 | - | 670.87 ± 46.26 | |
| VALLE CA’PASTA (112) | % | 1.8 | 76.8 | 7.1 | 14.3 |
| K | 1.52 ± 0.07 | 1.94 ± 0.17 | 2.18 ± 0.14 | 1.92 ± 0.18 | |
| GSI n. 11 | 0.46 ± 0.04 | 1.53 ± 0.26 | 1.77 ± 0.25 | 1.53 ± 0.18 | |
| Body Weight (BW) | 413.50 ± 8.77 | 747.43 ± 128.71 | 1020.48 ± 146.79 | 737.50 ± 90.23 | |
| Total Body Length (BL) | 648.50 ± 4.95 | 726.43 ± 43.26 | 775.13 ± 33.63 | 726.69 ± 35.60 | |
| VALLE SAN CARLO (168) | % | 1.8 | 55.3 | 3.0 | 39.9 |
| K | 1.66 ± 0.25 | 1.70 ± 0.17 | 1.99 ± 0.24 | 1.65 ± 0.20 | |
| GSI n. 17 | 0.70 ± 0.10 | 1.46 ± 0.33 | 2.08 ± 0.39 | 1.57 ± 0.13 | |
| Body Weight (BW) | 492.37 ± 57.53 | 616.80 ± 118.34 | 1143.90 ± 194.42 | 577.56 ± 112.70 | |
| Total Body Length (BL) | 668.67 ± 34.00 | 711.19 ± 41.61 | 831.80 ± 52.77 | 702.85 ± 39.52 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gentile, L.; Casalini, A.; Emmanuele, P.; Brusa, R.; Zaccaroni, A.; Mordenti, O. Gonadal Development in European Eel Populations of North Adriatic Lagoons at Different Silvering Stages. Appl. Sci. 2022, 12, 2820. https://doi.org/10.3390/app12062820
Gentile L, Casalini A, Emmanuele P, Brusa R, Zaccaroni A, Mordenti O. Gonadal Development in European Eel Populations of North Adriatic Lagoons at Different Silvering Stages. Applied Sciences. 2022; 12(6):2820. https://doi.org/10.3390/app12062820
Chicago/Turabian StyleGentile, Laura, Antonio Casalini, Pietro Emmanuele, Riccardo Brusa, Annalisa Zaccaroni, and Oliviero Mordenti. 2022. "Gonadal Development in European Eel Populations of North Adriatic Lagoons at Different Silvering Stages" Applied Sciences 12, no. 6: 2820. https://doi.org/10.3390/app12062820
APA StyleGentile, L., Casalini, A., Emmanuele, P., Brusa, R., Zaccaroni, A., & Mordenti, O. (2022). Gonadal Development in European Eel Populations of North Adriatic Lagoons at Different Silvering Stages. Applied Sciences, 12(6), 2820. https://doi.org/10.3390/app12062820